

Summary
Dorsal closure (DC) is an essential step during Drosophila development whereby a hole is sealed in the dorsal epidermis and serves as a model for cell sheet morphogenesis and wound healing. It involves the orchestrated interplay of transcriptional networks and dynamic regulation of cell machinery to bring about shape changes, mechanical forces, and emergent properties. Here we provide insight into the regulation of dorsal closure by describing novel autonomous and non-autonomous roles for U-shaped (Ush) in the amnioserosa, the epidermis, and in mediation of communication between the tissues. We identified Ush by gene expression microarray analysis of Dpp signaling targets and show that Ush mediates some DC functions of Dpp. By selectively restoring Ush function in either the AS or the epidermis in ush mutants, we show that the AS makes a greater (Ush-dependent) contribution to closure than the epidermis. A signal from the AS induces epidermal cell elongation and JNK activation in the DME, while cable formation requires Ush on both sides of the leading edge, i.e. in both the AS and epidermis. Our study demonstrates that the amnioserosa and epidermis communicate at several steps during the process: sometimes the epidermis instructs the amnioserosa, other times the AS instructs the epidermis, and still other times they appear to collaborate.
Keywords: JNK, Amnioserosa, Dorsal closure, Epidermis, U-shaped
Introduction
Morphogenesis, the self-assembly of cells into the spatial patterns that underpin tissues and organs, is a key feature of embryonic development. While the generation of different cell types relies on patterns of gene activity, morphogenesis uses the activities of the cytoskeleton and cell adhesion to power a gamut of behaviors of cellular ensembles. Spreading and fusion of epithelial cell sheets is one of these processes that serves to form various structures across phylogeny, e.g. during Drosophila dorsal closure (Martinez Arias, 1993; Jacinto et al., 2002; Gorfinkiel et al., 2011), C. elegans ventral enclosure (Williams-Masson et al., 1997), Zebrafish epibody (Keller and Trinkaus, 1987), vertebrate neurulation, palate and eyelid fusion (Taya et al., 1999; Colas and Schoenwolf, 2001), as well as playing a central role during epidermal wound healing (Martin and Parkhurst, 2004). Understanding the mechanics and molecular mechanisms of these events can, therefore, provide important insights into the emergence of form during organogenesis and repair.
The process of dorsal closure (DC) in Drosophila represents a good model system for the study of epithelial sheet morphogenesis. Half way through embryogenesis, the dorsal epidermis of Drosophila exhibits a discontinuity that is covered by the amnioserosa (AS), a transient epithelium which will not contribute to the larva. DC involves dorsalward extension of the lateral epidermal sheets and coordinated contraction of the AS cells to close the discontinuity and fuse the two epidermal flanks, thus forming a continuous epidermis. Over the last few years much has been learnt about the contributions of each of the epithelial sheets to DC. The pulsed contraction of the AS cells has been revealed as an essential driving force for the process while the lateral epidermis constrains closure (Kiehart et al., 2000; Hutson et al., 2003; Franke et al., 2005; Scuderi and Letsou, 2005; Fernandez et al., 2007; Gorfinkiel et al., 2009; Solon et al., 2009; Blanchard et al., 2010; David et al., 2010). The dorsal-most epidermal (DME) cells located at the interface of the epidermis and the AS, make a distinctive contribution to DC. First, they assemble a supracellular actomyosin cable, spanning the length of the epidermal leading edge, whose purse-string activity also contributes to closure (Hutson et al., 2003; Peralta et al., 2007). Dynamic actin-rich filopodia are projected from the leading edge (LE) and at the final stage of closure they interdigitate with filopodia from the opposing epidermal front, actively zipping the epidermal sheets together (Jacinto et al., 2000; Hutson et al., 2003; Peralta et al., 2007; Millard and Martin, 2008). Thus DC results from the joint contribution of AS and epidermis-specific forces, spanning multi-cellular distances, and several studies have suggested mechanical and chemical interactions between the two tissues (Kiehart et al., 2000; Reed et al., 2001; Stronach and Perrimon, 2002; Scuderi and Letsou, 2005; Fernandez et al., 2007; Gorfinkiel and Martínez Arias, 2007; Wada et al., 2007; Zahedi et al., 2008; Gorfinkiel et al., 2009; Solon et al., 2009). However, the control and coordination of these interactions has not been explored.
Dorsal closure commences at stage 13 of embryogenesis after germband retraction. Its onset is marked by two events in the DME cells: activation of JNK and dorsoventral elongation; however the initiation trigger remains uncharacterized (Knust, 1996; Sluss et al., 1996; Glise and Noselli, 1997; Kockel et al., 1997; Zeitlinger et al., 1997; Kaltschmidt et al., 2002; Jankovics and Brunner, 2006; Bates et al., 2008; VanHook and Letsou, 2008). JNK signaling induces expression of target genes puc and dpp. Dpp, a ligand of the TGF-β superfamily, is then secreted from the DME to promote autocrine and paracrine signaling with essential roles in morphogenesis of both epidermis and amnioserosa (Hou et al., 1997; Riesgo-Escovar and Hafen, 1997; Sluss and Davis, 1997; Su et al., 1998; Ricos et al., 1999; Arquier et al., 2001; Chen et al., 2002; Fernandez et al., 2007; Wada et al., 2007). The latter is particularly of interest since it represents a form of communication between the tissues: a chemical message (Dpp) is released from the epidermis and elicits contraction of the adjacent amnioserosa, in a parallel manner to paracrine TGF-β-induced wound bed contraction in mammals (Martin et al., 1993; Fernandez et al., 2007; Zahedi et al., 2008). Dpp has been shown to affect different cellular activities but little is known about its effectors.
Here we provide insight into the regulation of the process of DC by describing novel autonomous and non-autonomous roles for the Dpp target U-shaped (Ush) in the amnioserosa, the epidermis, and in mediation of communication between the tissues.
