

Abstract
In vivo pyruvate synthesis by malic enzyme (ME) and pyruvate kinase and in vivo malate synthesis by phosphoenolpyruvate carboxylase and the Krebs cycle were measured by 13C incorporation from [1-13C]glucose into glucose-6-phosphate, alanine, glutamate, aspartate, and malate. These metabolites were isolated from maize (Zea mays L.) root tips under aerobic and hypoxic conditions. 13C-Nuclear magnetic resonance spectroscopy and gas chromatography-mass spectrometry were used to discern the positional isotopic distribution within each metabolite. This information was applied to a simple precursor-product model that enabled calculation of specific metabolic fluxes. In respiring root tips, ME was found to contribute only approximately 3% of the pyruvate synthesized, whereas pyruvate kinase contributed the balance. The activity of ME increased greater than 6-fold early in hypoxia, and then declined coincident with depletion of cytosolic malate and aspartate. We found that in respiring root tips, anaplerotic phosphoenolpyruvate carboxylase activity was high relative to ME, and therefore did not limit synthesis of pyruvate by ME. The significance of in vivo pyruvate synthesis by ME is discussed with respect to malate and pyruvate utilization by isolated mitochondria and intracellular pH regulation under hypoxia.
The role of malate in many important metabolic processes contributing to energy production, biosynthesis, and mineral nutrition in plants has long been recognized (Lance and Rustin, 1984), yet our quantitative understanding of how the many reactions of malate metabolism contribute to plant function is rudimentary. One notable example concerns the fueling of mitochondria in plant cells oxidizing carbohydrate. It has been widely considered that malate is an important substrate for mitochondria, such that a significant fraction of glycolytic products enters the Krebs cycle via the combined action of PEPC, malate dehydrogenase, and ME rather than via PK (see Fig. 1) (Fowler, 1974; Day and Hanson, 1977; Wiskich, 1980; Bryce and ap Rees, 1985). However, the validity of this model remains to be unequivocally demonstrated in intact plant cells (for review, see ap Rees, 1990; Douce and Neuburger, 1990; Lambers, 1990).
Figure 1.

Schematic representation of glycolysis and the Krebs cycle highlighting the theoretical isotopic distribution (•) within specific metabolites derived from [1-13C]Glc. Numbers represent routes of synthesis for the specific metabolites discussed in the text. Enzymes: (a) PK; (b) PEPC; (c) Ala aminotransferase; (d) ME; (e) Glu dehydrogenase, Glu synthetase/Glu synthase, and aminotransferases; and (f) Asp aminotransferase. PPP, Pentose phosphate pathway.
One general strategy to solve this problem relies on using NMR and/or GC-MS to observe the metabolism of 13C-labeled substrates, and then analyzing labeling patterns by various models to deduce metabolic fluxes (for review, see Künnecke, 1995). We previously presented qualitative evidence that in [1-13C]Glc-labeled maize (Zea mays L.) root tips ME is active under hypoxia (Roberts et al., 1992), and may play a role in cytoplasmic pH regulation via proton consumption (Davies, 1986; Roberts et al., 1992). Dieuaide-Noubhani et al. (1995) compared the labeling pattern of Glu and Ala in oxygenated maize root tips labeled with [1-13C]Glc, and, using a complex metabolic model, deduced that little malate was converted to pyruvate via ME.
In the present study we measured 13C enrichment in precursors and products for reactions catalyzed by ME and PK, and deduced relative fluxes through each pathway using a simpler model. We show that ME has very low activity in respiring root tips, but is activated approximately 6-fold during the first few minutes of hypoxia. We also used this simple precursor-product approach to measure the in vivo activities of other important enzymes of malate metabolism, PEPC, and enzymes of the Krebs cycle, and discuss their role in respiration and biosynthesis.
MATERIALS AND METHODS
Maize (Zea mays L.) seeds (B73, Pioneer Hi-Bred International, Des Moines, IA) were soaked for approximately 24 h in flowing, deionized water, and were then allowed to germinate between wet paper towels in a foil-covered tray for approximately 48 h. Root tips 4 to 5 mm long were cut on ice with a razor blade and washed with deionized water to remove root cap slime. Washed root tips were transferred to 10-mL syringe barrels, each with a plastic mesh septum.
Perfusion Conditions
Four 10-mL syringes containing 2 to 6 g of root tips were connected in series using tubing and rubber stoppers. A peristaltic pump recirculated 100 mL of oxygenated medium for 12 to 15 h at 30 to 40 mL/min at room temperature. This medium contained 50 mm [1-13C]Glc in 10 mm Mes (brought to pH 6.5 with Tris), 0.1 mm CaSO4, 50 mg/L gentamycin, 2.5 mg/L amphotericin, and 2.5 mm (NH4)2SO4. [1-13C]Glc was obtained from Isotech (Miamisburg, OH). For experiments requiring hypoxic treatment, samples were perfused with N2-saturated 0.1 mm CaSO4 at 10 to 20 mL/min without recirculation.
Individual syringes were removed at the appropriate times and immediately frozen in liquid N2 and stored until extraction. The treatment conditions used here are similar to those used in earlier studies of metabolism in maize root tips (Roberts et al., 1992; Roberts and Xia, 1996). 13C labeling of root tip metabolites to steady state was done with recirculation to minimize the use of expensive [1-13C]Glc. (NH4)2SO4 and Glc were also omitted from the hypoxia treatment because no significant effects of these components on hypoxic metabolism were found in earlier experiments (Roberts et al., 1992).
Metabolite Extraction and Fractionation
Low-molecular-weight metabolites were extracted with 4% perchloric acid and centrifuged at 2000g. TABA (100 or 50 mm) was added to each sample just before extraction as an internal standard. The supernatant was neutralized with KOH, centrifuged, and placed on an H+-exchange column (model AG50W-X8, Bio-Rad). After being washed with water, amino acids were eluted with 4 n NH4OH, lyophilized, and reconstituted in 800 μL of 2H2O for NMR analysis.
