

Abstract
AIM: To study the association between inflammatory bowel disease (IBD) and genetic variations in eosinophil protein X (EPX) and eosinophil cationic protein (ECP).
METHODS: DNA was extracted from ethylene diamine tetraacetic acid blood of 587 patients with Crohn’s disease (CD), 592 with ulcerative colitis (UC) and 300 healthy subjects. The EPX405 (G > C, rs2013109), ECP434 (G > C, rs2073342) and ECP562 (G > C, rs2233860) gene polymorphisms were analysed, by the 5’-nuclease allelic discrimination assay. For determination of intracellular content of EPX and ECP in granulocytes, 39 blood samples was collected and extracted with a buffer containing cetyltrimethylammonium bromide. The intracellular content of EPX was analysed using an enzyme-linked immunosorbent assay. The intracellular content of ECP was analysed with the UniCAP® system as described by the manufacturer. Statistical tests for calculations of results were χ2 test, Fisher’s exact test, ANOVA, Student-Newman-Keuls test, and Kaplan-Meier survival curve with Log-rank test for trend, the probability values of P < 0.05 were considered statistically significant.
RESULTS: The genotype frequency for males with UC and with an age of disease onset of ≥ 45 years (n = 57) was for ECP434 and ECP562, GG = 37%, GC = 60%, CC = 4% and GG = 51%, GC = 49%, CC = 0% respectively. This was significantly different from the healthy subject’s genotype frequencies of ECP434 (GG = 57%, GC = 38%, CC = 5%; P = 0.010) and ECP562 (GG = 68%, GC = 29%,CC = 3%; P = 0.009). The genotype frequencies for females, with an age of disease onset of ≥ 45 years with CD (n = 62), was for the ECP434 and ECP562 genotypes GG = 37%, GC = 52%, CC = 11% and GG = 48%, GC = 47% and CC = 5% respectively. This was also statistically different from healthy controls for both ECP434 (P = 0.010) and ECP562 (P = 0.013). The intracellular protein concentration of EPX and ECP was calculated in μg/106 eosinophils and then correlated to the EPX 405 genotypes. The protein content of EPX was highest in the patients with the CC genotype of EPX405 (GG = 4.65, GC = 5.93, and CC = 6.57) and for ECP in the patients with the GG genotype of EPX405 (GG = 2.70, GC = 2.47 and CC = 1.90). ANOVA test demonstrated a difference in intracellular protein content for EPX (P = 0.009) and ECP (P = 0.022). The age of disease onset was linked to haplotypes of the EPX405, ECP434 and ECP562 genotypes. Kaplan Maier curve showed a difference between haplotype distributions for the females with CD (P = 0.003). The highest age of disease onset was seen in females with the EPX405CC, ECP434GC, ECP562CC haplotype (34 years) and the lowest in females with the EPX405GC, ECP434GC, ECP562GG haplotype (21 years). For males with UC there was also a difference between the highest and lowest age of the disease onset (EPX405CC, ECP434CC, ECP562CC, mean 24 years vs EPX405GC, ECP434GC, ECP562GG, mean 34 years, P = 0.0009). The relative risk for UC patients with ECP434 or ECP562-GC/CC genotypes to develop dysplasia/cancer was 2.5 (95%CI: 1.2-5.4, P = 0.01) and 2.5 (95%CI: 1.1-5.4, P = 0.02) respectively, compared to patients carrying the GG-genotypes.
CONCLUSION: Polymorphisms of EPX and ECP are associated to IBD in an age and gender dependent manner, suggesting an essential role of eosinophils in the pathophysiology of IBD.
Keywords: Eosinophil derived neurotoxin, RNase 2, RNase 3, Single nucleotide polymorphism, Inflammation bowel disease, Crohn’s disease, Ulcerative colitis
INTRODUCTION
The concept of inflammatory bowel disease (IBD) comprises the disorders Crohn’s disease (CD) and ulcerative colitis (UC). The aetiology of the diseases is unknown, but IBD is currently believed to develop in individuals with a hereditary predisposition. This is related to the reaction of the immune system to the bacterial challenge of the gut and/or defects in the gut wall, in addition to external environmental factors. IBD usually has an intermittent course of aggravations with common symptoms such as frequent diarrhoea, blood in stool, abdominal pain, and weight loss. The incidence of IBD has increased in the Western world in recent years, and new cases occur primarily in the group between 20 and 40 years of age[1,2]. CD seems to affect women more than men, while UC appears to be more prevalent among men, but some studies showed no gender differences[1,3,4]. A bimodal age distribution of disease onset has been reported and debated in population studies of CD and UC[2,5-8]. Patients with CD and UC have an increased risk of developing colorectal cancer. This risk is more pronounced in patients with UC with an overall 2.7-fold increased risk as compared with the general population[9].
Under normal conditions, the intestinal mucosa is in a state of “controlled inflammation”. Different subsets of T-cells are present in the healthy intestine, as are eosinophils and antigen-presenting cells with a predominance of anti-inflammatory and regulatory cytokine responses that keep the mucosal homeostasis intact[10]. However, in IBD the balance between mucosal responsiveness and tolerance towards antigens is disturbed, resulting in an exaggerated immune response to the commensal flora. It is well established that neutrophil granulocytes accumulate and infiltrate in the local processes in IBD. In recent years attention has been paid to the involvement of eosinophil granulocytes, since increased numbers of these cells are found in the intestinal mucosa of IBD[11,12]. Studies now emphasise a key role of the eosinophil granulocyte in the pathophysiology of IBD[13,14].
The eosinophil participate in a number of biological processes such as wound healing, defence against parasites and allergic inflammation[15]. The eosinophils granulocyte are characterised by specific cytoplasmic granules containing four major proteins; eosinophil cationic protein (ECP)[16], eosinophil peroxidase (EPO)[17], eosinophil protein X (EPX)/eosinophil derived neurotoxin[18], and major basic protein (MBP)[15]. Increased intraluminal release of EPX, ECP and EPO has been shown in patients with UC[19]. The cytotoxic activities of ECP and EPX as well as their potent RNase activities have been well documented[20] and are generally conceived as part of the host defence against viruses, parasites and helminths[21,22]. Furthermore both EPX and ECP have been associated with different types of cancer[23-25]. The presence of eosinophils in the intestinal mucosa of healthy subjects may reflect these host defence activities of the cell. In addition EPX has been identified as a member of the Alarmin family with the capacity to affect various aspects of the dendritic cells[26], and in aged asthma patients, relative to young, the degranulation of EPX was decreased[27]. Larger numbers of activated eosinophils, identified as an increased expression of the cell surface receptor CD44, have been reported both in CD and UC, but even more pronounced in quiescent than in active UC. Findings that might suggest a role of eosinophils in tissue remodelling and repair in IBD[28,29].
By linkage analysis and genome-wide association studies many genetic variations, linked to IBD, have been found[30]. The nucleotide sequences of EPX and ECP have a homology of 90%, and both genes have been located to chromosome 14q11.2[31,32] within the IBD4 locus, located at the 14q11-12 region[33]. Several single nucleotide polymorphisms (SNPs) were reported in the genes for EPX and ECP[34]. An intron SNP in the EPX gene, EPX405 G > C (rs2013109) was together with a SNP from the 3’ untranslated region in the ECP gene, ECP562 G > C (rs2233860)[35], shown to be closely linked to the eosinophil content of EPX and ECP (Jönsson et al[35], to be published). The C-allele of the nonsynonymous missense ECP434 G > C (rs2073342) polymorphism, gives rise to an arginine to threonine shift at amino acid position 97 in the mature protein[34,36]. As shown with recombinant ECP proteins this amino acid shift resulted in an alteration of the protein and its cytotoxic activity[37], but with no effect on its RNase activity[38]. The loss of cytotoxicity of ECP containing threonine at position 97 was confirmed using purified native proteins from genotyped blood donors[37]. The ECP434(G > C) polymorphism was shown to be associated with the expression of allergic symptoms[36], and with disease severity in Hodgkin lymphoma[39] in population based studies as well as to the prevalence and severity of Schistosoma mansoni infection in a Ugandan population[40].