Results and Discussion
The Dpp target Ush mediates dorsal closure functions of Dpp signaling
It has been suggested that Dpp is an important regulator of dorsal closure and other epithelial sheet movements (Affolter et al., 1994; Riesgo-Escovar and Hafen, 1997; Ricos et al., 1999; Harden, 2002; Beanes et al., 2003), and as a ligand secreted at the interface between the AS and epidermis it may travel to adjacent tissues and serve as a means of communication between them (Affolter and Basler, 2007; Fernandez et al., 2007; Zahedi et al., 2008). However its effectors are largely unknown and for this reason we carried out gene expression microarray analysis to identify targets of Dpp signaling during DC (K. Lada and A. Martinez Arias, unpublished). The expression of the transcription factor U-shaped (Ush), was robustly and reproducibly downregulated in loss of thickveins (tkv, the Dpp receptor) function and upregulated in gain of function conditions (Fernandez et al., 2007) and this work) suggesting that it is a bona fide target of Dpp signaling. Embryos mutant for ush exhibit defects in germband retraction (GBR) (Frank and Rushlow, 1996; Goldman-Levi et al., 1996), and cuticle preparations display a hole that spans the juxtaposed head and tail (Frank and Rushlow, 1996) (Fig. 1D) implying failure in both GBR and DC. We therefore decided to characterize the role of Ush in DC in an attempt to better understand the process.
Fig. 1. Dpp mediates some Dpp functions during Dorsal Closure.

(A,B) Wild type (A) and tkv8 (B) stage 14 embryos stained for Ush. (C) pAS/cAS ratio of Ush expression in wild type and tkv8 embryos. The difference in Ush expression between pAS and cAS is abolished in tkv8 embryos. (D–G) Phenotype of the cuticles layed by ush2 (D), daGAL4/UAS-ush (E, control cuticle), tkv8 (F), and tkv8, daGAL4/UAS-ush (G) embryos.
Immunostaining of stage 14 embryos with an antibody against Ush shows that the expression of Ush in the AS and dorsal epidermis is graded (Fig. 1A) as has previously been observed (Fernandez et al., 2007). Quantification of fluorescence intensity levels from confocal images (see Materials and Methods) reveals significant differences in Ush expression between the peripheral AS (pAS) cells (those that abut the DME) and the central AS (cAS) cells (those that do not contact the DME) (Fig. 1C). Indeed in wild-type embryos pAS cells expressed Ush 2.12 times more strongly than cAS cells (Fig. 1C, P ≈ 0.0003; 74 pAS and 80 cAS cells from 6 embryos). These differences correlate with variation in the behavior of these cells as shown before (Fernandez et al., 2007; Wada et al., 2007). The gradient-like patterns of Ush expression in the AS and the epidermis, highest close to the leading edge, are suggestive of regulation by the morphogen concentration gradient of Dpp. We confirmed that Ush is expressed in response to Dpp signaling, as in mutants in the Dpp receptor Thickveins (tkv8) the expression of Ush is significantly reduced in the epidermis (Fig. 1B, P ≈ 0.002; n = 4 embryos) and in the amnioserosa (P ≈ 0.016; 125 cells from 4 embryos). Furthermore, when we analyzed different regions of the AS from tkv8 mutants independently, we found that pAS cells were more affected than the cAS ones, i.e. Ush expression in pAS cells was reduced by half compared to wild-type (P ≈ 0.008; 34 cells from 4 embryos) while it was only 20% reduced in the cAS compared to wild-type (P ≈ 0.19; 91 cells from 4 embryos). Thus the gradient that normally exists was abolished, and the ratio of Ush expression between pAS and cAS cells evened out to 1.06 (86 cells from 3 embryos) from 1.67 in control siblings (114 cells from 3 embryos) (P ≈ 0.00016) (Fig. 1C). Since the epidermal expression of Ush also becomes homogenous in tkv8 mutants by virtue of being undetectable (Fig. 1B), it seems that the morphogen properties of Dpp are responsible for the graded expression of Ush in both tissues.
As ush appears to be activated by Dpp signaling during closure we also wanted to know whether it mediates some of the functions of Dpp during DC. We sought to do this by restoring Ush expression in tkv mutants using the ubiquitous driver daGal4. Mutants for tkv8 are 100% lethal and all of the cuticles exhibit a large dorsal hole, indicative of failure in DC (Fig. 1F; Table 1) (Fernandez et al., 2007). Compared to control cuticles (Fig. 1E), a range of rescue cuticular phenotypes were observed in the progeny of tkv8/CTG; da-GAL4/TM6B females crossed to tkv8 UAS-ush/CTG males, from smaller dorsal holes to complete closure of the dorsal epidermis (Fig. 1G; Table 2). Thus it appears that Ush may play an important role in mediating some of the function of Dpp during DC.
Table 1. Penetrance of cuticle defects seen in tkv8 lethal mutant embryos with respect to germband retraction (GBR) and dorsal closure (DC). n = 88.

Table 2. Ush rescues the dorsal closure defects of tkv8 mutants.
Range of cuticle defects observed in the progeny of tkv8/CTG; da-GAL4/TM6B females crossed to tkv8 UAS-ush/CTG; + males. The percentage of observed phenotypes is indicated as well as the percentage of expected phenotypes if Ush was rescuing the dorsal closure defects of tkv8 mutant embryos.

Ush is required for morphogenesis of both amnioserosa and epidermis during dorsal closure
Embryos mutant for the ush2 null allele exhibit a large proportion of dorsal holes (86.7%, n = 39; Table 3). Immunodetection of F-actin in DC-stage of these mutants revealed that the amnioserosa and epidermis are severely impaired. The AS is prematurely degraded by stage 13 (Frank and Rushlow, 1996), no actin cable is assembled at the leading edge and epidermal cells do not elongate dorso-ventrally but some of them are stretched antero-posteriorly (Fig. 3B,B′, compare with wild type stage 14 embryos in Fig. 3A). The severe disorganization of both tissues in ush2 mutants points to important cellular functions mediated by Ush. However the nature of these functions, whether autonomous or non-autonomous, remained to be addressed.
