

Abstract
Reaping the promise of human embryonic stem (hES) cells hinges on effective defined culture conditions. Efforts to identify chemically defined environments for hES cell propagation would benefit from understanding the relevant functional properties of the substratum. Biological materials are often employed as substrata, but their complexity obscures a molecular level analysis of their relevant attributes. Because the properties of hydrogels can be tuned and altered systematically, these materials can reveal the impact of substratum features on cell fate decisions. By tailoring the peptide displayed to cells and the substrate mechanical properties, a hydrogel was generated that binds hES cell surface glycosaminoglycans (GAGs) and functions robustly in a defined culture medium to support long-term hES cell self-renewal. A key attribute of the successful GAG-binding hydrogels is their stiffness. Only stiff substrates maintain hES cell proliferation and pluripotency. These findings indicate that cells can respond to mechanical information transmitted via GAG engagement. Additionally, we found the stiff matrices afforded activation of the paralogous proteins YAP/TAZ, which are transcriptional coactivators implicated in mechanosensing and hES cell pluripotency. These results indicate that the substratum mechanics can be tuned to activate specific pathways linked to pluripotency. Because several different hES and induced pluripotent stem cell lines respond similarly, we conclude that stiff substrata are more effective for the long term propagation of human pluripotent stem cells.
Keywords: hydrogel, human embryonic stem cells, pluripotency, YAP/TAZ, substrate mechanics, tissue engineering, glycosaminoglycans
Human pluripotent stem (hPS) cells, which consist of human embryonic stem (hES) cells and induced pluripotent stem (iPS) cells, have the capacity to self-renew indefinitely and differentiate into many cell types.1-3 They can serve as a virtually unlimited supply of cells for applications ranging from drug screening to cell therapies to understanding human developmental processes. Traditional methods for propagating hES cells, however, employ matrices derived from undefined animal sources.1, 4 Such ill-defined culture conditions can be irreproducible. They also limit the therapeutic potential of hES cells due to concerns with the transmission of animal pathogens and immunogens.5, 6 These problems underlie the intense interest in developing synthetic, chemically well-defined substrates that support hPS cell self-renewal.7-11 While some effective substrata have been identified, the molecular features of substrata that give rise to pluripotency have not.
Mounting evidence indicates that multiple factors in the cellular microenvironment can act to promote a specific cell fate decision; these include cell – soluble factor, cell – cell, and cell – matrix interactions. The extracellular matrix (ECM) provides adhesive, structural, and mechanical signals to the cell.12-15 Traditional tissue culture substrates (plastic and glass) do not display defined cell-binding groups, and they have mechanical properties outside the range of many functional tissues.16, 17 In contrast, synthetic materials provide the means to tailor the presentation of recognition epitopes to the cell scaffolds for culturing hES cells have been reported,26-29 yet these depend on the presence of undefined animal-derived components in the medium. These data suggest that the hydrogels reported to date lack some characteristics needed for hES self-renewal. In addition, the presence of these animal-derived components, which can unpredictably alter even a chemically defined substratum, obscures an understanding of what parameters are critical for hES self-renewal. For example, how substratum elasticity influences hES cell propagation was unknown. Here, we describe the preparation and evaluation of a series of hydrogels that vary systematically. Our investigations reveal that fully synthetic substrata can engage GAGs to transmit mechanical signals that promote hES cell pluripotency.
Results and Discussion
Synthesis of hydrogels presenting cell-binding peptides
Polyacrylamide hydrogels have been used in many applications, and their conversion to matrices has advanced the field of tissue engineering.17 We generated polyacrylamide hydrogels16, 30 using chemoselective reactions31 to ensure that the resulting materials display peptides in a defined orientation and at controlled substitution levels (density). To this end, we immobilized the peptides using the rapid reaction of maleimides and thiolates.32. To facilitate handling and analysis of the cells, the hydrogels were covalently linked to glass coverslips functionalized with amine and aldehyde groups.16, 17, 30 (Figure 1A). Specifically, the amine-decorated coverslips were exposed to polyacrylamide hydrogels bearing electrophilic succinimidyl ester groups. Coupling occurred to immobilize the hydrogel, and its remaining activated succinimidyl esters were exposed to a mixture of glucamine and an amine containing a maleimide group. Glucamine was used as a non-binding group, as glucamine-decorated surfaces are inert to cell adhesion.33 The maleimide group served as handle for peptide attachment, and the peptides, which contain, a terminal cysteine residue, were appended by conjugate addition (Figure 1A). The ratio of glucamine and the maleimide-containing amine was varied so that the final hydrogels would have different peptide substitution levels. The efficiency of peptide immobilization was assessed by treating the hydrogels with different ratios of the non-binding glucamine and a fluorophore-labeled peptide (FITC-Acp-GRGDSC). Analysis of the hydrogel fluorescence emission indicates the expected relationship between maleimide density and peptide substitution level (Figure 1B and S1A). The fluorescent reporter also facilitated the measurement of the hydrogel thickness, and a cross-sectional image indicates that the thickness of each gel was approximately 150 μm (Figure S1B). This method for hydrogel preparation is versatile. The cysteine residue can be installed either at the N- or C- terminus of a peptide, such that the peptide is displayed in appropriate orientation for cell recognition. This variation of standard synthetic methods can be used to optimize the distribution34 and orientation35 of peptides for cell adhesion strength and specificity.
Figure 1.

Production of polyacrylamide hydrogels with controlled presentation of peptides. A. Hydrogels were appended onto functionalized glass coverslips. Functionalization of these materials was conducted to introduce a non-binding group (glucamine) and peptide sequences of interest. B. Fluorescence microscopy images of hydrogels functionalized with fluorescein-labeled peptide (FITC-Acp-GRGDSC). C. Bright field images of embryonal carcinoma cells cultured on hydrogels under serum-free conditions. Role of density in cell binding (top) and results after 3 days of growth (bottom). Scale bars: (B) 500 μm, (C) 100 μm.
Cell adhesion and viability assay
To assess whether the synthetic hydrogels sustain cell attachment and proliferation, we cultured embryonic carcinoma (EC) cells on CGRGDS-substituted materials in a serum-free medium (see Supplementary Information). EC cells are pluripotent, and their adhesion profiles are similar to those of hES cells, but they are more tolerant than hES cells to different substrata. The serum-free medium was chosen to minimize nonspecific protein adsorption, and we employed short-term (1 h) cell adhesion to diminish any effect of cell-secreted proteins. Under these defined conditions, the attachment of EC cells to the hydrogels depends on the density of immobilized peptides (Figure 1C, top panel). Attached cells remain metabolically active as determined by a cell viability assay (Figure S2). After three days of culture with EC growth medium, the cells formed characteristic tight colonies (Figure 1C, bottom panel), indicating that the synthetic hydrogels bind pluripotent cells and promote their proliferation.
