

Abstract
Despite recent advances in the understanding of the biology of renal cell carcinoma (RCC) and the implementation of novel targeted therapies, the overall 5 years’ survival rate for RCC patients remains disappointing. Late presentation, tumor heterogeneity and in particular the lack of molecular biomarkers for early detection and classification represent major obstacles. Global, untargeted comparative analysis of RCC vs tumor adjacent renal epithelium (NN) samples by high throughput analyses both at the transcriptome and proteome level have identified signatures, which might further clarify the molecular differences of RCC subtypes and might allow the identification of suitable therapeutic targets and diagnostic/prognostic biomarkers, but none thereof has yet been implemented in routine clinical use. The increasing knowledge regarding the functional role of noncoding microRNA (miR) in physiological, developmental, and pathophysiological processes by shaping the protein expression profile might provide an important link to improve the definition of disease-relevant regulatory networks. Taking into account that miR profiling of RCC and NN provides robust signatures discriminating between malignant and normal tissues, the concept of evaluating and scoring miR/protein pairs might represent a strategy for the selection and prioritization of potential biomarkers and their translation into practical use.
Keywords: RCC, miR, proteomics, biomarker
Introduction
Over the last 20 years, the incidence of renal cell carcinoma (RCC) has increased in the Western world, thereby currently accounting for approximately 3% of cancers and 2% of all cancer-related deaths.1 RCC represents a highly aggressive tumor entity. At the time of diagnosis approximately 30% of the patients have developed metastases.2 Clear cell RCC (ccRCC) accounting for almost 70% of all kidney cancers is the most common histological subtype of this disease and is characterized by the loss of the von Hippel Lindau (VHL) tumor suppressor gene expression either due to deletions, mutations, loss of heterozygosity and/or hypermethylation.3 In addition, ccRCC exhibits a wide range of natural histories and varying response rates to targeted therapies.4 In particular, early stage tumors have a significantly better disease-free survival after resection than tumors of higher stage and grade.5–7 In addition, to structural abnormalities or epigenetic silencing of VHL other molecular factors directly associated with the initiation and progression of this disease are largely unknown. Therefore, the molecular basis for the diversity regarding histological grade, clinical behavior and response to targeted therapy has to be better characterized to gain further insights into the heterogeneity of RCC. The identification of deregulated molecules and their interactions is required to elucidate the pathophysiology of RCC and to improve the development of new treatment strategies.
Despite major efforts within the past decade to establish and implement genomic and proteomic profiling strategies into clinical routine so far only the MammaPrint has been approved by the Food and Drug Administration (FDA, FDA Clears Breast Cancer Specific Molecular Prognostic Test, 2007).8 Although (c)DNA microarrays represent the most advanced and robust high throughput technology, the assessment of expression profilings still face a number of challenges such as laboratory to laboratory and platform to platform reproducibility.9 Moreover, most expression patterns were established with rather small sizes of patients’ cohorts and/or numbers of samples analyzed. Nevertheless, gene expression profilings have been performed with samples from RCC patient-matched specimens and cell lines using different cDNA microarray platforms, which resulted in the development of preliminary expression signatures and array-comparative genomic hybridization patterns, but only in rare cases the potential targets were validated.10,11 Thus, in the clinical setting tumor stage and grade are hitherto still used as predictors for the outcome of RCC patients.
Since mRNA expression patterns can not account for frequently occurring posttranslational modifications at the protein level, which are often associated with key regulatory or functional consequences profilings restricted to the transcriptomic level allow only limited prediction of the disease. Therefore, side by side evaluations of the genomic/transcriptomic and proteomic status are critical to enhance the knowledge of disease pathogenesis and the underlying molecular mechanisms of the differential mRNA and protein expression levels. An important link within the regulatory network between the available genomic information and its outcome at the proteomic level is represented by a class of small noncoding RNAs, termed miRs. miRs have been shown to play a fundamental role in different physiological and pathophysiologcal processes including cell proliferation, cell motility, differentiation, apoptosis and carcinogenesis. At the functional level they regulate from few to up to thousands of genes in regard to their individual mRNA transcript and/or protein expression levels. With the development of miR arrays, systematic expression profilings could be performed, which may contribute (i) to define the effects of miRs on the regulation of mRNA transcripts and proteins under defined conditions and (ii) to elucidate their biological role by gaining further insights into relevant regulatory networks.12
Important Features of miRs
miRs are a class of small noncoding RNAs of approximately 22 nucleotides in length frequently conserved across species. They bind to complementary sequences in the 3′ untranslated region (UTR) of their target genes and thus regulate mRNA transcription levels by cleavage of the target mRNA or by repression of protein synthesis in a tissue-specific manner.13,14 However, it is noteworthy that translation of some proteins can also be up-regulated by miRs under some circumstances.15–17 This might be explained by oscillation of miRs between repression and activation in coordination with the cell cycle or by transcription of miRs as a polycistron, which preferentially coregulates proteins in close proximity.16
It is estimated that approximately 30% of the human genes are regulated by miRs.18 However, miRs not only serve as direct regulators of gene expression but are likely regulated themselves by multiple factors. Most miRs are located in genomic regions distant from the annotated target genes and also encoded in distinct transcription units.19 Approximately 25% of miR were processed from introns, whereas others are clustered in the genome and seem to exhibit a functional relationship. Although more than 900 miRs that have been identified so far in mammals, their biological relevance and functional targets still remain largely unknown. However, there is evidence that miRs have been shown to be involved in modulating key cellular processes which might also be influenced by oncogenes as well as tumor suppressor genes.20 In addition, miRs have also been investigated in several solid and hematologic malignancies revealing deregulations in many tumors, including renal cell carcinoma.21–26 This functional correlation is further strengthened by the finding that miR encoding genes are frequently located at fragile sites or chromosomal regions undergoing point mutations, amplifications, deletions or translocations in tumors.27
Analytical Issues Linked to the Profiling of miRs
High Throughput Analysis for miR Identification
Despite the existence of several high throughput approaches to identify and/or quantify miRs in tissue samples there exists no gold standard. cDNA oligonucleotide micro- and polymerase chain reaction (PCR)-based arrays are currently applied as global scale techniques for miR profiling.28 Recently, bead-based flow cytometric miR expression profiling has been developed as a new emerging technology, which uses xMAP beads with locked nucleic acid capture probes to detect target specific miRs.29 Furthermore, cloning of so far known miRs allows the identification of the all expressed miRs. However, these technologies have limitations regarding their specificity and/or ability to detect novel so far unknown RNAs. With the establishment of the next generation small RNA (smRNA) deep sequencing30,31 all smRNAs present in the samples analyzed including novel and under-expressed miRs, small nucleolar RNAs (snoRNAs), small cytoplasmic RNAs and small nuclear RNAs can be detected.32–34 Profiling of miRs by miR arrays and deep sequencing is nowadays also applicable for formalin-fixed paraffin-embedded tissues,35 allowing the retrospective analysis of large cohorts of clinical samples including material from clinical trial-based studies. This technology provides an extremely powerful tool in the identification of clinical relevant markers as well as therapeutic targets in cancer research.36 Although different strategies have been applied to identify specific miR profiles and functions, the question, how many of these changes are critical for the function of an individual miR, still remains elusive.