Table 3. Penetrance of cuticle defects seen in ush2 mutants embryos and in the progeny from rescue experiments (genotypes indicated in the table) with respect to germband retraction (GBR) and dorsal closure (DC).
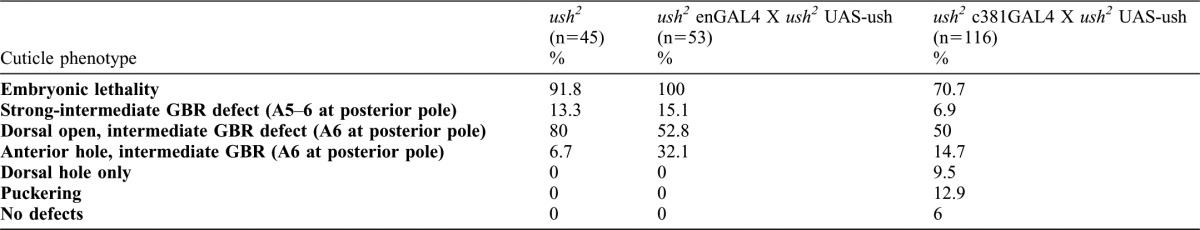
Fig. 3. An Ush-mediated signal from the AS induces elongation and D-V microtubule realignement of epidermal cells.

(A–D) Stage 14 embryos stained for F-actin. (E-G) Early stage 15 (E) and stage 14 (F,G) stained for α-tubulin. (A′–D′) Magnification of boxed region made by camera lucida tracing. (A) Wild type embryo. Epidermal cells are elongated in the D-V direction. (A′) DME cell are colored green, lateral epidermal cells blue. (B) ush2 embryos. Epidermal cells fail to elongate. (B′) DME and second row of epidermal cells are often stretched in the A-P direction while the bulk of epidermal cells tend to be isotropic. (C) Expression of Ush in epidermal stripes of ush2 mutants does not affect the shape of epidermal cells. (C′) Magnified region has been selected to cover a boundary between Ush-expressing and mutant epidermal cells and shows that the two first rows of epidermal cells still stretch in the A-P direction. (D) Expression of Ush in the amnioserosa of ush2 mutants rescues the elongation of epidermal cells. (D′) DME and lateral epidermal cells clearly elongate in the D-V direction. (E) Epidermal cell elongation is associated with formation of apical microtubule bundle that are oriented in the D-V direction, as shown here in wild type embryos. Right panel: magnification of the boxed region in (E). (F) Microtubules fail to form bundles and to orient in ush2 mutant embryos, Note their disorganized pattern in the corresponding magnification. (G) Restoring Ush in the amnioserosa of ush2 mutant embryos elicits correct bundling and D-V alignment of microtubules in the epidermis. All embryos are oriented with anterior to the left except embryo in G whose anterior end faces top left. Camera lucida drawings are 2× magnifications and the tubulin images are 2.5× magnifications of the corresponding boxed regions. All magnifications are as in Fig. 2 except for the insets in E, F and G, which are taken from the corresponding panels.
In order to analyze the cell autonomy of Ush function, we made use of the GAL4 UAS system (Brand and Perrimon, 1993) to create stripes of Ush expression in both the AS and epidermis of ush2 mutants. The prdGal4 driver expresses UASush in regularly arranged stripes, about four cells apart in the AS and about ten cells apart in the epidermis of stage 14 wild-type embryos (Fig. 2A). However, in ush2; prdGal4 > UASush stage 14 embryos the region in between expressing stripes is obliterated in the AS. It appears that ush2 mutant AS cells are eliminated from the tissue while the rescued Ush-expressing cells join together to maintain a continuous epithelium (Fig. 2B,B′). This observation suggests a cell autonomous role for Ush in maintaining AS integrity. In these embryos, the dorsal-most rows of epidermal cells ceased to be stretched in the A-P direction (Fig. 2B′). The rescue of this phenotype may be attributed to the presence of an intact, though smaller, AS which holds the embryo together and withstands the tension of other pulling events in the anterior (e.g. head involution) and posterior regions of the embryo. The actin cable could also potentially contribute to preventing epidermal cells from being stretched anteroposteriorly, and we observed that some regions of the LE regained actin cable and filopodia (Fig. 2B,B′). Despite rescue of the aforementioned features in ush2; prdGal4 > UASush embryos, another phenotype developed: the epidermis began to bunch at the LE in regions coinciding with mutant stripes (Fig. 2B′, arrowhead). Epidermal bunching at the LE could be an autonomous phentoype or it could be due to loss of adjacent AS cells. It was, however, possible to find mosaic mutant embryos that closed (Fig. 2C,D,D′), demonstrating the robustness of the system.
Fig. 2. Mosaics of ush in the AS and the epidermis.

(A) prd-GAL4/UAS-ush/UAS-GFP embryos stained for F-actin. GFP nuclei label the expression domain of the prd-GAL4 driver and thus the domain of ectopic Ush expression. (B–B′) ush2, prd-GAL4/UAS-ush embryos labelled for F-actin (B′). Ush is responsible for maintaining AS cell integrity. (C) ush2, prd-GAL4/UAS-ush embryos close. (D–D′) ush2, prd-GAL4 c381-GAL4/UAS-ush/UAS-GFP embryos stained for F-actin. Ush is responsible for maintaining dorsal epidermis organization. ush2 mutant stripes are bunched at the leading edge. (E–E″) ush2, prd-GAL4 c381-GAL4/UAS-ush/UAS-GFP embryos stained for F-actin. There is communication between the AS and the epidermis for LE cable formation. The cable only forms when Ush-expressing cells are juxtaposed to Ush-expressing epidermal cells. Scale bars are 20μm; the scale bar in B also applies to A, C and E.
