

Abstract
Efforts to develop animal models of memory are critical for understanding the neural substrate of memory. Memory is essential for daily life and enables information to be stored and retrieved after seconds to years. The ability to remember episodes from the past is thought to be related to the ability to plan for the future. Here we focus on a particular aspect of prospective cognition, namely the ability to remember to take action when a future scenario occurs. This review focuses on a recently developed method to evaluate prospective memory in the rat. Available evidence suggests that rats remember to take action in the future, but little is known about the temporal specificity of such memories or about the flexibility and limitations of prospective memories. Recent studies that suggest that rats remember a specific past episode are reviewed to underscore potential approaches that may be used to explore the range and limits of prospective cognition. The review highlights some directions to explore, including the temporal specificity of prospective cognition, the range of flexibility or creativity within prospective cognition, and the constraints imposed by multiple motivational systems.
Keywords: Prospective cognition, Prospective memory, Comparative cognition, Animal model, Episodic memory, Mental time travel
Prospective cognition in people
People remember the past and remember to take actions in the future. Reconstructing information from the past is critical to effectively plan to act in the future. Indeed, representing the future to simulate and predict possible future events depends on the same neural machinery that is used to remember the past (Schacter, Addis, & Buckner, 2007). These core brain systems include the medial prefrontal regions, posterior regions in the medial and lateral parietal cortex, the lateral temporal cortex, the medial temporal lobe, and hippocampus (Martin, Schacter, Corballis, & Addis, 2011; Schacter, et al., 2007). According to this view, integration of information from the past is used to construct simulations about possible future events. Hence, prospective cognition may involve episodic simulation, planning, prediction, and remembering intentions (Schacter, Addis, & Buckner, 2008). The temporal distance and elaboration of details regarding past and future events play an important role in episodic memory and prospective cognition (Addis & Schacter, 2008; Roberts, 2012, this issue; Roberts & Feeney, 2009; Schacter & Addis, 2007). Remote future events are characterized by disparateness of details. Moreover, the ability to specify a time point for episodic memories about the past and planning for a specific time in the future (rather than general knowledge about remoteness) plays an important role in comparative studies of episodic memory and planning (Roberts, 2012, this issue; Roberts & Feeney, 2009).
Because people remember to take actions in the future, we begin with some common examples of prospective memory that highlight the risk of memory failure. Parents need to remember to pick up children from daycare at an appropriate time, and failure to do so can have negative consequences (financial or de-enrollment). Patients need to remember to take medications at the appropriate time in the day, and memory failure can lead to under-dosing or over-medication (when forgetting to act or forgetting that the action has already been taken). Pilots need to remember to check instruments before commencing maneuvers or when other instruments reach a particular level, and memory failure can be catastrophic. A number of strategies support success in prospective memory. For example, smartphones are used to schedule appointments with audible or tactile prompts to check our schedules at appropriate times. Pills are organized into containers labeled for days of the week or times of day. Procedures that are of critical importance can be extensively trained (as is the case for pilots and healthcare personnel) together with redundant systems to detect potential errors.
Prospective memory is the ability to remember to take some action in the future (McDaniel & Einstein, 2007). The content of prospective memory includes a representation of an action to perform in the future. People “remember to remember” at an appropriate time (i.e., time-based prospective memory) or when a suitable event occurs (i.e., event-based prospective memory). The hallmark of prospective memory is that a deleterious effect on ongoing behavior occurs as the time to execute draws near because greater attentional resources are diverted to the now active prospective memory (Hicks, Marsh, & Cook, 2005; Kliegel, Martin, McDaniel, & Einstein, 2001; Marsh, Hicks, & Cook, 2006; Marsh, Hicks, & Landau, 1998; Smith, 2003; Smith, Hunt, McVay, & McConnell, 2007). According to this perspective, when people form a prospective memory, they temporarily put the memory representation into an inactive state while engaging in other activities. Later, the representation is reactivated at some point in the future. Ultimately, successful activation of the memory representation yields an action at an appropriate, yet temporally distant time. Prospective memory failures may occur when the memory representation fails to be reactivated or when reactivation occurs at an inappropriate time.
An important side effect of prospective memory is its deleterious effect on other, ongoing activity. Consider the childcare example described above. Remembering to pick up one’s child from daycare at the end of the day is an important action that needs to occur at some temporally distant point, rather than now. The intention to act is inactive (i.e., it is not actively rehearsed) throughout the day. As the appropriate time to act approaches, it becomes difficult to continue to engage in ongoing activities (e.g., concentrating on one’s work or participating in a meeting) likely because some cognitive resources are diverted to processing the prospective memory as the appropriate time approaches.
Prospective memory in rats
We recently developed an animal model of prospective memory (Wilson & Crystal, 2012). The basic insight for developing our model is that prospective memory is expected to produce a selective deficit in performance at the time when anticipation of a future event is greatest. Hence, our approach was to determine if anticipating a future event would produce a deleterious side effect on ongiong activity, specifically at a time when the representation of the event is most likely to be active.
Rats were trained in a temporal bisection task for 90 min per day. The bisection task began with the presentation of a 2- or 8-s signal followed by the opportunity to press one of two response levers. A small reward was delivered if the rat pressed the correct lever to classify the duration as short or long. Immediately after the bisection task each day, rats in the meal group received an 8-g meal whereas other rats in the no-meal group received no additional food. The meal was earned by interrupting a photobeam located inside a food trough, but photobeam breaks were only effective 90 min after the start of the bisection task. Rats in the meal group may remember to collect the meal, whereas rats in the no-meal group did not have an opportunity to learn to remember an additional action beyond the bisection task. Thus, we hypothesized that rats in the meal group would represent the meal as an initially inactive representation and only reactivate it at an appropriate time immediately before the start of the meal. Thus, we measured sensitivity to time in the bisection task at early and late time points. If rats have prospective memory, then we expect that they should exhibit a negative side effect on ongoing activity at the later time point (i.e., when the representation is most likely to be activated). We hypothesized that rats with prospective memory would both anticipate the start of the meal and show a selective deficit in performance at the time when meal anticipation is greatest. By contrast, if rats do not have prospective memory, then any change in sensitivity from early to late time points should be equivalent for both meal and no-meal groups.
Sensitivity to time in the ongoing task declined near the meal time in the meal group but not in the no-meal group, as predicted by the prospective-memory hypothesis. Performance in the ongoing task was examined at early and late time points (Figure 1a-b). The probability of long responses increased as a function of increasing durations and did not differ between early and late time points for both groups, as expected. However, temporal sensitivity (i.e., the steepness of the probability functions) decreased from early to late time points for the meal group but not for the no-meal group, as predicted by prospective memory. In particular, the slope of the psychophysical function was smaller at the late time point relative to the early time point, indicating a decline in performance as the meal approached. This decline in sensitivity is expected by prospective memory because it is more likely that the representation was active as the meal time approached. By contrast, bisection performance in the no-meal group did not show a decline in sensitivity over the same time points, which suggests that the sensitivity decline in the meal group was produced by the approaching meal. These observations suggest that sensitivity to time is relatively constant throughout the session when a representation of a meal is absent. Thus, we conclude that the approaching meal produced the decline in performance in ongoing activity in the meal group. Food-trough responses increased as a function of time in the meal group but not in the no-meal group (see Figure 1c), which suggests that the meal group anticipated the arrival of the meal, as expected.
Fig. 1.
