

Abstract
Please cite this paper as: Ducatez et al. (2012) Long‐term vaccine‐induced heterologous protection against H5N1 influenza viruses in the ferret model. Influenza and Other Respiratory Viruses 7(4), 506–512.
Background Highly pathogenic H5N1 influenza viruses reemerged in humans in 2003 and have caused fatal human infections in Asia and Africa as well as ongoing outbreaks in poultry. These viruses have evolved substantially and are now so antigenically varied that a single vaccine antigen may not protect against all circulating strains. Nevertheless, studies have shown that substantial cross‐reactivity can be achieved with H5N1 vaccines. These studies have not, however, addressed the issue of duration of such cross‐reactive protection.
Objectives To directly address this using the ferret model, we used two recommended World Health Organization H5N1 vaccine seed strains – A/Vietnam/1203/04 (clade 1) and A/duck/Hunan/795/02 (clade 2.1) – seven single, double, or triple mutant viruses based on A/Vietnam/1203/04, and the ancestral viruses A and D, selected from sequences at nodes of the hemagglutinin and neuraminidase gene phylogenies to represent antigenically diverse progeny H5N1 subclades as vaccine antigens.
Results All inactivated whole‐virus vaccines provided full protection against morbidity and mortality in ferrets challenged with the highly pathogenic H5N1 strain A/Vietnam/1203/04 5 months and 1 year after immunization.
Conclusion If an H5N1 pandemic was to arise, and with the hypothesis that one can extrapolate the results from three doses of a whole‐virion vaccine in ferrets to the available split vaccines for use in humans, the population could be efficiently immunized with currently available H5N1 vaccines, while the homologous vaccine is under production.
Keywords: Ferret, H5N1, influenza virus, pandemic, vaccine
Introduction
Highly pathogenic avian influenza (HPAI) A(H5N1) viruses continue to pose a pandemic threat. 1 Since its first detection in Guangdong, China, in 1996 (A/goose/Guangdong/1/1996 2 ), the pathogen has spread throughout Asia, Europe, and Africa. As of February 2012, it had caused more than 7000 outbreaks in poultry in 51 countries 3 and 583 human cases (344 of them fatal). 4
During its 15 years of circulation, the virus has evolved extensively at both genetic and antigenic levels, confounding pandemic preparedness efforts and the selection of a single vaccine seed. H5N1 lineages have now been classified into 10 phylogenetic clades (0–9) based on their hemagglutinin (HA) sequences, 5 and the World Health Organization has developed 17 candidate vaccine strains and has another five pending. 6 Because the rapid mutation of the virus poses the risk of development of drug resistance, vaccination remains the preferred pandemic preparedness strategy.
A few cross‐clade vaccine protection studies have been conducted in the ferret model, using adjuvanted or non‐adjuvanted whole‐virus or split vaccines. A/Hong Kong/213/03 (clade 1), A/Vietnam/1203/04 (VN1203, clade 1), A/Japanese white eyes/Hong Kong/1038/06 (clade 2.3.4), and A/Vietnam/1194/04 (clade 1) vaccines protected ferrets wholly or partially against mortality after challenge with A/Hong Kong/156/97 (clade 0), A/Indonesia/5/05 (clade 2.1), VN1203, and A/Indonesia/5/05 viruses, respectively. 7 We recently reported the use of the ‘most recent common ancestor’ computational method to design cross‐clade‐protective H5N1 vaccines containing ancestral HA antigens A or D, VN1203, or A/duck/Hunan/795/02 (DKHUN795, clade 2.1). These four vaccines fully protected ferrets against challenge with VN1203, DKHUN795, and A/turkey/Egypt/7/07 (clade 2.2.1) viruses. 8 Clinical trials in humans have shown promising antigenic cross‐clade reactivity: vaccination with adjuvanted A/duck/Singapore/1997 (H5N3) and boost with adjuvanted A/Vietnam/1194/2004 (clade 1) vaccines induced immune responses to clade 0, 1, and 2 viruses. 9