To test whether a defective AS was responsible for the bunching at the leading edge in ush2; prdGal4 > UASush embryos, we co-expressed prdGal4 with c381Gal4 in ush2 mutants to create embryos with homogeneous Ush expression in the AS and striped expression in the epidermis. In these embryos, the epidermis continued to bunch in regions lacking Ush (Fig. 2D,D′) despite rescue of the entire AS, suggesting that it is an autonomous response of the epidermis to the lack of ush. Interestingly, we noted that these embryos formed a discontinuous actin cable and that patches of cable coincided with rescued stripes, i.e. actin accumulated at the leading edge where Ush-expressing AS cells were juxtaposed to Ush-expressing epidermal cells (Fig. 2E,E′). The actin cable is associated with actin-nucleating centers, localized to adherens junctions, which exhibit phosphorylation of tyrosine (Kaltschmidt et al., 2002). We therefore examined phosphotyrosine (pTyr) levels and noticed that accordingly higher levels of pTyr can be seen in Ush-expressing areas of the LE whilst they are absent from the adjacent ush mutant areas (Fig. 2E″). The actin cable did not assemble in ush2 mutants in which Ush was expressed only in the AS (data not shown) nor only in the epidermis (en stripes, Fig. 3C), however it always formed when Ush is expressed in both the DME and in adjacent AS cells. This leads us to conclude that the AS and epidermis cooperate with each other to form the cable and that Ush plays an important role in this interaction.
A signal from the AS mediated by Ush induces epidermal cell elongation
To determine the possibility of further reciprocal influences of AS and epidermis on each other's behavior during DC, we analyzed DC in ush2 embryos with rescue only in the epidermis or the AS. Mutant embryos with Ush function restored in epidermal stripes exhibited a partial rescue of closure, i.e. a higher proportion of smaller dorsal holes were seen in cuticles (Table 3). However, the AS is still degraded and there is no difference in the cell morphology or LE cable formation between the rescued and mutant epidermal cells (Fig. 3B–C′, compare to wild type embryos in Figure 3A,A′). In contrast, when Ush is expressed only in the AS of ush2 mutants, we observe a wider range of phenotypes in cuticles, including complete rescue of GBR and DC, and a 30% increase in embryonic viability (Table 3). Analysis of these embryos at the cellular level revealed that restoring expression of Ush in the AS completely rescued epidermal cell elongation in ush2 mutants (Fig. 3D,D′, compare to wild type embryos in Fig. 3A,A′). Since epidermal cell elongation has been associated with microtubule realignment (Fig. 3E) (Kaltschmidt et al., 2002; Jankovics and Brunner, 2006), we analyzed the organization of α-tubulin and found that parallel bundles of microtubules formed apically and aligned correctly in the D-V direction throughout the epidermis of ush2; c381Gal4 > UASush embryos, which did not occur in ush2 mutants (Fig. 3E–G). As indicated above, the actin cable was not rescued (Fig. 3C–D) if Ush function was restored only in the epidermis or the AS, which supports our earlier findings that Ush is required on both sides of the leading edge for cable formation. These observations also show that the purse-string is not responsible for epidermal cell elongation, as has been suggested before (Knust, 1996; Harden et al., 2002), since all epidermal cells elongated in the absence of a complete cable. Thus, it is plausible that the elongation of epidermal cells is a response to the activity of the AS, and more precisely, to the pulling force exerted by the tissue. In agreement with this, we have observed that upon detachment of the AS from the epidermis, the elongation of epidermal cells is lost (N.G and A.M.A, unpublished results). Taken together, these observations provide direct evidence for the suggestion that the AS can influence the activity of the epidermis (Reed et al., 2001; Scuderi and Letsou, 2005) and also identify Ush as a major mediator of this interaction from the AS.
Activation of JNK in the DME is activated via Ush in the AS
One possible way the AS can influence the activity of the epidermis is through the contractile force generated by its constituent cells. There is strong evidence that the AS pulls on the epidermis since GBR and during DC (Kiehart et al., 2000; Harden et al., 2002; Schock and Perrimon, 2002; Hutson et al., 2003; Gorfinkiel et al., 2009; Solon et al., 2009). We thus considered whether the mechanical stress triggered by AS contraction may impinge on the epidermis through the regulation of JNK at the DME, since mechanical stress-induced JNK activation occurs in other systems, such as wound healing (Tran et al., 1999; Klepeis et al., 2001), endothelial shear stress (Azuma et al., 2000), and bladder smooth muscle stretching (Kushida et al., 2001). Previous work has referred to an unknown “signal X” which has represented the trigger for both epidermal elongation and for JNK activation in the DME (Stronach and Perrimon, 2001; Kaltschmidt et al., 2002; Bates et al., 2008; VanHook and Letsou, 2008). In light of these considerations and of our results, we thus explored whether signals from the AS could also be coupled with JNK activity in the DME.
The expression of puc in the DME is a readout of JNK activity (Glise and Noselli, 1997; Hou et al., 1997; Riesgo-Escovar and Hafen, 1997; Sluss and Davis, 1997; Zeitlinger et al., 1997; Martin-Blanco et al., 1998; Reed et al., 2001). For this reason we used the pucE69 enhancer trap that carries a LacZ reporter in the puc gene and is a reliable marker of JNK signaling (Ring and Martinez Arias, 1993; Martin-Blanco et al., 1998) that expresses β-gal cleanly in the dorsal-most epidermal cells during DC (Fig. 4A,A′). Expression of puclacZ is abolished in all DME cells of ush2 mutants but for one or two which are found at the bend of the germband (and serve as a convenient indicator of the presence of the transgenic construct) (Fig. 4B,B′). Interestingly, when we rescue ush expression in the AS of ush2 mutants, we observe that the expression of puc (β-gal) in restored in most DME cells (Fig. 4C–D′). This is an intriguing result particularly since previous work suggested JNK activation is independent of the AS, showing dpp enhancer trap expression in embryos with a toxin-ablated AS (Scuderi and Letsou, 2005). Our results may not be mutually exclusive, however, as Scuderi and Letsou (Scuderi and Letsou, 2005) tested dpp in earlier (pre-DC) stage 11–12 embryos while our work examined stage 14. Fernandez et al. (Fernandez et al., 2007) suggested that two rounds of Dpp signaling affect DC: the first, JNK-independent (pre-stage 13), is responsible for AS contractile behavior; and the second, JNK-dependent (starting stage 13) is necessary for epidermal morphogenesis. Our data is consistent with this idea and suggests that AS activity (perhaps elicited by the earlier round of Dpp signaling) activates JNK in the DME from stage 13.