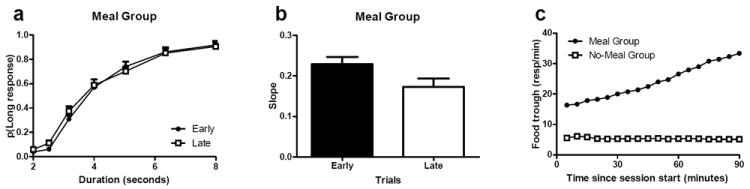
Anticipation of a meal reduced sensitivity to time in an ongoing interval-duration classification task near the meal time. Sensitivity to time in the ongoing task declined near the meal time in the meal group but not in the no-meal group. The probability of judging an interval as long (a) increased as a function of the interval duration. Sensitivity to time, as measured by the slope of the probability function (b) declined immediately before the end of the daily session in the meal group (a) but not in the no-meal group (not shown). Importantly, the interaction between early and late variable is significant in the meal group (a, p < 0.001) but not in the no-meal group (p = 0.1), and these differences are significant as shown by the three-way interaction (p < 0.009). Similarly, the slope of the psychophysical function was smaller at the late relative to early time points (p = 0.009) in the meal group but not in the no-meal group (p = 0.8), and these differences are significant as shown by the interaction (p = 0.03). The meal group anticipated the arrival of the meal, as shown by the increase in food-trough responses before the meal whereas the increase in food-trough responses was absent in the no-meal group (c). (a-c) Error bars indicate SEM. Reproduced from Wilson, A. G. & Crystal, J. D. (2012). Prospective memory in the rat. Animal Cognition. 15, 349-358. © 2011 Springer-Verlag.
Wilson and Crystal (2012) proposed that prospective memory produced the decline in sensitivity as the meal approached. According to this proposal, rats formed a representation of the meal but inactivated it at the early time point, when the meal was temporally distant. As the expectation of the meal grew, more attentional resources were recruited to maintain the representation of the forthcoming meal, which impaired temporal sensitivity on the bisection task. This hypothesis predicts that sensitivity to time would decline as the meal approached whereas the no-meal group’s sensitivity would remain constant throughout the session.
To support the representational account, a number of alternative, non-representational hypotheses were examined and ruled out by aspects of the data (e.g., attentional limit, response competition, contrast, fatigue; for review see Wilson & Crystal, 2012). As an example, consider response competition. Notice that the rats in the meal group were engaged in much more lever pressing than rats in the no-meal group. Perhaps the impairment in sensitivity to time occurred because the rats in the meal group suffered from response competition between pressing levers to express a judgment about intervals and investigating the food trough in anticipation of the meal. According to this non-prospective memory alternative explanation, sensitivity to time may have suffered because these rats may be less likely to hear the duration stimuli, process the interval duration, or remember the to-be-selected duration classification response while they were simultaneously investigating the food trough. Each version of this response-competition hypothesis proposes that engaging in food-trough responses causes the observed decline in temporal sensitivity. To test the hypothesis that robust behavior at the food trough as the meal approached causes the decrease in sensitivity to time, we examined temporal sensitivity and the number of food-trough responses on individual days. A response competition hypothesis predicts that a high number of responses produces low temporal sensitivity whereas sensitivity is high when few competing food-trough responses occur. Thus, response competition predicts a significant negative correlation between temporal sensitivity and the number of food-trough responses in the meal group immediately before the meal. Contrary to this prediction, the observed correlation was -0.006 ± 0.061 (mean ± SEM), which was not significantly different from zero. Response competition explains less than one hundredth of one percent, and a Bayesian analysis suggests that the null hypothesis of a zero correlation is a reasonably safe bet (Wilson & Crystal, 2012).
Evaluation of prospective memory in rats: strengths and limitations
The development of an animal model is likely characterized by a pattern of strengths and weaknesses, particularly at the early stages of development (Crystal, 2012). In this section, a number of strengths and potential weaknesses of the prospective-memory model are examined.
The major strength of the model is that it provides a method for evaluating the existence of a representation of a future event that would otherwise be behaviorally silent. Anticipating the arrival of a meal, in itself, is not evidence of prospective memory. By contrast, the observation that anticipating the arrival of the meal produces a deleterious effect on ongoing behavior suggests that rats form a prospective memory of the future meal.
One benefit of studying cognition in animals is that it may provide insight into impairments in cognition observed in people. Cognitive impairments in people are debilitating, and developing insight into the origins of such impairments may improve the effectiveness of treatments. Significant obstacles impede the translation of animal models to clinical conditions. Although there is a long history of studying learning and memory in animals, the types of cognitive processes involved in many cases of learning and memory may not match the types of impairments observed clinically. Thus, it is possible that treatments such as drug-development programs may be effective at the pre-clinical level but may not be effective when translated to clinical conditions in people. Therefore, the expansion of the suite of cognitive processes that may be modeled in animals may ultimately translate to improved therapies for debilitating memory impairments observed clinically.
Failures of prospective memory (i.e., forgetting to act on an intention at an appropriate time in the future) are a common feature of aging (Aberle, Rendell, Rose, McDaniel, & Kliegel, 2010; Craik, 1986; d’Ydewalle, Bouckaert, & Brunfaut, 2001; Driscoll, McDaniel, & Guynn, 2005; Henry, MacLeod, Phillips, & Crawford, 2004). In addition, prospective memory is impaired in a number of clinical populations, including mild cognitive impairment (Schmitter-Edgecombe, Woo, & Greeley, 2009; Troyer & Murphy, 2007), Alzheimer’s disease (Blanco-Campal, Coen, Lawlor, Walsh, & Burke, 2009; Jones, Livner, & Bäckman, 2006; Troyer & Murphy, 2007), Parkinson disease (Foster, McDaniel, Repovš, & Hershey, 2009; Raskin et al., 2011), and traumatic brain injury (Henry et al., 2007; McCauley, McDaniel, Pedroza, Chapman, & Levin, 2009). Prospective memory is dependent on the integrity of the prefrontal cortex and Brodmann’s area 10 in particular (Burgess, Scott, & Frith, 2003; Hashimoto, Umeda, & Kojima, 2011; Simons, Schölvinck, Gilbert, Frith, & Burgess, 2006). Thus, an animal model of prospective memory may be used to explore neuroanatomical, neurochemical, and genetic mechanisms for representations of the future in future research. Such work would exploit the extensive knowledge about the neuroanatomy and neurophysiology of the rodent hippocampus and other parts of the core network and utilize neuroscience (e.g., pharmacological, electrophysiological, RNA interference, and targeted gene expression) techniques (Eriksen & Janus, 2007; Hwang et al., 2004; Jankowsky et al., 2005; Keri et al., 2009; Maxwell, 2009; Ueberham et al., 2006). Thus, identifying mechanisms that govern prospective memory holds enormous potential to significantly benefit society by providing insights into deficits in memory associated with aging, brain injuries, amnesia, Alzheimer’s disease and other human memory pathologies.