After NMR analysis, amino acids were further fractionated to separate Glu and Asp from other amino acids, because the high temperatures of derivatization and GC-MS converts Gln and Asn into derivatives of Glu and Asp, respectively. This was accomplished by conversion of an AG 1-X8 chloride column (Bio-Rad) to an acetate column using 3 m sodium acetate. The column was then washed with three bed loads of deionized water. Amino acids were removed from the NMR tube and placed on the column. One-milliliter fractions were eluted with 0.5 m acetic acid, which resulted in three ninhydrin peaks. Peaks were pooled, lyophilized, and reconstituted in 700 μL of 2H2O.
Glu and Asp were identified as the second and third peaks, respectively, by NMR. All other amino acids were eluted in the first peak (in column void volume). These fractions were frozen at −20°C and saved for derivatization and GC-MS analysis. The total enrichment of Glu, as measured by GC-MS, was not statistically different from the total enrichment of unseparated Glu and Gln measured in the total amino acid fractions (data not shown). Organic acids, including Glc-6-P, eluted in the void volume of the H+-exchange column were separated from neutral metabolites by anion-exchange chromatography (AG 1-X8 formate, Bio-Rad). Organic acids and Glc-6-P were eluted in bulk with 5 n formic acid. The eluant was split into aliquots for either GC-MS analysis of malate or dephosphorylation and derivatization of Glc-6-P before drying and further processing (described below). Ala, Asp, Glu, and malate were assayed enzymatically (Bergmeyer, 1974).
NMR Spectroscopy
All NMR spectra were obtained using a spectrometer (model GN500, General Electric). 13C-NMR data were collected at 125.7 MHz with pulses every 15 s, a spectral width of 24 kHz, and 32,000 data points. TABA, which has distinct signals across the 13C chemical-shift range, was used as an internal standard, and proton decoupling was applied only during data acquisition, which, together with the long pulse interval, alleviated the need for nuclear Overhauser corrections (Roberts and Xia, 1995). Assignments are based on correspondence with chemical shifts of standards (Roberts et al., 1992; Roberts and Xia, 1995).
GC-MS
Amino acid samples (25 μL) were converted to heptafluorobutyryl isobutyl esters using a protocol modified from MacKenzie and Tenaschuk (1979). Samples were lyophilized in Teflon-capped vials to which 200 μL of 3 n HCl in isobutanol was added. Reaction vials were then placed in a silicon oil bath at 120°C for 20 min. Samples were allowed to return to room temperature, and then unreacted reagents were evaporated in an N2 stream. Next, 140 μL of ethyl acetate and 60 μL of heptafluorobutyric anhydride (Sigma) were added to dried samples, which were then heated at 150°C in the oil bath for 10 min. Samples were again evaporated in an N2 stream and resuspended in ethyl acetate to a suitable concentration for GC-MS. Malate was derivatized as a heptafluorobutyryl isobutyl ester, as described above. This novel derivative of malate provided three ions (m/e 443, 387, and 331) that were similar in abundance.
Glc-6-P was dephosphorylated by the addition of 20 to 100 units of alkaline phosphatase (Sigma) for 2 h at room temperature. Samples were then deproteinized with 4% perchloric acid (as described above under “Metabolite Extraction and Fractionation”). The released Glc was derivatized to an aldonitrile penta-acetate, as described by Katz et al. (1989). Lyophilized samples were mixed with 0.5% (w/v) hydroxylamine hydrochloride in pyridine and heated in an oil bath at 100°C for 1 h. Unreacted reagents were evaporated in a stream of N2. One hundred microliters of pyridine and 20 μL of acetic anhydride were added to the evaporated sample and left to react for 20 min at room temperature. Excess reagents were again evaporated in a N2 stream and samples were brought to volume with ethyl acetate for analysis by GC-MS.
One microliter of each derivatized sample (100– 500 ng/μL) was injected into a single-quadrapole gas chromatograph-mass spectrometer (models HP5890 and HP5989, respectively, Hewlett-Packard), and the data were analyzed by the Chem Station program (HP59940, Hewlett-Packard). The source temperature was maintained at 200°C. Methane chemical ionization resulted in the molecular ion or the molecular ion-57 (the isobutyl group) as the base ion for all metabolites measured. The mass scan range was m/e 60 to 600. Single-ion monitoring was then used to measure the abundances of m/e 286, 287, 288, and 289 for Ala; m/e 442, 443, and 444 for Asp; m/e 456, 457, and 458 for Glu; m/e 443, 444, and 445 for malate; and m/e 328, 329, and 330 for Glc. Derivatized products were confirmed with standards and, except for malate, published values (MacKenzie and Hogge, 1977; MacKenzie and Tenaschuk, 1979; Katz et al., 1989). Peak areas were used for quantitation. All analyses were performed on a 30-m × 0.25-mm capillary column of 5% phenyl (DB 5, J&W Scientific, Folsom, CA) with He carrier gas at 4 mL/min. The temperature program for amino acids and malate was 60 to 250°C at 20°C/min, and was isothermal for Glc at 235°C. The injector temperature was maintained at 240°C for all applications.
Isotopically labeled standards mixed with natural abundance standards to four different 13C enrichments were used to generate standard curves. The known atom percent of the excess 13C was plotted against the relative abundances of the specific ion clusters represented as:
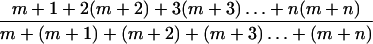 |
1 |
where the number of 13C atoms contributing to each ionic abundance within the ion cluster of a given molecule was divided by the total abundance of the cluster. The standard curves were within 5% of each value of theoretical curves, and correlation coefficients of the regression lines were higher than 0.999 (Beylot et al., 1986). The isotopic enrichments of maize root samples were then determined by the equation y = mx + b, where y represents Equation 1, m is the slope of the standard curve, and b is the y intercept. The distribution of 13C within Asp was found to be statistically identical to that in malate, reflecting the rapidity of transaminase and malate dehydrogenase action in vivo.