Fig. 4. Non-autonomous rescue of puc expression in DME cells.
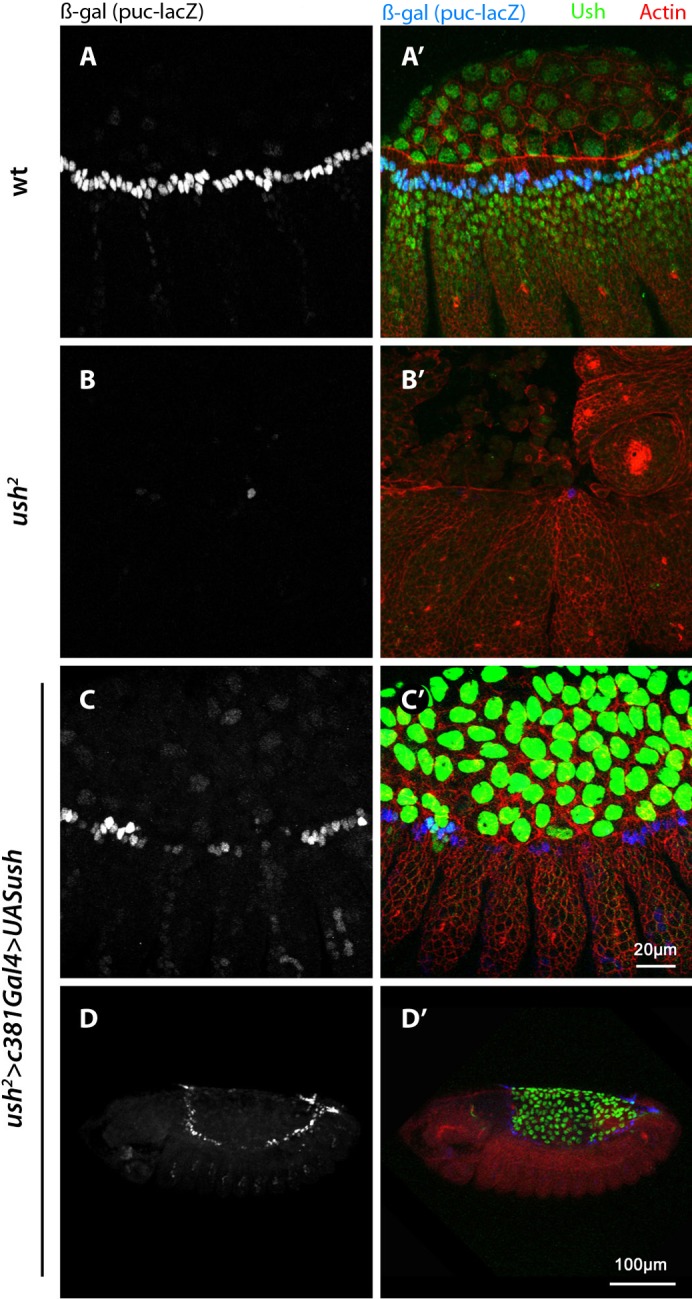
(A–D′) Stage 14 embryos stained for F-actin (red), Ush (green) and β-gal (blue). (A–A′) Wild type expression of the puc enhancer trap in the dorsal most row of epidermal cells. (B–B′) Puc reporter expression is abolished in ush2 mutant embryos. Here, only one DME cell expresses β-gal. In this region of ush2 mutant embryos remnant AS cells persist the longest before they degrade. (C–C′) β-gal expression is restored in DME of ush2 mutant embryos where Ush function in the AS has been restored. Detection is only weaker in about 10 cells around the central region. (D–D′) Lower magnification of C shows β-gal is expressed along the entire leading edge. The scale bar in C applies to A and B.
Conclusion
Our work provides direct evidence for crosstalk between the AS and the epidermis during DC. Dpp secreted from the DME activates Ush in both AS and epidermis. Mosaic rescue analysis of ush mutants suggests distinct autonomous and non-autonomous roles for Ush in both AS and epidermis, summarized in the model in Fig. 5A–C. Autonomous functions involve the maintenance of AS integrity and epidermal organization, including the capacity for DME cells to assemble the LE cable. Non-autonomous functions are manifest by a normal AS being able to pull and close a ush mutant epidermis. One interesting observation of these experiments is the demonstration that a functional AS can promote cell shape changes and coordinated behavior in the otherwise mutant epidermis. Activation of JNK in the DME cells in these circumstances suggests that this might be a mechanical response of the epidermis.
Fig. 5. Distinct roles of Ush in the amnioserosa and the epidermis.

Ush regulates autonomously and non-autonomously multiple morphogenetic properties during dorsal closure. (A) Most of Ush activity in the AS observed in this work occurs prior to or at the initiation of DC. (B) Ush activity in the epidermis is manifested during the later phases of DC. (C) Ush function in the AS and the epidermis act together to allow for the accumulation of actin at the leading edge.
While there have been reports of interactions between the epidermis and the AS (Lamka and Lipshitz, 1999; Stronach and Perrimon, 2001) and also suggestions that the AS must act on the epidermis (Kiehart et al., 2000; Reed et al., 2001; Fernandez et al., 2007; Zahedi et al., 2008), our work provides the first evidence of a signal from the AS that can shape the epidermis. This signal is likely to be a mechanical signal, as has been shown in other systems (Kolega, 1986; Leppa and Bohmann, 1999; Tran et al., 1999; Klepeis et al., 2001; Kushida et al., 2001; Ramet et al., 2002; Kaunas et al., 2006). In addition the work highlights that there are cooperative interactions between the two tissues and that some of these are mediated by Ush.