Several studies have suggested pre‐pandemic vaccination of populations as a use for stockpiled vaccines. 9 , 10 , 11 , 12 , 13 The success of such an approach, however, would rely directly on the duration of cross‐reactive immunity, something that has not been adequately measured in the H5N1 model. To conserve resources, most research teams indeed challenge the animals 2–4 weeks after the final immunization. However, two groups recently reported long‐term protection of ferrets against homologous challenge. Baras et al. 14 challenged ferrets with A/Indonesia/5/05 virus 10, 16, or 19 weeks after vaccination and observed various levels of protection, depending on the number of vaccine doses and the use of adjuvant. An adjuvanted A/Vietnam/1194/2004 vaccine was found to protect ferrets against homologous challenge for at least 15 months. 15 Long‐term heterologous protection has been shown only in mice, 5 months after immunization with a virus‐like particle vaccine containing A/Puerto Rico/8/34 (H1N1) HA and matrix 1 proteins 16 and 1 year after vaccination with a vesicular stomatitis virus‐based avian influenza vaccine. 17
It is not possible to predict which virus clade might produce a future pandemic strain. Further, it would take an estimated 2 months after the emergence of a pandemic to produce the vaccine seed, another 3 months to produce the first vaccine doses, 18 and many more months to generate a sufficient supply, 19 even in the absence of technical difficulties. 18 However, if long‐lasting heterologous protection can be induced, the population could be rapidly primed with currently available vaccine antigens in the event of an H5N1 pandemic. Here, we investigated long‐term H5N1 heterologous protection in the ferret model. We vaccinated the ferrets with a number of whole‐virus A(H5N1) vaccines and challenged them with VN1203 5 months and 1 year after the initial vaccination.
Materials and methods
Ferret immunization and challenge
Young adult male ferrets of 3–4 months of age and seronegative for currently circulating influenza A H1N1, H3N2, and influenza B viruses were obtained from Triple F farms (Sayre, PA, USA). Three doses of vaccine (0·25 ml each) were injected intramuscularly at 3‐week intervals into groups of four ferrets. Each dose contained approximately 7·5 μg of HA, giving a total of approximately 22·5 μg HA protein given to each ferret. Vaccines were based on reverse genetics (rg)‐generated 6+2 VN1203 (clade 1), DKHUN795 (clade 2.1), or ancestral A or D HA viruses. A and D were selected from sequences at nodes of the HA and NA gene phylogenies to represent antigenically diverse progeny H5N1 subclades. 8 In this previous work, we had detected slight increases in the immunogenicity of the D and DKHUN795 antigens as compared with the A and VN1203 antigens. We therefore also included VN1203 antigens with specific HA mutations common to the D and DKHUN795 antigens to avoid any potential issues with differences in immunogenicity. The viruses produced were VN1203 D94N, VN1203 S124D, VN1203 K189R, VN1203 D94N:S124D, VN1203 D94N:K189R, VN1203 S124D:K189R, and VN1203 D94N:S124D:K189R. The vaccines were prepared as previously described. 8 The total protein content of the vaccines was assayed (Bio‐Rad, Hercules, CA, USA) using the Bradford method 20 ; HA was estimated to make up approximately 25% of the total protein content of our H5 vaccines.
A group of four ferrets was vaccinated with each vaccine. Four control ferrets were injected with PBS only. Five months or 1 year after the first dose, ferrets were anesthetized with isoflurane (respiratory route) and inoculated intranasally with 106 egg infectious dose 50 (EID50) of the HPAI H5N1 challenge virus, VN1203, in 1 ml of sterile PBS in a BSL3+ laboratory. In total, four animals were available for serology assays prior to challenge, and two ferrets per group were challenged either 5 months or a year after first immunization. Weight, temperature, and clinical disease signs were monitored daily for 14 days. Body temperature was measured via transponders subcutaneously implanted between the shoulder blades (BioMedic Data Systems, Inc., Seaford, DE, USA). This study adhered to the guidelines of the St. Jude Children’s Research Hospital Animal Care and Use Committee.