Determination of Relative C Fluxes
The absolute enrichment of each C in Ala, Glu, Asp, and malate was determined as follows: First, relative amounts of 13C in each position of these metabolites were directly determined from the relative peak areas of each 13C-NMR signal (e.g. Ala C1, C2, or C3; see Roberts and Xia, 1995). Relative percent 13C enrichments so obtained were then multiplied by the total isotopic enrichment determined from GC-MS spectra to give the absolute-enrichment 13C at each C position. C in metabolites not derived from [1-13C]Glc has a 13C enrichment of 1.1% (natural abundance). Positional isotopic enrichment of Glc-6-P was not determined, only its total enrichment. These measurements do not depend on percentage of recovery of metabolites through various procedures described above, because recovery is not discriminatory between different isotopomers.
To determine the sensitivity of NMR for determination of natural-abundance 13C enrichment, Ala was isolated from control maize root tips (fed only natural-abundance Glc under normoxic or hypoxic conditions). 13C abundance was determined using the ratio of peak heights from 13C-NMR spectra of each individual C of Ala and TABA (internal standard). The ratio of total amounts of Ala and TABA was measured from GC-MS spectra. Measurement of the 13C enrichment for each individual C of Ala was determined to be 1.021 ± 0.103% 13C (mean ± sd, n = 27, from nine separate samples). Relative NMR peak heights for each individual C of natural-abundance Ala were shown to be equal at 32 ± 3.4, 36 ± 1.7, and 32 ± 2.6%, respectively (mean ± sd), in root-tip extracts.
From these data on relative and absolute 13C enrichments, relative C fluxes through specific pathways were determined using the following assumptions: (a) Metabolic and isotopic steady state has been reached. This assumption was validated by measurements of 13C enrichment in metabolites (data not shown). (b) Exchange of mitochondrial, cytosolic, and vacuolar metabolites is rapid relative to metabolic fluxes. Studies of malate metabolism support this assumption (see Chang and Roberts, 1989, 1991; Kalt et al., 1990). (c) There is insignificant channeling of metabolites down specific pathways, i.e. the isotopic composition of enzyme substrates is the same as the bulk isotopic composition of metabolites. There is little information available with which to judge the validity of this assumption. Studies such as those presented here may lead to insights on the direct transfer of metabolites between enzymes (see Voet and Voet, 1995). (d) The pathways of synthesis for the metabolites studied here are the only significant metabolic reactions occurring in vivo. This assumption is justified by the fact that pathways such as gluconeogenesis and the glyoxylate cycle are not significant in excised maize root tips.
Under these conditions, the observed enrichment of the product C is equal to the percent contribution V1 from precursor A and the percent contribution V2 from precursor B:
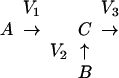 |
Scheme 1 |
where
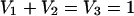 |
2 |
and V3 may represent the sum of more than one outgoing reaction (e.g. consumption of pyruvate for oxidation and fermentation reactions).
If there is a difference between the positional isotopic enrichments in molecules A and B, the contribution of A and B to C can be deduced using the expression:
 |
3 |
where Ce is the observed isotopic enrichment of a specific C atom within molecule C, and Ae and Be are the observed isotopic enrichments of the precursor atoms in molecules A and B, respectively, that are converted to the C atom having enrichment Ce. This analysis can be applied to each atom in the precursor and the product. The sensitivity with which V1 and V2 can be measured increases as the difference between Ae and Be increases.
During hypoxia Ala levels increase, so we cannot assume steady state. Therefore, a correction for the dilution of newly synthesized label incorporated into the Ala pool was applied. The correction determines the actual isotopic enrichment of the Ala synthesized between sequential time points i and f, Aa, by the following equation:
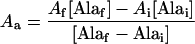 |
4 |
where Af and Ai are the 13C isotopic enrichments of Ala from the newest time point and the previous time point, respectively. This correction represents the extreme case in which Ala synthesized during the designated time interval is simply added to the preexisting and inert pool of Ala. In contrast, the steady-state model assumes that there is no inert pool of Ala. These two models represent the two extreme possibilities; the actual relative fluxes under nonsteady-state conditions lie somewhere in between.
RESULTS
ME Activity Is Negligible Relative to PK in Respiring Maize Root Tips
We determined fluxes through different pathways of pyruvate synthesis in [1-13C]Glc-fed root tips from the isotopic labeling of Ala. High transaminase activity in vivo relative to other enzymatic activities results in equivalent labeling of pyruvate and Ala (Dieuaide-Noubhani et al., 1995). The relative distribution of 13C label within Ala depends on the pathway used for pyruvate synthesis, as shown in Table I (see also Fig. 1). The predicted 13C enrichments shown in Table I were determined from the 13C enrichments of the precursors Glc-6-P and malate, measured as described in Methods. Ala synthesized from Glc-6-P by the classic glycolytic pathway via PK is labeled at C3 (Voet and Voet, 1995). In contrast, Ala synthesized from malate via ME is labeled at all three Cs (Fig. 1, reactions 1, 2, and 3); C2 and C3 of malate are similarly labeled as a result of randomization by fumarase activity (Osmond and Holtum, 1981), and C1 is labeled after multiple turns of the Krebs cycle (Chance et al., 1983). Ala synthesis from [1-13C]Glc that cycles through the pentose phosphate pathway (Fig. 1) will not change the labeling pattern of Ala, but dilutes the label incorporated into C3 Ala relative to Glc-6-P enrichment (Dieuaide-Noubhani et al., 1995).
Table I.