Materials and Methods
Drosophila strains
The following strains were used: ush2, tkv8, UAS-ush (Fossett et al., 2000), c381Gal4 (Manseau et al., 1997), da-GAL4, puc-lacZ (Ring and Martinez Arias, 1993). UAS lines were expressed using the UAS/GAL4 system (Brand and Perrimon, 1993). Wild-type stocks are Oregon R. Also, heterozygous flies which did not manifest any mutant phenotype were used as wild-type internal controls for homozygous mutant siblings. The following crosses were analyzed:
-tkv8/CTG; da-GAL4/TM6B X tkv8 UAS-ush/CTG; +
-ush2 UAS-ush/CyO X ush2 c381-GAL4/CyO
-ush2 UAS-ush/CyO X ush2/CTG; prd-GAL4/TM6B
-ush2 UAS-ush/CyO X ush2 c381-GAL4/CyO; prd-GAL4/TM6B
- ush2 UAS-ush/CyO X ush2 c381-GAL4/CyO; puc-lacZ/MKRS
Cuticle preparations
In all cases where mutant phenotypes were quantified, the balancer chromosome was outcrossed with a wild-type chromosome in order to eliminate homozygous balancer lethal embryos. This facilitates analysis, as subsequently all lethals should theoretically be homozygous mutants. At least 200 embryos were collected and aged 48 hours at 25°C, allowing the viable larvae to hatch and crawl out of the basket. Lethal specimens were dechorionated under a light microscope in 50% sodium hypochlorite and washed thoroughly 3 times with dH2O. They were transferred with a Pasteur pipette onto a cleaned slide, arranged so that each could be viewed individually, excess water wiped away, and mounted in Hoyers/acetic acid 1∶1. The slide was incubated overnight on a 60°C mini hotplate and specimens examined using Nomarski DIC imaging.
Quantitative embryonic lethality assays
Out of a minimum of 200 eggs total, hatched versus unhatched embryos were counted and the latter class was then processed for cuticle analysis as described above. Unhatched eggs were scored as lethal (those with cuticle deposition) or unfertilized (white and undeveloped). Embryonic lethality was then calculated as [number of lethal embryos]/([total eggs in basket] – [unfertilized eggs]). Since the flies were outcrossed to wild-type, and heterozygous parents predictably yield a quarter homozygous mutant offspring, 25% embryonic lethality can be assumed to be equivalent to 100% of the mutant class. The percent embryonic lethality of the mutant class is thus calculated as the [embryonic lethality]/0.25 * 100 (Reed et al., 2001).
The degree of germband retraction was measured by noting which abdominal segment was located at the posterior tip of the embryo; half-segment measures were used to increase resolution. Complete retraction occurred when the telson is found at the tip (Yip et al., 1997). Dorsal closure defects were manifest with a conspicuous hole in the dorsal cuticle. For simplicity and accuracy of dorsal closure defect comparison across different genotypes, only cuticles which displayed holes in their epidermis were counted as DC failure.
Immunostainings
Embryos were fixed and stained as described previously (Kaltschmidt et al., 2002). The following antibodies were used: rat anti-α-tubulin (D. Glover, 1∶200), mouse anti-Armadillo (Developmental Studies Hybridoma Bank (DSHB), University of Iowa, developed by E. Wieschaus, 1∶50), mouse anti-β-Galactosidase (Promega, 1∶200), rabbit anti-β-Galactosidase (Cappel, 1∶1000), rat anti-DECadherin (DSHB, University of Iowa, developed by T. Uemura, 1∶50), mouse anti-Fasciclin III (DSHB, University of Iowa, developed by C. Goodman, 1∶50), rabbit anti-Ush (kindly provided by R. A. Schulz, 1∶1000) mouse anti-Hindsight (DSHB, University of Iowa, developed by H. D. Lipshitz, 1∶50), mouse anti-phospho-Tyrosine (Santa Cruz, 1∶1000). Alexa fluor 488, 568 and 647 (Molecular Probes, 1∶200) were used as secondary antobodies.
Quantification of fluorescence intensities
For quantification of fluorescence intensity, the polygon selection tool from ImageJ was used to draw around an object (nuclei) and the mean grey value was obtained. Several nuclei from at least 3 different embryos were measured to obtain the mean intensity value for each embryo (M1). A mean background value was measured (B1) and subtracted from the expression value. A Student's t-test was used for all statistical comparisons of fluorescence intensities.
Confocal microscopy
Confocal imaging was performed using a Nikon Eclipse E800 microscope equipped with a Bio-Rad MRC 1024ES laser scanning confocal system (with emission lines at 488 nm, 568 nm and 647 nm) and processed with Bio-Rad Lasersharp 2000 software. 8-bit 1024 × 1024 pixel images were collected using a 63×/1.4 NA, 40×/1.30 NA, or 20×/0.75 NA oil immersion objective lens and Kalman filter. RGB channels were merged and Z-sections projected (at maximal intensity) using ImageJ and then image brightness and contrast adjusted using Adobe Photoshop CS3.
Acknowledgments
We thank members of our lab for fruitful discussions. This work was supported by a Biotechnology and Biological Sciences Research Council grant to NG and AMA.
Footnotes
Competing interests: The authors declare no competing interests.