There are potential limitations of the model. One perspective on prospective memory in animals is that it taps into planning for the future or is a precursor to planning. Maintaining a representation of a future event is a prerequisite for planning yet it need not involve fully developed planning. For example, other studies of planning (Cheke & Clayton, 2012; Correia, Dickinson, & Clayton, 2007; Naqshbandi & Roberts, 2006; Raby, Alexis, Dickinson, & Clayton, 2007) fully meet criteria for planning according to which the animal takes action now for a future need that is fully dissociated from current motivational needs (Suddendorf & Corballis, 1997; Suddendorf & Corballis, 2007). According to this mental time travel approach, an animal forms a representation in which it envisions itself in a future scenario; moreover, the representation is posited to be about a specific point in the future (Roberts & Feeney, 2009). By contrast, in our approach, rats were food restricted and participated in two tasks that both provided food. Hence, our approach clearly did not seek to dissociate motivational states. Moreover, in experiments that sought to demonstrate planning in rats using techniques that did dissociate motivational states, no evidence for planning was obtained (Naqshbandi & Roberts, 2006). Therefore, it is possible that rats exhibit a precursor to planning only in a limited sense, and they may not be capable of more robust planning; alternatively, refinements in techniques may reveal more robust planning in future research. Although considerable progress has been made within the mental time travel framework, it has recently been argued that future-oriented abilities should also be evaluated outside this framework (Raby, et al., 2007; Zentall, 2006, 2010). One advantage of our attempt to model prospective memory in rats outside the mental-time-travel framework is that it may provide insight into the evolution of planning to act in the future by focusing on deleterious side-effects of a prospective memory representation that may be evaluated across a wide array of species; see Beran, Perdue, Bramlett, Menzel and Evans (2012, this issue) for a demonstration of prospective cognition in a language-trained chimpanzee. Because a precursor to planning may exist in the absence of other aspects of planning, it will be valuable to determine how motivational states constrain prospective memory in future research.
Aside from the issues of dissociating motivational states, our approach is a precursor to planning rather than robust planning in another respect. A hallmark of planning is flexibility or creativity. However, our approach involved extensive training and testing in the same conditions throughout the experiment. Efforts to probe flexibility will need to examine the ability to apply a learned representation of a future event to novel situations in future research. Hence, it will be valuable to determine the range or limits of flexibility or creative deployment of a plan in prospective memory.
An important issue to evaluate in candidates for mental time travel in animals is the ability to pinpoint the representation to a rather specific event in time (Roberts & Feeney, 2009; Roberts et al., 2008). Roberts and colleagues have argued that a candidate for mental time travel in animals is qualitatively comparable to that of human mental time travel only if the representation is clearly demonstrated to be for a specific time. In the case of episodic memory, Roberts and colleagues have argued that it is necessary to show that an animal remembers the specific time when an earlier event occurred, and similarly in the case of prospective cognition, it is necessary to show that an animal represents a specific time in the future. Figure 2 illustrates Roberts and Feeney’s (2009) proposed conceptualization of mental time travel. In the next section, experiments that meet this criterion for memory of a specific past episode are reviewed to highlight techniques that may prove similarly useful to evaluate the hypothesis that a specific time in the future is represented in prospective cognition.
Fig. 2.
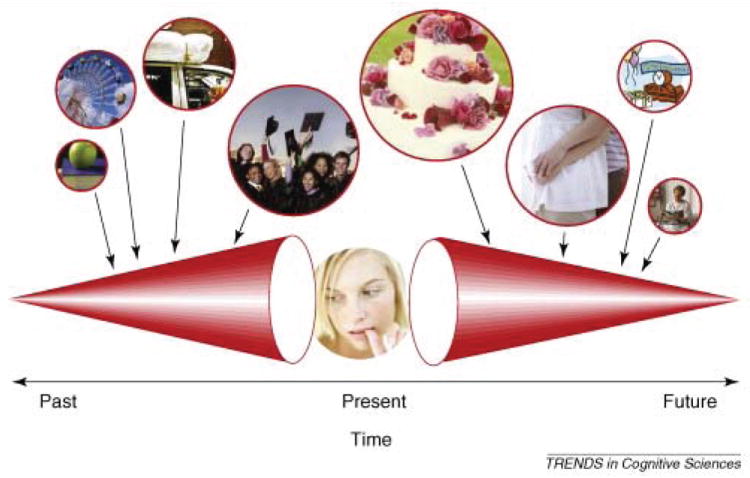
Roberts and Feeney’s (2009) proposed conceptualization of mental time travel. Memories of the past and planned events for the future are shown at varying temporal distances into the past or future. Temporal remoteness determines disparateness of detail. The size of the cones and balloons represent the clarity of the memories or plans. Because remoteness plays a critical role according to this proposal, Roberts and Feeney further propose that tests for mental time travel in animals must show that animals remember when a specific event occurred in the past and that they are planning for a specific time in the future. Reproduced from Roberts, W. A., & Feeney, M. C. (2009). The comparative study of mental time travel. Trends in Cognitive Sciences, 13, 271-277. © 2009 Elsevier.
Rats remember when an earlier episode occurred
The central hypothesis in animal models of episodic memory is that, at the time of a memory assessment, the animal remembers a specific earlier event. One approach to isolate memory of a specific earlier event is to focus on what-where-when memory (Clayton, Bussey, & Dickinson, 2003; Clayton & Dickinson, 1998); that is, memory for what happened, where it took place, and when in time it occurred. Importantly, availability of alternative strategies by which to produce putative episodic-memory performance without memory of the specific earlier event represents a threat to the episodic-memory hypothesis (Crystal, 2009, 2010, 2011; Roberts, et al., 2008; Zentall, 2005, 2006). Other approaches provide independent evidence for episodic memory in rats (Eacott & Easton, 2012, this issue; Eacott, Easton, & Zinkivskay, 2005; Zhou, Hohmann, & Crystal, in press) but are beyond the scope of this review. Efforts to document memory for a specific earlier event in rats have been complicated by some initial attempts that did not produce robust memory (Bird, Roberts, Abroms, Kit, & Crupi, 2003; Roberts & Roberts, 2002). Subsequent studies suggested that rats remember what-where-when (Babb & Crystal, 2005, 2006a, 2006b; Eacott, et al., 2005; Eacott & Norman, 2004; Kart-Teke, De Souza Silva, Huston, & Dere, 2006; Naqshbandi, Feeney, McKenzie, & Roberts, 2007). These may be regarded as proof of concept studies, which require validation by ruling out alternative explanations. An important attempt to validate episodic memory provided both episodic memory and non-episodic memory solutions and found that, in this situation, rats more readily rely on the non-episodic memory solution (Roberts, et al., 2008). Hence, we sought to eliminate the availability of non-episodic memory solutions to develop a working model of episodic memory in rats (Zhou & Crystal, 2009, 2011). A series of experiments by Zhou and Crystal (2009, 2001) that document memory of a specific earlier event in rats are described below.
In the Zhou and Crystal (2009) study, replenishment of a distinctive flavor at a recently presented location could be predicted by remembering the time at which an earlier episode occurred. Thus, what-where-when memory could be used to predict replenishment (or non-replenishment) only if the animals remembered when the earlier episode occurred. Rats’ memory was assessed once per day, either in the morning or in the afternoon (see Figure 3a). Chocolate replenished at a daily unique location at only one of these times of day (morning for some rats; afternoon for other rats). Another flavor (regular chow) was available at all other locations but never replenished. The interval between memory encoding (study phase) and memory assessment (test phases) was constant (approximately 2 min). Because the location of chocolate varied randomly across days and the morning and afternoon sessions were presented in random order, what-where-when memory would be implicated if the rats visited the chocolate location selectively on occasions when chocolate was about to replenish. Indeed, when the chocolate location was about to replenish, the rats revisited that location at a higher rate relative to equivalent trials in which chocolate did not replenish (Figure 4a). Differential rates of revisiting chocolate-flavored locations was accomplished while rats accurately avoided revisits to depleted chow-flavored locations. These data are consistent with the hypothesis that rats used what-where-when memories to adjust revisit rates to the daily-unique chocolate location. Importantly, what-where-when memory in this study could not be based on the delay between study and test (i.e., it could not be based on judging relative familiarity of the study items, judging how long ago the study occurred, or timing an interval between study and test) because the retention interval was constant in replenish and non-replenish conditions. Hence, an important non-episodic memory solution was controlled throughout these experiments.
Fig. 3.