Peptide-substituted hydrogels that promote hES cell self-renewal
A challenging test of the hydrogels is whether they can maintain hES cell pluripotency. We examined the utility of synthetic hydrogels presenting the integrin binding sequence RGDS peptide for culturing hES cells.36 Human ES cells were cultured on the RGDS-functionalized hydrogels with a defined medium consisting of mTeSR137 supplemented with Y-27632,38 a small molecule inhibitor of Rho-associated protein kinase or ROCK. This small molecule was included in the medium because it improves the survival of dissociated hES cells, and cells were dissociated prior to plating to ensure that our assessment of the hydrogels as substrata was stringent.8 In addition, ROCK inhibitors can facilitate hES cell culture.8 Under these defined conditions, synthetic hydrogels presenting the RGDS peptide bound hES cells. Within in 7 days, however, most of the cells failed to produce markers of pluripotency (Figure 2A and B). These observations are consistent with a report that another type of RGDS-modified hydrogel cannot maintain hES cell self-renewal beyond 5 days, even in the presence of serum proteins and mouse embryonic fibroblast-conditioned media.26
Figure 2.
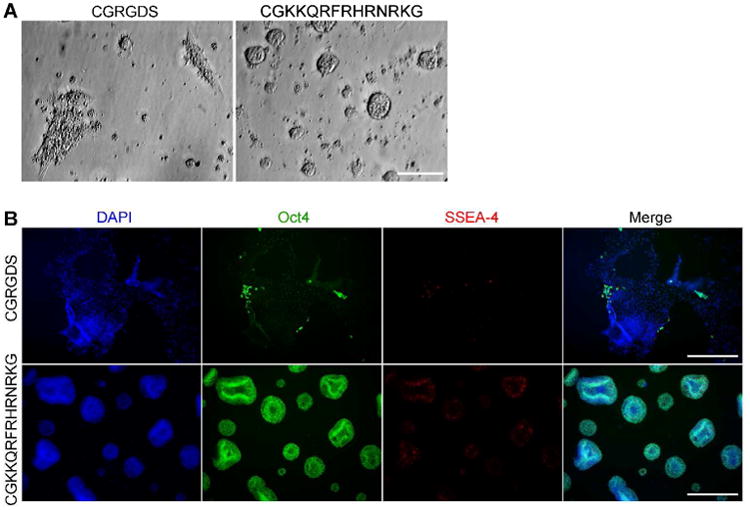
Adhesion and growth of human embryonic stem cells on defined hydrogels presenting integrin or glycosaminoglycan-binding peptides. A. Bright field images of human embryonic stem (hES) cell (H9) after 24 h culture on hydrogels functionalized with CGRGDS (integrin ligand) or CGKKQRFRHRNRKG (GAG ligand). Hydrogels were functionalized with a 1:1 ratio of aminomaleimide to glucamine. B. Immunostaining of hES cells (H9) after 7 days of culture on hydrogels presenting either the CGRGDS or CGKKQRFRHRNRKG peptide. Cells were immunostained for pluripotency markers Oct-4 (green) and SSEA-4 (red), and counterstained with DAPI (blue). Scale bars: A: 100 μm, B: 500 μm.
Another means of anchoring cells is through glycans on their surfaces. An exciting target for engaging and facilitating hES cell propagation is a class of glycans termed glycosaminoglycans (GAGs). GAGs include cell-surface polysaccharides, whose engagement has been linked to hES cell self-renewal.8 We therefore synthesized a hydrogel displaying the GAG-binding peptide GKKQRFRHRNRKG, which is derived from vitronectin.39 In contrast to what was observed with integrin-binding hydrogels, cells cultured on this substratum maintained expression of pluripotency markers Oct-4 and SSEA-4 (Figure 2A and B). In addition, the hydrogel presenting the GAG-binding peptide was more effective at supporting cell proliferation than those displaying the RGDS peptide (Figure S3). Thus, the GAG-binding hydrogel is a substrate for hES cell self-renewal under completely defined conditions. With this finding, we set out to examine whether the mechanical properties of the matrix would influence cell propagation.
Matrix mechanics impact hES cell proliferation
Having identified a hydrogel that functions in a chemically defined medium, we next examined the influence of hydrogel mechanical properties on hES cell self-renewal. Hydrogels have been used previously to show that matrix mechanics influence the cell fate decisions of specialized and progenitor cell types.17, 40-42 Substratum elasticity has also been shown to impact murine embryonic stem (mES) cell propagation; compliant surfaces were more effective than stiffer surfaces.43 Extrapolation would suggest that their human counterparts should have similar preferences for compliant matrices. With our hydrogel that serves as a robust substratum, we were poised to examine the effects of matrix mechanics on hES cell self-renewal.
Our initial experiments with hES cells were carried out using hydrogels corresponding to an elasticity of 3 kPa. To examine the role of matrix elasticity, we synthesized polyacrylamide hydrogels of various elasticities (Figure S4A) and examined their ability to support hES cell adhesion and proliferation. Hydrogels with different levels of cross-linking (Figure S4 A and B) were functionalized to display the GAG-binding peptide (Figure 2B) and used for hES cell culture in a defined medium. We observed that hES cells bound to all of the hydrogels presenting the GAG-binding peptide (Figure S4C). Over seven days of culture, however, differences in cell behavior emerged. Specifically, on a more compliant hydrogel (approximately 0.7 kPa elastic modulus, E), the bound hES cells detached during the course of the experiment. For those cells that did remain bound, colony formation was compromised. When a hydrogel of intermediate stiffness (approximately 3 kPa) was employed, most cells remained bound and colonies formed. Over the course of days, however, some cells within the colonies began to disengage (Figure 3A, white arrow). Human ES cells cultured on a stiffer hydrogel (approximately 10 kPa) attached, spread, and proliferated into robust colonies (Figure 3A, S4C and S5). These data indicate that hES cells prefer stiffer substrates.
Figure 3.