The aim of this study was to investigate the impact of SNPs in the EPX and ECP genes in a cohort of patients with IBD. The hypothesis was that alterations in the cytotoxic activities of ECP and/or the altered expression of EPX might affect the propensity to acquire IBD and also might affect different features of IBD. The influence of polymorphisms in the EPX and ECP genes was first analysed in a pilot cohort of patients with IBD from Uppsala University Hospital and then in a second step in a bigger Swedish cohort. The possible altered expression of EPX was in addition analysed by measuring the intracellular eosinophil content of EPX in a subset of the patients.
MATERIALS AND METHODS
Study population
The patient group consisted of 1179 individuals recruited from Uppsala University Hospital, Sweden (95 CD and 100 UC), and an established Swedish cohort recruited at Karolinska University Hospital and Örebro University Hospital (492 CD and 492 UC). Diagnoses of CD and UC was based on clinical, histological and endoscopic findings, according to standardized criteria[41]. Bimodal patterns as to age of disease diagnosis were observed for both CD and UC irrespective of gender (Figure 1). The major peak of age at diagnosis of both diseases was seen at 20-25 years, with a second peak around 40-50 years.
Figure 1.
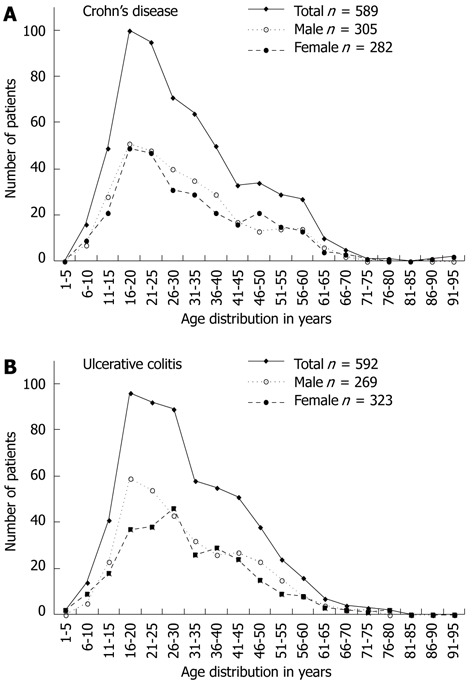
Incidence curves, given as a mean of 5 consecutive years, are shown for the inflammatory bowel disease study cohort. Curves separated by gender and displaying the total inflammatory bowel disease cohort are included.
A group of 300 healthy Swedish blood donors from the Uppsala blood bank served as reference population.
The study was approved by the local Ethics Committees of the Medical Faculties, of Uppsala University and Karolinska University Hospital, Stockholm, and all patients gave their informed consent to participate in the study (Table 1).
Table 1.
Clinical characteristic of study populations
| Crohn’s disease (n = 587) | Ulcerative colitis (n = 592) | Reference group (n = 300) | |
| Female | 282 | 269 | 132 |
| Male | 305 | 323 | 168 |
| Median age1 | 48 (18-95) | 48 (14-102) | 44 (19-73) |
| Age at disease diagnosis2 | 28 (6-93) | 28 (4-78) |
Range at sampling;
Median (range).
DNA extraction
Genomic DNA from all IBD patients and the reference group were extracted from whole ethylene diamine tetraacetic acid (EDTA) blood, by the use of the QIAamp DNA Blood Mini Kit from QIAGEN (QIAGEN Inc, Valencia, CA, United States), according to the manufactures description.
Genotyping
All IBD samples and controls were genotyped for the EPX405 (G > C), ECP434 (G > C) and the ECP562 (G > C) polymorphisms by use of the 5’-nuclease allelic discrimination assay. The assay has previously been described in detail[42]. Oligonucleotide primers were designed by the use of Primer Express® (Applied Biosystems, Foster City, CA, United States) based on the sequence of the EPX (X16546) and ECP (X16545) genes available in GenBank. Sequences for primers and probes are displayed in Table 2.
Table 2.
Primers and probes
| EPX405 (G > C): | |
| Forward primer | 5’ AAG AGA GCT GAC GTT AGT GCT TAG G 3’ |
| Reverse primer | 5’ GGT CTT GGT TAT GAC ACA CAC TGT AGT 3’ |
| Probe 1 | 5’ VIC-ACG TTG CAC ACT TT 3’ |
| Probe 2 | 5’ 6-FAM-CGT TCC ACA CTT TG 3’ |
| ECP434 (G > C): | |
| Forward primer | 5’ GAG TAG ATT CCG GGT GCC TTT ACT 3’ |
| Reverse primer | 5’ CGT GGA GAA TCC CGT GGA T 3’ |
| Probe 1 | 5’ VIC-AAA CTG CAG GTA TGC AGA 3’ |
| Probe 2 | 5’ 6-FAM-AAA CTG CAC GTA TGC AG 3’ |
| ECP562 (G > C): | |
| Forward primer | 5’ GGT TCC AGT TCA CCT GGA TAC C 3’ |
| Reverse primer | 5’ GGT ATG GAG ACT GAT GAG GAC AGT 3’ |
| Probe 1 | 5’ VIC-TCA GCA GTC CTC ATC 3’ |
| Probe 2 | 5’ FAM-TCA GCA CTC CTC ATC 3’ |
EPX: Eosinophil protein X; ECP: Eosinophil cationic protein.
Polymerase chain reaction (PCR) cycling was carried out in an ABI PRISM 7000 (Applied Biosystems) with the recommended thermal profile of: 50 °C for 2 min, 95 °C for 10 min followed by a total of 40 cycles of 95 °C for 15 s and 60 °C for 1 min. After the PCR cycling the genotypes were determined according to the application Allelic Discrimination of the ABI Prism 7000 SDS software (Applied Biosystems).
Blood cell count
Blood eosinophil count (B-Eos) (reference interval 0.0 × 109/L-0.5 × 109/L) and total white blood cell count (reference interval 3.5 × 109/L-9.0 × 109/L) were determined by an automated haematology analyser (Celldyn Sapphire; Abbott Laboratories, CA, United States) at the Department of Clinical Chemistry, Uppsala University Hospital, Uppsala, Sweden.
Whole blood extraction
The EPX and ECP polymorphisms were first only analysed in the Uppsala cohort. There was a difference in the EPX content between CD and UC patients, which lead to an interest in measuring the intracellular content of EPX and ECP in eosinophils. For the determination of intracellular content of EPX and ECP in granulocytes, EDTA blood was collected from 39 patient samples with CD. The subjects were selected based on their EPX405 (G > C) genotypes (GG = 15, GC = 16, and CC = 8). The method for whole blood extraction using cetyltrimethylammonium bromide has previously been described in detail (Jönsson et al[35], to be published). The supernatants were stored at -20 °C until assayed for EPX and ECP concentrations.
Protein assays
EPX whole cell extract was analysed using an enzyme-linked immunosorbent assay (Diagnostics Development, Uppsala, Sweden), according to the recommendations of the manufacturer with minor modifications. The concentration of ECP was analysed with the UniCAP® system, (Phadia AB, Uppsala, Sweden) as described by the manufacturer.