Microneutralization assay and enzyme‐linked immunosorbent assay
Filtered sera were tested by microneutralization (MN) assay in Madin‐Darby canine kidney (MDCK) cells as previously described. 21 Neutralizing titers were expressed as the reciprocal of the serum dilution that inhibited 50% of the growth of 100 50% tissue culture infectious doses (TCID50) of virus. Enzyme‐linked immunosorbent assays (ELISA) were carried out as described previously 8 using 50 ng per well of A/Vietnam/1203/04 (aa 18‐530, eEnzyme LLC, Montgomery Village, MD, USA) as coating antigen.
Nasal washes
On days 1, 3, 5, and 7 after virus inoculation, ferrets were anesthetized with ketamine (intramuscular administration, 25 mg/kg; Hospira Inc., Lake Forest, IL, USA), and nasal washes were collected by standard procedures. Bovine serum albumin (Sigma, St. Louis, MO, USA) was added at a ratio of 1:20 v/v as a stabilizing agent. Virus was titrated in embryonated chicken eggs, and the log10 EID50/ml was calculated by the method of Reed and Muench. 22 The lower limit of virus detection was 0·75 log10 EID50/ml. A value of 0 (log101) was assigned to titers below the lower limit of detection.
Results
Long‐term heterologous immunogenicity of the H5N1 vaccines
To assess the long‐term immune responses induced by vaccination with the two ancestral viruses antigens A and D, the reference H5N1 isolates VN1203 (clade 1) and DKHUN795 (clade 2.1), and VN1203 mutants, we measured the cross‐reactivity of their antisera toward VN1203 by MN assay. The VN1203‐vaccinated animals showed MN titers ≥40 after the initial vaccination. Ferrets vaccinated with DKHUN795, A, and D antigens had much lower initial MN titers against VN1203, as expected (Table 1). The peak MN titer for all vaccines was observed after the second boost dose (week 8 after initial vaccination), at which time DKHUN795 and D antigen induced titers ≥40. However, while ferrets vaccinated with VN1203 maintained the same homologous MN titers at 1 year post‐vaccination, ferrets vaccinated with the other vaccines did not, and ferrets vaccinated with antigen A did not show protective titers at any point (Table 1). The ferrets vaccinated with the three heterologous vaccines had undetectable to very low MN titers against VN1203 (0–15; Table 1) at the time of challenge (5 months or 1 year after initial vaccination). Similarly, ELISA titers to VN1203 just before challenge ranged from undetectable to 20 for the ferrets immunized with heterologous vaccines and from 40 to 80 for the VN1203 immunized animals. Animals vaccinated with VN1203 HA mutants had an intermediate pre‐challenge serological response: MN titers ranged from 20 to 40 after the first immunization, peak titers were observed after the third vaccine boost and ranged from 40 to 640, and low titers (undetectable to 80) were measured right before challenge (Table 1).
Table 1.
Pre‐challenge serology
| Vaccine regimen* | Weeks after first immunization | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 6 | 8 | 10 | 12 | 16 | 20 | 51 | |||||||||
| VN1203 | 52·5 | 52·5 | 260 | 260 | 580 | 580 | 100 | 100 | 150 | 150 | 135 | 135 | 180 | 180 | 180 | 180 |
| VN1203 HA D94N | 12·5 | 22·5 | 35 | 40 | 140 | 230 | 52·5 | 60 | 35 | 70 | 12·5 | 30 | 27·5 | 32·5 | 20 | 20 |
| VN1203 HA S124D | 12·5 | 25 | 30 | 80 | 90 | 560 | 60 | 240 | 40 | 90 | 10 | 50 | 17·5 | 45 | 10 | 15 |
| VN1203 HA K189R | 12·5 | 25 | 100 | 130 | 160 | 140 | 80 | 60 | 40 | 70 | 20 | 25 | 27·5 | 27·5 | 25 | 22·5 |