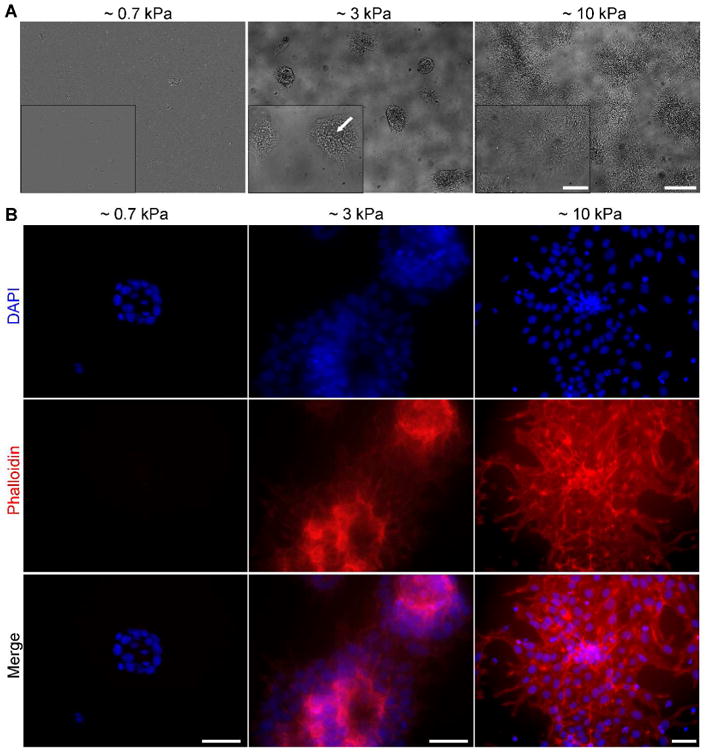
Evaluating hES cell adhesion and spreading on synthetic hydrogels of different elasticity. A. Bright field images of H9 hES cells cultured for 7 days on polyacrylamide hydrogels of the indicated elasticities. All hydrogels were functionalized with CGKKQRFRHRNRKG from a 1:1 ratio of aminomaleimide to glucamine. Arrow (inset) indicates areas where some cells have detached. B. H9 hES cells cultured for 7 days on hydrogels of variable elasticities and stained for actin filaments with Alexa Fluor 594-conjugated phalloidin (red) and counterstained with DAPI (blue). Scale bars: A: 250 μm, 100 μm for inset, B: 50 μm for all.
To examine the generality of these observations regarding substrate stiffness, we tested hydrogels coated with Matrigel. Matrigel is a complex mixture of extracellular matrix proteins, which can engage multiple cell surface targets.44 It is commonly used for hES cell culture45 and is therefore known to promote hES cell adhesion and pluripotency. If substrate stiffness is critical, hES cells should show similar preferences when presented with different cell adhesive ligands. As expected, adhesion, spreading, and growth of hES cells cultured on Matrigel-functionalized hydrogels also varied with hydrogel stiffness; The stiff materials were most effective at promoting hES cell proliferation (Figure S6). These data indicate that the response of hES cells on the different surfaces is not dependent on the affinity of the ligand for its cell surface target.
hES cell lines prefer stiff surfaces
In our initial examination of the role of substratum elasticity, we employed hES cell line H9.1 To determine whether other pluripotent cell lines respond similarly, we examined the self-renewal of a diverse set of hES cell lines on the GAG-binding hydrogels that vary in elasticity. All hES cell lines tested preferred the stiffest hydrogel. It is noteworthy that within two weeks of culture, the few hES cells that remained on the soft hydrogel undergo differentiation, as indicated by loss of Oct4 expression (data not shown). Thus the soft hydrogel (0.7 kPa) is unable to support hES cell self-renewal over prolonged culture period. The 10 kPa hydrogel was the only scaffold that consistently afforded large and well-spread colonies for all hES cell lines examined (H9, H1, H7, H14, and SA02; see Figure 3A and S5). It was effective not only for expanding hES cell lines but also for propagating diverse induced pluripotent stem cell lines (IMR-90-1, iPS-BM1i, iPS-CBT4, iPS vector free and iPS-Foreskin-1). All cell lines cultured on the stiffest hydrogels maintain the expression of pluripotency markers Oct-4 and SSEA-4 (Figure 4 and S8). These results highlight the generality of stiff hydrogels that bind GAGs.
Figure 4.

Pluripotency marker expression in hES cells grown short-term on GAG-binding stiff hydrogel. H9 hES cells were cultured for 7 days on the 10 kPa hydrogel and immunostained for pluripotency markers Oct-4 (green) and SSEA-4 (red), and counterstained with DAPI (blue). Scale bar: 100 μm.
Our observation that hES cells prefer the stiffer surfaces contrasts with the preference of murine ES cells. This difference is intriguing given that mES and hES cells represent different developmental states.46, 47 We therefore set out to explore molecular mechanisms that could shed light on the origin of these differences.
Substrate mechanics regulate cytoskeletal organization and transcriptional activation
The inability of the soft materials to support hES cell propagation could be due to insufficient cell adhesion to the matrix, enhanced cell – cell interactions that lead to colony dissociation, or changes that result from mechanosensing.48 One key cellular attribute that is altered in response to changes in substrate mechanics is the actin cytoskeleton. Actin polymerization is favored in cells that respond to stiffer matrices, and cells that adhere and spread typically exhibit higher levels of F-actin.49 After 7 days of culture on hydrogels, hES cells (H9 cell line) were stained to visualize actin filaments (F-actin). Most cells on the compliant hydrogel detached by the end of the culture period. In the cells remaining, little F-actin could be detected (Figure 3B). These results indicate that hES cells cultured on the soft hydrogel fail to assemble the cell signaling components that regulate adhesion complex formation and cell division.49, 50 Cells cultured on the stiffer hydrogels (3 or 10 kPa) exhibited higher levels of F-actin. Notably, cells plated on hydrogels of intermediate stiffness grow in tight colonies but form multilayer aggregates (as indicated by overlapping nuclei, DAPI staining, and F-actin localization). With hES cells cultured on the stiff hydrogels, there was less stacking of cells within the colonies and a more extensive network of F-actin. These observations indicate that the stiff hydrogels facilitate a robust cell -substratum interaction. Responses of this type might be expected when the substratum binds cells via integrins, but GAGs have not been implicated in this type of mechanosensing. Our data indicate that mechanical signals can also be conveyed via GAG engagement.
Our observation that stiff surfaces are superior for hES propagation is intriguing in light of emerging functional data regarding the transcriptional coactivators YAP (Yes-associated protein) and TAZ (transcriptional coactivator with PDZ-binding motif, also known as WWTR1). One set of investigations has linked these paralogous proteins to cellular mechanosensing while another suggests that they function in hES cell self-renewal.51-55 With regard to the former, experiments with mesenchymal stem cells indicate that stiff materials coated with the integrin binding protein fibronectin activate We therefore tested whether the differences in hES cell responses to matrix elasticity would be manifested in the subcellular localization of YAP/TAZ.
We predicted that only the stiff substrates would promote YAP/TAZ localization in the nucleus. We employed short-term (24 h) adhesion of individualized hES cells (Figure S7A) to prevent cell–cell interactions from influencing YAP/TAZ localization52, 57. The hES cells that attached to the most compliant substratum, the 0.7 kPa hydrogel, exhibit low levels and diffuse cytoplasmic staining of YAP/TAZ. This observation is consistent with putative degradation of the inactive (cytoplasmic) YAP/TAZ.58 In contrast, hES cells attached to the hydrogel of intermediate stiffness display higher levels of nuclear YAP/TAZ. The stiffest hydrogel, 10 kPa, was the most effective at inducing YAP/TAZ nuclear localization (Figure S7B, S7C). These data provide additional evidence of the importance of active YAP/TAZ in hES cell pluripotency. They also reaffirm our conclusion that GAG engagement can contribute to mechanosensing. Finally, they highlight the value of using synthetic materials to dissect and optimize the properties required for robust hES cell propagation.