miRNA Target Prediction
So far, computational algorithms have been the major driving force in predicting miR targets, which are primarily based on sequence complementarities between the 5′ end of most miRs and the 3′ UTR of target genes and free energy predictions. Due to the ubiquitous nature of miR/mRNA targeting, independent prediction databases using 11 different algorithms have been developed. The miRWalk database (http://www.ma.uni-heidelberg.de/apps/zmf/mirwalk/), is covering information based on eight established miR target prediction algorithms, such as e. g. RNA22,37 miRanda,38 miRDB,39 TargetScan,40 RNAhybrid,41 PITA,42 PICTAR43 and Diana-microT.44
miRs can target from few to more than thousand mRNAs causing significant expression changes of the targeted transcripts and/or proteins.40,45 However, in silico approaches might not necessarily be reliable predictors for miR target interactions due to the high rates of false positive and false negative miR targets.
miR Experimental Target Validation
The lack of validation data for predicted miR target genes currently represents the bottleneck in the functional characterization of miR-dependent regulatory networks. However, this information is essential to gain further insights into the molecular mechanisms of miR-controlled pathogenic processes such as tumorigenesis, invasiveness of tumors and metastasis formation.46 The regulatory role of miRs makes them per se strong candidate oncogenes and tumor suppressor genes.47,48 Examples of both oncogenic miRs and those that suppress tumors have been reported.49–51 In addition, mutations and single nucleotide polymorphisms (SNPs) within the seed sequence of miRs or alterations in their epigenetic control might affect miR processing leading to reduced miR expression.52–55
Expression and Function of miRs in RCC
There exists a growing body of literature focusing on the analysis and the role of miRs in RCC, in particular in clear cell RCC. Studies investigating global changes in the miR pattern of RCC lesions compared to normal kidney epithelium focusing on the effects of individual miR on the given RCC subtype.21–23,25,54,56 Hierarchical cluster analysis of established miR expression profiles allowed the classification of matched tissue pairs into RCC lesions and nontumoral kidney parenchyma and in some cases even according to histological distinct RCC subtypes as well as tumor staging and grading.21–23,25,35,57,58 Thus, there exist unique miR signatures for RCC lesions, each renal tumor subtype, autologous kidney parenchyma, primary and metastatic lesions.25 Furthermore, epigenetic changes might also modulate the miR expression and function as demonstrated for the miR-9 family exhibiting an altered methylation status in primary RCC lesions.54 Moreover, genetic polymorphisms of the miR processing machinery might contribute to the risk of RCC as demonstrated by Horikawa and co-workers (2008).55 These data suggest that miRs are involved in the initiation as well as progression of RCC and might serve as diagnostic, prognostic and therapeutic tools. Until now 184 miRs have been identified to be differentially expressed between RCC and NN.26,35,56–60 The numbers of tissue samples analyzed ranged from 3 to >72, but not for all of these studies pairs of RCC and normal corresponding kidney epithelium were available. A similar miR regulation pattern was also defined in various other tumor types, such as ovarian, breast, pancreas, lung and stomach cancer. In order to determine differentially expressed miRs the independently identified miRs were subjected to strict selection criteria such as the availability of at least 3 independent identifications along with a consistent regulation pattern leading to a subset of 12 down-regulated and 8 up-regulated miRs (Table 1). The group of down-regulated miRs in RCC lesions comprised the members miR-30a, –133b, –138, –141, –200a, –200b, –200c, –204, –429, –510, and –514 (Table 1), whereas the panel of up-regulated miRs is represented by miR 34a, –34b, 106a, 106b, –155, –185, and –224 (Table 1). Although predominantly classified as up-regulated (Table 1)56 miR-21 is also included in a set of down-regulated miRs.
Table 1.
Comparison of Differentially Regulated miRs with the Counter Regulated Proteins Identified
| miRs
|
target proteins
|
||||
|---|---|---|---|---|---|
| name | reference | DNA locus | RCC related region | name | reference |
| Down-regulated miRNA | |||||
| miR-21a | 56 | 17q23 | 17q21 loss of het.101 | Retinal dehydrogenase 1 | 78, 79, 82, 84 |
| Phosphoglucomutase-1 | 69 | ||||
| miR-30a | 26, 58, 59 | 6q13 | loss 6q66,103 | Dihydropyrimidinase-related protein 2 | 77, 84 |
| miR-133b | 58, 59 | 6p12 | loss 6p65 | Ferritin light chain | 86 |
| N-myc downstream-regulated gene 1 protein | 84 | ||||
| miR-138 | 35, 59 | 3p21 | loss of 3p,65,103 loss of 3p21-pter66 |
3′(2′),5′-bisphosphate nucleotidase 1 | 83 |
| miR-141 | 26, 35, 56, 58–60 | 12p13 | Glutaminase kidney isoform | 77 | |
| Nucleoside diphosphate kinase A | 86 | ||||
| miR-200a | 56, 58, 59 | 1p36 | loss 1p102 | Aspartyl-tRNA synthetase | 84 |
| Nucleoside diphosphate kinase A | 86 | ||||
| miR-200b | 26, 57–60 | 1p36 | loss 1p102 | Chloride intracellular channel protein 4 | 69 |
| Fibronectin | 82 | ||||
| Stress-70 protein | 79 | ||||
| Moesin | 77 | ||||
| miR-200c | 26, 35, 56–60 | 12p13 | Chloride intracellular channel protein 4 | 69 | |
| Stress-70 protein | 79 | ||||
| Moesin | 77 | ||||
| miR-204 | 26, 56, 58, 59 | 9q21 | loss 966 | Ezrin | 82 |
| Lamin-A | 82 | ||||
| miR-429 | 26, 56, 58, 59 | 1p36 | loss 1p102 | Chloride intracellular channel protein 4 | 69 |
| Four and a half LIM domains protein 1 | 82 | ||||
| Fibronectin | 82, 84 | ||||
| Moesin | 77 | ||||
| miR-510 | 26, 58, 59 | Xq27 | loss Xq65,66,102 | Peptidyl-prolyl cis-trans isomerase A | 78 |
| miR-514 | 26, 35, 58, 59 | Xq27 | loss Xq65,66,102 | UDP-glucose 6-dehydrogenase | 83 |
| Up-regulated miRNA | |||||
| miR-21 | 26, 57–59 | 17q23 | gain 17q65 | ||
| miR-34a | 56, 58, 59 | 1p36 | add(1)(p34)104 | Calbindin | 81, 85 |
| miR-34b | 56, 58, 59 | 11q23 | Calbindin | 81, 85 | |
| miR-106a | 57, 59, 60 | Xq26 | gain Xq65 | Guanosine-diphosphatase | 85 |
| Sorting nexin-9 | 69 | ||||
| miR-106b | 57–60 | 7q22 | trisomy 7,103 gain 765,66 | 6-phosphofructokinase type C | 78 |
| Sorting nexin-9 | 69 | ||||
| miR-155 | 26, 35, 58–60 | 21q21 | del(21)(q22)104 | 4-aminobutyrate aminotransferase | 75, 82, 84 |
| Glycine amidinotransferase | 81, 82, 84, 85 | ||||
| miR-185 | 26, 58, 59 | 22q11 | ATP synthase subunit beta | 83, 84 | |
| Guanosine-diphosphatase | 85 | ||||
| 14-3-3 protein gamma | 85 | ||||
| miR-224 | 26, 35, 58, 59 | Xq28 | gain Xq65 | Argininosuccinate synthase | 75, 81, 84 |
| Fibrinogen beta chain | 82 | ||||
miR-21: 6 times detected, only one time a down-regulation; het.: heterozygosity.