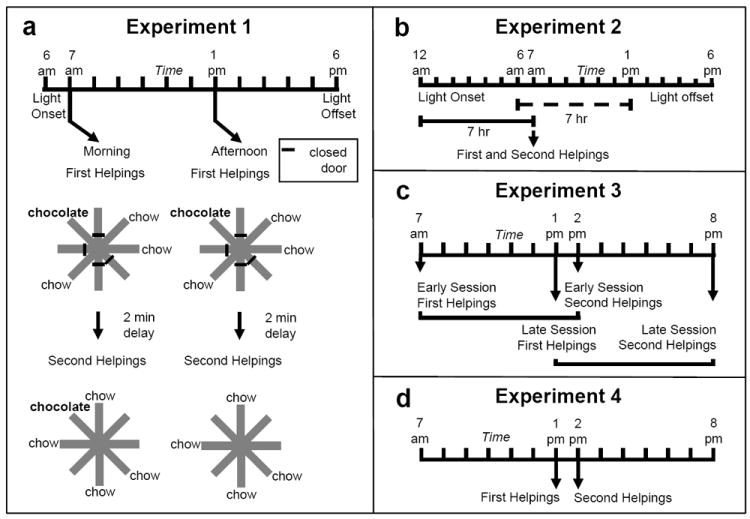
Experimental design of Zhou and Crystal’s (2009) study. a. Design of Experiment 1. The morning or afternoon was randomly selected for presentation of first helpings (study phase; encoding) and second helpings (test phase; memory assessment) of food. An example of the accessible arms and flavors in study and test phases is shown. Chocolate or chow flavored pellets were available at four arms in the study phase (randomly selected). After a 2-min retention interval, chow-flavored pellets were available at previously inaccessible locations in the test phase. Chocolate replenished in the test phase conducted in the morning (7 a.m.) but not in the afternoon (1 p.m.) for half of the rats; these contingencies were reversed for the remaining rats (not shown). For each rat, one session (i.e., study and test phases) was conducted per day. b. Phase-shift design of Experiment 2. Light onset occurred at 12 a.m. (i.e., 6 hr earlier than in Experiment 1) and the study and test phases occurred at the time of a typical morning session. Note that 7 hr elapsed between light onset and the study-test sequence (solid horizontal line), which is comparable to the time between the typical light onset and a typical afternoon session (dashed horizontal line) in Experiment 1. The design of the experiment puts predictions for time-of-day and how-long-ago cues in conflict. Thus, a rat would be expected to behave as in its morning baseline (based on time of day) or as in its afternoon baseline (based on how long ago). c. Transfer-test design of Experiment 3. The time of day at which the study phase occurred was the same as in Experiment 1. The introduction of 7-hr retention intervals in Experiment 3 produced test phases that occurred at novel times of day. Early and late sessions had study times (but not test times) that corresponded to those in Experiment 1. The first two sessions in Experiment 3 consisted of one replenishment and one non-replenishment condition. An early or late session was randomly selected on subsequent days. Differential revisits to the chocolate location is expected if the rats were adjusting revisit rates based on the time of day at which the study episode occurred; revisit rates are expected to be equal in early and late sessions if the rats used time of day at which the test phase occurred. Study and test phases were as in Experiment 1, except that they were separated by 7-hr delays (shown by horizontal brackets). d. Conflict-test design of Experiment 4. The study and test phases occurred at 1 p.m. and 2 p.m., respectively. These times correspond to the typical time of day at which a late-session study phase and early-session test phase occurred in Experiment 3. The design of the experiment put predictions for time of day at study and time of day at test in conflict. A rat would be expected to behave as in its early-session, second-helpings baseline (based on test time of day) or as in its late-session, second-helpings baseline (based on study time of day). Reproduced with permission from Zhou, W., & Crystal, J. D. (2009). Evidence for remembering when events occurred in a rodent model of episodic memory. Proceedings of the National Academy of Sciences of the United States of America, 106, 9525-9529. © 2009 National Academy of Sciences, U.S.A.
Fig. 4.
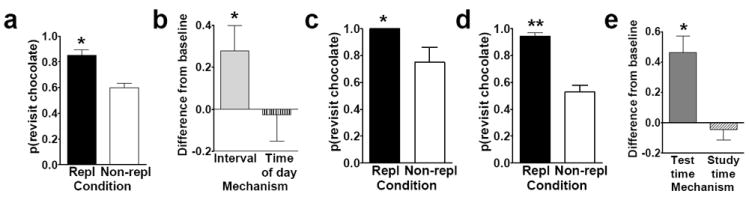
a. Rats preferentially revisit the chocolate location when it is about to replenish in Experiment 1. The probability of a revisit to the chocolate location in the first four choices of a test phase is shown for replenishment and non-replenishment conditions. b. Rats used time of day, rather than an interval, to adjust revisit rates in Experiment 2. The figure plots the difference between observed and baseline revisit rates. For the bar labeled interval, the baseline was the probability of revisiting chocolate in the afternoon; thus, the significant elevation above baseline shown in the figure suggests that the rats did not use an interval mechanism. For the bar labeled time of day, the baseline was the probability of revisiting chocolate in the morning; thus, the absence of a significant elevation above baseline is consistent with the use of time of day. The horizontal line corresponds to the baseline revisit rate to the chocolate location from Experiment 1. Positive difference scores correspond to evidence against the hypothesis indicated on the horizontal axis. c. and d. Rats preferentially revisited the chocolate location when it was about to replenish when the study, but not the test, time of day was familiar in Experiment 3. The probability of a revisit to the chocolate location in the first four choices of a test phase is shown for first replenishment and first non-replenishment conditions (c; initial) and for subsequent sessions (d; terminal). e. Rats remembered the time of day at which the study episode occurred in Experiment 4. Rats treated the novel study-test sequence as a late-session test phase, suggesting memory of the time of day at study rather than discriminating time of day at test. The figure plots the difference between observed and baseline revisit rates. For the bar labeled test time, the baseline was the probability of revisiting chocolate in the test phase of the early session in Experiment 3; thus, the significant elevation above baseline suggests that the rats did not use the time of day at test to adjust revisit rates. For the bar labeled study time, the baseline was the probability of revisiting chocolate in the test phase of the late session in Experiment 3; thus, the absence of a significant elevation above baseline is consistent with memory of the time of day at study. The horizontal line corresponds to the baseline revisit rate to the chocolate location from Experiment 3 (terminal). Positive difference scores correspond to evidence against the hypothesis indicated on the horizontal axis. a-e. Error bars indicate SEM. a, c, and d. The probability expected by chance is 0.41. Repl = replenishment condition. Non-repl = non-replenishment condition. a. * P < 0.001 difference between conditions. b. * P < 0.04 different from baseline. c and d. * P < 0.04 and ** P < 0.0001 difference between conditions. e. * P < 0.001 different from baseline. Reproduced with permission from Zhou, W., & Crystal, J. D. (2009). Evidence for remembering when events occurred in a rodent model of episodic memory. Proceedings of the National Academy of Sciences of the United States of America, 106, 9525-9529. ©2009 National Academy of Sciences, U.S.A.