Long-term hES cell self-renewal
To test whether the 10 kPa hydrogel displaying the GAG-binding peptide can support the long-term self-renewal of hES cells, we cultured H9 hES cells on the hydrogel scaffold for up to 60 days (12 passages). A defined medium was employed, and cells were passaged every 4-7 days onto newly synthesized hydrogels. The status of the cultured cells was assessed by profiling their expression of genes implicated in the maintenance of pluripotency. This analysis indicated that cells propagated on the hydrogel had an expression pattern similar to those cultured on Matrigel-coated plates (Figure 5A and Table S1A). Flow cytometry and immunostaining analyses revealed that the majority of cultured cells maintained high levels of pluripotency markers Oct-4 (85%), SSEA-4 (86%), and alkaline phosphatase (90%) (Figure S9A, B). Additionally, cytogenetic testing revealed that the long-term cultured cells were karyotypically normal (Figure S9C).
Figure 5.

Long-term culture of hES cells on 10 kPa hydrogels. A. Gene expression analysis of hES cells (H9) cultured for 60 days on hydrogels functionalized with CGKKQRFRHRNRKG using quantitative PCR (qPCR). The level of gene expression is compared to cells cultured on Matrigel-coated plates. B. Gene expression analysis of embryoid bodies generated from the long-term (60 days) cultured hES cells. The level of gene expression is relative to undifferentiated cells cultured on the peptide-bearing hydrogels. For both qPCR scatter plots, black lines represent a four-fold increase or decrease between control and experimental conditions. C. Microscopy images of embryoid bodies developed from long-term (38 days) cultured cells that were immunostained for markers of all three embryonic germ layers; ectoderm (nestin and β-III tubulin), mesoderm (FABP4 and α-SMA), and endoderm (AFP and Sox17). Scale bars: 100 μm for nestin, 50 μm for all others.
The pluripotency of the hES cells propagated long-term was evaluated by assaying their capacity to differentiate into derivatives of all three embryonic germ layers (ectoderm, mesoderm, and endoderm). Suspension culture to facilitate embryoid body (EB) formation induces spontaneous hES cell differentiation. Because the majority of hES cell culture substrates are susceptible to nonspecific cell adhesion, hES cells are typically transferred to a tissue culture vessel treated with poly(2-hydroxyethyl methacrylate), (poly-HEMA), which minimizes nonspecific cell adhesion. We postulated that the peptide-free polyacrylamide hydrogels should also resist protein adsorption and therefore cell adhesion (Figure 1C and S2). The non-fouling property of our hydrogels treated with glucamine should allow for EB formation. This idea was tested by removing the cells from the glycosaminoglycan-binding hydrogels, which was effected by treatment with the protease dispase; we then transferred the cells onto peptide-free hydrogels (10 kPa) for differentiation into EBs. After 14 days of culture with EB medium, the cells formed well-organized EBs, which remained in suspension (Figure S9D). The heterogeneous cell population obtained from the EBs included cells that stain positive for proteins (Figure 5C) and expressed key genes (Figure 5B and Table S1B) associated with all three embryonic germ layers.
These results highlight two features of our hydrogels. First, they are highly specific in their interactions with cells. The success of the EB formation protocol underscores the ability of the unmodified hydrogel scaffold to resist nonspecific cell adhesion. Second, the hES cells cultured over extended periods on the glycosaminoglycan-binding hydrogels differentiate into all three germ layers, a finding that underscores the utility of our materials for expanding hES cells that retain their full developmental potential.
Properties of Effective Substrates for hES Cell Propagation
Our data identify key attributes of matrices that promote hES cell self-renewal. Specifically, our investigations emphasize the utility of GAG engagement. We previously found that self-assembled monolayers that could bind cell-surface GAGs were effective for hES cell propagation.8 Here, we demonstrate the generality of GAG – matrix interactions, as we found that GAG-binding peptides can be appended to hydrogels to afford excellent hES cell substrates. Moreover, hES cells respond to the mechanical properties of GAG-binding hydrogels. Although integrin engagement is known to convey mechanical signals,13, 59-61 our results suggest that cells can sense matrix stiffness through GAG interactions. With regard to this finding, it is intriguing that proteoglycans can collaborate with integrins to activate cellular signaling pathways.62, 63 Together, our findings indicate that stiff substrata should be more effective for the propagation of hES and iPS cells.
Conclusions
In summary, we devised a synthetic hydrogel that performs in chemically defined condtions to sustain the long-term self-renewal of hES cells. Unlike most previously reported scaffolds26-29 or polymeric surfaces,64-66 cells bind to our synthetic hydrogels by virtue of the peptide epitopes displayed. The key attributes of our synthetic hydrogels are the functional epitope they display (the GAG-binding peptide) and their mechanical properties. Unlike murine ES cells, hES cells prefer a stiff hydrogel. This preference arises from the ability of the hydrogels to promote YAP/TAZ activation, which presumably upregulates genes associated with pluripotency. In this way, our studies link the external environment (i.e., the substratum) to changes in hES gene expression. We envision that the blueprint that emerges from our findings can be used to devise new materials, including three-dimensional hydrogels67, 68 for hPS cell propagation and differentiation. The development of chemically defined and tunable microenvironments could facilitate the design of synthetic nanomaterials to guide hES cell differentiation into specialized and functional cell types.
Methods
Synthesis of polyacrylamide hydrogels with controlled functionalization of peptides
Polyacrylamide hydrogels were synthesized by modification of a known protocol.16, 30 The following stock solutions and reagents were freshly prepared for hydrogel synthesis: 40% w/v acrylamide (Sigma), 2% w/v bis-acrylamide (Promega Corp.), 10% w/v ammonium persulfate (APS) from Aldrich Chemical Company Inc., saturated solution of acrylic acid N-hydroxy-succinimide ester (acrylic-NHS) from Aldrich Chemical Company Inc., and N, N, N′, N′-tetramethylethylenediamine (TEMED) from Sigma. Using these stock solutions and reagents, hydrogel precursor solutions were prepared by mixing 143 μL of H2O, 75 μL acrylamide, 60 μL bisacrylamide, 0.3 μL TEMED, 4 μL APS and 118 μL acrylic-NHS. To synthesize hydrogels with different elasticities, 10 μL, 60 μL, or 120 μL of the bisacrylamide solution corresponding to final w/v concentrations of 0.05, 0.3, and 0.6%, respectively, were brought to a final volume of 400 μL with water. The hydrogel solution was briefly vortexed and a fixed volume of 120 μL was pipetted onto the “reactive” coverslip. A siliconized coverslip was then gently placed atop of the hydrogel solution. Polymerization was allowed to proceed for approximately 10 min, and the siliconized coverslip was removed while leaving the polymerized hydrogel bound to the “reactive” coverslip. Hydrogels were transferred to 6-well plates and washed three times with phosphate buffered saline (PBS). Each wash was conducted for 5 min with rocking.