miR-141 (6 identifications) and miR-200c (7 identifications) were not only the most frequently down-regulated miRs when compared to normal renal epithelium but also mapped to the same locus. Moreover, the down-regulation of both miRs seems to be involved in the induction of epithelial-to-mesenchymal transition (EMT) by up-regulating the expression of zinc finger E box-binding homeobox 2 (ZFHXIB, ZEB2), a transcriptional repressor of CDH1/E-cadherin.61 This is associated with an increased aggressiveness of the tumor, which is in line with the report of Park and coauthors demonstrating that the miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2.62
The most frequently up-regulated miRs in RCC were miR-155 and miR-224, which are mapped to distinct chromosomal loci. Some of the up-regulated miRs (miR-21, miR-34) have so far been associated with cancer-related processes such as proliferation, invasion or metastases formation,63,64 others like miR-106b have been linked to disease progression, in particular with prediction of metastatic recurrence after nephrectomy and to poor patients survival.60
Matching patterns between chromosomal alterations in RCC lesions and miR deregulation was found for 84% of the selected miRs (refs 26, 35, 56–60 and Table 1). The panel of selected miRs is distributed over 11 chromosomes. Four of the differentially expressed miRs were located on chromosome 1 (miR-200a, - 200b, -429 and -34a) or X (miR-510, -514, -106b and -224), whereas miR-30a and -133b were mapped to chromosome 6 and the EMT-associated miR-141 and -200c to chromosome 12, respectively. The remaining miRs were mapped to chromosome 3 (miR-138), 7 (miR-106b), 9 (miR-204), 11 (miR-34b), 17 (miR-21), 21 (miR-155) and 22 (miR-185). Their regulation pattern was mostly in line with genetic alterations previously defined in RCC by comparative genomic hybridization65,66 with the exception of chromosome 12, for which rather gains in a frequency of 20% for both 12p and 12q was described and of chromosome 21 for which no frequent genetic alterations yet exist in RCC. In terms of the miRs mapped to chromosome X a 10% frequency of Xp loss as well as Xp and/or Xq gain was reported.65
Concerning validation and functional studies, the distinct miR expression pattern detected in RCC vs NN was only confirmed in a limited number of samples analyzed by quantitative real-time PCR,56,59,67 even less information is available in terms of the functional characterization of the differentially expressed miRs.
Proteomic Profiling of RCC
Protein profiling is a complementary approach to the mRNA-based analysis, which has several advantages despite its limited high throughput capacity.68 Studies at the proteome level not only allow to bridge potential gaps in respect to correlations between mRNA and protein expression levels, but also allow the detection and subsequent identification of posttranslational modifications (PTM), such as e. g. glycosylation and phosphorylation in combination with mass spectrometric analyses. This is of great interest because in most cases PTMs have an enormous impact on the protein function and/or its biological activity.
Various proteome-based strategies have been employed for the proteomic profiling of RCC, mostly in the context of identification of candidate biomarkers in RCC,69–86 which despite certain advantages and limitations complement for the most part each other.68,87 Next to gel-based technologies using conventional two-dimensional polyacrylamide gel electrophoresis (2-DE), a number of gel-free approaches, in particular liquid chromatography coupled with matrix-assisted laser desortion ionization or electro-spray ionization MS, have been employed to determine changes in the protein expression profiles between RCC and normal renal tissues and/or respective primary or established RCC cell cultures. The latter approach complements 2D-PAGE-based profilings by covering not only low abundant or membrane proteins but also proteins excluded from gel-based approaches due to limitations such as size or extreme isoelectric points. In analogy to miR studies, the number of RCC lesions and corresponding epithelium analyzed significantly varied from 5 to >50 paired samples. Although the comparative analysis of RCC lesions vs patient-matched NN or representative RCC cell lines generated a pool of 336 differentially expressed proteins, only a limited number of proteins have been validated by RT-PCR, Western blot and/ or immunohistochemistry and demonstrated a general RCC or RCC subtype-specific expression.68,86–88 Furthermore, the functional relevance of candidate biomarkers has only been addressed for a limited number of candidate targets.87
Can the Correlation of miR and Proteomic Profiling Data Lead to a Further Improvement in the Search for Biomarkers?
Using distinct high throughput platforms, a number of studies have identified panels of transcripts, miRs, and proteins that are differentially expressed between normal renal tissue and RCC lesions or between lesions of distinct RCC subtypes.26,68,77,81,89–93 The simultaneous profiling of the miR and mRNA expression pattern was recently performed and demonstrated an inverse relationship between the miR expression profile and that of a number of putative targets.94–96 The linkage between miR and mRNA expression levels led to the identification of deregulated miR/mRNA pairs involved in the initiation or maintenance of disease processes.26,97 However, this approach has its limitations since it cannot identify targets that are regulated at the posttranscriptional level. For such targets the changes at the mRNA levels remain unaffected, whereas the corresponding protein levels will be frequently reduced.98
In contrast, it could be postulated that proteome-based approaches can detect and quantify changes at the protein expression level induced by specific miRs. Since there exists increasing evidence that translational repression instead of mRNA degradation is the dominant miR regulatory mechanism the analysis of the correlation between miR and protein expression levels is gaining more importance.97,99 It is suggested that broad changes in protein synthesis are induced by miRs.45,100 Therefore, the concept to perform parallel miR and proteomic profiling represents a novel experimental strategy to define the protein output45,100 and might be further applied to define potential biomarkers in any disease, including RCC. On the basis of this assumption, the determination of the inverse correlation between differentially expressed miRs and proteins in tumor lesions and corresponding normal tissues might lead to the optimization of the selection process for candidate biomarkers.
Despite the majority of miRs exhibit an impact on the general protein expression profile,45,100 a comparative global proteomic and miR profiling in malignancies has not yet been performed. It is postulated that the comparison of proteome-based data to available miR profiling data sets will demonstrate a significant inverse regulation between differentially expressed miRs and their potential protein targets (Table 1). However, the disadvantage of this selection strategy is the elimination of indirect regulations since only those miR/protein pairs with strictly inverse correlated expression levels will be retained. This approach will reduce the number of putative, pair-matched-based protein targets extracted from databases and might provide highly robust subsets of relevant functional targets. However, the complex biology and tissue specificity of miRs make it still difficult to elucidate their precise role in the initiation and progression of disease and their link to deregulated genes and/or protein expression.
Identification of Correlative Signatures between miR and Protein Pairs in RCC and their Association to Pathways
To determine a link between miR and protein expression the subset of frequently and consistently deregulated miRs was subjected to a miRWalk search to collect their predicted targets. Only miR targets (Table 1), which were found in at least four of the eight prediction algorithms, were subsequently stored in an in-house database. These were then aligned to the experimentally defined differentially expressed proteins in RCC samples. With the exception of the up-regulated miR-21, counter-regulated miR/protein pairs could be defined (Table 1) for the subset of miRs fulfilling the selection criteria. In addition, the chromosomal location of the miRs are in line with chromosomal alterations in RCC.65,66,101–104
In the next step, the identified pairs of counter-regulated miRs and proteins were dedicated to their respective pathway according to the KEGG database (Table 2 and Figure 1). Interestingly several metabolic pathways known to be involved in cancer seemed to be altered by different miRs such as the nucleotide, energy as well as amino acid metabolism. Regarding the purine metabolism, the guanine deaminase protein was down-regulated in RCC,85 which was accompanied by an up-regulation of miRs-106a and –185. Since this enzyme is responsible for the guanine degradation the diminished expression might be a prerequisite for tumor cells to reach an increased proliferation rate as large amounts of DNA are required for cell division. In contrast, the up-regulation of the nucleoside diphosphate kinase might lead to an increased RCC proliferation since the required nucleoside triphosphates required for DNA synthesis can be provided. Concerning the energy metabolism miR-21 down-regulation influencing the expression of phosphoglucomutase was detected.56,69 This enzyme is up-regulated at the proteome level, suggesting that miR-21 has potential regulatory effects for the glycolytic flux and might therefore contribute to the metabolic transformed phenotype. In this context it is noteworthy that miR-106b targeting phosphofructokinase (PFKP), the rate-determining enzyme of glycolysis, is up-regulated in RCC,59 but there also exist controversial results regarding the expression of this enzyme82,84 in RCC lesions.79 Furthermore the observed up-regulation of glutaminase in RCC is accompagnied by a decreased expression of miR-141, which is in line with the enhanced glutaminolysis frequently observed during metabolic transformation of tumor cells.26,35,56,58–60,77 The decreased expression of miR-200b, –200c, –429 as well as miR-204, which targets moesin and ezrin, respectively, might be the underlying molecular mechanism for the increased cell migration of RCC caused by an up-regulation of the moesin/ezrin/radixin family frequently detected in proteome studies of RCC. Taken together miRs are potential key regulators for the expression of different metabolic enzymes that are required for the altered metabolic demands of RCC.