Predicted and observed 13C enrichments of Ala, reflecting dual pathways of synthesis in oxygenated [1-13C]Glc-labeled maize root tips
| C | Pathway
|
||
|---|---|---|---|
| Glycolysis only (Glc-6-P → Ala)a | ME (Malate → Ala)a | Glycolysis + MEb | |
| % | |||
| C1 | 1.1 | 7.42 ± 1.6 | 2.16 ± 0.36 |
| C2c | 1.1 | 17.45 ± 1.2 | 1.59 ± 0.37 |
| C3 | 31.92 ± 3.88 | 17.54 ± 2.0 | 25.73 ± 2.1 |
Predicted enrichments of Ala synthesized exclusively via either glycolysis or ME activity were deduced from 13C enrichments of Glc-6-P and malate, respectively. Values are means ± sd (n = 4).
Predicted.
Observed in vivo.
Only C2 Ala was used to measure in vivo ME activity (see text).
By comparing the 13C enrichment at C1, C2, and C3 Ala derived from the alternative precursors Glc-6-P and malate via the indicated pathway (Table I), it is clear that C2 Ala is most suited for measuring relative activities of ME and PK. First, 13C enrichment at C2 Ala differs by more than 15-fold when comparing synthesis via PK or ME, and so provides a more sensitive indicator than enrichment at either C1 or C3, which differ by less than 7- and 2-fold, respectively (Table I). Second, 13C enrichment at C2 Ala is not sensitive to operation of the pentose phosphate pathway, unlike labeling at C3 Ala (Dieuaide-Noubhani et al., 1995). We therefore used the 13C enrichment of C2 Ala and its precursors to measure the fractional contribution of PK and ME to pyruvate synthesis, as described in Methods (Scheme 1 and Eqs. 2 and 3). In oxygenated maize root tips, 13C enrichment at C2 Ala was found to be 1.59 ± 0.37% (Table I), whereas the precursors C2 and C5 of Glc-6-P were both 1.1.% 13C (natural abundance) (Dieuaide-Noubhani et al., 1995), and the precursor C2 of malate was 17.47 ± 1.2%. Solving Equations 2 and 3 with these values indicates that only 3 ± 1.1% of the pyruvate in vivo was synthesized via ME, the balance being synthesized by PK.
ME Is Activated during Early Hypoxia
We previously described an increase in 13C incorporation into C2 Ala in hypoxic root tips, and presented qualitative evidence that this increase was caused by the action of ME (Roberts et al., 1992). Our goal here was to quantify ME activity after the onset of hypoxia to test the validity of our previous conclusions. Ala levels increase during hypoxia (Roberts et al., 1992; Xia and Roberts, 1994), and so the approach to measurement of ME activity taken in the previous section was adapted to the nonsteady-state condition, as described in Methods. We observed a greater than 2-fold increase in the incorporation of 13C into C2 Ala in root tips within 20 min of the onset of hypoxia (Fig. 2A).
Figure 2.

A, 13C incorporation into C2 Ala in maize root tips during hypoxia. Data points are mean values ± se (n = 4). B, Relative contribution of ME and PK to in vivo pyruvate synthesis determined using a steady-state (•) or a nonsteady-state (▴) model (see Methods). Data points are mean values ± ranges, the latter calculated using extreme values from sd values from 13C enrichments.
Flux analysis using Equations 2, 3, and 4 indicates that the activity of ME relative to PK increases approximately 6-fold during the first few minutes of hypoxia (Fig. 2B). The two data sets shown in Figure 2B show that analysis using either the steady-state model or the nonsteady-state model yields very similar results, indicating that the changes in levels of Ala do not cause significant changes in isotopic enrichment relative to the steady-state condition before the onset of hypoxia. The activation of ME early in hypoxia followed the rapid depletion of intracellular Asp (Fig. 3B), and roughly paralleled the changes in intracellular malate (Fig. 3A).
Figure 3.

Tissue contents of malate (A) and Asp (B) in maize root tips during hypoxia. Values are means ± sd and were determined by enzymatic analysis of root-tip extracts (see Methods) (n = 4).
Anaplerotic Malate Synthesis via PEPC in Respiring Root Tips
The 13C isotopic distribution in malate and Glu was used to estimate the contribution of the anaplerotic C flux to malate via PEPC. Malate is enriched differently depending on whether it is synthesized anaplerotically via PEPC or via the Krebs cycle, as shown in Table II. If malate is synthesized via PEPC, its precursor is PEP (Fig. 1, reaction 4); if it is synthesized via the Krebs cycle, it is derived from αKG (Fig. 1, reaction 5). The labeling of PEP was determined as described in Table II, whereas the labeling pattern in Glu reflects that in αKG (Dieuaide-Noubhani et al., 1995). We found that the 13C enrichment of malate lies between predicted values for each pathway (Table II), indicating that both are active sources of malate in vivo.
Table II.
Predicted and observed 13C enrichments of malate, reflecting dual pathways of synthesis in oxygenated [1-13C]Glc-labeled maize root tips
| Ca | Pathway
|
||
|---|---|---|---|
| TCA only (Glu → malate)b | PEPC only (PEP → malate)b | PEPC + TCAc | |
| % | |||
| C1 | 9.97 ± 0.99 | 5.8 | 7.42 ± 1.6 |
| C2 | 22.3 ± 1.82 | 12.8 ± 1.1 | 17.47 ± 1.2 |
| C3 | 22.3 ± 1.82 | 12.8 ± 1.1 | 17.54 ± 1.9 |
| C4 | 9.97 ± 0.99 | 5.8 | 6.31 ± 0.62 |
Predicted enrichments of malate synthesized exclusively via the TCA cycle or PEPC were deduced from 13C enrichments of Glu and PEP, respectively. 13C of PEP was determined as described by Dieuaide-Noubhani et al. (1995). The enrichment of carboxyl groups of malate synthesized exclusively from PEP via PEPC was deduced from enrichment of 13CO2 that would be generated from respired Glc-6-P. Values are means ± sd (n = 4).