To control the density and orientation of peptide epitopes presented on the hydrogels, the activated succinimidyl ester on the polymerized gel was exposed for 2 h to 1.5 mL of a solution containing a ratio of N-methyl-D-glucamine (Aldrich) to N-(2-aminoethyl)maleimide trifluoroacetate (Fluka) (ratios included 1:0, 20:1, 4:1, 1:1, and 0:1) from 5 mM stock solutions. The pH of the mixture was adjusted to 7.5. The hydrogels were washed with PBS for 10 min (3×). A solution of 0.1 mM of the peptide of interest at pH 7.5 was added to the hydrogels, and the reaction was allowed to proceed overnight at rt. All peptides were custom synthesized by Biomatik Corp. For functionalization with Matigel, a manufacturer recommended stock solution of Matrigel (BD Biosciences) was diluted to 1:10 in PBS and reacted with the succinimidyl ester-presenting hydrogels overnight at rt.
Cell culture
Human ES and iPS cells were cultured on plates coated with Matrigel (BD Biosciences) per manufacture instructions using mTeSR1 medium (Stem Cell Technologies).37 Cells were passaged every 5-7 days by treatment with dispase (2 mg/mL). For spontaneous differentiation, cells were cultured in EB medium (described below) for at least 7 days. Human embryonic carcinoma cell line, NCCIT (ATCC) were cultured in the presence of RPMI-1640 (Gibco) supplemented with 10% heat-inactivated fetal bovine serum (FBS) from Gibco, and 1% non-essential amino acids (Gibco). NCCIT medium was refreshed every three days during passaging.
For hES and iPS cell culture on synthetic hydrogels, the hydrogels were first prepared for cell culture by sterilizing with 70% ethanol and washed with DMEM/F12 (see supplementary methods). Human hES and iPS cells at ∼70% confluency were detached from Matrigel with Hank's-based enzyme-free cell dissociation buffer (Sigma) for about 10 min at 37 °C. Cells were resuspended in mTeSR1 medium (Stem Cell Technologies) and centrifuged twice at 1200 rpm for 5 min. Cells were then resuspended in mTeSR1 medium supplemented with 5 μM ROCK inhibitor Y-27632 (Calbiochem)38 and plated on hydrogels at 15×104 cell/mL. Cells were incubated at 37 °C with 5% CO2, and the medium was refreshed daily. For long-term culture, cells were passaged onto newly synthesized hydrogels every 4-7 days at a splitting ratio of 1:4 in mTeSR1 medium supplemented with 10 μM ROCK inhibitor Y-27632. Detachment was effected using Hank's-based enzyme-free cell dissociation buffer.
Immunostaining and microscopy analysis
Cells were fixed by incubating with 4% paraformaldehyde in PBS for 20 min at rt, and permeabilized with 0.125% Triton X-100 in PBS for 5 min. Cells were blocked with a solution of 1% BSA and 0.125% Triton X-100 in PBS for 30 min at rt, then washed with permeabilization buffer (3×). Cell staining was performed with antibodies at 1:400 dilutions (1:300 for YAP antiboby) in permeabilization buffer overnight at 4 °C. Cells were exposed to secondary antibodies at 1:1000 dilutions in 0.125% Triton X-100/PBS for 1 h at rt, and then washed with permeabilization buffer (3×). The cells were counterstained with 4′, 6-diamidino-2-phenylindole (DAPI, Invitrogen) at 1:1000 dilution in H2O for 5 min at rt. The primary antibodies used include Oct-4 (R&D Systems), SSEA-4 (Santa Cruz Biotechnology), β-III tubulin (R&D Systems), nestin (R&D Systems), α-smooth muscle actin (Sigma), α-fetoprotein (Sigma), Sox17 (R&D Systems), fatty acid binding protein 4 (R&D Systems), and YAP (Sc-101199, Santa Cruz Biotechnology); Alexa Fluor 488 and Alexa Fluor 594-conjugated antibodies (Invitrogen) were used as secondary antibodies. For cytoskeletal staining, cells were incubated for 20 min with Alexa Fluor 594-conjugated phalloidin (Invitrogen) at 1:40 dilution in PBS at rt. Immunostained cells were visualized with Olympus IX81 microscope equipped with Hamamatsu digital camera. Confocal images were acquired on a Nikon Eclipse TE2000-U microscope with a Plan Apo VC x60 oil objective. Images were colored and overlayed in Adobe Photoshop CS3. Quantification of YAP/TAZ colocalization with DAPI was performed using Fiji for ImageJ.69
Gene expression analysis
Cells cultured on peptide-bearing hydrogels and Matrigel-coated plates were detached by treatment with 0.05% trypsin-EDTA (Gibco), and embryoid bodies were isolated from suspension by centrifugation. Total RNA was isolated by using RNeasy Plus Kit (Qiagen). For each sample, 725 ng of RNA was reverse transcribed by using RT2 First Strand Kit (SABiosciences). Human embryonic stem cell RT2 Profiler PCR Arrays (SABiosciences) were performed using a 7500 FAST Real-Time PCR system (Applied Biosystems Inc.) according to manufacturer protocol. Data were analyzed with the PCR array analysis web portal (pcrdataanalysis.sabiosciences.com). For clarity, genes related to signaling rather than specific lineages were omitted. The results represent the threshold cycle (Ct) for each gene normalized to the arithmetic mean of the Ct values (2-ΔCt) of five housekeeping genes (RPL13A, B2M, HPRT1, GAPDH, and ACTB). Black lines on scatter plots represent a four-fold change between control and experimental conditions. A default Ct value of 35 was assigned to genes that were detected in one sample but not the other. Genes with Ct values that are greater than 35 under both experimental conditions were considered undetected and omitted.
Supplementary Material
Acknowledgments
This research was supported by the NIH (Grant No. GM49975). S. Musah thanks the NSF Graduate Research Fellowship (ID 2007058921) and the NIH Chemical-Biology Training Grant (T32 GM008505) for fellowships. P. J. Wrighton was supported by the NIH Molecular Biosciences Training Grant (5T32 GM00721535). S. A. Morin thanks 3M for a fellowship and S. Jin acknowledges the Sloan Research Fellowship for support. We thank L. Li, J. R. Klim, A. H. Courtney, and J. M. Fishman for helpful comments, the W. M. Keck Foundation for support of the Center for Chemical Genomics, the WiCell Research Institute for providing hES and iPS cells, and the UWCCC Flow Cytometry Facility for technical assistance. We thank I. Slukvin for providing the BM1i and CBT4 iPS cell lines.