Table 2.
Differentially Regulated miR/Protein Pairs Identified in RCC Associated to Their Metabolic Pathwaysa
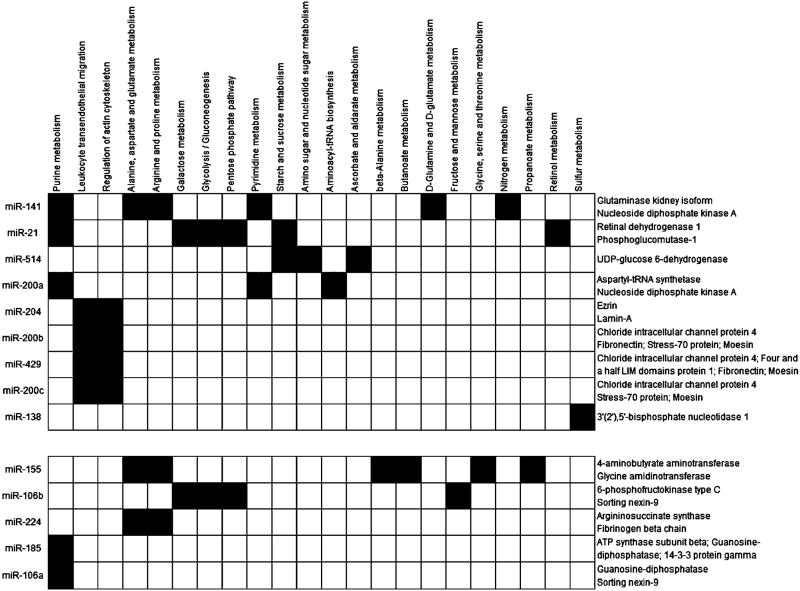
|
Upper part, down- regulated miRs; lower part, up-regulated miRs.
Figure 1.

Overview of influenced metabolic pathways. The scheme assigns the inverse correlated miRs and proteins to their respective metabolic pathways according to the KEGG database. The gray boxes represent metabolic parts. ↑, up-regulated; ↓, down-regulated.
Conclusions
Until now, high throughput RNA, DNA, and protein analyses of tumors have not given comprehensive information about the initiation and progression of malignancies. In addition, there exist only a limited number of reports analyzing the miR expression pattern in RCC lesions.26,35,56–60,69–86 The link of the miR profile to respective protein expression patterns of RCC lesions and normal kidney parenchyma might lead to an increased understanding of the molecular mechanisms involved in the pathogenesis of this disease.
In the future, the correlative expression profile of miRs and proteins have to be extended to a larger sample number to use the coordinate by regulated miR and protein pairs as diagnostic and prognostic tools not only for RCC but also for other cancers. Nevertheless, the number of tumor samples analyzed so far is too low to draw general conclusions or to refer to a common RCC-specific miR/protein profile. In addition, there exists only limited information about (i) the clinical significance of miRs by correlating miR expression levels with the outcome of RCC patients and (ii) about the relevance of miR in the alteration of fundamental processes in RCC.
Acknowledgments
This work was funded by the Bun-desministerium für Bildung und Forschung, NGFN 031U101H, the DFG GUP GRK 1591, and the Else Kroener Fresenius Stiftung. We thank Sylvi Magdeburg and Nicole Ott for excellent secretarial assistance.
Abbreviations
- ccRCC
clear cell RCC
- HIF1-α
hypoxia inducible factor
- miR
microRNA
- NN
tumor adjacent renal epithelium
- PFKP
phosphofructokinase
- RCC
renal cell carcinoma
- UTR
untranslated region
- VHL
von Hippel Lindau
References
- 1.Ljungberg B, Hanbury DC, Kuczyk MA, Merseburger AS, Mulders PF, Patard JJ, Sinescu IC. Renal cell carcinoma guideline. Eur Urol. 2007;51(6):1502–10. doi: 10.1016/j.eururo.2007.03.035. [DOI] [PubMed] [Google Scholar]
- 2.Campbell D, Walker R, Mathew T, Craig J. Commentary on “Influence of industry on renal guideline development. Clin J Am Soc Nephrol. 2007;2(2):211. doi: 10.2215/CJN.00300107. [DOI] [PubMed] [Google Scholar]
- 3.Yoshimoto T, Matsuura K, Karnan S, Tagawa H, Nakada C, Tanigawa M, Tsukamoto Y, Uchida T, Kashima K, Akizuki S, Takeuchi I, Sato F, Mimata H, Seto M, Moriyama M. High-resolution analysis of DNA copy number alterations and gene expression in renal clear cell carcinoma. J Pathol. 2007;213(4):392–401. doi: 10.1002/path.2239. [DOI] [PubMed] [Google Scholar]
- 4.Lam JS, Leppert JT, Belldegrun AS, Figlin RA. Novel approaches in the therapy of metastatic renal cell carcinoma. World J Urol. 2005;23(3):202–12. doi: 10.1007/s00345-004-0466-0. [DOI] [PubMed] [Google Scholar]
- 5.Fuhrman SA, Lasky LC, Limas C. Prognostic significance of morphologic parameters in renal cell carcinoma. Am J Surg Pathol. 1982;6(7):655–63. doi: 10.1097/00000478-198210000-00007. [DOI] [PubMed] [Google Scholar]
- 6.Janzen NK, Kim HL, Figlin RA, Belldegrun AS. Surveillance after radical or partial nephrectomy for localized renal cell carcinoma and management of recurrent disease. Urol Clin North Am. 2003;30(4):843–52. doi: 10.1016/s0094-0143(03)00056-9. [DOI] [PubMed] [Google Scholar]
- 7.Ficarra V, Guille F, Schips L, de la Taille A, Prayer Galetti T, Tostain J, Cindolo L, Novara G, Zigeuner R, Bratti E, Li G, Altieri V, Abbou CC, Zanolla L, Artibani W, Patard JJ. Proposal for revision of the TNM classification system for renal cell carcinoma. Cancer. 2005;104(10):2116–23. doi: 10.1002/cncr.21465. [DOI] [PubMed] [Google Scholar]
- 8.FDA. [last accessed 12/ 15/10.];FDA Clears Breast Cancer Specific Molecular Prognostic Test. 2007 http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/2007/ucm108836.htm.