Next Zhou and Crystal (2009) sought to rule out remaining non-episodic memory explanations. First, we determined the type of timing mechanism used in what-where-when memory. According to an episodic memory explanation, at the time of memory assessment, the rats remembered the earlier study episode and adjusted revisits to chocolate at test accordingly. Alternatively, the rats may have been reactive at the time of test based on other available cues without remembering the study episode. Hence, we tested the following two proposals. According to the circadian time-of-day hypothesis, the rats used a circadian signal (i.e., morning vs. afternoon) to adjust revisit rates at the daily-unique chocolate location; this view is consistent with the episodic-memory hypothesis that the rats remember the specific time of day at which the study episode occurred. Alternatively, according to the interval-timing hypothesis, the rats timed the interval from light onset in the colony to the morning and afternoon sessions. Morning and afternoon sessions occurred 1 and 7 hr, respectively, after light onset in the colony. Importantly, adjusting the revisit strategy based on the passage of time since light onset can be done without remembering the time at which the study episode occurred, which makes the interval-timing proposal a non-episodic memory hypothesis. To test these hypotheses, we used a 6-hr phase shift of light onset. The lights in the colony were turned on 6 hr early and the probe session was conducted at the usual time in the morning (see Figure 3b). According to the circadian time-of-day hypothesis, the rats would treat the probe as a morning session because an endogenous circadian oscillator is not expected to adjust immediately to a phase shift. Alternatively, according to the interval-timing hypothesis, the rats would treat the probe as an afternoon session because afternoon sessions typically occur 7 hr after light onset in the colony. The rats did not use the interval between light onset and the session, suggesting that they used circadian time of day (Figure 4b).
Next, we sought to determine if rats remember the time at which the earlier episode occurred (an episodic-memory hypothesis) or, alternatively, if they were merely selectively reactive at the different times of test. Importantly, reactivity at the time of test can occur without a memory of the earlier episode, making this a non-episodic memory alternative. Hence, we determined if it was the time of day at study or at test that was responsible for the different rates of revisiting the chocolate location. Because a 2-min delay between study and test is too small for rats to discriminate based on a circadian oscillator (Pizzo & Crystal, 2004), we increased the delay to 7 hr (see Figure 3c). Importantly, the time of day at study was familiar from earlier training, but the time of day at test was unfamiliar (approximately 7 hr later than usual). Consequently, if the rats remembered the study episode (i.e., they used time of day at study), then they should continue to differentially revisit the chocolate locations when their memory was assessed at novel times. Alternatively, if the rats were merely reactive to the time of day at test (i.e., they used time of day at test without remembering the earlier study episode), then there is no basis for them to revisit chocolate locations at different rates in the morning and afternoon because the test times were unfamiliar. When tested with novel test times of day after familiar morning or afternoon study times of day, we observed complete transfer (i.e., the differential rates of revisiting occurred on the very first trial in the morning and afternoon; Figure 4c-d). These data suggest that at the time of memory assessment, the rats remembered the time of day at which the study episode occurred.
We obtained an additional line of evidence for the same episodic-memory conclusion by putting episodic (study time) and non-episodic (test time) hypotheses into conflict. We used a novel combination of study and test times to determine if the rats remember the study episode or are merely reactive at the time of test. The 7-hr delays between study and test phases produced a 1-hr overlap between the two types of trials, which allowed us to start a trial with a late study phase and end the trial with an early test phase (see Figure 3d). Again we sought to determine if the rats were adjusting revisit rates in the test phase based on the time of day at test (test-time hypothesis; a non-episodic memory proposal) or based on memory of the time of day at which the study phase occurred (study-time hypothesis; an episodic memory proposal). According to the test-time hypothesis, the rats should revisit at the usual baseline rate that typically occurred on tests at that time of day. Alternatively, according to the study-time hypothesis, the rats should revisit at the usual time of day that occurred after a later study time (which usually is followed by a test 7 hr later, rather than 1 hr later). The rats adjusted chocolate revisits based on the time of day at study rather than the time of day at test (Figure 4e). These data suggest that rats remembered the study episode, and the time of day at which the study episode occurred, providing a second line of evidence that converges on the conclusion that rats remember when the earlier study episode occurred.
The data from Zhou and Crystal (2009) are consistent with the hypothesis that rats have specific knowledge about earlier episodes, including when the episode occurred, what happened, and where it took place. However, the rats might have detected that encoding the chocolate location was not required in some time-of-day conditions, which we refer to as the encoding failure hypothesis. For example, a rat might solve the task by selectively encoding the location of chocolate only at one time of day (e.g., when chocolate replenished in the afternoon) but not encoding the location of chocolate at the other time of day (e.g., when chocolate did not replenish in the morning); this is a non-episodic-memory alternative because differential rates of revisiting chocolate would occur in this situation without remembering the episode. The encoding failure hypothesis could also explain data from other studies (Naqshbandi, et al., 2007).
To address this potential problem, Zhou and Crystal (2011) conducted a what-where-when study that required encoding in every single trial. We provided rats with daily information about a preferred food type (chocolate) that replenished or did not replenish at its previously encountered location (Figure 5a). Another flavor (regular chow) was available at all other locations but never replenished. Importantly, although some of the information needed to predict replenishment was available at the time of encoding (location, time of day, food flavor), one critical piece of information needed to predict replenishment was not presented at encoding. Instead, this piece of information was provided only immediately before the memory assessment. The presence or absence of additional chocolate pellets in a central location could be used to predict replenishment when combined with time of day. Critically, although time of day was known at memory encoding, the subsequent baiting of the central location could not be predicted at encoding. Thus, to solve this task, it was necessary to always encode the location of chocolate and time of day at study, but we prevented the rats from decoding replenishment until immediately prior to memory assessment. To preferentially revisit the chocolate location when it was about to replenish at the memory assessment phase, the rats needed to remember where they found it during their earlier encoding phase. For example, for some rats, the presence of chocolate in the hub in the morning and the absence of chocolate in the hub in the afternoon allowed the rat to predict the forthcoming replenishment of chocolate. For other rats, the role played by presence and absence of food in the hub was reversed to counterbalance assignment of conditions across the rats. Because it was impossible to predict whether chocolate would replenish later, rats had to encode the episode on each study occasion. If the encoding failure hypothesis explained the results from previous studies, it would be impossible for rats to solve the current task. By contrast, if rats retrieved an episodic memory about the what-where-and-when of their earlier encounter with chocolate, they should revisit chocolate when it was about to replenish more than when it was not about to replenish. We conducted two tests of the encoding-failure hypothesis. Initially, we used a constant, minimal (2-min) retention interval. Next, we conducted a transfer test with a much longer retention interval (approximately 1 h).
Fig. 5.