Statistical analysis
All the statistical analyses were performed with the Statistica 9.0 software for Windows (StatSoft, Tulsa, United States), and MedCalc software (V.10, Mariakerke, Belgium). The statistical tests in use for the calculations of results were χ2 and Fisher’s exact test, ANOVA, Student-Newman-Keuls test, and Kaplan-Meier survival curve with the Log-rank test for trend. P < 0.05 were considered statistically significant.
RESULTS
Genotype distributions in healthy population vs patients with IBD
The distribution of the EPX405, ECP434 and ECP562 genotypes was investigated in the population of 1179 subjects with CD (n = 587) or UC (n = 592) (Table 1).
For comparison the distributions of the same genotypes were investigated in a group of apparently healthy blood donors (n = 300). In the healthy reference population males had a higher prevalence of the EPX405 GG-genotype (P = 0.049) and the G-allele (P = 0.021) compared to females (Table 3). All continuing data has been calculated gender stratified for the EPX405 genotype. No difference was seen between genders for the ECP434 and ECP 562 genotype in the control cohort (Table 3). No genotype differences were seen between younger (n = 150; median age: 31 years; range: 19-43 years) and older (n = 150; median age: 54 years; range: 44-73 years) healthy references. At first only the Uppsala cohort with CD (n = 95) and UC (n = 100) was examined. Here there was a difference between patients with CD and UC (P = 0.048) with more EPX405 GC and CC in CD. These results could not be verified in the whole cohort where no general differences were found between the healthy and the patient cohorts, or between the CD and the UC cohorts, for any of the genotypes. When dividing the patient cohorts by their median (28 years) age at disease diagnosis (ADD) and comparing them, no difference was seen in any of the genotypes for younger vs older patients with CD (median ADD: 20 years; range: 6-28 years vs median ADD: 40 years; range: 28-93 years) or UC (median ADD: 20 years; range: 4-28 years vs median ADD: 40 years; range: 28-78 years). In the total cohort, no difference was seen in the genotype frequencies related to gender in patients with CD or UC. When the patient populations were separated by age at disease diagnosis, according to the Montreal classification[43] (< 16 years, 17-40 years and > 40 years) and gender, a pattern with significant differences was observed. Since the incidence curve (Figure 1) of women with CD indicated a second peak of disease onset around 45 years the age range > 45 years was also included in the study. Thus, a lower prevalence of the ECP434 and ECP562-GG genotypes and higher prevalence of the ECP434 and ECP562-GC genotypes were found in female patients with CD, compared to the healthy reference population (Table 4). In males similar differences in the prevalence of the ECP434 and ECP562 genotypes were observed, but only in patients with UC (Table 4).
Table 3.
Genotype and allelic frequency distribution among 300 healthy individuals and the total cohorts of Crohn’s disease and ulcerative colitis
| GG-genotype (%) | GC-genotype (%) | CC-genotype (%) | G-allele (%) | C-allele (%) | |
| Healthy | |||||
| ECP562 | 68 | 29 | 3 | 83 | 17 |
| ECP434 | 57 | 38 | 5 | 76 | 24 |
| EPX405 M1 | 64 | 32 | 4 | 80 | 20 |
| EPX405 F1 | 53 | 38 | 9 | 72 | 28 |
| CD | |||||
| ECP562 | 64 | 33 | 4 | 80 | 20 |
| ECP434 | 53 | 40 | 7 | 73 | 27 |
| EPX405 M | 55 | 38 | 6 | 74 | 26 |
| EPX405 F | 58 | 34 | 8 | 75 | 25 |
| UC | |||||
| ECP562 | 64 | 33 | 4 | 80 | 20 |
| ECP434 | 55 | 38 | 7 | 74 | 26 |
| EPX405 M | 62 | 31 | 7 | 78 | 22 |
| EPX405 F | 57 | 38 | 5 | 76 | 24 |
For ECP434 and ECP562 there was no difference between males and females.
In EPX405 there was a difference between the sexes according their genotype (P = 0.049) and allelic distribution (P = 0.021). UC: Ulcerative colitis; CD: Crohn’s disease; EPX: Eosinophil protein X; ECP: Eosinophil cationic protein; F: Female; M: Male.
Table 4.
Gender specific difference
|
Crohn’s disease |
Ulcerative colitis |
|||||||||||||
| Genotype distribution | Allelic frequency | Genotype distribution | Allelic frequency | |||||||||||
| Age at diagnosis | Patients | EPX | ECP | ECP | EPX | ECP | ECP | EPX | ECP | ECP | EPX | ECP | ECP | |
| (yr) | CD/UC (n) | 405 | 434 | 562 | 405 | 434 | 562 | 405 | 434 | 562 | 405 | 434 | 562 | |
| Female | ≥ 40 | 80/71 | NS | NS | NS | NS | 0.043 | 0.034 | NS | NS | NS | NS | NS | NS |
| ≥ 45 | 42/62 | NS | 0.009 | 0.013 | NS | 0.004 | 0.007 | NS | NS | NS | NS | NS | NS | |
| Male | ≥ 40 | 77/85 | NS | NS | NS | NS | NS | NS | 0.048 | 0.025 | 0.008 | 0.026 | 0.024 | 0.045 |
| ≥ 45 | 57/52 | NS | NS | NS | NS | NS | NS | NS | 0.01 | 0.009 | NS | NS | NS | |
Difference (χ2, P-values displayed) between the genotype distributions and allelic frequencies, related to age at diagnosis, in individuals with CD or UC compared with the healthy population. UC: Ulcerative colitis; CD: Crohn’s disease; NS: Not significant; EPX: Eosinophil protein X; ECP: Eosinophil cationic protein.
For both genders the genotype differences were observed in the older cohort of the population with an age at diagnosis above 40 years (Table 4). In the cohort of males with UC a higher prevalence of the EPX405 GC-genotype, compared to healthy controls, was observed in patients with an ADD of > 40 years (P = 0.048).
Genotype distribution within different cohorts
Among females with CD and ADD of > 45 years, the GC-genotype of the ECP434 (P = 0.020) and ECP562 (P = 0.014) gene polymorphisms were more common than in females with ADD < 45 years. In females with UC, no such differences were observed. In males with UC there was a difference between those with an ADD over and under 40 and 45 years, concerning all three genotypes (40 years; EPX405 P = 0.003, ECP434 P = 0.003 and ECP562 P = 0.004). The differences were, in all genotypes, due to a higher amount of the GC-genotypes among males > 40 years and > 45 years.
In Figure 2, a comparison of the ECP434 genotype distributions between patients with CD and UC and an age of disease diagnosis of > 45 years is shown. In the comparison the population is separated by gender. Among female patients with CD there was a significantly lower prevalence of the ECP434GG genotype and a prevalence of the heterozygote genotype almost twice as high as in those women having UC (P = 0.010). This was contrasted by the findings in males, since those patients having CD had a predominance of the ECP434 GG-genotype, as compared to patients with UC, who had a predominance of the GC-genotype (P = 0.004). Highly significant differences were found in ECP434 genotype distributions between females and males (> 45 years) with CD (P = 0.001) and with UC (P = 0.001). Similar patterns were found for the ECP562 genotypes, but not for the EPX405 genotypes (not shown).
Figure 2.