| VN1203 HA D94N:S124D | 17·5 | 30 | 90 | 110 | 280 | 480 | 130 | 90 | 55 | 90 | 10 | 17·5 | 35 | 65 | 30 | 60 |
| VN1203 HA D94N:K189R | 17·5 | 35 | 120 | 30 | 300 | 110 | 110 | 60 | 85 | 45 | 35 | 10 | 55 | 30 | 20 | 5 |
| VN1203 HA S124D:K189R | 20 | 25 | 90 | 72·5 | 360 | 320 | 200 | 55 | 160 | 60 | 70 | 32·5 | 60 | 27·5 | 50 | 20 |
| VN1203 HA D94N:S124D:K189R | 17·5 | 20 | 90 | 35 | 160 | 120 | 150 | 20 | 120 | 55 | 35 | 17·5 | 75 | 45 | 20 | 7·5 |
| DKHUN795 | 22·5 | 20 | 70 | 22·5 | 340 | 55 | 110 | 10 | 130 | 27·5 | 62·5 | 17·5 | 35 | 15 | 25 | 5 |
| A | 12·5 | 22·5 | 45 | 7·5 | 640 | 27·5 | 120 | 10 | 140 | 22·5 | 45 | 0 | 65 | 7·5 | 60 | 0 |
| D | 15 | 17·5 | 85 | 7·5 | 680 | 42·5 | 160 | 16·7 | 240 | 20 | 120 | 12·5 | 120 | 7·5 | 180 | 5 |
| PBS | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Mean microneutralization titers (n = 4 ferrets except at week 51 n = 2 ferrets) against the homologous strain in plain font and against VN1203 (the challenge virus) in bold, italic font.
HA, hemagglutinin.
*Ferrets were immunized on weeks 0, 3, and 6 (after the bleed weeks 3 and 6).
Long‐term protection of ferrets in vivo
We tested the long‐term protectiveness induced by the homologous vaccine antigen VN1203, VN1203 HA mutants, and of the heterologous vaccine antigens DKHUN795 and ancestral strains A and D against VN1203. Groups of vaccinated ferrets were challenged with wild‐type VN1203 (H5N1) virus 5 months or 1 year after the initial vaccination. All control (PBS‐vaccinated) ferrets died or developed paralysis in the hind legs and were euthanized within 5 days post‐inoculation, while all vaccinated animals survived irrespective of the vaccine and time of challenge. These homologous and heterologous H5N1 vaccines therefore protected ferrets from mortality for at least 1 year after initial vaccination. Further, none of the vaccinated animals showed clinical signs of disease, although control ferrets were lethargic and had diarrhea, and some had nasal discharge and/or neurologic complications (data not shown). Vaccinated ferrets lost 0–6·5% of their body weight post‐challenge, irrespective of the vaccine and time of challenge, but weight loss did not differ substantially according to vaccine or time of challenge (Table 2). Maximum body temperature did not differ between vaccine regimens, although fever was higher after the 1‐year challenge than after the 5‐month challenge (groups were too small to allow meaningful statistical comparison). Control (PBS‐vaccinated) animals lost 16%–23% of their body weight and had an increase of 1·2–1·7°C in body temperature 1 day post‐inoculation (Table 2). Therefore, all of the test vaccines protected ferrets against mortality and reduced morbidity after challenge with VN1203.
Table 2.
Clinical signs in vaccinated ferrets after lethal challenge with VN1203 (H5N1) virus
| Vaccine | Maximum % weight loss within 10 days of inoculation* | Maximum temperature increase (°C) 1 day after inoculation*,** | ||
|---|---|---|---|---|
| Challenge after first vaccination | Challenge after first vaccination | |||
| 5 months | 1 year | 5 months | 1 year | |
| VN1203 | 4·3 | 5·4*** | 0·1 | 1·4 |
| DKHUN795 | 4·6 | 6·5 | 0·0 | 1·4 |
| A | 4·3 | 4·0 | 0·6 | 1·4 |
| D | 0·0 | 2·1 | 0·3 | 1·0 |
| PBS† | 23·0 | 16·0 | 1·2 | 1·7 |
*Mean maximum weight loss or temperature increase per group (n = 2 per group).