All four C's were used to measure in vivo PEPC activity (see text).
Predicted.
Observed in vivo.
Quantitation of the relative C fluxes to malate via each pathway was determined individually from C1, C2, C3, and C4 malate using both predicted and actual enrichment values for each C (Table II). All four Cs of malate were used because they are similarly sensitive to changes in relative flux between the two pathways, in contrast to the measurements of ME/PK, described above. By solving Equations 2 and 3 using the values given in Table II, we determined that PEPC contributed 62 ± 5.2% (mean ± se) of the malate synthesized in respiring root tips.
The large PEPC flux, comparable to that through the Krebs cycle, can also be inferred from the pattern of 13C enrichment observed in Glu, as previously demonstrated by Dieuaide-Noubhani et al. (1995). In the absence of anaplerotic activity, steady-state enrichment of Glu C is a simple reflection of labeling of pyruvate, as described by Chance et al. (1983) (Table III). The observation of lower enrichments in C1, C2, and C3 than in C4 of Glu (Table III) reflects the action of PEPC, which dilutes the 13C label in these first three C's (Dieuaide-Noubhani et al., 1995). We found that the predicted 13C enrichment in Glu, calculated from observed enrichments of the precursors malate and pyruvate/Ala, matches the observed enrichment in Glu (Table III). This observation supports the validity of the model of Dieuaide-Noubhani et al. (1995).
Table III.
Predicted and observed 13C enrichments of Glu, reflecting dual pathways of synthesis in oxygenated [1-13C]Glc-labeled maize root tips
| C | Pathway
|
||
|---|---|---|---|
| PDH/TCA only (Ala → Glu)a | PEPC = PDH (Malate + Ala → Glu)a | PEPC + PDHb | |
| C1 | 12.86 | 6.31 ± 0.62 | 7.07 ± 1.14 |
| C2 | 25.73 ± 2.1 | 17.54 ± 1.9 | 18.2 ± 1.79 |
| C3 | 25.73 ± 2.1 | 17.47 ± 1.6 | 19.29 ± 1.76 |
| C4 | 25.73 ± 2.1 | 25.73 ± 2.1 | 25.26 ± 1.89 |
| C5 | 1.59 ± 0.37 | 1.59 ± 0.37 | 1.75 ± 0.19 |
Predicted enrichments of Glu synthesized via the TCA cycle operating either exclusively in catabolic mode (PEPC flux = 0) or exclusively in anabolic mode (PEPC flux = PDH flux) were deduced from 13C enrichments of Ala alone (from the steady-state result described by Chance et al., 1983) or malate plus Ala (ignoring continued cycling of C beyond α-KG/Glu), respectively. Values are means ± sd (n = 4).
Predicted.
Observed in vivo.
PEPC Exceeds ME Activity by More Than 10-Fold in Respiring Root Tips
To relate the two paired flux measurements described above (ME/PK and PEPC/KrebsαKG→malate), it is first necessary to consider how individual C fluxes around the complete Krebs cycle will differ depending on the particular biosynthetic output. When biosynthesis is absent, the net flux of C through each Krebs cycle step will be equal, and likewise if the Krebs cycle's biosynthetic output is exclusively from oxaloacetate to Asp. In contrast, if the biosynthetic output of the Krebs cycle is exclusively from αKG to Glu, then the flux from malate to αKG will be proportionately higher than the flux from αKG to malate (Fig. 1).
Similarly, the rate of entry of pyruvate into the Krebs cycle (equal to the combined action of PK and ME) will increase relative to the flux from αKG to malate when the biosynthetic flux to Glu increases (Fig. 1). From these considerations, we determined the limits of the relative fluxes via PK, ME, and PEPC in respiring root tips (Table IV). It is clear that the anaplerotic flux via PEPC is comparable in magnitude to the rate of pyruvate entry into the Krebs cycle. Furthermore, the flux to malate via PEPC is at least 1 order of magnitude greater than the flux from malate via ME.
Table IV.
Summary of relative enzyme fluxes in oxygenated maize root tips
| Enzymes | Relative Flux | Defined Flux | Constraint |
|---|---|---|---|
| ME/PK | 0.03 ± 0.011 | Malate to pyruvate/PEP to pyruvate | None |
| PEPC/KrebsαKG→malate | 0.62 ± 0.05 | PEP to malate/α-KG to malate | None |
| PK/PEPC | 0.67a | PEP to pyruvate/PEP to malate | Synthesis of Asp only (Fig. 1) |
| PK/PEPC | 1.8b | PEP to pyruvate/PEP to malate | Synthesis of Glu only (Fig. 1) |
| PK/PEPC | 1 | PEP to pyruvate/PEP to malate | Synthesis of Asp and Glu equal |
| PEPC/ME | 20c | PEP to malate/malate to pyruvate | Synthesis of Asp only |
| PEPC/ME | 12d | PEP to malate/malate to pyruvate | Synthesis of Glu only |
| PEPC/ME | 17 | PEP to malate/malate to pyruvate | Synthesis of Asp and Glu equal |
Relative enzyme activities were either measured or deduced under specific physiological restrictions based on the results shown in Tables I–III. Errors represent se.
From line 2 and the relationship KrebsαKG→malate = PK + ME.
From line 2 and the relationship KrebsαKG→malate + PEPC = PK + ME.
From lines 1 to 3.
From lines 1, 2, and 4.
DISCUSSION
The Contribution of ME to Respiratory Activity
Using the combined analytical methods of GC-MS and NMR, we have measured in vivo activities of the enzymes ME and PK. We found that during respiration, ME activity accounts for approximately 3% of the pyruvate synthesized in respiring maize root tips (Table IV). These results are consistent with the analysis of Dieuaide-Noubhani et al. (1995), and support their model relating labeling patterns of Glu and malate. However, our results are contrary to results obtained in isolated mitochondria. For example, Day and Hanson (1977) determined that in isolated mitochondria from maize, ME accounted for 42% of the pyruvate used by the Krebs cycle.