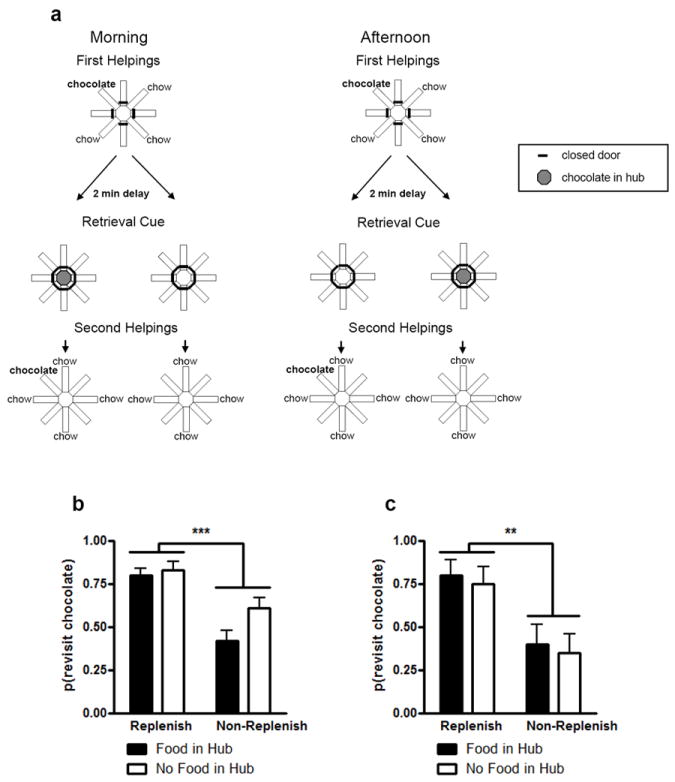
a. Schematic representation of Zhou and Crystal’s (2011) study. The morning or afternoon was randomly selected for presentation of study and test phases. An example of the accessible arms and flavors is shown in encoding and the corresponding memory assessment phases that would occur after a 2-min retention interval. The presence or absence of food in the hub, immediately prior to memory assessment, served as a cue that could be used to predict the replenishment or non-replenishment of chocolate. In the replenishment conditions, chocolate replenished at the location that recently delivered chocolate, which was predicted by the presence or absence of food (e.g., presence of chocolate in the central hub immediately prior to second helpings memory assessment in the morning but absence of chocolate in the hub in the afternoon); these contingencies were reversed in the non-replenishment conditions. These conditions were counterbalanced across rats (not shown). For each rat, one session (i.e., study phase, hub-baiting retrieval cue, and test phase) was conducted per day. Rats preferentially revisit the chocolate location when it is about to replenish when the retention interval was approximately b. 2 min and c. 1hr. The probability of a revisit to the chocolate location in the first four choices of a test phase is shown for replenishment and non-replenishment conditions. Error bars represent 1 SEM. b. *** p < 0.001 difference between replenishment and non-replenishment conditions. c. Each condition was tested once, in random order. ** p = 0.009 difference between replenishment and non-replenishment conditions. Reproduced from Zhou, W., & Crystal, J. D.(2011). Validation of a rodent model of episodic memory. Animal Cognition, 14, 325-340. © 2011 Springer-Verlag.
The results rule out the encoding failure hypothesis, suggesting that rats remember the specific earlier study episode. The rats revisited the chocolate location when it was about to replenish and avoided revisits in the non-replenishment condition (see Figures 5b and 5c). Importantly, rats were more likely to revisit the chocolate location in the replenishment conditions compared to the non-replenishment conditions. Revisit probabilities were similar for both retrieval cues and the effect of replenishment condition did not depend on the retrieval cue. Differential rates of revisiting chocolate-flavored locations were accomplished while rats accurately avoided revisits to depleted chow-flavored locations. To successfully solve this task, rats had to encode the episode at study, because the critical information about whether or not chocolate would be replenished at the recently visited location was not available until immediately before the memory assessment.
Application to prospective cognition
As described above, Roberts and Feeney (Roberts, 2012, this issue; Roberts & Feeney, 2009) have argued that it is necessary to show in prospective cognition that an animal represents a specific time in the future. It will be valuable to examine the temporal specificity of prospective memory in rats in future research. Our initial attempt to document prospective memory in rats used a broad-ranged temporal signal. We described the availability of the meal as 90-min after the start of the bisection task, and the animal may have timed the 90-min interval. However, rats may have used other temporal cues. Meals occurred at approximately a constant time of day because the daily sessions began at an approximately constant time of day. Moreover, other non-temporal cues were available to the animals including the number of food pellets earned, physiological signals of satiation, and number of trials. Additional experiments are needed to determine if anticipation of the meal was based on interval, circadian, and/or other cues.
It is currently unknown what type(s) of temporal representations are used by rats in prospective memory. The approach used by Zhou and Crystal (2009) may be deployed to examine temporal representations in prospective memory. The light cycle may be shifted and an immediate test may be used to identify the role of time of day (i.e., a circadian representation) in prospective memory; this situation is akin to the predictable bias in one’s sense of time of day that occurs immediately upon flying across several time zones. A less widely used technique is to permanently shift the light cycle and allow the animals to adjust completely to the new light regime for several days before assessing performance; this situation is akin to the elimination of temporal bias that occurs after adjusting to a new time zone. Incubation under a new light cycle in the absence of any further behavioral training predicts that the deleterious effect would shift in the direction of the light-cycle shift in this type of assessment if prospective-memory representations include information about a specific future time.
Our initial demonstration of prospective memory intentionally confounded interval timing and time of day cues to maximize the predictability of the forthcoming meal. However, some of the techniques used by Zhou and Crystal (2009) may be used to selectively provide one type of temporal cue to predict a forthcoming meal. It remains to be determined which type of temporal cue(s) are critical to the production of a prospective-memory deleterious effect on ongoing activity.
Our initial demonstration of prospective memory used two types of timing tasks - a short-interval time discrimination task (judgments about 2 vs. 8 s) and anticipation of a meal in the future. Timing two targets may be more cognitively demanding than timing a single target. Similarly, there may be important consequences of a processing bottleneck that may be unique to timing two targets. Hence, it will be important to determine if the observation of deleterious effects is limited to situations that use related tasks.
Our initial demonstration used two food-motivated tasks. The rats earned single pellets intermittently while completing the bisection task, whereas they rapidly obtained many more pellets when the meal occurred. It is possible that rats may only represent future events within a single motivational system. Alternatively, rats may represent a wide profile of future events. Similarly, deleterious effects on ongoing activity may be quite specific to a given motivational system, or alternatively may involve interactions across multiple motivational systems. Careful selection of motivational systems may allow future research to sketch out a comprehensive description of the elements of prospective cognition in rats.
Conclusions
A multi-method approach is needed to fully explore the range and limits of prospective cognition in rats. It is possible that rats have some aspects of prospective cognition, but in some significant ways it may be limited relative to prospective cognition in humans or other animals. The use of multiple approaches is likely to provide a more complete picture of the representations used in prospective cognition. This review has highlighted some directions to explore, including the temporal specificity of prospective cognition, the range of flexibility or creativity within prospective cognition, and the constraints imposed by multiple motivational systems.
Highlights.