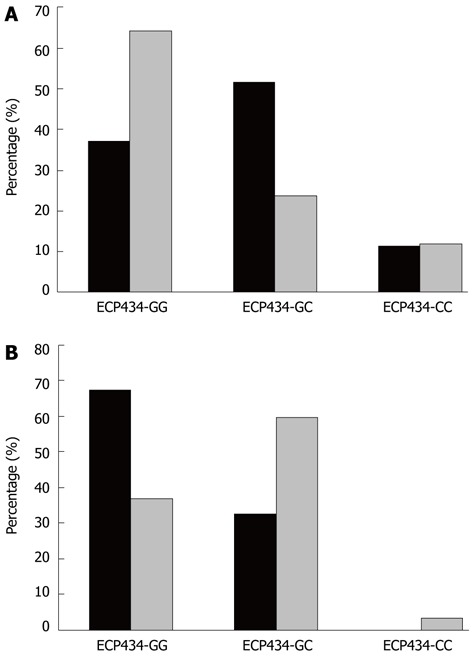
Prevalence of ECP434 genotypes in Crohn’s disease (black bars) and ulcerative colitis (grey bars). A: In females, age at disease diagnosis (ADD) > 45 years, a significant difference between Crohn’s disease (CD) and ulcerative colitis (UC) concerning ECP434 (P = 0.01, χ2) was observed; B: In men, ADD > 45 years a significant difference between CD and UC concerning ECP434 (P = 0.004) was observed. ECP: Eosinophil cationic protein.
Relationships of age at disease diagnosis to genotypes and haplotypes
The relationships of the EPX405, ECP435 and ECP562 genotypes and haplotypes (Table 5) to ADD (females with CD and males with UC) are illustrated in Figure 3. The haplotypes analysed were chosen to include the wild type (WT) (haplotype 1; GG in all three genotypes) and the opposite extreme with CC in all genotypes (haplotype 27). The criterion for other haplotypes to be included was to differ from the WT and to contain at least 10 individuals in the group. We found significant associations between the respective genotypes and ADD for men with UC; EPX405 (P = 0.007), ECP434 (P = 0.002) and ECP562 (P = 0.010) with the lowest ADD seen in those carrying the CC-genotypes and the highest ADD in those carrying the heterozygous genotypes (Figure 3C-E). To attain a better understanding of the significance of the genetic diversity of the polymorphisms in the EPX and ECP genes, four haplotypes were related to ADD. The haplotypes analysed in men with UC were haplotype 1, haplotype 13, haplotype 14 and haplotype 27 (Table 5). For females with CD the haplotypes analysed were haplotype 1, haplotype 4, haplotype 13 and haplotype 27 (Table 5).
Table 5.
Haplotype compositions and their frequencies in the studied population with inflammatory bowel disease
| Gender | Haplotype | EPX405 | ECP434 | ECP562 | Frequency % |
| Males with UC | 1 | GG | GG | GG | 48 |
| 13 | GC | GC | GG | 5 | |
| 14 | GC | GC | GC | 20 | |
| 27 | CC | CC | CC | 3 | |
| Females with CD | 1 | GG | GG | GG | 46 |
| 4 | GG | GC | GG | 6 | |
| 13 | GC | GC | GG | 5 | |
| 27 | CC | CC | CC | 5 |
UC: Ulcerative colitis; CD: Crohn’s disease; EPX: Eosinophil protein X; ECP: Eosinophil cationic protein.
Figure 3.
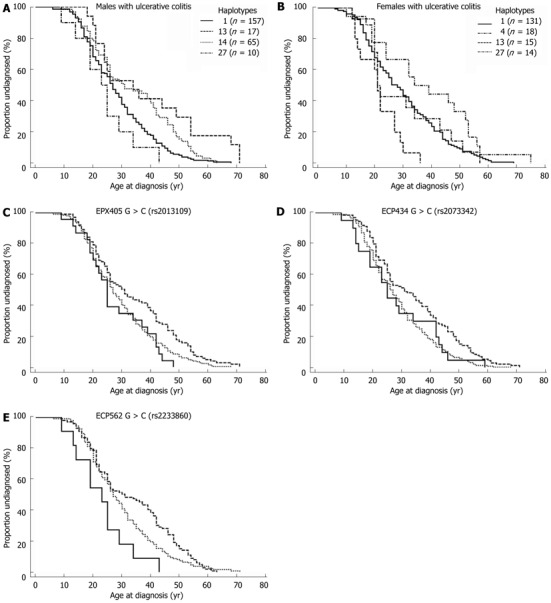
Kaplan-Meier Curve of four groups haplotypes and EPX405, ECP562 genotypes. A: Kaplan-Meier Curve of four groups of haplotypes in male patients with ulcerative colitis (UC) (P = 0.0009); B: Kaplan-Meier Curve of four groups of haplotypes in female patients with Crohn’s disease (P = 0.003); C: Kaplan-Meier curve of the EPX405 genotype in men with UC (P = 0.007); D: Curve of the ECP434 genotype in men with UC (P = 0.002); E: Curve of the ECP562 genotype in males with UC (P = 0.01). Dotted line: GG genotype; Dashed line: GC genotype; Continuous line: CC genotype. ECP: Eosinophil cationic protein; EPX: Eosinophil protein X.
As shown in Figure 3A, ADD differed significantly depending on haplotype (P = 0.0009) in men with UC and in females with CD (P = 0.003) (Figure 3B). In the female cohort extreme outliers (ADD > 80 years) were excluded from the analysis. The ADD was markedly lower in males with UC who carried the haplotype 27 as compared to those who carried the haplotype 13. Almost reverse results were obtained in females with CD since ADD in those carrying haplotype 13 was the lowest and haplotype 4 the highest.
Intracellular protein content
As noted in the introduction the eosinophil content of the alarmin EPX is associated to the EPX405 genotypes in healthy and allergic subjects. We therefore investigated the association of the cellular content in a selected subset of patients with CD from the Uppsala cohort. The eosinophil content of EPX was closely linked to the EPX405 genotype with the highest content in the subjects carrying the EPX405CC genotype (ANOVA P = 0.009) (Figure 4). The eosinophil content of ECP was also linked to the EPX405 genotype (ANOVA P = 0.022), but in a reverse mode (Figure 4).
Figure 4.
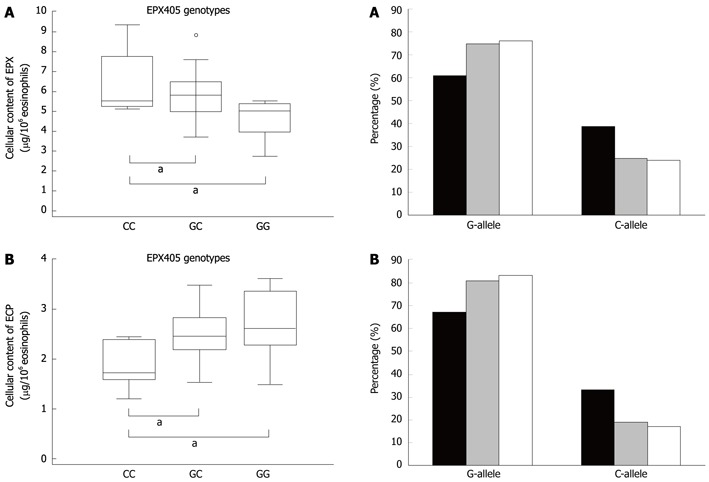
The box plots displays the association between the EPX405 genotypes and the intracellular content of eosinophil protein X and eosinophil cationic protein in eosinophils. The box plots display the distribution of the protein content within each class of genotype. A: Intracellular content of eosinophil protein X (EPX) (ANOVA P = 0.009). aP < 0.05 vs EPX405 GG genotype (Student-Newman-Keuls test); B: Intracellular content of eosinophil cationic protein (ANOVA P = 0.022). aP < 0.05 vs the EPX405 CC genotype (Student-Newman-Keuls test).