- 9.Fan X, Shi L, Fang H, Cheng Y, Perkins R, Tong W. DNA microarrays are predictive of cancer prognosis: a re-evaluation. Clin Cancer Res. 2010;16(2):629–36. doi: 10.1158/1078-0432.CCR-09-1815. [DOI] [PubMed] [Google Scholar]
- 10.Osunkoya AO, Yin-Goen Q, Phan JH, Moffitt RA, Stokes TH, Wang MD, Young AN. Diagnostic biomarkers for renal cell carcinoma: selection using novel bioinformatics systems for microarray data analysis. Hum Pathol. 2009;40(12):1671–8. doi: 10.1016/j.humpath.2009.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ohno Y, Izumi M, Tachibana M, Kawamura T, Yoshioka K, Aoyagi T, Ohori M, Namiki K, Sakamoto N, Nakagami Y, Hatano T, Akimoto S, Nishimura T. Characterization and gene expression analysis of novel matched primary and metastatic renal cell carcinoma cell lines. Oncol Rep. 2008;20(3):501–9. [PubMed] [Google Scholar]
- 12.Liu CG, Calin GA, Meloon B, Gamliel N, Sevignani C, Ferracin M, Dumitru CD, Shimizu M, Zupo S, Dono M, Alder H, Bullrich F, Negrini M, Croce CM. An oligonucleotide microchip for genome-wide microRNA profiling in human and mouse tissues. Proc Natl Acad Sci US A. 2004;101(26):9740–4. doi: 10.1073/pnas.0403293101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Engels BM, Hutvagner G. Principles and effects of microRNA-mediated post-transcriptional gene regulation. Oncogene. 2006;25(46):6163–9. doi: 10.1038/sj.onc.1209909. [DOI] [PubMed] [Google Scholar]
- 14.Zeng Y, Cullen BR. Recognition and cleavage of primary microRNA transcripts. Methods Mol Biol. 2006;342:49–56. doi: 10.1385/1-59745-123-1:49. [DOI] [PubMed] [Google Scholar]
- 15.Cheng J, Zhou L, Xie QF, Xie HY, Wei XY, Gao F, Xing CY, Xu X, Li LJ, Zheng SS. The impact of miR-34a on protein output in hepatocellular carcinoma HepG2 cells. Proteomics. 2010;10(8):1557–72. doi: 10.1002/pmic.200900646. [DOI] [PubMed] [Google Scholar]
- 16.Vasudevan S, Tong Y, Steitz JA. Switching from repression to activation: microRNAs can up-regulate translation. Science. 2007;318(5858):1931–4. doi: 10.1126/science.1149460. [DOI] [PubMed] [Google Scholar]
- 17.Yang Y, Chaerkady R, Beer MA, Mendell JT, Pandey A. Identification of miR-21 targets in breast cancer cells using a quantitative proteomic approach. Proteomics. 2009;9(5):1374–84. doi: 10.1002/pmic.200800551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Esquela-Kerscher A, Slack FJ. Oncomirs - microRNAs with a role in cancer. Nat Rev Cancer. 2006;6(4):259–69. doi: 10.1038/nrc1840. [DOI] [PubMed] [Google Scholar]
- 19.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–97. doi: 10.1016/s0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 20.Ambros V. The functions of animal microRNAs. Nature. 2004;431(7006):350–5. doi: 10.1038/nature02871. [DOI] [PubMed] [Google Scholar]
- 21.Gottardo F, Liu CG, Ferracin M, Calin GA, Fassan M, Bassi P, Sevignani C, Byrne D, Negrini M, Pagano F, Gomella LG, Croce CM, Baffa R. Micro-RNA profiling in kidney and bladder cancers. Urol Oncol. 2007;25(5):387–92. doi: 10.1016/j.urolonc.2007.01.019. [DOI] [PubMed] [Google Scholar]
- 22.Huang Y, Dai Y, Yang J, Chen T, Yin Y, Tang M, Hu C, Zhang L. Microarray analysis of microRNA expression in renal clear cell carcinoma. Eur J Surg Oncol. 2009;35(10):1119–23. doi: 10.1016/j.ejso.2009.04.010. [DOI] [PubMed] [Google Scholar]
- 23.Jung M, Mollenkopf HJ, Grimm C, Wagner I, Albrecht M, Waller T, Pilarsky C, Johannsen M, Stephan C, Lehrach H, Nietfeld W, Rudel T, Jung K, Kristiansen G. MicroRNA profiling of clear cell renal cell cancer identifies a robust signature to define renal malignancy. J Cell Mol Med. 2009;13(9B):3918–28. doi: 10.1111/j.1582-4934.2009.00705.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA, Downing JR, Jacks T, Horvitz HR, Golub TR. MicroRNA expression profiles classify human cancers. Nature. 2005;435(7043):834–8. doi: 10.1038/nature03702. [DOI] [PubMed] [Google Scholar]
- 25.Petillo D, Kort EJ, Anema J, Furge KA, Yang XJ, Teh BT. MicroRNA profiling of human kidney cancer subtypes. Int J Oncol. 2009;35(1):109–14. doi: 10.3892/ijo_00000318. [DOI] [PubMed] [Google Scholar]
- 26.Liu H, Brannon AR, Reddy AR, Alexe G, Seiler MW, Arreola A, Oza JH, Yao M, Juan D, Liou LS, Ganesan S, Levine AJ, Rathmell W, Bhanot GV. Identifying mRNA targets of microRNA dysregulated in cancer: with application to clear cell Renal Cell Carcinoma. BMC Syst Biol. 2010;4:51. doi: 10.1186/1752-0509-4-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M, Croce CM. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci US A. 2004;101(9):2999–3004. doi: 10.1073/pnas.0307323101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jiang J, Lee EJ, Gusev Y, Schmittgen TD. Real-time expression profiling of microRNA precursors in human cancer cell lines. Nucleic Acids Res. 2005;33(17):5394–403. doi: 10.1093/nar/gki863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Biscontin A, Casara S, Cagnin S, Tombolan L, Rosolen A, Lanfranchi G, De Pitta C. New miRNA labeling method for bead-based quantification. BMC Mol Biol. 2010;11:44. doi: 10.1186/1471-2199-11-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Morin RD, Aksay G, Dolgosheina E, Ebhardt HA, Magrini V, Mardis ER, Sahinalp SC, Unrau PJ. Comparative analysis of the small RNA transcriptomes of Pinus contorta and Oryza sativa. Genome Res. 2008;18(4):571–84. doi: 10.1101/gr.6897308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rajagopalan R, Vaucheret H, Trejo J, Bartel DP. A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes Dev. 2006;20(24):3407–25. doi: 10.1101/gad.1476406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kasschau KD, Fahlgren N, Chapman EJ, Sullivan CM, Cumbie JS, Givan SA, Carrington JC. Genome-wide profiling and analysis of Arabidopsis siRNAs. PLoS Biol. 2007;5(3):e57. doi: 10.1371/journal.pbio.0050057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lu YC, Smielewska M, Palakodeti D, Lovci MT, Aigner S, Yeo GW, Graveley BR. Deep sequencing identifies new and regulated microRNAs in Schmidtea mediterranea. RNA. 2009;15(8):1483–91. doi: 10.1261/rna.1702009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Moxon S, Jing R, Szittya G, Schwach F, Rusholme Pilcher RL, Moulton V, Dalmay T. Deep sequencing of tomato short RNAs identifies microRNAs targeting genes involved in fruit ripening. Genome Res. 2008;18(10):1602–9. doi: 10.1101/gr.080127.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Weng L, Wu X, Gao H, Mu B, Li X, Wang JH, Guo C, Jin JM, Chen Z, Covarrubias M, Yuan YC, Weiss LM, Wu H. MicroRNA profiling of clear cell renal cell carcinoma by whole-genome small RNA deep sequencing of paired frozen and formalin-fixed, paraffin-embedded tissue specimens. J Pathol. 2010;222(1):41–51. doi: 10.1002/path.2736. [DOI] [PubMed] [Google Scholar]