Footnotes
Supporting Information Available: Methods for coverslip preparation, AFM measurement, post-synthesis preparation of hydrogels for cell culture, cell viability and proliferation assays, flow cytometry analysis, embryoid body formation, karyotype analysis. Figures for fluorence intensity measurement, hydrogel thickness, cell viability and proliferation, and representative AFM force-indentation plot. Subcellular localization of YAP/TAZ, microscopy images for additional hES and iPS cells. Flow cytometry analysis, karayogram, EB characterization, and gene expression levels for long tem cultures. This material is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science. 1998;282:1145–1147. doi: 10.1126/science.282.5391.1145. [DOI] [PubMed] [Google Scholar]
- 2.Yu JY, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA, Ruotti V, Stewart R, et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science. 2007;318:1917–1920. doi: 10.1126/science.1151526. [DOI] [PubMed] [Google Scholar]
- 3.Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell. 2007;131:861–872. doi: 10.1016/j.cell.2007.11.019. [DOI] [PubMed] [Google Scholar]
- 4.Xu CH, Inokuma MS, Denham J, Golds K, Kundu P, Gold JD, Carpenter MK. Feeder-Free Growth of Undifferentiated Human Embryonic Stem Cells. Nat Biotechnol. 2001;19:971–974. doi: 10.1038/nbt1001-971. [DOI] [PubMed] [Google Scholar]
- 5.Hisamatsu-Sakamoto M, Sakamoto N, Rosenberg AS. Embryonic Stem Cells Cultured in Serum-Free Medium Acquire Bovine Apolipoprotein B-100 from Feeder Cell Layers and Serum Replacement Medium. Stem Cells. 2008;26:72–78. doi: 10.1634/stemcells.2007-0744. [DOI] [PubMed] [Google Scholar]
- 6.Martin MJ, Muotri A, Gage F, Varki A. Human Embryonic Stem Cells Express an Immunogenic Nonhuman Sialic Acid. Nat Med. 2005;11:228–232. doi: 10.1038/nm1181. [DOI] [PubMed] [Google Scholar]
- 7.Derda R, Musah S, Orner BP, Klim JR, Li L, Kiessling LL. High-Throughput Discovery of Synthetic Surfaces That Support Proliferation of Pluripotent Cells. J Am Chem Soc. 2010;132:1289–1295. doi: 10.1021/ja906089g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Klim JR, Li L, Wrighton PJ, Piekarczyk MS, Kiessling LL. A Defined Glycosaminoglycan-Binding Substratum for Human Pluripotent Stem Cells. Nat Methods. 2010;7:989–994. doi: 10.1038/nmeth.1532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Melkoumian Z, Weber JL, Weber DM, Fadeev AG, Zhou Y, Dolley-Sonneville P, Yang J, Qiu L, Priest CA, Shogbon C, et al. Synthetic Peptide-Acrylate Surfaces for Long-Term Self-Renewal and Cardiomyocyte Differentiation of Human Embryonic Stem Cells. Nat Biotechnol. 2010;28:606–610. doi: 10.1038/nbt.1629. [DOI] [PubMed] [Google Scholar]
- 10.Saha K, Mei Y, Reisterer CM, Pyzocha NK, Yang J, Muffat J, Davies MC, Alexander MR, Langer R, Anderson DG, et al. Surface-Engineered Substrates for Improved Human Pluripotent Stem Cell Culture under Fully Defined Conditions. Proc Natl Acad Sci USA. 2011;108:18714–18719. doi: 10.1073/pnas.1114854108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Derda R, Li LY, Orner BP, Lewis RL, Thomson JA, Kiessling LL. Defined Substrates for Human Embryonic Stem Cell Growth Identified from Surface Arrays. Acs Chemical Biology. 2007;2:347–355. doi: 10.1021/cb700032u. [DOI] [PubMed] [Google Scholar]
- 12.Lutolf MP, Hubbell JA. Synthetic Biomaterials as Instructive Extracellular Microenvironments for Morphogenesis in Tissue Engineering. Nat Biotechnol. 2005;23:47–55. doi: 10.1038/nbt1055. [DOI] [PubMed] [Google Scholar]
- 13.Discher DE, Mooney DJ, Zandstra PW. Growth Factors, Matrices, and Forces Combine and Control Stem Cells. Science. 2009;324:1673–1677. doi: 10.1126/science.1171643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vogel V, Sheetz M. Local Force and Geometry Sensing Regulate Cell Functions. Nat Rev Mol Cell Biol. 2006;7:265–275. doi: 10.1038/nrm1890. [DOI] [PubMed] [Google Scholar]
- 15.Mammoto T, Ingber DE. Mechanical Control of Tissue and Organ Development. Development. 2010;137:1407–1420. doi: 10.1242/dev.024166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yeung T, Georges PC, Flanagan LA, Marg B, Ortiz M, Funaki M, Zahir N, Ming W, Weaver V, Janmey PA. Effects of Substrate Stiffness on Cell Morphology, Cytoskeletal Structure, and Adhesion. Cell Motil Cytoskeleton. 2005;60:24–34. doi: 10.1002/cm.20041. [DOI] [PubMed] [Google Scholar]
- 17.Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell. 2006;126:677–689. doi: 10.1016/j.cell.2006.06.044. [DOI] [PubMed] [Google Scholar]
- 18.Langer R, Tirrell DA. Designing Materials for Biology and Medicine. Nature. 2004;428:487–492. doi: 10.1038/nature02388. [DOI] [PubMed] [Google Scholar]
- 19.Wichterle O, Lim D. Hydrophilic Gels for Biological Use. Nature. 1960;185:117–118. [Google Scholar]
- 20.Jia XQ, Kiick KL. Hybrid Multicomponent Hydrogels for Tissue Engineering. Macromol Biosci. 2009;9:140–156. doi: 10.1002/mabi.200800284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gauvin R, Parenteau-Bareil R, Dokmeci MR, Merryman WD, Khademhosseini A. Hydrogels and Microtechnologies for Engineering the Cellular Microenvironment. Wiley Interdisciplinary Rev-Nanomed Nanobiotech. 2012;4:235–246. doi: 10.1002/wnan.171. [DOI] [PubMed] [Google Scholar]
- 22.Cushing MC, Anseth KS. Hydrogel Cell Cultures. Science. 2007;316:1133–1134. doi: 10.1126/science.1140171. [DOI] [PubMed] [Google Scholar]