References
- Affolter M., Basler K. (2007). The Decapentaplegic morphogen gradient: from pattern formation to growth regulation. Nat. Rev. Genet. 8, 663–674 10.1038/nrg2166 [DOI] [PubMed] [Google Scholar]
- Affolter M., Nellen D., Nussbaumer U., Basler K. (1994). Multiple requirements for the receptor serine/threonine kinase thick veins reveal novel functions of TGF beta homologs during Drosophila embryogenesis. Development 120, 3105–3117. [DOI] [PubMed] [Google Scholar]
- Arquier N., Perrin L., Manfruelli P., Semeriva M. (2001). The Drosophila tumor suppressor gene lethal(2)giant larvae is required for the emission of the Decapentaplegic signal. Development 128, 2209–2220. [DOI] [PubMed] [Google Scholar]
- Azuma N., Duzgun S. A., Ikeda M., Kito H., Akasaka N., Sasajima T., Sumpio B. E. (2000). Endothelial cell response to different mechanical forces. J. Vasc. Surg. 32, 789–794 10.1067/mva.2000.107989 [DOI] [PubMed] [Google Scholar]
- Bates K. L., Higley M., Letsou A. (2008). Raw mediates antagonism of AP-1 activity in Drosophila. Genetics 178, 1989–2002 10.1534/genetics.107.086298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beanes S. R., Dang C., Soo C., Ting K. (2003). Skin repair and scar formation: the central role of TGF-beta. Expert Rev. Mol. Med. 5, 1–22 10.1017/S1462399403005817 [DOI] [PubMed] [Google Scholar]
- Blanchard G. B., Murugesu S., Adams R. J., Martinez-Arias A., Gorfinkiel N. (2010). Cytoskeletal dynamics and supracellular organisation of cell shape fluctuations during dorsal closure. Development 137, 2743–2752 10.1242/dev.045872 [DOI] [PubMed] [Google Scholar]
- Brand A. H., Perrimon N. (1993). Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118, 401–415. [DOI] [PubMed] [Google Scholar]
- Chen H. W., Marinissen M. J., Oh S. W., Chen X., Melnick M., Perrimon N., Gutkind J. S., Hou S. X. (2002). CKA, a novel multidomain protein, regulates the JUN N-terminal kinase signal transduction pathway in Drosophila. Mol. Cell. Biol. 22, 1792–1803 10.1128/MCB.22.6.1792-1803.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colas J. F., Schoenwolf G. C. (2001). Towards a cellular and molecular understanding of neurulation. Dev. Dyn. 221, 117–145 10.1002/dvdy.1144 [DOI] [PubMed] [Google Scholar]
- David D. J., Tishkina A., Harris T. J. (2010). The PAR complex regulates pulsed actomyosin contractions during amnioserosa apical constriction in Drosophila. Development 137, 1645–1655 10.1242/dev.044107 [DOI] [PubMed] [Google Scholar]
- Fernandez B. G., Arias A. M., Jacinto A. (2007). Dpp signalling orchestrates dorsal closure by regulating cell shape changes both in the amnioserosa and in the epidermis. Mech. Dev. 124, 884–897 10.1016/j.mod.2007.09.002 [DOI] [PubMed] [Google Scholar]
- Fossett N., Zhang Q., Gajewski K., Choi C. Y., Kim Y., Schulz R. A. (2000). The multitype zinc-finger protein U-shaped functions in heart cell specification in the Drosophila embryo. Proc. Natl. Acad. Sci. USA 97, 7348–7353 10.1073/pnas.97.13.7348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank L. H., Rushlow C. (1996). A group of genes required for maintenance of the amnioserosa tissue in Drosophila. Development 122, 1343–1352. [DOI] [PubMed] [Google Scholar]
- Franke J. D., Montague R. A., Kiehart D. P. (2005). Nonmuscle myosin II generates forces that transmit tension and drive contraction in multiple tissues during dorsal closure. Curr. Biol. 15, 2208–2221 10.1016/j.cub.2005.11.064 [DOI] [PubMed] [Google Scholar]
- Glise B., Noselli S. (1997). Coupling of Jun amino-terminal kinase and Decapentaplegic signaling pathways in Drosophila morphogenesis. Genes Dev. 11, 1738–1747 10.1101/gad.11.13.1738 [DOI] [PubMed] [Google Scholar]
- Goldman-Levi R., Miller C., Greenberg G., Gabai E., Zak N. B. (1996). Cellular pathways acting along the germband and in the amnioserosa may participate in germband retraction of the Drosophila melanogaster embryo. Int. J. Dev. Biol. 40, 1043–1051. [PubMed] [Google Scholar]
- Gorfinkiel N., Martínez Arias A. (2007). Requirements for Adherens Junctions components in the interaction between epithelial tissues during Dorsal Closure in Drosophila. J. Cell Sci. 120, 3289–3298 10.1242/jcs.010850 [DOI] [PubMed] [Google Scholar]
- Gorfinkiel N., Blanchard G. B., Adams R. J., Martinez-Arias A. (2009). Mechanical control of global cell behaviour during Dorsal Closure in Drosophila. Development 136, 1889–1898 10.1242/dev.030866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorfinkiel N., Schamberg S., Blanchard G. B. (2011). Integrative approaches to morphogenesis: lessons from dorsal closure. Genesis 49, 522–533 10.1002/dvg.20704 [DOI] [PubMed] [Google Scholar]
- Harden N. (2002). Signaling pathways directing the movement and fusion of epithelial sheets: lessons from dorsal closure in Drosophila. Differentiation 70, 181–203 10.1046/j.1432-0436.2002.700408.x [DOI] [PubMed] [Google Scholar]
- Harden N., Ricos M., Yee K., Sanny J., Langmann C., Yu H., Chia W., Lim L. (2002). Drac1 and Crumbs participate in amnioserosa morphogenesis during dorsal closure in Drosophila. J. Cell Sci. 115, 2119–2129. [DOI] [PubMed] [Google Scholar]
- Hou X. S., Goldstein E. S., Perrimon N. (1997). Drosophila Jun relays the Jun amino-terminal kinase signal transduction pathway to the Decapentaplegic signal transduction pathway in regulating epithelial cell sheet movement. Genes Dev. 11, 1728–1737 10.1101/gad.11.13.1728 [DOI] [PubMed] [Google Scholar]