- 36.Tam W. The emergent role of microRNAs in molecular diagnostics of cancer. J Mol Diagn. 2008;10(5):411–4. doi: 10.2353/jmoldx.2008.080067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Miranda KC, Huynh T, Tay Y, Ang YS, Tam WL, Thomson AM, Lim B, Rigoutsos I. A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell. 2006;126(6):1203–17. doi: 10.1016/j.cell.2006.07.031. [DOI] [PubMed] [Google Scholar]
- 38.John B, Enright AJ, Aravin A, Tuschl T, Sander C, Marks DS. Human MicroRNA targets. PLoS Biol. 2004;2(11):e363. doi: 10.1371/journal.pbio.0020363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wang X. miRDB: a microRNA target prediction and functional annotation database with a wiki interface. RNA. 2008;14(6):1012–7. doi: 10.1261/rna.965408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005;120(1):15–20. doi: 10.1016/j.cell.2004.12.035. [DOI] [PubMed] [Google Scholar]
- 41.Kruger J, Rehmsmeier M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006;34(Web Server issue):W451–4. doi: 10.1093/nar/gkl243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kertesz M, Iovino N, Unnerstall U, Gaul U, Segal E. The role of site accessibility in microRNA target recognition. Nat Genet. 2007;39(10):1278–84. doi: 10.1038/ng2135. [DOI] [PubMed] [Google Scholar]
- 43.Krek A, Grun D, Poy MN, Wolf R, Rosenberg L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M, Rajewsky N. Combinatorial microRNA target predictions. Nat Genet. 2005;37(5):495–500. doi: 10.1038/ng1536. [DOI] [PubMed] [Google Scholar]
- 44.Maragkakis M, Alexiou P, Papadopoulos GL, Reczko M, Dalamagas T, Giannopoulos G, Goumas G, Koukis E, Kourtis K, Simossis VA, Sethupathy P, Vergoulis T, Koziris N, Sellis T, Tsanakas P, Hatzigeorgiou AG. Accurate microRNA target prediction correlates with protein repression levels. BMC Bioinform. 2009;10:295. doi: 10.1186/1471-2105-10-295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Baek D, Villen J, Shin C, Camargo FD, Gygi SP, Bartel DP. The impact of microRNAs on protein output. Nature. 2008;455(7209):64–71. doi: 10.1038/nature07242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ma L, Weinberg RA. Micromanagers of malignancy: role of microRNAs in regulating metastasis. Trends Genet. 2008;24(9):448–56. doi: 10.1016/j.tig.2008.06.004. [DOI] [PubMed] [Google Scholar]
- 47.Dykxhoorn DM. MicroRNAs and Metastasis: Little RNAs Go a Long Way. Cancer Res. 2010;70(16):6401–6. doi: 10.1158/0008-5472.CAN-10-1346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Varol N, Konac E, Gurocak OS, Sozen S. The realm of microRNAs in cancers. Mol Biol Rep. 2010 doi: 10.1007/s11033-010-0205-0. [DOI] [PubMed] [Google Scholar]
- 49.He L, Thomson JM, Hemann MT, Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe SW, Hannon GJ, Hammond SM. A microRNA polycistron as a potential human oncogene. Nature. 2005;435(7043):828–33. doi: 10.1038/nature03552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hammond SM. MicroRNA therapeutics: a new niche for antisense nucleic acids. Trends Mol Med. 2006;12(3):99–101. doi: 10.1016/j.molmed.2006.01.004. [DOI] [PubMed] [Google Scholar]
- 51.Ortholan C, Puissegur MP, Ilie M, Barbry P, Mari B, Hofman P. MicroRNAs and lung cancer: new oncogenes and tumor suppressors, new prognostic factors and potential therapeutic targets. Curr Med Chem. 2009;16(9):1047–61. doi: 10.2174/092986709787581833. [DOI] [PubMed] [Google Scholar]
- 52.Iwai N, Naraba H. Polymorphisms in human pre-miRNAs. Biochem Biophys Res Commun. 2005;331(4):1439–44. doi: 10.1016/j.bbrc.2005.04.051. [DOI] [PubMed] [Google Scholar]
- 53.Kanwar JR, Mahidhara G, Kanwar RK. MicroRNA in human cancer and chronic inflammatory diseases. Front Biosci (Schol Ed) 2010;2:1113–26. doi: 10.2741/s121. [DOI] [PubMed] [Google Scholar]
- 54.Hildebrandt MA, Gu J, Lin J, Ye Y, Tan W, Tamboli P, Wood CG, Wu X. Hsa-miR-9 methylation status is associated with cancer development and metastatic recurrence in patients with clear cell renal cell carcinoma. Oncogene. 2010;29(42):5724–8. doi: 10.1038/onc.2010.305. [DOI] [PubMed] [Google Scholar]
- 55.Horikawa Y, Wood CG, Yang H, Zhao H, Ye Y, Gu J, Lin J, Habuchi T, Wu X. Single nucleotide polymorphisms of microRNA machinery genes modify the risk of renal cell carcinoma. Clin Cancer Res. 2008;14(23):7956–62. doi: 10.1158/1078-0432.CCR-08-1199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yi Z, Fu Y, Zhao S, Zhang X, Ma C. Differential expression of miRNA patterns in renal cell carcinoma and nontumorous tissues. J Cancer Res Clin Oncol. 2010;136(6):855–62. doi: 10.1007/s00432-009-0726-x. [DOI] [PubMed] [Google Scholar]
- 57.Chow TF, Youssef YM, Lianidou E, Romaschin AD, Honey RJ, Stewart R, Pace KT, Yousef GM. Differential expression profiling of microRNAs and their potential involvement in renal cell carcinoma pathogenesis. Clin Biochem. 2010;43(1–2):150–8. doi: 10.1016/j.clinbiochem.2009.07.020. [DOI] [PubMed] [Google Scholar]
- 58.Juan D, Alexe G, Antes T, Liu H, Madabhushi A, Delisi C, Ganesan S, Bhanot G, Liou LS. Identification of a microRNA panel for clear-cell kidney cancer. Urology. 2010;75(4):835–41. doi: 10.1016/j.urology.2009.10.033. [DOI] [PubMed] [Google Scholar]
- 59.White NM, Bui A, Mejia-Guerrero S, Chao J, Soosaipillai A, Youssef Y, Mankaruos M, Honey RJ, Stewart R, Pace KT, Sugar L, Diamandis EP, Dore J, Yousef GM. Dysregulation of kallikrein-related peptidases in renal cell carcinoma: potential targets of miRNAs. Biol Chem. 2010;391(4):411–23. doi: 10.1515/BC.2010.041. [DOI] [PubMed] [Google Scholar]
- 60.Slaby O, Jancovicova J, Lakomy R, Svoboda M, Poprach A, Fabian P, Kren L, Michalek J, Vyzula R. Expression of miRNA-106b in conventional renal cell carcinoma is a potential marker for prediction of early metastasis after nephrectomy. J Exp Clin Cancer Res. 2010;29:90. doi: 10.1186/1756-9966-29-90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Nakada C, Matsuura K, Tsukamoto Y, Tanigawa M, Yoshimoto T, Narimatsu T, Nguyen LT, Hijiya N, Uchida T, Sato F, Mimata H, Seto M, Moriyama M. Genome-wide microRNA expression profiling in renal cell carcinoma: significant down-regulation of miR-141 and miR-200c. J Pathol. 2008;216(4):418–27. doi: 10.1002/path.2437. [DOI] [PubMed] [Google Scholar]