**Highest temperature was observed 1 day post‐inoculation in all groups.
***N = 1 in this group only.
†All PBS‐vaccinated animals died or were euthanized by day 5 post‐inoculation.
After challenge at 1 year, nasal wash titers differed between vaccinated and control ferrets at days 3 and 5 after inoculation (Figure 1). After challenge at 5 months, the same general trend was observed, but only ferrets vaccinated with VN1203 or DKHUN795 had lower nasal wash titers than control animals on day 5 post‐inoculation. Again, because of the small number of animals per group, statistical analysis was not meaningful. All ferrets but one vaccinated with DKHUN795 and challenged 1 year later cleared the virus by day 7 (Figure 1). The VN1203 HA mutant‐vaccinated animals followed the same pattern as VN1203‐, DKHUN795‐, A‐, and D‐vaccinated ferrets: all vaccine antigens induced an appropriate long‐term protective response.
Figure 1.
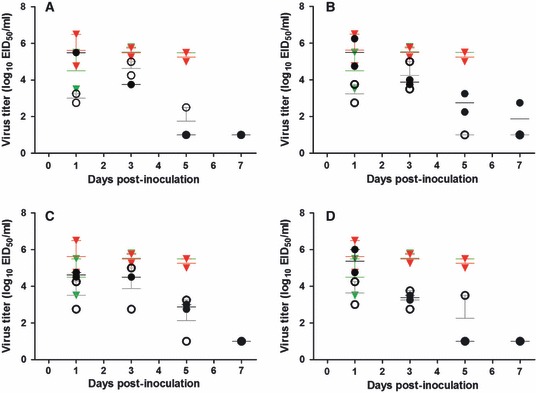
Ferrets’ nasal wash titers after challenge with VN1203. Post‐challenge virus titers of ferrets vaccinated with (A) VN1203 (clade 1), (B) DKHUN795 (clade 2.1), (C) ancestral A antigen, or (D) ancestral D antigen are expressed as mean log10 EID50/ml (horizontal lines) ± standard error (vertical lines). Vaccinated ferrets are represented by black (challenged at 1 year) and gray (challenged at 5 months) circles. Control (PBS‐vaccinated) animals are represented by red (challenged at 1 year) and green (challenged at 5 months) symbols. All control ferrets died or were euthanized before day 7 post‐challenge.
Discussion
The development of pre‐pandemic H5N1 vaccines has been a hallmark of some countries preparedness for a potential pandemic caused by these viruses. In the absence of a fully human transmissible H5N1, how such vaccines might be used is not entirely clear. One suggestion has been that these vaccines be used to prime the population in advance of an outbreak. Supporting this suggested use is evidence from human clinical trials, showing that vaccinating with one H5N1 antigen can provide substantial priming for a subsequent immunization with an antigenically distinct H5N1 antigen. Goji et al. 23 indeed reported higher hemagglutination inhibition (HI) titers in subjects primed with a dose of A/Hong Kong/156/1997 (clade 0) vaccine and given a boost dose of A/Vietnam/1203/2004 (clade 1) vaccine than in subjects who received one or two doses of A/Vietnam/1203/2004 alone (about ½ less responders with a fourfold increase in antibody level to a titer ≥40). Two studies have investigated the benefit of using adjuvants for H5N1 pre‐pandemic vaccine priming. MF59 was shown to increase the antibody response to A/duck/Singapore/97 (H5N3) vaccine as measured by HI titers to viruses of different H5N1 clades. 24 Leroux‐Roels et al. 25 reported that priming with AS03(A)‐adjuvanted A/Vietnam/1194/2004 (clade 1) vaccine followed by a boost dose of AS03(A)‐adjuvanted A/Indonesia/5/2005 (clade 2.1) 14 months later induced a more robust and more rapid immune response than two doses of AS03(A)‐adjuvanted A/Indonesia/5/2005 vaccine. Three doses of an alum‐adjuvanted inactivated whole‐virus H5N1 vaccine were also shown to significantly boost the immune response in humans. 26 Taken together, these studies support the benefit of a priming strategy for H5N1 pandemic preparedness. The other potential benefit of pre‐priming the human population with an H5N1 vaccine is that it could provide some level of cross‐protection against future infection, albeit with an antigenically distinct virus. The underlying mechanisms of the observed cross‐protection are not yet fully understood. B cells play a role in the heterosubtypic immunity to influenza infection. 27 Inactivated whole‐virus vaccines also produce a good cross‐clade cellular immune response, 28 which may well explain the cross‐clade protection observed in the present study. CD4+ T cells have indeed been shown to recognize H5N1 viral antigens in human: peptides from matrix 1, nucleocapsid, neuraminidase, and H5 HA proteins. 29 , 30 , 31 A key variable in such an approach is the duration of cross‐protective immunity, a variable that we measured in this study. Our results suggest that H5N1 vaccine‐induced cross‐protective response can indeed be long‐lived.