It has been widely accepted that mitochondrial ME may serve to compensate for limited pyruvate transport across the mitochondrial membrane (Day and Hanson, 1977; Brailsford et al., 1986; Hill et al., 1994) by providing the Krebs cycle with pyruvate under high-energy demands (for review, see Wiskich, 1980; ap Rees, 1990; Douce and Neuburger, 1990; Lambers, 1990). However, our work here indicates that this does not appear to be the case in excised maize root tips. Although we cannot at this point exclude the possibility that plant tissues other than maize root tips might rely more on ME to fuel respiration, we suggest the possibility that the greater activity of ME observed in isolated maize mitochondria reflects nothing more than its apparent activation by low pH, as discussed in the next section.
Regulation of ME Activity in Vivo and in Vitro
We observed that in respiring root tips the flux to malate via PEPC is a least 1 order of magnitude greater than the rate of malate consumption by ME (Table IV). This indicates that ME activity is not limited by the supply of malate in vivo. Furthermore, we demonstrate that ME is activated approximately 6-fold during the first few minutes of hypoxia (Fig. 2B), which is consistent with our earlier hypothesis (Roberts et al., 1992). The activation of ME in hypoxia, which we measured relative to flux through PK (Table IV), represents an increase in absolute activity, not an inhibition of PK, given that glycolytic flux increases during hypoxia (Roberts et al., 1984).
These observations indicate that ME activity is somehow suppressed in oxygenated, respiring root tips, and we speculate that this inhibition is primarily caused by the high cytoplasmic pH (approximately 7.6) in oxygenated root tips (Roberts et al., 1992). There is significant circumstantial evidence that the behavior of ME in root tips under high O2 and hypoxia is regulated by pH. First, ME activity in vitro is strongly affected by pH, increasing when the reaction pH decreases between 7.6 and 6.5 (Davies and Patil, 1974; Wedding and Black, 1983). Second, cytoplasmic pH decreases rapidly from approximately 7.5 to 6.9 during the first few minutes of hypoxia (Roberts et al., 1984, 1992). Third, ME activity in isolated mitochondria is activated at low pH (Neuburger and Douce, 1980; Wedding and Whatley, 1984), indicating that cytoplasmic acidosis can be sensed by mitochondrial ME. Fourth, studies with isolated mitochondria show that significant pyruvate synthesis via ME was obtained with buffers of pH 6.8 to 7.2 (Day and Hanson, 1977; Brailsford et al., 1986; Hill et al., 1994).
These observations provide an explanation for our result that very little pyruvate synthesis occurs via ME in respiring root tips (Table IV). Although ME is activated by cytoplasmic acidosis, in vivo this condition is met only under severe hypoxia, in which respiration becomes negligible. Hence, it would seem that only under unusual conditions will both high respiratory demand and cytoplasmic acidosis exist together in plant cells, and allow ME to fulfill its widely considered role in plant respiration (for review, see Wiskich, 1980; ap Rees, 1990; Lambers, 1990; Douce and Neuburger, 1990).
This narrow view clearly awaits experimental studies of ME activities in other plant tissues and under different physiological conditions. In addition to regulation by pH, ME activity may also be influenced by levels of malate (Davies and Patil, 1974; Wedding and Black, 1983; Davies, 1984), given that the concentration of cytoplasmic malate in excised maize root tips is approximately 3.5 mm (Chang and Roberts, 1991). Consistent with this possibility is the coincident activation of ME (Fig. 2B) and the increase in malate during the first few minutes of hypoxia (Fig. 3A). This increase in malate is at least in part attributable to its synthesis from Asp, which is depleted (Fig. 3B) (Roberts et al., 1992).
The decrease in ME activity after approximately 20 min of hypoxia (Fig. 2B) may be partially caused by a slight increase in cytoplasmic pH after the initial large acidification (Roberts et al., 1992; Roberts and Xia, 1996). Indeed, we have suggested that the action of ME early in hypoxia may serve to offset cytoplasmic acidification, because this reaction consumes protons (Davies and Patil, 1974; Davies, 1980; Roberts et al., 1992). Ultimately, however, ME activity in hypoxic root tips is limited by cytoplasmic malate, because both intracellular Asp and malate become depleted (Fig. 3). The residual intracellular malate found after 1 h or more of hypoxia is predominantly located in the vacuole (Roberts, 1993), and is therefore essentially unavailable for decarboxylation by ME.
The Relationship of PEPC to the Krebs Cycle in Maize Root Tips
The flux through PEPC is comparable in magnitude to the rate of glycolysis (which we measured at the level of PK) (Table IV) and the rate of malate synthesis via the Krebs cycle (Table IV). In contrast, very little malate synthesized by PEPC is converted to pyruvate via ME (Table IV). We therefore conclude that the principal role of PEPC in oxygenated maize root tips is anaplerotic, and therefore sustains biosynthesis rather than respiration.
What is the contribution of the Krebs cycle in biosynthesis associated with PEPC activity? If the Krebs cycle is drained via oxaloacetate, PEPC activity requires support from neither PK nor any part of the Krebs cycle. In contrast, if PEPC activity sustains synthesis of Glu, there is a requirement for stoichiometric participation of PK and the Krebs cycle (from oxaloacetate to αKG) (see Fig. 1). Hence, the nature of the biosynthetic output resulting from anaplerotic action of PEPC determines the relative activities of PEPC, PK, and different parts of the Krebs cycle. This interdependence is evident in Table IV, in which we describe the activity of PEPC with respect to the flux from αKG to malate. Given this ratio of activities, we show that the accompanying flux through PK will vary from 0.67 to 1.8 times the PEPC flux (Table IV), depending on whether Asp or Glu is the principal product.