- 23.Kloxin AM, Kasko AM, Salinas CN, Anseth KS. Photodegradable Hydrogels for Dynamic Tuning of Physical and Chemical Properties. Science. 2009;324:59–63. doi: 10.1126/science.1169494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Seliktar D. Designing Cell-Compatible Hydrogels for Biomedical Applications. Science. 2012;336:1124–1128. doi: 10.1126/science.1214804. [DOI] [PubMed] [Google Scholar]
- 25.Levental I, Georges PC, Janmey PA. Soft Biological Materials and Their Impact on Cell Function. Soft Matter. 2007;3:299–306. doi: 10.1039/b610522j. [DOI] [PubMed] [Google Scholar]
- 26.Li YJ, Chung EH, Rodriguez RT, Firpo MT, Healy KE. Hydrogels as Artificial Matrices for Human Embryonic Stem Cell Self-Renewal. J Biomed Mater Res A. 2006;79:1–5. doi: 10.1002/jbm.a.30732. [DOI] [PubMed] [Google Scholar]
- 27.Gerecht S, Burdick JA, Ferreira LS, Townsend SA, Langer R, Vunjak-Novakovic G. Hyaluronic Acid Hydrogel for Controlled Self-Renewal and Differentiation of Human Embryonic Stem Cells. Proceedings of the National Academy of Sciences of the United States of America; 2007; pp. 11298–11303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Siti-Ismail N, Bishop AE, Polak JM, Mantalaris A. The Benefit of Human Embryonic Stem Cell Encapsulation for Prolonged Feeder-Free Maintenance. Biomaterials. 2008;29:3946–3952. doi: 10.1016/j.biomaterials.2008.04.027. [DOI] [PubMed] [Google Scholar]
- 29.Li Z, Leung M, Hopper R, Ellenbogen R, Zhang M. Feeder-Free Self-Renewal of Human Embryonic Stem Cells in 3d Porous Natural Polymer Scaffolds. Biomaterials. 2010;31:404–412. doi: 10.1016/j.biomaterials.2009.09.070. [DOI] [PubMed] [Google Scholar]
- 30.Klein EA, Yung Y, Castagnino P, Kothapalli D, Assoian RK. Integrins. Vol. 426. Elsevier Academic Press Inc; San Diego: 2007. Cell Adhesion, Cellular Tension, and Cell Cycle Control; pp. 155–175. [DOI] [PubMed] [Google Scholar]
- 31.Gauthier MA, Klok HA. Peptide/Protein-Polymer Conjugates: Synthetic Strategies and Design Concepts. Chem Commun (Camb) 2008:2591–2611. doi: 10.1039/b719689j. [DOI] [PubMed] [Google Scholar]
- 32.Underbakke ES, Zhu Y, Kiessling LL. Isotope-Coded Affinity Tags with Tunable Reactivities for Protein Footprinting. Angewandte Chemie International Ed In English. 2008;47:9677–9680. doi: 10.1002/anie.200803378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mrksich M. Using Self-Assembled Monolayers to Model the Extracellular Matrix. Acta Biomater. 2009;5:832–841. doi: 10.1016/j.actbio.2009.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Maheshwari G, Brown G, Lauffenburger DA, Wells A, Griffith LG. Cell Adhesion and Motility Depend on Nanoscale RGD Clustering. J Cell Sci. 2000;113:1677–1686. doi: 10.1242/jcs.113.10.1677. [DOI] [PubMed] [Google Scholar]
- 35.Behanna HA, Donners JJ, Gordon AC, Stupp SI. Coassembly of Amphiphiles with Opposite Peptide Polarities into Nanofibers. J Am Chem Soc. 2005;127:1193–1200. doi: 10.1021/ja044863u. [DOI] [PubMed] [Google Scholar]
- 36.Pytela R, Pierschbacher MD, Ruoslahti E. Identification and Isolation of a 140-Kd Cell-Surface Glycoprotein with Properties Expected of a Fibronectin Receptor. Cell. 1985;40:191–198. doi: 10.1016/0092-8674(85)90322-8. [DOI] [PubMed] [Google Scholar]
- 37.Ludwig TE, Levenstein ME, Jones JM, Berggren WT, Mitchen ER, Frane JL, Crandall LJ, Daigh CA, Conard KR, Piekarczyk MS, et al. Derivation of Human Embryonic Stem Cells in Defined Conditions. Nat Biotechnol. 2006;24:185–187. doi: 10.1038/nbt1177. [DOI] [PubMed] [Google Scholar]
- 38.Watanabe K, Ueno M, Kamiya D, Nishiyama A, Matsumura M, Wataya T, Takahashi JB, Nishikawa S, Muguruma K, Sasai Y. A Rock Inhibitor Permits Survival of Dissociated Human Embryonic Stem Cells. Nat Biotechnol. 2007;25:681–686. doi: 10.1038/nbt1310. [DOI] [PubMed] [Google Scholar]
- 39.Vogel BE, Lee SJ, Hildebrand A, Craig W, Pierschbacher MD, Wong-Staal F, Ruoslahti E. A Novel Integrin Specificity Exemplified by Binding of the Alpha V Beta 5 Integrin to the Basic Domain of the HIV Tat Protein and Vitronectin. J Cell Biol. 1993;121:461–468. doi: 10.1083/jcb.121.2.461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gilbert PM, Havenstrite KL, Magnusson KEG, Sacco A, Leonardi NA, Kraft P, Nguyen NK, Thrun S, Lutolf MP, Blau HM. Substrate Elasticity Regulates Skeletal Muscle Stem Cell Self-Renewal in Culture. Science. 2010;329:1078–1081. doi: 10.1126/science.1191035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Holst J, Watson S, Lord MS, Eamegdool SS, Bax DV, Nivison-Smith LB, Kondyurin A, Ma L, Oberhauser AF, Weiss AS, et al. Substrate Elasticity Provides Mechanical Signals for the Expansion of Hemopoietic Stem and Progenitor Cells. Nat Biotechnol. 2010;28:1123–1128. doi: 10.1038/nbt.1687. [DOI] [PubMed] [Google Scholar]
- 42.Adamo L, Naveiras O, Wenzel PL, McKinney-Freeman S, Mack PJ, Gracia-Sancho J, Suchy-Dicey A, Yoshimoto M, Lensch MW, Yoder MC, et al. Biomechanical Forces Promote Embryonic Haematopoiesis. Nature. 2009;459:1131–1135. doi: 10.1038/nature08073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chowdhury F, Li Y, Poh YC, Yokohama-Tamaki T, Wang N, Tanaka TS. Soft Substrates Promote Homogeneous Self-Renewal of Embryonic Stem Cells Via Downregulating Cell-Matrix Tractions. Plos One. 2010;5:e15655. doi: 10.1371/journal.pone.0015655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kleinman HK, Martin GR. Matrigel: Basement Membrane Matrix with Biological Activity. Semin Cancer Biol. 2005;15:378–386. doi: 10.1016/j.semcancer.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 45.Xu C, Inokuma MS, Denham J, Golds K, Kundu P, Gold JD, Carpenter MK. Feeder-Free Growth of Undifferentiated Human Embryonic Stem Cells. Nat Biotechnol. 2001;19:971–974. doi: 10.1038/nbt1001-971. [DOI] [PubMed] [Google Scholar]