Relationship to dysplasia/cancer
In the study UC cohort there were several patients (n = 27; females: 8; males: 19) that through continuous endoscopic examination were shown to have developed dysplasia and/or cancer (D/C) (median age at cancer diagnosis: 49 years; range: 27-67 years). The D/C cohort was compared to all UC patients, females and males who had not developed D/C, since no difference was shown between older and younger UC patients in the mixed cohort. The distributions of the ECP434 and ECP562 alleles in patients with UC, with and without findings of D/C of the intestine are shown in Figure 5. The C-allele of both genotypes was more prevalent among patients with D/C (ECP434 P = 0.034 and ECP562 P = 0.019). The D/C patients also differed from the healthy controls (ECP434 P = 0.027 and ECP562 P = 0.007) (Figure 5). The relative risk (RR), for UC patients with the ECP434 GC or CC genotypes, to develop D/C was 2.5 (95%CI, 1.2-5.4) compared to other patients with UC. In patients with the ECP562 GC or CC genotypes the RR was 2.5 (95%CI, 1.2-5.8) as compared to UC patients. Considering the whole cohort, no associations to the EPX405 (G > C) genotypes were detected.
Figure 5.
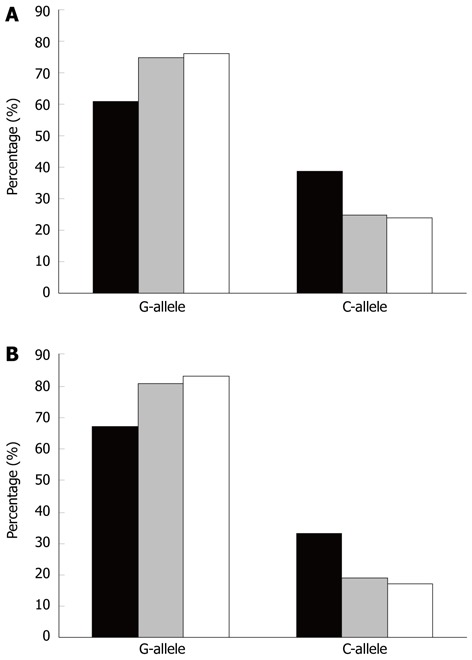
Distribution of allele frequencies, displaying the difference in patients with ulcerative colitis and dysplasia/cancer. Bars represent the allele frequency (%) of ECP434 (A) and ECP562 (B) single nucleotide polymorphism in patients with dysplasia/cancer (black bar), all patients with ulcerative colitis (grey bar), and healthy individuals (white bar). ECP: Eosinophil cationic protein.
DISCUSSION
Inflammatory bowel disease, including CD and UC, are idiopathic diseases with chronic inflammation of the gut. A key role of the eosinophil in the pathophysiology of IBD has been suggested. Our findings in this report, of intriguing associations between gene polymorphisms of the two major eosinophil granule proteins ECP and EPX, emphasize this notion. The primary results gained from the Uppsala cohort gave a hint that there was some association between the EPX and ECP polymorphisms and IBD. The associations, however, were in the larger cohort mainly found when the patients were separated into gender, age groups and diagnosis, since the associations were most apparent in older women with CD and in older men with UC. Another interesting observation was the higher prevalence of the ECP434C-allele in those patients with dysplasia/cancer, since this allele codes for the non-cytotoxic variant of ECP. We also confirmed the relationship between the EPX405 (G > C) polymorphism and the eosinophil content of EPX, since an increased content of EPX was found in subjects carrying the C-allele. The C-allele was also related to the age of disease diagnosis suggesting an impact of the alarmin EPX on the course of the disease in IBD.
In our study, we show that older females with CD had a relatively higher amount of the ECP434 GC and ECP562 GC genotypes as compared to healthy individuals, and younger females with CD. Our novel findings show that in healthy individuals and females diagnosed with CD at a young age the GG genotype of ECP434 and ECP562 was more dominating than in older women with CD, implying a more cytotoxic ECP and a higher content of ECP. This interesting finding could have a bearing on granuloma formation in CD. Granuloma formation is one of the pathological hallmarks of CD and is detected in the gut wall in 37%-60% of the patients[44]. Patients with granulomas, and particularly females, are diagnosed early in their course of disease[45]. In CD, the production of tumor growth factor (TGF)-β appears critical in CD-associated fibrosis and in the formation of granulomas[46,47]. ECP is known to promote TGF-β production and migration of fibroblasts[48,49] and the ECP434 GG genotype has been linked to increased fibrosis[40]. All together this suggests a role of ECP in the production of granulomas in younger females with CD. Furthermore high numbers of bacteria, and mainly gram negative bacteria as Escherichia coli (E. coli), can induce secretion of ECP[50]. By the use of laser capture microscopy and PCR it was shown that DNA from E. coli was present in 80% of granulomas from patients with CD[51].
There are few studies associating genetic risk factors and gender to falling ill with IBD. For patients with UC, there is a gender-specific risk associated with the interleukin-10 promoter. Females with a particular genotype and haplotype have an increased risk of contracting the disease earlier in life[52]. In patients with CD the DLG5 R30Q variant was shown to be associated with an increased risk for male patients to develop CD. This risk has been described both in the paediatric[53] and the adult age group[54]. We showed that age at diagnosis in females with CD is linked to the EPX405 genotype. Females with haplotype 13, including EPX405 GC, are diagnosed earlier in life. The longest span of years before onset of disease was observed in females carrying haplotype 4, including EPX405 GG. The C-allele of EPX405 is linked to a higher content of EPX in the eosinophils. The increased availability of EPX (in combination with unchanged amount of ECP) may result in an earlier disease onset in life. EPX was previously shown to present characteristics of an endogenous alarmin, which strengthens the immune systems Th2 immune response[26]. CD has often been regarded as a disease with a dominant Th1 immune response, but a Th2 response in the early stages of the disease with active ileitis has been shown[55,56]. EPX uses the toll-like receptor 2 (TLR2)-MyD88 signal transduction pathway to induce dendritic cell maturation and activation of NF-κB[26] and also acts as a chemotactic activator of dendritic cells[57]. Few of the intestinal dendritic cells in healthy controls express TLR2, whereas in patients with IBD the TLR2 receptor density is increased[58]. NOD2 also induces nuclear factor κB (NF-κB) activation through the MyD88 pathway[59]. The potentiated activation of NF-κB through the co-operation of EPX and NOD2 in CD, therefore, is an interesting possibility and may explain the differences found between CD and UC, since the 405C-allele was more common in CD in patients with shorter onset of disease.
In the cohort of older men with UC, the ECP434 and ECP562 GC genotypes were more common than in men with CD, females with UC, younger men with UC and the healthy population. Compared to younger men and the healthy reference population the EPX405 genotype was also significantly different, with more GC genotype in the older cohort. Thus, older men had a lower content of less cytotoxic ECP and a higher content of EPX. These results are similar to the cohort of older females with CD. Thus, from a genetic point of view, females with CD show similar distributions of the ECP and EPX SNPs as men with UC. These are interesting findings and suggest major gender differences in the pathogenesis of the two diseases.
In men with UC the Kaplan Meier analysis showed the lowest ADD in subjects carrying haplotype 27, (EPX405 CC, ECP434 CC and ECP562 CC) which codes for elevated production of EPX, less cytotoxic ECP and reduced production of ECP protein. The highest ADD was seen in those carrying haplotype 13 (EPX405 GC, ECP434 GC and ECP562 GG), which codes for increased production of EPX and cytotoxic ECP. These findings suggest a protective role of ECP, putatively against the microbial challenge, in the development of UC, whereas the role of the increased availability of EPX may be involved in the initiation of the disease. In contrast to the findings in men with UC, haplotype 13 was associated with the lowest ADD in women with CD, which should result in the increased production of cytotoxic ECP. As noted above this finding may relate to early granuloma formation. Our findings emphasize major differences in the pathophysiology and role of eosinophils in CD and UC. A notion supported by previous studies showing differences in the morphology and function of eosinophils in CD and UC[12,27].