- 62.Park SM, Gaur AB, Lengyel E, Peter ME. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Dev. 2008;22(7):894–907. doi: 10.1101/gad.1640608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Dutta KK, Zhong Y, Liu YT, Yamada T, Akatsuka S, Hu Q, Yoshihara M, Ohara H, Takehashi M, Shinohara T, Masutani H, Onuki J, Toyokuni S. Association of microRNA-34a overexpression with proliferation is cell type-dependent. Cancer Sci. 2007;98(12):1845–52. doi: 10.1111/j.1349-7006.2007.00619.x. [DOI] [PubMed] [Google Scholar]
- 64.Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, Prueitt RL, Yanaihara N, Lanza G, Scarpa A, Vecchione A, Negrini M, Harris CC, Croce CM. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci US A. 2006;103(7):2257–61. doi: 10.1073/pnas.0510565103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kallio JP, Mahlamaki EH, Helin H, Karhu R, Kellokumpu-Lehtinen P, Tammela TL. Chromosomal gains and losses detected by comparative genomic hybridization and proliferation activity in renal cell carcinoma. Scand J Urol Nephrol. 2004;38(3):225–30. doi: 10.1080/00365590310025399. [DOI] [PubMed] [Google Scholar]
- 66.Moch H, Presti JC, Jr, Sauter G, Buchholz N, Jordan P, Mihatsch MJ, Waldman FM. Genetic aberrations detected by comparative genomic hybridization are associated with clinical outcome in renal cell carcinoma. Cancer Res. 1996;56(1):27–30. [PubMed] [Google Scholar]
- 67.Slaby O, Jancovicova J, Lakomy R, Svoboda M, Poprach A, Fabian P, Kren L, Michalek J, Vyzula R. Correction: Expression of miRNA-106b in conventional renal cell carcinoma is a potential marker for prediction of early metastasis after nephrectomy. J Exp Clin Cancer Res. 2010;29(1):105. doi: 10.1186/1756-9966-29-90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Seliger B, Dressler SP, Wang E, Kellner R, Recktenwald CV, Lottspeich F, Marincola FM, Baumgartner M, Atkins D, Lichtenfels R. Combined analysis of transcriptome and proteome data as a tool for the identification of candidate biomarkers in renal cell carcinoma. Proteomics. 2009;9(6):1567–81. doi: 10.1002/pmic.200700288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Seliger B, Menig M, Lichtenfels R, Atkins D, Bukur J, Halder TM, Kersten M, Harder A, Ackermann A, Beck J, Muehlenweg B, Brenner W, Melchior S, Kellner R, Lottspeich F. Identification of markers for the selection of patients undergoing renal cell carcinoma-specific immunotherapy. Proteomics. 2003;3(6):979–90. doi: 10.1002/pmic.200300404. [DOI] [PubMed] [Google Scholar]
- 70.Sarto C, Frutiger S, Cappellano F, Sanchez JC, Doro G, Catanzaro F, Hughes GJ, Hochstrasser DF, Mocarelli P. Modified expression of plasma glutathione peroxidase and manganese superoxide dismutase in human renal cell carcinoma. Electrophoresis. 1999;20(17):3458–66. doi: 10.1002/(SICI)1522-2683(19991101)20:17<3458::AID-ELPS3458>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 71.Lichtenfels R, Kellner R, Bukur J, Beck J, Brenner W, Ackermann A, Seliger B. Heat shock protein expression and anti-heat shock protein reactivity in renal cell carcinoma. Proteomics. 2002;2(5):561–70. doi: 10.1002/1615-9861(200205)2:5<561::AID-PROT561>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 72.Balabanov S, Zimmermann U, Protzel C, Scharf C, Klebingat KJ, Walther R. Tumour-related enzyme alterations in the clear cell type of human renal cell carcinoma identified by two-dimensional gel electrophoresis. Eur J Biochem. 2001;268(22):5977–80. doi: 10.1046/j.0014-2956.2001.02546.x. [DOI] [PubMed] [Google Scholar]
- 73.Zimmermann U, Balabanov S, Giebel J, Teller S, Junker H, Schmoll D, Protzel C, Scharf C, Kleist B, Walther R. Increased expression and altered location of annexin IV in renal clear cell carcinoma: a possible role in tumour dissemination. Cancer Lett. 2004;209(1):111–8. doi: 10.1016/j.canlet.2003.12.002. [DOI] [PubMed] [Google Scholar]
- 74.Seliger B, Lichtenfels R, Atkins D, Bukur J, Halder T, Kersten M, Harder A, Ackermann A, Malenica B, Brenner W, Zobawa M, Lottspeich F. Identification of fatty acid binding proteins as markers associated with the initiation and/or progression of renal cell carcinoma. Proteomics. 2005;5(10):2631–40. doi: 10.1002/pmic.200401264. [DOI] [PubMed] [Google Scholar]
- 75.Unwin RD, Craven RA, Harnden P, Hanrahan S, Totty N, Knowles M, Eardley I, Selby PJ, Banks RE. Proteomic changes in renal cancer and co-ordinate demonstration of both the glycolytic and mitochondrial aspects of the Warburg effect. Proteomics. 2003;3(8):1620–32. doi: 10.1002/pmic.200300464. [DOI] [PubMed] [Google Scholar]
- 76.Dallmann K, Junker H, Balabanov S, Zimmermann U, Giebel J, Walther R. Human agmatinase is diminished in the clear cell type of renal cell carcinoma. Int J Cancer. 2004;108(3):342–7. doi: 10.1002/ijc.11459. [DOI] [PubMed] [Google Scholar]
- 77.Craven RA, Stanley AJ, Hanrahan S, Dods J, Unwin R, Totty N, Harnden P, Eardley I, Selby PJ, Banks RE. Proteomic analysis of primary cell lines identifies protein changes present in renal cell carcinoma. Proteomics. 2006;6(9):2853–64. doi: 10.1002/pmic.200500549. [DOI] [PubMed] [Google Scholar]
- 78.Poznanovic S, Wozny W, Schwall GP, Sastri C, Hunzinger C, Stegmann W, Schrattenholz A, Buchner A, Gangnus R, Burgemeister R, Cahill MA. Differential radioactive proteomic analysis of microdissected renal cell carcinoma tissue by 54 cm isoelectric focusing in serial immobilized pH gradient gels. J Proteome Res. 2005;4(6):2117–25. doi: 10.1021/pr050218q. [DOI] [PubMed] [Google Scholar]
- 79.Craven RA, Hanrahan S, Totty N, Harnden P, Stanley AJ, Maher ER, Harris AL, Trimble WS, Selby PJ, Banks RE. Proteomic identification of a role for the von Hippel Lindau tumour suppressor in changes in the expression of mitochondrial proteins and septin 2 in renal cell carcinoma. Proteomics. 2006;6(13):3880–93. doi: 10.1002/pmic.200500811. [DOI] [PubMed] [Google Scholar]
- 80.Hwa JS, Park HJ, Jung JH, Kam SC, Park HC, Kim CW, Kang KR, Hyun JS, Chung KH. Identification of proteins differentially expressed in the conventional renal cell carcinoma by proteomic analysis. J Korean Med Sci. 2005;20(3):450–5. doi: 10.3346/jkms.2005.20.3.450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Perroud B, Lee J, Valkova N, Dhirapong A, Lin PY, Fiehn O, Kultz D, Weiss RH. Pathway analysis of kidney cancer using proteomics and metabolic profiling. Mol Cancer. 2006;5:64. doi: 10.1186/1476-4598-5-64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Okamura N, Masuda T, Gotoh A, Shirakawa T, Terao S, Kaneko N, Suganuma K, Watanabe M, Matsubara T, Seto R, Matsumoto J, Kawakami M, Yamamori M, Nakamura T, Yagami T, Sakaeda T, Fujisawa M, Nishimura O, Okumura K. Quantitative proteomic analysis to discover potential diagnostic markers and therapeutic targets in human renal cell carcinoma. Proteomics. 2008;8(15):3194–203. doi: 10.1002/pmic.200700619. [DOI] [PubMed] [Google Scholar]