Although the flux measurements in the present study do not allow distinction between these possibilities, consideration of previous work on inorganic C metabolism is useful. First, it has long been known that the respiratory quotient in oxygenated root tips is close to 1 (Beevers, 1961). A large flux from Glc to Asp will give less net CO2 evolution relative to O2 consumption, whereas inorganic C fixed by PEPC to sustain synthesis of Glu is accompanied by decarboxylation, and so in this case O2 consumption (associated with recycling of pyridine nucleotides) approximates CO2 release. This analysis indicates that in maize root tips the flux to Asp is much smaller than that to Glu. Second, comparison of measurements of PEPC activity, obtained by following the fate of 14C- and 13C-labeled bicarbonate (Chang and Roberts, 1992) with measurements of respiration (Roberts et al., 1984), suggest that net CO2 production is severalfold higher than PEPC activity, an inference similar to that drawn by Dieuaide-Noubhani et al. (1995). As in the first case, these data appear to preclude biosynthesis of Asp as the major output of the Krebs cycle, because under the relative flux conditions shown in Table IV, line 3, CO2 evolution from pyruvate oxidation (via PK) would be considerably offset by HCO3− fixation attributable to PEPC activity. In contrast, biosynthesis of Glu under the relative flux conditions shown in Table IV, line 4, requires CO2 evolution to be more than 3-fold higher than PEPC activity, a result in greater accordance with the separate measurements just noted. Hence, these considerations both indicate that Glu is a much more important biosynthetic product than Asp in maize root tips and, as a corollary to this conclusion, that the net flux of C through the Krebs cycle is faster from malate to αKG than from αKG to malate. Furthermore, they point to the potential value of measurements of CO2 evolution, simultaneous with isotope analysis of metabolites as performed in this study, to an understanding of these important metabolic fluxes in plants.
ACKNOWLEDGMENT
We thank Dr. Philippe Raymond for helpful discussions during the preparation of the manuscript.
Abbreviations:
- αKG
α-ketoglutarate
- ME
malic enzyme
- m/e
mass-to-charge ratio
- PEPC
PEP carboxylase
- PK
pyruvate kinase
- TABA
2-aminobutylrate
Footnotes
This work was supported by National Science Foundation (NSF) grant no. IBN 9310850. S.E. was supported by a NSF Minority Graduate Research Fellowship in Plant Biochemistry.
LITERATURE CITED
- ap Rees T. Carbon metabolism in mitochondria. In: Dennis DT, Turpin DH, editors. Plant Physiology Biochemistry and Molecular Biology. Harlow, UK: Longman Scientific & Technical; 1990. pp. 106–123. [Google Scholar]
- Beevers H (1961) Respiratory Metabolism in Plants. Row, Peterson, New York
- Bergmeyer HU (1974) Methods of Enzymatic Analysis, Ed 2. Academic Press, New York
- Beylot M, Beaufrere B, Normand S, Riou JP, Cohen R, Mornex R. Determination of human ketone body kinetics using stable-isotope labelled tracers. Diabetologia. 1986;29:90–96. doi: 10.1007/BF00456116. [DOI] [PubMed] [Google Scholar]
- Brailsford MA, Thompson AG, Kaderbhai N, Beechey RB. Pyruvate metabolism in castor-bean mitochondria. Biochem J. 1986;239:355–361. doi: 10.1042/bj2390355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryce JH, ap Rees T. Rapid decarboxylation of the products of dark fixation of CO2 in roots of Pisum and Plantago. Phytochemistry. 1985;24:1635–1638. [Google Scholar]
- Chance EM, Seeholzer SH, Kobayashi K, Williamson JR. Mathematical analysis of isotope labeling in the citric acid cycle with applications to 13C NMR studies in perfused rat hearts. J Biol Chem. 1983;258:13785–13794. [PubMed] [Google Scholar]
- Chang K, Roberts JKM. Cytoplasmic malate levels in maize root tips during K+ ion uptake determined by 13C-NMR spectroscopy. Biochim Biophys Acta. 1991;1092:29–34. doi: 10.1016/0167-4889(91)90174-v. [DOI] [PubMed] [Google Scholar]
- Chang K, Roberts JKM. Quantification of rates of transport, metabolic fluxes, and cytoplasmic levels of inorganic carbon in maize root tips during K+ ion uptake. Plant Physiol. 1992;99:291–297. doi: 10.1104/pp.99.1.291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang KJ, Roberts JKM. Observation of cytoplasmic and vacuolar malate in maize root tips by 13C-NMR spectroscopy. Plant Physiol. 1989;89:197–203. doi: 10.1104/pp.89.1.197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies DD (1980) Anaerobic metabolism and the production of organic acids. In DD Davies, ed, The Biochemistry of Plants, Vol 2. Academic Press, New York, pp 581–607
- Davies DD. The co-ordination and integration of metabolite pathways. In: Palmer JM, editor. The Physiology and Biochemistry of Plant Respiration. Cambridge, UK: Cambridge University Press; 1984. pp. 159–170. [Google Scholar]
- Davies DD. The fine control of cytosolic pH. Physiol Plant. 1986;67:702–706. [Google Scholar]
- Davies DD, Patil KD. Regulation of “malic” enzyme of Solanum tuberosum by metabolites. Biochem J. 1974;137:45–53. doi: 10.1042/bj1370045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Day DA, Hanson JB. Pyruvate and malate transport and oxidation in corn mitochondria. Plant Physiol. 1977;59:630–635. doi: 10.1104/pp.59.4.630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dieuaide-Noubhani M, Raffard G, Canioni P, Pradet A, Raymond P. Quantification of compartmented metabolic fluxes in maize root tips using isotope distribution from 13C- or 14C- labeled glucose. J Biol Chem. 1995;270:13147–13159. doi: 10.1074/jbc.270.22.13147. [DOI] [PubMed] [Google Scholar]