Our findings of an association between the ECP434 and 562 SNPs and UC with complication of dysplasia/cancer suggest a role of ECP in the defence against malignant cell transformation, since the results indicate a lower production of ECP and that the produced protein is non-cytotoxic and unable to kill malignant cells[37]. The relative risk for a patient with UC and a C-allele in ECP434 or 562 to develop D/C is 2.5. When separating the cohort due to gender the EPX405 genotype was also significantly associated to D/C in males (data not shown). It is noticed that the number of patients with D/C was limited in our cohort, but recent studies have pointed out the association of ECP and EPX and cancer[23-25].
It is important to remember that the polymorphisms EPX405, ECP434 and ECP562 are in linkage disequilibrium (Jönsson et al to be published). Because of the profound linkage disequilibrium between the alleles, it is difficult to determine which allele that is the determining allele.
In conclusion, the present study has identified a gender and age related difference between polymorphisms in the EPX and ECP genes in CD and UC, and showed that the haplotype distributions are associated to the age of disease diagnosis. We also confirmed a link between the eosinophil content of EPX and the EPX405 gene polymorphism. Finally our results suggested a link between polymorphisms in the ECP gene and the risk in patients with UC of developing dysplasia/cancer. The role of neutrophils in IBD is a hallmark of the disease, and the results presented in this study suggest a key role of eosinophil granulocytes and their major granule proteins in the pathophysiology of inflammatory bowel disease.
ACKNOWLEDGMENTS
We acknowledge Inger Gustavsson and Gunilla Jansson for their skilful technical assistance.
COMMENTS
Background
A role of eosinophils is implicated in inflammatory bowel disease. The intraluminal release of eosinophil derived cytotoxic proteins is increased in patients with ulcerative colitis (UC). Larger numbers of activated eosinophils, identified as CD44-high, are seen in quiescent as opposed to active UC. Gene polymorphisms in eosinophil protein X (EPX)/eosinophil derived neurotoxin and eosinophil cationic protein (ECP) affect the eosinophil content of EPX and ECP.
Research frontiers
The cytotoxic activities of ECP and EPX as well as their potent RNase activities have been well documented and are generally conceived as part of the host defence against viruses, parasites and helminths exerted by eosinophils. Polymorphisms in the genes of EPX and ECP are known to be linked to changes of cytotoxic affect and protein content. Increased release of EPX and ECP has been shown in patients with UC.
Innovations and breakthroughs
There are no previous studies focusing on the genetics of the eosinophil cationic proteins and inflammatory bowel disease (IBD). Studies of IBD and genetics in a gender and age dependent manner are few. With the present study, the authors have combined the two, enhancing the relation between the eosinophils and IBD, and revealing the possible risk for cancer.
Applications
The study results have helped to attain an increased knowledge of the eosinophils role in IBD with focus on age and gender related differences. The results also suggest that the determination of the EPX and ECP genotypes may contribute in patients risk stratification for cancer development.
Terminology
Eosinophil granulocyte: the eosinophil granulocyte is a pro inflammatory cell. The cell participates in wound healing and host defence against parasites and helminths. The cell is also present in the process of the allergic inflammation. Eosinophil cationic proteins: the cationic proteins are granula proteins secreted by activated eosinophils. The proteins are eosinophil specific and are therefore often used as markers for the eosinophil activities. Single nucleotide polymorphism (SNP): this is a variation in the DNA sequence, occurring when a single nucleotide (A, C, G or T) in the genome, differs between paired chromosomes in an individual or in between individuals. If a SNP is present in the coding region of a gene this may result in an amino acid change, influencing the phenotype of a specific gene.
Peer review
The authors of this paper report that polymorphisms in the EPX and ECP genes are associated with IBD and contribute to the risk for the disease development in a gender-specific manner. Furthermore, these polymorphisms seem to be specifically related to the late-onset disease and to increased risk for colorectal cancer in UC patients. The results of this large study are certainly interesting, especially with the attention paid to gender-and age-stratified analysis which is a typical shortcoming of most of the genetic studies in IBD.
Footnotes
Supported by Grants from the Swedish Research Council-Medicine; the Hedlunds Foundation and from the Medical Faculty, Uppsala University, Uppsala, Sweden
Peer reviewer: Dr. Zuzana Zelinkova, Institute of Erasmus MC, Postbus 2040, 3000 CA Rotterdam, The Netherlands
S- Editor Gou SX L- Editor A E- Editor Zhang DN
References
- 1.Hanauer SB. Inflammatory bowel disease: epidemiology, pathogenesis, and therapeutic opportunities. Inflamm Bowel Dis. 2006;12 Suppl 1:S3–S9. doi: 10.1097/01.mib.0000195385.19268.68. [DOI] [PubMed] [Google Scholar]
- 2.Rönnblom A, Samuelsson SM, Ekbom A. Ulcerative colitis in the county of Uppsala 1945-2007: incidence and clinical characteristics. J Crohns Colitis. 2010;4:532–536. doi: 10.1016/j.crohns.2010.03.003. [DOI] [PubMed] [Google Scholar]
- 3.Brant SR, Nguyen GC. Is there a gender difference in the prevalence of Crohn’s disease or ulcerative colitis? Inflamm Bowel Dis. 2008;14 Suppl 2:S2–S3. doi: 10.1002/ibd.20540. [DOI] [PubMed] [Google Scholar]
- 4.Russel MG, Stockbrügger RW. Epidemiology of inflammatory bowel disease: an update. Scand J Gastroenterol. 1996;31:417–427. doi: 10.3109/00365529609006759. [DOI] [PubMed] [Google Scholar]
- 5.Evans JG, Acheson ED. An epidemiological study of ulcerative colitis and regional enteritis in the Oxford area. Gut. 1965;6:311–324. doi: 10.1136/gut.6.4.311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Johnston RD, Logan RF. What is the peak age for onset of IBD? Inflamm Bowel Dis. 2008;14 Suppl 2:S4–S5. doi: 10.1002/ibd.20545. [DOI] [PubMed] [Google Scholar]
- 7.Loftus CG, Loftus EV, Harmsen WS, Zinsmeister AR, Tremaine WJ, Melton LJ, Sandborn WJ. Update on the incidence and prevalence of Crohn’s disease and ulcerative colitis in Olmsted County, Minnesota, 1940-2000. Inflamm Bowel Dis. 2007;13:254–261. doi: 10.1002/ibd.20029. [DOI] [PubMed] [Google Scholar]