The effectiveness of vaccine priming can be confirmed only in human clinical trials. However, using the ferret model, which closely recapitulates human influenza infection, 7 we were able to assess the response to lethal challenge. The limits of the extrapolation of our study reside in the small number of animals used (four ferrets per vaccine regimen, two of which challenged at a given time point). The number of vaccines tested (11), all providing protection against morbidity and mortality 5 months and 1 year after initial immunization of ferrets, however, allows confidence on the long‐term heterologous protection conferred by inactivated whole‐virus H5N1 vaccines. We found that even ferrets with undetectable MN titers were protected against heterologous H5N1 viruses for at least 1 year post‐immunization. These results suggest that in the event of an H5N1 pandemic, the population could be protected by priming with an available heterologous vaccine strain until a homologous vaccine is available. There are, however, caveats to the extrapolation of our study to pandemic preparedness. Foremost, we administered three doses of vaccine to the ferrets to allow for a better serological comparison between animal groups prior to challenge, a scenario that would very unlikely occur in a pandemic situation. Additionally, most current inactivated vaccines available for human use are split or subunit products, but these are less immunogenic than whole‐virus vaccines and may trigger a different outcome. A priming vaccine should also be optimized to ensure maximal effect. Leroux‐Roels et al. 25 recently observed a low anti‐H5 antibody response after priming without adjuvant and boosting with a heterologous strain, suggesting that an adjuvant should be added to the available priming vaccine.
We observed little correlation between pre‐challenge MN and ELISA titers and protection. Most of the ferrets vaccinated with the DKHUN795, A, or D antigens had undetectable MN titers against VN1203 before challenge (and none had a titer ≥40 in MN or ELISA), but all were protected against mortality and morbidity. Non‐neutralizing antibodies that are not directed against HA (and that hence would not have been detected by MN, HI, or HA‐specific ELISA) may contribute to the protective effect. 32 , 33 Although the pre‐challenge MN titer would be expected to predict the level of protection, our results and others reported previously 8 , 9 , 10 , 11 suggest otherwise.
In summary, we demonstrated that long‐term homologous and heterologous protection against HPAI virus is conferred by whole‐virus inactivated vaccines in the ferret model. While the risk/benefit ratio of priming against a putative H5N1 pandemic may not be currently favorable, the long‐term protection conferred by heterologous priming can provide a crucial public health benefit if such a pandemic arises.
Acknowledgements
We thank Jennifer DeBeauchamp (St. Jude Department of Infectious Diseases) for help in vaccinating animals; Lisa Kercher, David Carey, Beth Little, and William Martin (St. Jude Animal Resource Center) for excellent technical assistance; and Sharon Naron (St. Jude) for editing the manuscript. This work was supported by Contract No. HHSN266200700005C from the National Institute of Allergy and Infectious Disease, National Institute of Health, Department of Health and Human Services, and by the American Lebanese Syrian Associated Charities (ALSAC).
References
- 1. Yen HL, Webster RG. Pandemic influenza as a current threat. Curr Top Microbiol Immunol 2009; 333:3–24. [DOI] [PubMed] [Google Scholar]
- 2. Xu X, Subbarao K, Cox NJ, Guo Y. Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virology 1999; 261:15–19. [DOI] [PubMed] [Google Scholar]
- 3. World Organization for Animal Health . Update on highly pathogenic avian influenza in animals (type H5 and H7). Available at http://www.oie.int/downld/AVIAN%20INFLUENZA/A_AI‐Asia.htm (Accessed 6 February 2012).