- 46.Nichols J, Smith A. Naive and Primed Pluripotent States. Cell Stem Cell. 2009;4:487–492. doi: 10.1016/j.stem.2009.05.015. [DOI] [PubMed] [Google Scholar]
- 47.Chou YF, Chen HH, Eijpe M, Yabuuchi A, Chenoweth JG, Tesar P, Lu J, McKay RDG, Geijsen N. The Growth Factor Environment Defines Distinct Pluripotent Ground States in Novel Blastocyst-Derived Stem Cells. Cell. 2008;135:449–461. doi: 10.1016/j.cell.2008.08.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chowdhury F, Na S, Li D, Poh YC, Tanaka TS, Wang F, Wang N. Material Properties of the Cell Dictate Stress-Induced Spreading and Differentiation in Embryonic Stem Cells. Nat Mater. 2010;9:82–88. doi: 10.1038/nmat2563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chamaraux F, Fache S, Bruckert F, Fourcade B. Kinetics of Cell Spreading. Phys Rev Lett. 2005;94:158102. doi: 10.1103/PhysRevLett.94.158102. [DOI] [PubMed] [Google Scholar]
- 50.Rao JY, Hurst RE, Bales WD, Jones PL, Bass RA, Archer LT, Bell PB, Hemstreet GP., III Cellular F-Actin Levels as a Marker for Cellular Transformation: Relationship to Cell Division and Differentiation. Cancer Res. 1990;50:2215–2220. [PubMed] [Google Scholar]
- 51.Lian I, Kim J, Okazawa H, Zhao J, Zhao B, Yu J, Chinnaiyan A, Israel MA, Goldstein LSB, Abujarour R, et al. The Role of Yap Transcription Coactivator in Regulating Stem Cell Self-Renewal and Differentiation. Genes Dev. 2010;24:1106–1118. doi: 10.1101/gad.1903310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhao B, Tumaneng K, Guan KL. The Hippo Pathway in Organ Size Control, Tissue Regeneration and Stem Cell Self-Renewal. Nat Cell Biol. 2011;13:877–883. doi: 10.1038/ncb2303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ramos A, Camargo FD. The Hippo Signaling Pathway and Stem Cell Biology. Trends Cell Biol. 2012;22:339–346. doi: 10.1016/j.tcb.2012.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Varelas X, Sakuma R, Samavarchi-Tehrani P, Peerani R, Rao BM, Dembowy J, Yaffe MB, Zandstra PW, Wrana JL. Taz Controls Smad Nucleocytoplasmic Shuttling and Regulates Human Embryonic Stem-Cell Self-Renewal. Nat Cell Biol. 2008;10:837–848. doi: 10.1038/ncb1748. [DOI] [PubMed] [Google Scholar]
- 55.Mendez MG, Janmey PA. Transcription Factor Regulation by Mechanical Stress. Int J Biochem Cell Biol. 2012;44:728–732. doi: 10.1016/j.biocel.2012.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Dupont S, Morsut L, Aragona M, Enzo E, Giulitti S, Cordenonsi M, Zanconato F, Le Digabel J, Forcato M, Bicciato, et al. Role of Yap/Taz in Mechanotransduction. Nature. 2011;474:179–183. doi: 10.1038/nature10137. [DOI] [PubMed] [Google Scholar]
- 57.Hong W, Guan KL. The Yap and Taz Transcription Co-Activators: Key Downstream Effectors of the Mammalian Hippo Pathway. Sem Cell Dev Biol. 2012 doi: 10.1016/j.semcdb.2012.05.004. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zhao B, Li L, Tumaneng K, Wang CY, Guan KL. A Coordinated Phosphorylation by Lats and Ck1 Regulates Yap Stability through Scfβ-Trcp. Genes Dev. 2010;24:72–85. doi: 10.1101/gad.1843810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hynes RO. The Extracellular Matrix: Not Just Pretty Fibrils. Science. 2009;326:1216–1219. doi: 10.1126/science.1176009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Giancotti FG, Ruoslahti E. Integrin Signaling. Science. 1999;285:1028–1033. doi: 10.1126/science.285.5430.1028. [DOI] [PubMed] [Google Scholar]
- 61.Wang N, Tytell JD, Ingber DE. Mechanotransduction at a Distance: Mechanically Coupling the Extracellular Matrix with the Nucleus. Nat Rev Mol Cell Biol. 2009;10:75–82. doi: 10.1038/nrm2594. [DOI] [PubMed] [Google Scholar]
- 62.Couchman JR. Transmembrane Signaling Proteoglycans. Annu Rev Cell Dev Biol. 2010;26:89–114. doi: 10.1146/annurev-cellbio-100109-104126. [DOI] [PubMed] [Google Scholar]
- 63.Morgan MR, Humphries MJ, Bass MD. Synergistic Control of Cell Adhesion by Integrins and Syndecans. Nat Rev Mol Cell Biol. 2007;8:957–969. doi: 10.1038/nrm2289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mei Y, Saha K, Bogatyrev SR, Yang J, Hook AL, Kalcioglu ZI, Cho SW, Mitalipova M, Pyzocha N, Rojas F, et al. Combinatorial Development of Biomaterials for Clonal Growth of Human Pluripotent Stem Cells. Nat Mater. 2010;9:768–778. doi: 10.1038/nmat2812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Villa-Diaz LG, Nandivada H, Ding J, Nogueira-de-Souza NC, Krebsbach PH, O'Shea KS, Lahann J, Smith GD. Synthetic Polymer Coatings for Long-Term Growth of Human Embryonic Stem Cells. Nat Biotechnol. 2010;28:581–583. doi: 10.1038/nbt.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Irwin EF, Gupta R, Dashti DC, Healy KE. Engineered Polymer-Media Interfaces for the Long-Term Self-Renewal of Human Embryonic Stem Cells. Biomaterials. 2011;32:6912–6919. doi: 10.1016/j.biomaterials.2011.05.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kraehenbuehl TP, Langer R, Ferreira LS. Three-Dimensional Biomaterials for the Study of Human Pluripotent Stem Cells. Nat Meth. 2011;8:731–736. doi: 10.1038/nmeth.1671. [DOI] [PubMed] [Google Scholar]
- 68.Huh D, Hamilton GA, Ingber DE. From 3D Cell Culture to Organs-on-Chips. Trends Cell Biol. 2011;21:745–754. doi: 10.1016/j.tcb.2011.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, et al. An Open-Source Platform for Biological-Image Analysis. Nat Meth. 2012;9:676–682. doi: 10.1038/nmeth.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.