- 8.Sonnenberg A. Age distribution of IBD hospitalization. Inflamm Bowel Dis. 2010;16:452–457. doi: 10.1002/ibd.21058. [DOI] [PubMed] [Google Scholar]
- 9.Söderlund S, Brandt L, Lapidus A, Karlén P, Broström O, Löfberg R, Ekbom A, Askling J. Decreasing time-trends of colorectal cancer in a large cohort of patients with inflammatory bowel disease. Gastroenterology. 2009;136:1561–1567; quiz 1561-1567. doi: 10.1053/j.gastro.2009.01.064. [DOI] [PubMed] [Google Scholar]
- 10.Coombes JL, Robinson NJ, Maloy KJ, Uhlig HH, Powrie F. Regulatory T cells and intestinal homeostasis. Immunol Rev. 2005;204:184–194. doi: 10.1111/j.0105-2896.2005.00250.x. [DOI] [PubMed] [Google Scholar]
- 11.Carvalho AT, Elia CC, de Souza HS, Elias PR, Pontes EL, Lukashok HP, de Freitas FC, Lapa e Silva JR. Immunohistochemical study of intestinal eosinophils in inflammatory bowel disease. J Clin Gastroenterol. 2003;36:120–125. doi: 10.1097/00004836-200302000-00006. [DOI] [PubMed] [Google Scholar]
- 12.Coppi LC, Thomazzi SM, de Ayrizono ML, Coy CS, Fagundes WJ, Goes JR, Franchi GC, Nowill AE, Montes CG, Antunes E, et al. Comparative study of eosinophil chemotaxis, adhesion, and degranulation in vitro in ulcerative colitis and Crohn’s disease. Inflamm Bowel Dis. 2007;13:211–218. doi: 10.1002/ibd.20018. [DOI] [PubMed] [Google Scholar]
- 13.Wedemeyer J, Vosskuhl K. Role of gastrointestinal eosinophils in inflammatory bowel disease and intestinal tumours. Best Pract Res Clin Gastroenterol. 2008;22:537–549. doi: 10.1016/j.bpg.2007.12.001. [DOI] [PubMed] [Google Scholar]
- 14.Jeziorska M, Haboubi N, Schofield P, Woolley DE. Distribution and activation of eosinophils in inflammatory bowel disease using an improved immunohistochemical technique. J Pathol. 2001;194:484–492. doi: 10.1002/path.904. [DOI] [PubMed] [Google Scholar]
- 15.Venge P, Bergstrand H, Hakansson L. Neutrophils and Eosinophils. In: Kelley WN, Harris ED, Ruddy S, Sledge CB, editors. Textbook of Rheumatology. Philadelphia: WB Saunders; 1996. pp. 146–160. [Google Scholar]
- 16.Olsson I, Venge P. Cationic proteins of human granulocytes. II. Separation of the cationic proteins of the granules of leukemic myeloid cells. Blood. 1974;44:235–246. [PubMed] [Google Scholar]
- 17.Carlson MG, Peterson CG, Venge P. Human eosinophil peroxidase: purification and characterization. J Immunol. 1985;134:1875–1879. [PubMed] [Google Scholar]
- 18.Peterson CG, Venge P. Purification and characterization of a new cationic protein--eosinophil protein-X (EPX)--from granules of human eosinophils. Immunology. 1983;50:19–26. [PMC free article] [PubMed] [Google Scholar]
- 19.Carlson M, Raab Y, Peterson C, Hällgren R, Venge P. Increased intraluminal release of eosinophil granule proteins EPO, ECP, EPX, and cytokines in ulcerative colitis and proctitis in segmental perfusion. Am J Gastroenterol. 1999;94:1876–1883. doi: 10.1111/j.1572-0241.1999.01223.x. [DOI] [PubMed] [Google Scholar]
- 20.Gullberg U, Widegren B, Arnason U, Egesten A, Olsson I. The cytotoxic eosinophil cationic protein (ECP) has ribonuclease activity. Biochem Biophys Res Commun. 1986;139:1239–1242. doi: 10.1016/s0006-291x(86)80310-2. [DOI] [PubMed] [Google Scholar]
- 21.Gleich GJ, Loegering DA, Bell MP, Checkel JL, Ackerman SJ, McKean DJ. Biochemical and functional similarities between human eosinophil-derived neurotoxin and eosinophil cationic protein: homology with ribonuclease. Proc Natl Acad Sci USA. 1986;83:3146–3150. doi: 10.1073/pnas.83.10.3146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.McLaren DJ, Peterson CG, Venge P. Schistosoma mansoni: further studies of the interaction between schistosomula and granulocyte-derived cationic proteins in vitro. Parasitology. 1984;88(Pt 3):491–503. doi: 10.1017/s0031182000054755. [DOI] [PubMed] [Google Scholar]
- 23.Sun JY, Huang Q, Wang AD, Dong J, Shao NY, Lan Q. [Bioinformatic analysis of glioma development relative genes] Ai Zheng. 2003;22:225–229. [PubMed] [Google Scholar]
- 24.Trulson A, Nilsson S, Venge P. The eosinophil granule proteins in serum, but not the oxidative metabolism of the blood eosinophils, are increased in cancer. Br J Haematol. 1997;98:312–314. doi: 10.1046/j.1365-2141.1997.2203035.x. [DOI] [PubMed] [Google Scholar]
- 25.Ye B, Skates S, Mok SC, Horick NK, Rosenberg HF, Vitonis A, Edwards D, Sluss P, Han WK, Berkowitz RS, et al. Proteomic-based discovery and characterization of glycosylated eosinophil-derived neurotoxin and COOH-terminal osteopontin fragments for ovarian cancer in urine. Clin Cancer Res. 2006;12:432–441. doi: 10.1158/1078-0432.CCR-05-0461. [DOI] [PubMed] [Google Scholar]
- 26.Yang D, Chen Q, Su SB, Zhang P, Kurosaka K, Caspi RR, Michalek SM, Rosenberg HF, Zhang N, Oppenheim JJ. Eosinophil-derived neurotoxin acts as an alarmin to activate the TLR2-MyD88 signal pathway in dendritic cells and enhances Th2 immune responses. J Exp Med. 2008;205:79–90. doi: 10.1084/jem.20062027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mathur SK, Schwantes EA, Jarjour NN, Busse WW. Age-related changes in eosinophil function in human subjects. Chest. 2008;133:412–419. doi: 10.1378/chest.07-2114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lampinen M, Rönnblom A, Amin K, Kristjansson G, Rorsman F, Sangfelt P, Säfsten B, Wagner M, Wanders A, Winqvist O, et al. Eosinophil granulocytes are activated during the remission phase of ulcerative colitis. Gut. 2005;54:1714–1720. doi: 10.1136/gut.2005.066423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lampinen M, Backman M, Winqvist O, Rorsman F, Rönnblom A, Sangfelt P, Carlson M. Different regulation of eosinophil activity in Crohn’s disease compared with ulcerative colitis. J Leukoc Biol. 2008;84:1392–1399. doi: 10.1189/jlb.0807513. [DOI] [PubMed] [Google Scholar]
- 30.Ishihara S, Aziz MM, Yuki T, Kazumori H, Kinoshita Y. Inflammatory bowel disease: review from the aspect of genetics. J Gastroenterol. 2009;44:1097–1108. doi: 10.1007/s00535-009-0141-8. [DOI] [PubMed] [Google Scholar]
- 31.Zhang J, Dyer KD, Rosenberg HF. RNase 8, a novel RNase A superfamily ribonuclease expressed uniquely in placenta. Nucleic Acids Res. 2002;30:1169–1175. doi: 10.1093/nar/30.5.1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cho S, Beintema JJ, Zhang J. The ribonuclease A superfamily of mammals and birds: identifying new members and tracing evolutionary histories. Genomics. 2005;85:208–220. doi: 10.1016/j.ygeno.2004.10.008. [DOI] [PubMed] [Google Scholar]