Rats remember to take action in the future
Rats remember specific past episodes
Directions to explore include: the temporal specificity of prospective cognition;
the range of flexibility or creativity within prospective cognition;
the constraints imposed by multiple motivational systems
Acknowledgments
Supported by National Institute of Mental Health grant R01MH080052.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Aberle I, Rendell PG, Rose NS, McDaniel MA, Kliegel M. The age prospective memory paradox: Young adults may not give their best outside of the lab. Developmental Psychology. 2010;46:1444–1453. doi: 10.1037/a0020718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Addis DR, Schacter DL. Constructive episodic simulation: Temporal distance and detail of past and future events modulate hippocampal engagement. Hippocampus. 2008;18:227–237. doi: 10.1002/hipo.20405. [DOI] [PubMed] [Google Scholar]
- Babb SJ, Crystal JD. Discrimination of what, when, and where: Implications for episodic-like memory in rats. Learning & Motivation. 2005;36:177–189. [Google Scholar]
- Babb SJ, Crystal JD. Discrimination of what, when, and where is not based on time of day. Learning & Behavior. 2006a;34:124–130. doi: 10.3758/bf03193188. [DOI] [PubMed] [Google Scholar]
- Babb SJ, Crystal JD. Episodic-like memory in the rat. Current Biology. 2006b;16:1317–1321. doi: 10.1016/j.cub.2006.05.025. [DOI] [PubMed] [Google Scholar]
- Beran MJ, Perdue BM, Bramlett JL, Menzel CR, Evans TA. Prospective memory in a language-trained chimpanzee (Pan troglodytes) Learning and Motivation. 2012 doi: 10.1016/j.lmot.2012.05.002. This issue. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bird LR, Roberts WA, Abroms BD, Kit KA, Crupi C. Spatial memory for food hidden by rats (Rattus norvegicus) on the radial maze: Studies of memory for where, what, and when. Journal of Comparative Psychology. 2003;117:176–187. doi: 10.1037/0735-7036.117.2.176. [DOI] [PubMed] [Google Scholar]
- Blanco-Campal A, Coen RF, Lawlor BA, Walsh JB, Burke TE. Detection of prospective memory deficits in mild cognitive impairment of suspected Alzheimer?s disease etiology using a novel event-based prospective memory task. Journal of the International Neuropsychological Society. 2009;15:154–159. doi: 10.1017/S1355617708090127. [DOI] [PubMed] [Google Scholar]
- Burgess PW, Scott SK, Frith CD. The role of the rostral frontal cortex (area 10) in prospective memory: a lateral versus medial dissociation. Neuropsychologia. 2003;41:906–918. doi: 10.1016/s0028-3932(02)00327-5. [DOI] [PubMed] [Google Scholar]
- Cheke LG, Clayton NS. Eurasian jays (Garrulus glandarius) overcome their current desires to anticipate two distinct future needs and plan for them appropriately. Biology Letters. 2012;8:171–175. doi: 10.1098/rsbl.2011.0909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clayton NS, Bussey TJ, Dickinson A. Can animals recall the past and plan for the future? Nature Reviews Neuroscience. 2003;4:685–691. doi: 10.1038/nrn1180. [DOI] [PubMed] [Google Scholar]
- Clayton NS, Dickinson A. Episodic-like memory during cache recovery by scrub jays. Nature. 1998;395:272–274. doi: 10.1038/26216. [DOI] [PubMed] [Google Scholar]
- Correia SPC, Dickinson A, Clayton NS. Western scrub-jays anticipate future needs independently of their current motivational state. Current Biology. 2007;17:856–861. doi: 10.1016/j.cub.2007.03.063. [DOI] [PubMed] [Google Scholar]
- Craik FIM. A functional account of age differences in memory. In: Hagendorf FKH, editor. Human memory and cognitive capabilities: Mechanisms and performances. North Holland: Elsevier; 1986. pp. 409–422. [Google Scholar]
- Crystal JD. Elements of episodic-like memory in animal models. Behavioural Processes. 2009;80:269–277. doi: 10.1016/j.beproc.2008.09.009. [DOI] [PubMed] [Google Scholar]
- Crystal JD. Episodic-like memory in animals. Behavioural Brain Research. 2010;215:235–243. doi: 10.1016/j.bbr.2010.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crystal JD. Navigating the interface between learning and cognition. International Journal of Comparative Psychology. 2011;24:412–436. [PMC free article] [PubMed] [Google Scholar]
- Crystal JD. Animal models of human cognition. In: Vonk J, Shackelford T, editors. Oxford Handbook of Comparative Evolutionary Psychology. Oxford: Oxford University Press; 2012. pp. 261–270. [Google Scholar]
- d’Ydewalle G, Bouckaert D, Brunfaut E. Age-related differences and complexity of ongoing activities in time- and event-based prospective memory. American Journal of Psychology. 2001;114:411–423. [PubMed] [Google Scholar]
- Driscoll I, McDaniel MA, Guynn MJ. Apolipoprotein E and prospective memory in normally aging adults. Neuropsychology. 2005;19:28–34. doi: 10.1037/0894-4105.19.1.28. [DOI] [PubMed] [Google Scholar]
- Eacott MJ, Easton A. Remembering the past and thinking about the future: Is it really about time? Learning and Motivation. 2012 This issue. [Google Scholar]
- Eacott MJ, Easton A, Zinkivskay A. Recollection in an episodic-like memory task in the rat. Learning & Memory. 2005;12:221–223. doi: 10.1101/lm.92505. [DOI] [PubMed] [Google Scholar]
- Eacott MJ, Norman G. Integrated memory for object, place, and context in rats: A possible model of episodic-like memory? The Journal of Neuroscience. 2004;24:1948–1953. doi: 10.1523/JNEUROSCI.2975-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksen JL, Janus CG. Plaques, Tangles, and Memory Loss in Mouse Models of Neurodegeneration. Behavior Genetics. 2007;37:79–100. doi: 10.1007/s10519-006-9118-z. [DOI] [PubMed] [Google Scholar]
- Foster ER, McDaniel MA, Repovš G, Hershey T. Prospective memory in Parkinson disease across laboratory and self-reported everyday performance. Neuropsychology. 2009;23:347–358. doi: 10.1037/a0014692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimoto T, Umeda S, Kojima S. Neural substrates of implicit cueing effect on prospective memory. Neuroimage. 2011;54:645–652. doi: 10.1016/j.neuroimage.2010.07.047. [DOI] [PubMed] [Google Scholar]
- Henry JD, MacLeod MS, Phillips LH, Crawford JR. A meta-analytic review of prospective memory and aging. Psychology and Aging. 2004;19:27–39. doi: 10.1037/0882-7974.19.1.27. [DOI] [PubMed] [Google Scholar]
- Henry JD, Phillips LH, Crawford JR, Kliegel M, Theodorou G, Summers F. Traumatic brain injury and prospective memory: Influence of task complexity. Journal of Clinical and Experimental Neuropsychology. 2007;29:457–466. doi: 10.1080/13803390600762717. [DOI] [PubMed] [Google Scholar]
- Hicks JL, Marsh RL, Cook GI. Task interference in time-based, event-based, and dual intention prospective memory conditions. Journal of Memory and Language. 2005;53:430–444. doi: 10.1016/j.jml.2005.04.001. [DOI] [Google Scholar]
- Hwang DY, Cho JS, Lee SH, Chae KR, Lim HJ, Min SH, Seo SJ, Song YS, Song CW, Paik SG, Sheen YY, Kim YK. Aberrant expressions of pathogenic phenotype in Alzheimer’s diseased transgenic mice carrying NSE-controlled APPsw. Experimental Neurology. 2004;186:20–32. doi: 10.1016/j.expneurol.2003.09.021. [DOI] [PubMed] [Google Scholar]
- Jankowsky JL, Slunt HH, Gonzales V, Savonenko AV, Wen JC, Jenkins NA, Copeland NG, Younkin LH, Lester HA, Younkin SG, Borchelt DR. Persistent Amyloidosis following Suppression of Aβ Production in a Transgenic Model of Alzheimer Disease. PLoS Medicine. 2005;2:e355–1333. doi: 10.1371/journal.pmed.0020355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones S, Livner Å, Bäckman L. Patterns of prospective and retrospective memory impairment in preclinical Alzheimer’s disease. Neuropsychology. 2006;20:144–152. doi: 10.1037/0894-4105.20.2.144. [DOI] [PubMed] [Google Scholar]
- Kart-Teke E, De Souza Silva MA, Huston JP, Dere E. Wistar rats show episodic-like memory for unique experiences. Neurobiology of Learning and Memory. 2006;85:173–182. doi: 10.1016/j.nlm.2005.10.002. [DOI] [PubMed] [Google Scholar]