- 4. World Health Organization . Cumulative number of confirmed human cases of avian influenza A/(H5N1) reported to WHO. 2011. Available at http://www.who.int/influenza/human_animal_interface/H5N1_cumulative_table_archives/en/index.html (Accessed 6 February 2012).
- 5. Donis RO, Smith GJ, Perdue ML et al. Toward a unified nomenclature system for highly pathogenic avian influenza virus (H5N1). Emerg Infect Dis 2008; 14:e1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. World Health Organization . Antigenic and genetic characteristics of influenza A(H5N1) and influenza A(H9N2) viruses for development of candidate vaccines viruses for pandemic preparedness – February 2011. Wkly Epidemiol Rec 2011; 86:93–100. [PubMed] [Google Scholar]
- 7. Govorkova EA, Webby RJ, Humberd J, Seiler JP, Webster RG. Immunization with reverse‐genetics‐produced H5N1 influenza vaccine protects ferrets against homologous and heterologous challenge. J Infect Dis 2006; 194:159–167. [DOI] [PubMed] [Google Scholar]
- 8. Ducatez MF, Bahl J, Griffin Y et al. Feasibility of reconstructed ancestral H5N1 influenza viruses for cross‐clade protective vaccine development. Proc Natl Acad Sci USA 2011; 108:349–354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Lipatov AS, Hoffmann E, Salomon R, Yen HL, Webster RG. Cross‐protectiveness and immunogenicity of influenza A/Duck/Singapore/3/97(H5) vaccines against infection with A/Vietnam/1203/04(H5N1) virus in ferrets. J Infect Dis 2006; 194:1040–1043. [DOI] [PubMed] [Google Scholar]
- 10. Hoschler K, Gopal R, Andrews N et al. Cross‐neutralisation of antibodies elicited by an inactivated split‐virion influenza A/Vietnam/1194/2004 (H5N1) vaccine in healthy adults against H5N1 clade 2 strains. Influenza Other Respi Viruses 2007; 1:199–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Levie K, Leroux‐Roels I, Hoppenbrouwers K et al. An adjuvanted, low‐dose, pandemic influenza A (H5N1) vaccine candidate is safe, immunogenic, and induces cross‐reactive immune responses in healthy adults. J Infect Dis 2008; 198:642–649. [DOI] [PubMed] [Google Scholar]
- 12. Lu X, Edwards LE, Desheva JA et al. Cross‐protective immunity in mice induced by live‐attenuated or inactivated vaccines against highly pathogenic influenza A (H5N1) viruses. Vaccine 2006; 24:6588–6593. [DOI] [PubMed] [Google Scholar]
- 13. Suguitan AL Jr, McAuliffe J, Mills KL et al. Live, attenuated influenza A H5N1 candidate vaccines provide broad cross‐protection in mice and ferrets. PLoS Med 2006; 3:e360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Baras B, Stittelaar KJ, Kuiken T et al. Longevity of the protective immune response induced after vaccination with one or two doses of AS03A‐adjuvanted split H5N1 vaccine in ferrets. Vaccine 2011; 29:2092–2099. [DOI] [PubMed] [Google Scholar]
- 15. Middleton D, Rockman S, Pearse M et al. Evaluation of vaccines for H5N1 influenza virus in ferrets reveals the potential for protective single‐shot immunization. J Virol 2009; 83:7770–7778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Quan FS, Huang C, Compans RW, Kang SM. Virus‐like particle vaccine induces protective immunity against homologous and heterologous strains of influenza virus. J Virol 2007; 81:3514–3524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Schwartz JA, Buonocore L, Suguitan AL Jr et al. Potent vesicular stomatitis virus‐based avian influenza vaccines provide long‐term sterilizing immunity against heterologous challenge. J Virol 2010; 84:4611–4618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Wood JS, Robertson JS. Reference viruses for seasonal and pandemic influenza vaccine preparation. Influenza Other Respi Viruses 2007; 1:5–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Abelin A, Colegate T, Gardner S, Hehme N, Palache A. Lessons from pandemic influenza A(H1N1): the research‐based vaccine industry’s perspective. Vaccine 2011; 29:1135–1138. [DOI] [PubMed] [Google Scholar]
- 20. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein‐dye binding. Anal Biochem 1976; 72:248–254. [DOI] [PubMed] [Google Scholar]
- 21. World Health Organization . WHO manual on animal diagnosis and surveillance. 2011. Available at http://whqlibdoc.who.int/publications/2011/9789241548090_eng.pdf (Accessed 21 October 2011).