- 33.Duerr RH, Barmada MM, Zhang L, Pfützer R, Weeks DE. High-density genome scan in Crohn disease shows confirmed linkage to chromosome 14q11-12. Am J Hum Genet. 2000;66:1857–1862. doi: 10.1086/302947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zhang J, Rosenberg HF. Sequence variation at two eosinophil-associated ribonuclease loci in humans. Genetics. 2000;156:1949–1958. doi: 10.1093/genetics/156.4.1949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Jönsson UB, Byström J, Stålenheim G, Venge P. A (G->C) transversion in the 3' UTR of the human ECP (eosinophil cationic protein) gene correlates to the cellular content of ECP. J Leukoc Biol. 2006;79:846–851. doi: 10.1189/jlb.0904517. [DOI] [PubMed] [Google Scholar]
- 36.Jönsson UB, Byström J, Stålenheim G, Venge P. Polymorphism of the eosinophil cationic protein-gene is related to the expression of allergic symptoms. Clin Exp Allergy. 2002;32:1092–1095. doi: 10.1046/j.1365-2222.2002.01410.x. [DOI] [PubMed] [Google Scholar]
- 37.Trulson A, Byström J, Engström A, Larsson R, Venge P. The functional heterogeneity of eosinophil cationic protein is determined by a gene polymorphism and post-translational modifications. Clin Exp Allergy. 2007;37:208–218. doi: 10.1111/j.1365-2222.2007.02644.x. [DOI] [PubMed] [Google Scholar]
- 38.Rubin J, Zagai U, Blom K, Trulson A, Engström A, Venge P. The coding ECP 434(G>C) gene polymorphism determines the cytotoxicity of ECP but has minor effects on fibroblast-mediated gel contraction and no effect on RNase activity. J Immunol. 2009;183:445–451. doi: 10.4049/jimmunol.0803912. [DOI] [PubMed] [Google Scholar]
- 39.Molin D, Glimelius B, Sundström C, Venge P, Enblad G. The serum levels of eosinophil cationic protein (ECP) are related to the infiltration of eosinophils in the tumours of patients with Hodgkin’s disease. Leuk Lymphoma. 2001;42:457–465. doi: 10.3109/10428190109064602. [DOI] [PubMed] [Google Scholar]
- 40.Eriksson J, Reimert CM, Kabatereine NB, Kazibwe F, Ireri E, Kadzo H, Eltahir HB, Mohamed AO, Vennervald BJ, Venge P. The 434(G>C) polymorphism within the coding sequence of Eosinophil Cationic Protein (ECP) correlates with the natural course of Schistosoma mansoni infection. Int J Parasitol. 2007;37:1359–1366. doi: 10.1016/j.ijpara.2007.04.001. [DOI] [PubMed] [Google Scholar]
- 41.Sands BE. From symptom to diagnosis: clinical distinctions among various forms of intestinal inflammation. Gastroenterology. 2004;126:1518–1532. doi: 10.1053/j.gastro.2004.02.072. [DOI] [PubMed] [Google Scholar]
- 42.Livak KJ. Allelic discrimination using fluorogenic probes and the 5’ nuclease assay. Genet Anal. 1999;14:143–149. doi: 10.1016/s1050-3862(98)00019-9. [DOI] [PubMed] [Google Scholar]
- 43.Satsangi J, Silverberg MS, Vermeire S, Colombel JF. The Montreal classification of inflammatory bowel disease: controversies, consensus, and implications. Gut. 2006;55:749–753. doi: 10.1136/gut.2005.082909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Simillis C, Jacovides M, Reese GE, Yamamoto T, Tekkis PP. Meta-analysis of the role of granulomas in the recurrence of Crohn disease. Dis Colon Rectum. 2010;53:177–185. doi: 10.1007/DCR.0b013e3181b7bfb0. [DOI] [PubMed] [Google Scholar]
- 45.Freeman HJ. Granuloma-positive Crohn’s disease. Can J Gastroenterol. 2007;21:583–587. doi: 10.1155/2007/917649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Burke JP, Mulsow JJ, O’Keane C, Docherty NG, Watson RW, O’Connell PR. Fibrogenesis in Crohn’s disease. Am J Gastroenterol. 2007;102:439–448. doi: 10.1111/j.1572-0241.2006.01010.x. [DOI] [PubMed] [Google Scholar]
- 47.Appleton I, Tomlinson A, Colville-Nash PR, Willoughby DA. Temporal and spatial immunolocalization of cytokines in murine chronic granulomatous tissue. Implications for their role in tissue development and repair processes. Lab Invest. 1993;69:405–414. [PubMed] [Google Scholar]
- 48.Zagai U, Lundahl J, Klominek J, Venge P, Sköld CM. Eosinophil cationic protein stimulates migration of human lung fibroblasts in vitro. Scand J Immunol. 2009;69:381–386. doi: 10.1111/j.1365-3083.2009.02233.x. [DOI] [PubMed] [Google Scholar]
- 49.Zagai U, Dadfar E, Lundahl J, Venge P, Sköld CM. Eosinophil cationic protein stimulates TGF-beta1 release by human lung fibroblasts in vitro. Inflammation. 2007;30:153–160. doi: 10.1007/s10753-007-9032-4. [DOI] [PubMed] [Google Scholar]
- 50.Svensson L, Wennerås C. Human eosinophils selectively recognize and become activated by bacteria belonging to different taxonomic groups. Microbes Infect. 2005;7:720–728. doi: 10.1016/j.micinf.2005.01.010. [DOI] [PubMed] [Google Scholar]
- 51.Ryan P, Kelly RG, Lee G, Collins JK, O’Sullivan GC, O’Connell J, Shanahan F. Bacterial DNA within granulomas of patients with Crohn’s disease--detection by laser capture microdissection and PCR. Am J Gastroenterol. 2004;99:1539–1543. doi: 10.1111/j.1572-0241.2004.40103.x. [DOI] [PubMed] [Google Scholar]
- 52.Tedde A, Laura Putignano A, Bagnoli S, Congregati C, Milla M, Sorbi S, Genuardi M, Papi L. Interleukin-10 promoter polymorphisms influence susceptibility to ulcerative colitis in a gender-specific manner. Scand J Gastroenterol. 2008;43:712–718. doi: 10.1080/00365520701885507. [DOI] [PubMed] [Google Scholar]
- 53.Biank V, Friedrichs F, Babusukumar U, Wang T, Stoll M, Broeckel U, Kugathasan S. DLG5 R30Q variant is a female-specific protective factor in pediatric onset Crohn’s disease. Am J Gastroenterol. 2007;102:391–398. doi: 10.1111/j.1572-0241.2006.01011.x. [DOI] [PubMed] [Google Scholar]
- 54.Cho JH. Inflammatory bowel disease: genetic and epidemiologic considerations. World J Gastroenterol. 2008;14:338–347. doi: 10.3748/wjg.14.338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Desreumaux P, Brandt E, Gambiez L, Emilie D, Geboes K, Klein O, Ectors N, Cortot A, Capron M, Colombel JF. Distinct cytokine patterns in early and chronic ileal lesions of Crohn’s disease. Gastroenterology. 1997;113:118–126. doi: 10.1016/s0016-5085(97)70116-1. [DOI] [PubMed] [Google Scholar]
- 56.Bamias G, Martin C, Mishina M, Ross WG, Rivera-Nieves J, Marini M, Cominelli F. Proinflammatory effects of TH2 cytokines in a murine model of chronic small intestinal inflammation. Gastroenterology. 2005;128:654–666. doi: 10.1053/j.gastro.2004.11.053. [DOI] [PubMed] [Google Scholar]
- 57.Oppenheim JJ, Yang D. Alarmins: chemotactic activators of immune responses. Curr Opin Immunol. 2005;17:359–365. doi: 10.1016/j.coi.2005.06.002. [DOI] [PubMed] [Google Scholar]
- 58.Hart AL, Al-Hassi HO, Rigby RJ, Bell SJ, Emmanuel AV, Knight SC, Kamm MA, Stagg AJ. Characteristics of intestinal dendritic cells in inflammatory bowel diseases. Gastroenterology. 2005;129:50–65. doi: 10.1053/j.gastro.2005.05.013. [DOI] [PubMed] [Google Scholar]
- 59.Kufer TA, Sansonetti PJ. Sensing of bacteria: NOD a lonely job. Curr Opin Microbiol. 2007;10:62–69. doi: 10.1016/j.mib.2006.11.003. [DOI] [PubMed] [Google Scholar]