- Keri RA, Siegel RE, Donald WP, Arthur PA, Susan EF, Anne ME, Robert TR. Transgenic and Genetic Animal Models. In: Donald APA, Pfaff W, Fahrbach Susan E, Etgen Anne M, Rubin Robert T, editors. Hormones, Brain and Behavior. San Diego: Academic Press; 2009. pp. 2673–2708. [Google Scholar]
- Kliegel M, Martin M, McDaniel MA, Einstein GO. Varying the importance of a prospective memory task: Differential effects across time - and event-based prospective memory. Memory. 2001;9:1–11. doi: 10.1080/09658210042000003. [DOI] [PubMed] [Google Scholar]
- Marsh RL, Hicks JL, Cook GI. Task interference from prospective memories covaries with contextual associations of fulfilling them. Memory & Cognition. 2006;34:1037–1045. doi: 10.3758/bf03193250. [DOI] [PubMed] [Google Scholar]
- Marsh RL, Hicks JL, Landau JD. An investigation of everyday prospective memory. Memory & Cognition. 1998;26:633–643. doi: 10.3758/bf03211383. [DOI] [PubMed] [Google Scholar]
- Martin VC, Schacter DL, Corballis MC, Addis DR. A role for the hippocampus in encoding simulations of future events. Proceedings of the National Academy of Sciences. 2011;108:13858–13863. doi: 10.1073/pnas.1105816108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maxwell MM. RNAi applications in therapy development for neurodegenerative disease. Current Pharmaceutical Design. 2009;15:3977–3991. doi: 10.2174/138161209789649295. [DOI] [PubMed] [Google Scholar]
- McCauley SR, McDaniel MA, Pedroza C, Chapman SB, Levin HS. Incentive effects on event-based prospective memory performance in children and adolescents with traumatic brain injury. Neuropsychology. 2009;23:201–209. doi: 10.1037/a0014192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDaniel MA, Einstein GO. Prospective memory: An overview and synthesis of an emerging field. Thousand Oaks, CA: Sage Publications; 2007. [Google Scholar]
- Naqshbandi M, Feeney MC, McKenzie TLB, Roberts WA. Testing for episodic-like memory in rats in the absence of time of day cues: Replication of Babb and Crystal. Behavioural Processes. 2007;74:217–225. doi: 10.1016/j.beproc.2006.10.010. [DOI] [PubMed] [Google Scholar]
- Naqshbandi M, Roberts WA. Anticipation of Future Events in Squirrel Monkeys (Saimiri sciureus) and Rats (Rattus norvegicus): Tests of the Bischof-Kohler Hypothesis. Journal of Comparative Psychology. 2006;120:345–357. doi: 10.1037/0735-7036.120.4.34. 10.103710735-7036. 20.4.345. [DOI] [PubMed] [Google Scholar]
- Raby CR, Alexis DM, Dickinson A, Clayton NS. Planning for the future by western scrub-jays. Nature. 2007;445:919–921. doi: 10.1038/nature05575. [DOI] [PubMed] [Google Scholar]
- Raskin SA, Woods SP, Poquette AJ, McTaggart AB, Sethna J, Williams RC, Tröster AI. A differential deficit in time- versus event-based prospective memory in Parkinson’s disease. Neuropsychology. 2011;25:201–209. doi: 10.1037/a0020999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts WA. Evidence for future cognition in animals. Learning and Motivation. 2012 This issue. [Google Scholar]
- Roberts WA, Feeney MC. The comparative study of mental time travel. Trends in Cognitive Sciences. 2009;13:271–277. doi: 10.1016/j.tics.2009.03.003. [DOI] [PubMed] [Google Scholar]
- Roberts WA, Feeney MC, MacPherson K, Petter M, McMillan N, Musolino E. Episodic-like memory in rats: Is it based on when or how long ago? Science. 2008;320:113–115. doi: 10.1126/science.1152709. [DOI] [PubMed] [Google Scholar]
- Roberts WA, Roberts S. Two tests of the stuck in-time hypothesis. Journal of General Psychology. 2002;129:415–429. doi: 10.1080/00221300209602105. [DOI] [PubMed] [Google Scholar]
- Schacter DL, Addis DR. The cognitive neuroscience of constructive memory: remembering the past and imagining the future. Philosophical Transactions of the Royal Society B: Biological Sciences. 2007;362:773–786. doi: 10.1098/rstb.2007.2087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schacter DL, Addis DR, Buckner RL. Remembering the past to imagine the future: the prospective brain. Nature Reviews Neuroscience. 2007;8:657–661. doi: 10.1038/nrn2213. [DOI] [PubMed] [Google Scholar]
- Schacter DL, Addis DR, Buckner RL. Episodic Simulation of Future Events. Annals of the New York Academy of Sciences. 2008;1124:39–60. doi: 10.1196/annals.1440.001. [DOI] [PubMed] [Google Scholar]
- Schmitter-Edgecombe M, Woo E, Greeley DR. Characterizing multiple memory deficits and their relation to everyday functioning in individuals with mild cognitive impairment. Neuropsychology. 2009;23:168–177. doi: 10.1037/a0014186. [DOI] [PubMed] [Google Scholar]
- Simons JS, Schölvinck ML, Gilbert SJ, Frith CD, Burgess PW. Differential components of prospective memory?: Evidence from fMRI. Neuropsychologia. 2006;44:1388–1397. doi: 10.1016/j.neuropsychologia.2006.01.005. [DOI] [PubMed] [Google Scholar]
- Smith RE. The cost of remembering to remember in event-based prospective memory: investigating the capacity demands of delayed intention performance. Journal of Experimental Psychology-Learning Memory and Cognition. 2003;29:347–361. doi: 10.1037/0278-7393.29.3.347. [DOI] [PubMed] [Google Scholar]
- Smith RE, Hunt RR, McVay JC, McConnell MD. The cost of event-based prospective memory: Salient target events. Journal of Experimental Psychology-Learning Memory and Cognition. 2007;33:734–746. doi: 10.1037/0278-7393.33.4.734. [DOI] [PubMed] [Google Scholar]
- Suddendorf T, Corballis MC. Mental time travel and the evolution of the human mind. Genetic, Social, & General Psychology Monographs. 1997;123:133–167. [PubMed] [Google Scholar]
- Suddendorf T, Corballis MC. The evolution of foresight: What is mental time travel, and is it unique to humans? Behavioral and Brain Sciences. 2007;30:299–313. doi: 10.1017/S0140525X07001975. [DOI] [PubMed] [Google Scholar]
- Troyer AK, Murphy KJ. Memory for intentions in amnestic mild cognitive impairment: Time- and event-based prospective memory. Journal of the International Neuropsychological Society. 2007;13:365–369. doi: 10.1017/s1355617707070452. [DOI] [PubMed] [Google Scholar]
- Ueberham U, Zobiak B, Ueberham E, Brückner MK, Boriss H, Arendt T. Differentially expressed cortical genes contribute to perivascular deposition in transgenic mice with inducible neuron-specific expression of TGF-β1. International Journal of Developmental Neuroscience. 2006;24:177–186. doi: 10.1016/j.ijdevneu.2005.11.007. [DOI] [PubMed] [Google Scholar]
- Wilson AG, Crystal JD. Prospective memory in the rat. Animal Cognition. 2012;15:349–358. doi: 10.1007/s10071-011-0459-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zentall TR. Animals may not be stuck in time. Learning & Motivation. 2005;36:208–225. [Google Scholar]
- Zentall TR. Mental time travel in animals: A challenging question. Behavioural Processes. 2006;72:173–183. doi: 10.1016/j.beproc.2006.01.009. [DOI] [PubMed] [Google Scholar]
- Zentall TR. Coding of stimuli by animals: Retrospection, prospection, episodic memory and future planning. Learning and Motivation. 2010;41:225–240. doi: 10.1016/j.lmot.2010.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou W, Crystal JD. Evidence for remembering when events occurred in a rodent model of episodic memory. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:9525–9529. doi: 10.1073/pnas.0904360106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou W, Crystal JD. Validation of a rodent model of episodic memory. Animal Cognition. 2011;14:325–340. doi: 10.1007/s10071-010-0367-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou W, Hohmann AG, Crystal JD. Rats answer an unexpected question after incidental encoding. Current Biology. doi: 10.1016/j.cub.2012.04.040. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]