- Douce R, Neuburger M. Metabolite exchange between the mitochondrion and the cytosol. In: Dennis DT, Turpin DH, editors. Plant Physiology Biochemistry and Molecular Biology. Harlow, UK: Longman Scientific & Technical; 1990. pp. 173–190. [Google Scholar]
- Fowler MW. Role of the malic enzyme reaction in plant roots, utilization of [2,3-14C]malate, [4-14C]malate, and [1-14C]pyruvate by pea root apices and measurement of enzyme activity. Biochim Biophys Acta. 1974;372:245–254. [Google Scholar]
- Hill AS, Bryce JH, Leaver CJ. Pyruvate metabolism in mitochondria from cucumber cotyledons during early seedling development. J Exp Bot. 1994;45:1489–1491. [Google Scholar]
- Kalt W, Osmond CB, Siedow JN. Malate metabolism in the dark after 13CO2 fixation in the Crassulacean plant Kalanchoe tubiflora. Plant Physiol. 1990;94:826–832. doi: 10.1104/pp.94.2.826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katz J, Lee W-NP, Wals PA, Bergner EA. Studies of glycogen synthesis and the Krebs cycle by mass isotopomer analysis with [U-13C]glucose in rats. J Biol Chem. 1989;264:12994–13001. [PubMed] [Google Scholar]
- Künnecke B. Application of 13C NMR spectroscopy to metabolic studies on animals. In: Beckmann N, editor. Carbon-13 NMR Spectroscopy of Biological Systems. San Diego, CA: Academic Press; 1995. pp. 159–267. [Google Scholar]
- Lambers H. Oxidation of mitochondrial NADH and the synthesis of ATP. In: Dennis DT, Turpin DH, editors. Plant Physiology Biochemistry and Molecular Biology. Harlow, UK: Longman Scientific & Technical; 1990. pp. 124–143. [Google Scholar]
- Lance C, Rustin P. The central role of malate in plant metabolism. Physiol Veg. 1984;22:625–641. [Google Scholar]
- MacKenzie SL, Hogge LR. Gas chromatography-mass spectrometry of the N(O)-hepta-fluorobutyryl isobutyl esters of the protein amino acids using electron impact ionization. J Chromatogr. 1977;132:485–493. doi: 10.1016/s0021-9673(00)82912-x. [DOI] [PubMed] [Google Scholar]
- MacKenzie SL, Tenaschuk D. Quantitative formation of N(O,S)-heptafluorobutyryl isobutyl amino acids for gas chromatographic analysis. J Chromatogr. 1979;171:195–208. [Google Scholar]
- Neuburger M, Douce R. Effect of bicarbonate and oxaloacetate on malate oxidation by spinach leaf mitochondria. Biochim Biophys Acta. 1980;589:176–189. doi: 10.1016/0005-2728(80)90036-5. [DOI] [PubMed] [Google Scholar]
- Osmond CB, Holtum JAM. Crassulacean acid metabolism. Biochem Plants. 1981;8:283–328. [Google Scholar]
- Roberts JKM. Interaction between cytoplasmic fermentation reactions and transport of protons between cytoplasm and vacuoles in maize root tips studied in vivo by NMR spectroscopy. In: Hochachka PW, Lutz PL, Sick T, Rosenthal M, van den Thillart G, editors. Surviving Hypoxia: Mechanisms of Control and Adaptation. Boca Raton, FL: CRC Press; 1993. pp. 187–200. [Google Scholar]
- Roberts JKM, Hooks MA, Miaullis AP, Edwards S, Webster C. Contribution of malate and amino acid metabolism to cytoplasmic pH regulation in hypoxic maize root tips studied using nuclear magnetic resonance spectroscopy. Plant Physiol. 1992;98:480–487. doi: 10.1104/pp.98.2.480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts JKM, Wemmer D, Jardetzky O. Measurement of mitochondrial ATPase activity in maize root tips by saturation transfer 31P nuclear magnetic resonance. Plant Physiol. 1984;74:632–639. doi: 10.1104/pp.74.3.632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts JKM, Xia JH. High-resolution NMR methods for study of higher plants. In: Galbraith DW, Bohnert HJ, Bourque DP, editors. Methods in Plant Cell Biology, Vol 49, Part A. San Diego, CA: Academic Press; 1995. pp. 245–258. [DOI] [PubMed] [Google Scholar]
- Roberts JKM, Xia JH (1996) NMR contributions to understanding of plant responses to low oxygen stress. In Y Shachar-Hill, PE Pfeffer, eds, Nuclear Magnetic Resonance in Plant Biology, Vol 16. American Society of Plant Physiologists, Rockville, MD, pp 155–180
- Voet D, Voet J. Biochemistry, Ed 2. New York, NY: John Wiley & Sons; 1995. [Google Scholar]
- Wedding RT. Malic enzymes of higher plants. Plant Physiol. 1989;90:367–371. doi: 10.1104/pp.90.2.367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wedding RT, Black K. Physical and kinetic properties and regulation of the NAD malic enzyme purified from leaves of Crassula argentea. Plant Physiol. 1983;72:1021–1028. doi: 10.1104/pp.72.4.1021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wedding RT, Whatley FR. Malate oxidation by Arum spadix mitochondria: participation and characterization of NAD-malic enzyme. New Phytol. 1984;96:505–517. [Google Scholar]
- Wiskich JT (1980) Control of the kreb cycle. In DD Davies, ed, The Biochemistry of Plants, Vol 2. Academic Press, New York, pp 243–278
- Xia JH, Roberts JKM. Improved cytoplasmic pH regulation, increased lactate efflux, and reduced cytoplasmic lactate levels are biochemical traits expressed in root tips of whole maize seedlings acclimated to a low-oxygen environment. Plant Physiol. 1994;105:651–657. doi: 10.1104/pp.105.2.651. [DOI] [PMC free article] [PubMed] [Google Scholar]