- 83.Siu KW, DeSouza LV, Scorilas A, Romaschin AD, Honey RJ, Stewart R, Pace K, Youssef Y, Chow TF, Yousef GM. Differential protein expressions in renal cell carcinoma: new biomarker discovery by mass spectrometry. J Proteome Res. 2009;8(8):3797–807. doi: 10.1021/pr800389e. [DOI] [PubMed] [Google Scholar]
- 84.Perroud B, Ishimaru T, Borowsky AD, Weiss RH. Grade-dependent proteomics characterization of kidney cancer. Mol Cell Proteomics. 2009;8(5):971–85. doi: 10.1074/mcp.M800252-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Lichtenfels R, Dressler SP, Zobawa M, Recktenwald CV, Ackermann A, Atkins D, Kersten M, Hesse A, Puttkammer M, Lottspeich F, Seliger B. Systematic comparative protein expression profiling of clear cell renal cell carcinoma: a pilot study based on the separation of tissue specimens by two-dimensional gel electrophoresis. Mol Cell Proteomics. 2009;8(12):2827–42. doi: 10.1074/mcp.M900168-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Kim DS, Choi YP, Kang S, Gao MQ, Kim B, Park HR, Choi YD, Lim JB, Na HJ, Kim HK, Nam YP, Moon MH, Yun HR, Lee DH, Park WM, Cho NH. Panel of candidate biomarkers for renal cell carcinoma. J Proteome Res. 2010;9(7):3710–9. doi: 10.1021/pr100236r. [DOI] [PubMed] [Google Scholar]
- 87.Seliger B, Dressler SP, Lichtenfels R, Kellner R. Candidate biomarkers in renal cell carcinoma. Proteomics. 2007;7(24):4601–12. doi: 10.1002/pmic.200700415. [DOI] [PubMed] [Google Scholar]
- 88.Craven RA, Banks RE. Understanding and managing renal cell carcinoma: can proteomic studies contribute to clinical practice. Contrib Nephrol. 2008;160:88–106. doi: 10.1159/000125936. [DOI] [PubMed] [Google Scholar]
- 89.Higgins JP, Shinghal R, Gill H, Reese JH, Terris M, Cohen RJ, Fero M, Pollack JR, van de Rijn M, Brooks JD. Gene expression patterns in renal cell carcinoma assessed by complementary DNA microarray. Am J Pathol. 2003;162(3):925–32. doi: 10.1016/S0002-9440(10)63887-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Tan MH, Rogers CG, Cooper JT, Ditlev JA, Maatman TJ, Yang X, Furge KA, Teh BT. Gene expression profiling of renal cell carcinoma. Clin Cancer Res. 2004;10(18 Pt 2):6315S–21S. doi: 10.1158/1078-0432.CCR-050002. [DOI] [PubMed] [Google Scholar]
- 91.Wang E, Lichtenfels R, Bukur J, Ngalame Y, Panelli MC, Seliger B, Marincola FM. Ontogeny and oncogenesis balance the transcriptional profile of renal cell cancer. Cancer Res. 2004;64(20):7279–87. doi: 10.1158/0008-5472.CAN-04-1597. [DOI] [PubMed] [Google Scholar]
- 92.Yang XJ, Sugimura J, Schafernak KT, Tretiakova MS, Han M, Vogelzang NJ, Furge K, Teh BT. Classification of renal neoplasms based on molecular signatures. J Urol. 2006;175(6):2302–6. doi: 10.1016/S0022-5347(06)00255-2. [DOI] [PubMed] [Google Scholar]
- 93.Young AN, Amin MB, Moreno CS, Lim SD, Cohen C, Petros JA, Marshall FF, Neish AS. Expression profiling of renal epithelial neoplasms: a method for tumor classification and discovery of diagnostic molecular markers. Am J Pathol. 2001;158(5):1639–51. doi: 10.1016/S0002-9440(10)64120-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Farh KK, Grimson A, Jan C, Lewis BP, Johnston WK, Lim LP, Burge CB, Bartel DP. The widespread impact of mammalian MicroRNAs on mRNA repression and evolution. Science. 2005;310(5755):1817–21. doi: 10.1126/science.1121158. [DOI] [PubMed] [Google Scholar]
- 95.Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, Bartel DP, Linsley PS, Johnson JM. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature. 2005;433(7027):769–73. doi: 10.1038/nature03315. [DOI] [PubMed] [Google Scholar]
- 96.Huang JC, Babak T, Corson TW, Chua G, Khan S, Gallie BL, Hughes TR, Blencowe BJ, Frey BJ, Morris QD. Using expression profiling data to identify human microRNA targets. Nat Methods. 2007;4(12):1045–9. doi: 10.1038/nmeth1130. [DOI] [PubMed] [Google Scholar]
- 97.Margolin AA, Ong SE, Schenone M, Gould R, Schreiber SL, Carr SA, Golub TR. Empirical Bayes analysis of quantitative proteomics experiments. PLoS One. 2009;4(10):e7454. doi: 10.1371/journal.pone.0007454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Yang Y, Chaerkady R, Kandasamy K, Huang TC, Selvan LD, Dwivedi SB, Kent OA, Mendell JT, Pandey A. Identifying targets of miR-143 using a SILAC-based proteomic approach. Mol Biosyst. 2010;6(10):1873–82. doi: 10.1039/c004401f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Li J, Min R, Bonner A, Zhang Z. A probabilistic framework to improve microrna target prediction by incorporating proteomics data. J Bioinform Comput Biol. 2009;7(6):955–72. doi: 10.1142/s021972000900445x. [DOI] [PubMed] [Google Scholar]
- 100.Selbach M, Schwanhausser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N. Widespread changes in protein synthesis induced by microRNAs. Nature. 2008;455(7209):58–63. doi: 10.1038/nature07228. [DOI] [PubMed] [Google Scholar]
- 101.Diakoumis E, Sourvinos G, Kiaris H, Delakas D, Cranidis A, Spandidos DA. Genetic instability in renal cell carcinoma. Eur Urol. 1998;33(2):227–32. doi: 10.1159/000019541. [DOI] [PubMed] [Google Scholar]
- 102.Jiang F, Richter J, Schraml P, Bubendorf L, Gasser T, Sauter G, Mihatsch MJ, Moch H. Chromosomal imbalances in papillary renal cell carcinoma: genetic differences between histological subtypes. Am J Pathol. 1998;153(5):1467–73. doi: 10.1016/S0002-9440(10)65734-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Klatte T, Rao PN, de Martino M, LaRochelle J, Shuch B, Zomorodian N, Said J, Kabbinavar FF, Belldegrun AS, Pantuck AJ. Cytogenetic profile predicts prognosis of patients with clear cell renal cell carcinoma. J Clin Oncol. 2009;27(5):746–53. doi: 10.1200/JCO.2007.15.8345. [DOI] [PubMed] [Google Scholar]
- 104.Kuroda N, Tamura M, Shiotsu T, Nakamura S, Taguchi T, Tominaga A, Hes O, Michal M, Kawada C, Shuin T, Lee GH. Chromosomal abnormalities of clear cell renal cell carcinoma: frequent gain of chromosome 7. Pathol Int. 2010;60(1):9–13. doi: 10.1111/j.1440-1827.2009.02467.x. [DOI] [PubMed] [Google Scholar]