- 22. Reed LJ, Muench H. A simple method for estimating fifty percent endpoints. Am J Hyg 1938; 27:493–497. [Google Scholar]
- 23. Goji NA, Nolan C, Hill H et al. Immune responses of healthy subjects to a single dose of intramuscular inactivated influenza A/Vietnam/1203/2004 (H5N1) vaccine after priming with an antigenic variant. J Infect Dis 2008; 198:635–641. [DOI] [PubMed] [Google Scholar]
- 24. Stephenson I, Bugarini R, Nicholson KG et al. Cross‐reactivity to highly pathogenic avian influenza H5N1 viruses after vaccination with nonadjuvanted and MF59‐adjuvanted influenza A/Duck/Singapore/97 (H5N3) vaccine: a potential priming strategy. J Infect Dis 2005; 191:1210–1215. [DOI] [PubMed] [Google Scholar]
- 25. Leroux‐Roels I, Roman F, Forgus S et al. Priming with AS03 A‐adjuvanted H5N1 influenza vaccine improves the kinetics, magnitude and durability of the immune response after a heterologous booster vaccination: an open non‐randomised extension of a double‐blind randomised primary study. Vaccine 2010; 28:849–857. [DOI] [PubMed] [Google Scholar]
- 26. Lin JT, Li CG, Wang X et al. Antibody persistence after 2‐dose priming and booster response to a third dose of an inactivated, adjuvanted, whole‐virion H5N1 vaccine. J Infect Dis 2009; 199:184–187. [DOI] [PubMed] [Google Scholar]
- 27. Nguyen HH, van Ginkel FW, Vu HL, McGhee JR, Mestecky J. Heterosubtypic immunity to influenza A virus infection requires B cells but not CD8+ cytotoxic T lymphocytes. J Infect Dis 2001; 183:368–376. [DOI] [PubMed] [Google Scholar]
- 28. Crowe BA, Brühl P, Gerencer M et al. Evaluation of the cellular immune responses induced by a non‐adjuvanted inactivated whole virus A/H5N1/VN/1203 pandemic influenza vaccine in humans. Vaccine 2010; 29:166–173. [DOI] [PubMed] [Google Scholar]
- 29. Roti M, Yang J, Berger D, Huston L, James EA, Kwok WW. Healthy human subjects have CD4+ T cells directed against H5N1 influenza virus. J Immunol 2008; 180:1758–1768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Lee LY, Ha do LA, Simmons C et al. Memory T cells established by seasonal human influenza A infection cross‐react with avian influenza A (H5N1) in healthy individuals. J Clin Invest 2008; 118:3478–3490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Goy K, Von Bibra S, Lewis J et al. Heterosubtypic T‐cell responses against avian influenza H5 haemagglutinin are frequently detected in individuals vaccinated against or previously infected with human subtypes of influenza. Influenza Other Respi Viruses 2008; 2:115–125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Dormitzer PR, Galli G, Castellino F et al. Influenza vaccine immunology. Immunol Rev 2011; 239:167–177. [DOI] [PubMed] [Google Scholar]
- 33. LaMere MW, Lam HT, Moquin A et al. Contributions of antinucleoprotein IgG to heterosubtypic immunity against influenza virus. J Immunol 2011; 186:4331–4339. [DOI] [PMC free article] [PubMed] [Google Scholar]