- Hutson M. S., Tokutake Y., Chang M. S., Bloor J. W., Venakides S., Kiehart D. P., Edwards G. S. (2003). Forces for morphogenesis investigated with laser microsurgery and quantitative modeling. Science 300, 145–149 10.1126/science.1079552 [DOI] [PubMed] [Google Scholar]
- Jacinto A., Wood W., Balayo T., Turmaine M., Martinez-Arias A., Martin P. (2000). Dynamic actin-based epithelial adhesion and cell matching during Drosophila dorsal closure. Curr. Biol. 10, 1420–1426 10.1016/S0960-9822(00)00796-X [DOI] [PubMed] [Google Scholar]
- Jacinto A., Woolner S., Martin P. (2002). Dynamic analysis of dorsal closure in Drosophila: from genetics to cell biology. Dev. Cell 3, 9–19 10.1016/S1534-5807(02)00208-3 [DOI] [PubMed] [Google Scholar]
- Jankovics F., Brunner D. (2006). Transiently reorganized microtubules are essential for zippering during dorsal closure in Drosophila melanogaster. Dev. Cell 11, 375–385 10.1016/j.devcel.2006.07.014 [DOI] [PubMed] [Google Scholar]
- Kaltschmidt J. A., Lawrence N., Morel V., Balayo T., Fernandez B. G., Pelissier A., Jacinto A., Martinez Arias A. (2002). Planar polarity and actin dynamics in the epidermis of Drosophila. Nat. Cell Biol. 4, 937–944 10.1038/ncb882 [DOI] [PubMed] [Google Scholar]
- Kaunas R., Usami S., Chien S. (2006). Regulation of stretch-induced JNK activation by stress fiber orientation. Cell. Signal. 18, 1924–1931 10.1016/j.cellsig.2006.02.008 [DOI] [PubMed] [Google Scholar]
- Keller R. E., Trinkaus J. P. (1987). Rearrangement of enveloping layer cells without disruption of the epithelial permeability barrier as a factor in Fundulus epiboly. Dev. Biol. 120, 12–24 10.1016/0012-1606(87)90099-6 [DOI] [PubMed] [Google Scholar]
- Kiehart D. P., Galbraith C. G., Edwards K. A., Rickoll W. L., Montague R. A. (2000). Multiple forces contribute to cell sheet morphogenesis for dorsal closure in Drosophila. J. Cell Biol. 149, 471–490 10.1083/jcb.149.2.471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klepeis V. E., Cornell-Bell A., Trinkaus-Randall V. (2001). Growth factors but not gap junctions play a role in injury-induced Ca2+ waves in epithelial cells. J. Cell Sci. 114, 4185–4195. [DOI] [PubMed] [Google Scholar]
- Knust E. (1996). Drosophila morphogenesis: follow-my-leader in epithelia. Curr. Biol. 6, 379–381 10.1016/S0960-9822(02)00500-6 [DOI] [PubMed] [Google Scholar]
- Kockel L., Zeitlinger J., Staszewski L. M., Mlodzik M., Bohmann D. (1997). Jun in Drosophila development: redundant and nonredundant functions and regulation by two MAPK signal transduction pathways. Genes Dev. 11, 1748–1758 10.1101/gad.11.13.1748 [DOI] [PubMed] [Google Scholar]
- Kolega J. (1986). Effects of mechanical tension on protrusive activity and microfilament and intermediate filament organization in an epidermal epithelium moving in culture. J. Cell Biol. 102, 1400–1411 10.1083/jcb.102.4.1400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kushida N., Kabuyama Y., Yamaguchi O., Homma Y. (2001). Essential role for extracellular Ca(2+) in JNK activation by mechanical stretch in bladder smooth muscle cells. Am. J. Physiol. Cell Physiol. 281, C1165–C1172. [DOI] [PubMed] [Google Scholar]
- Lamka M. L., Lipshitz H. D. (1999). Role of the amnioserosa in germ band retraction of the Drosophila melanogaster embryo. Dev. Biol. 214, 102–112 10.1006/dbio.1999.9409 [DOI] [PubMed] [Google Scholar]
- Leppa S., Bohmann D. (1999). Diverse functions of JNK signaling and c-Jun in stress response and apoptosis. Oncogene 18, 6158–6162 10.1038/sj.onc.1203173 [DOI] [PubMed] [Google Scholar]
- Manseau L., Baradaran A., Brower D., Budhu A., Elefant F., Phan H., Philp A. V., Yang M., Glover D., Kaiser K. et al. (1997). GAL4 enhancer traps expressed in the embryo, larval brain, imaginal discs, and ovary of Drosophila. Dev. Dyn. 209, 310–322 [DOI] [PubMed] [Google Scholar]
- Martin P., Parkhurst S. M. (2004). Parallels between tissue repair and embryo morphogenesis. Development 131, 3021–3034 10.1242/dev.01253 [DOI] [PubMed] [Google Scholar]
- Martin P., Dickson M. C., Millan F. A., Akhurst R. J. (1993). Rapid induction and clearance of TGF beta 1 is an early response to wounding in the mouse embryo. Dev. Genet. 14, 225–238 10.1002/dvg.1020140309 [DOI] [PubMed] [Google Scholar]
- Martin-Blanco E., Gampel A., Ring J., Virdee K., Kirov N., Tolkovsky A. M., Martinez-Arias A. (1998). puckered encodes a phosphatase that mediates a feedback loop regulating JNK activity during dorsal closure in Drosophila. Genes Dev. 12, 557–570 10.1101/gad.12.4.557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez Arias A. (1993). Development and patterning of the larval epidermis of Drosophila. The development of Drosophila melanogaster 1, (Martinez Arias A, Bate M.) Plainview, NY: Cold Spring Harbor Laboratory Press. [Google Scholar]
- Millard T. H., Martin P. (2008). Dynamic analysis of filopodial interactions during the zippering phase of Drosophila dorsal closure. Development 135, 621–626 10.1242/dev.014001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peralta X. G., Toyama Y., Hutson M. S., Montague R., Venakides S., Kiehart D. P., Edwards G. S. (2007). Upregulation of forces and morphogenic asymmetries in dorsal closure during Drosophila development. Biophys. J. 92, 2583–2596 10.1529/biophysj.106.094110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramet M., Lanot R., Zachary D., Manfruelli P. (2002). JNK signaling pathway is required for efficient wound healing in Drosophila. Dev. Biol. 241, 145–156 10.1006/dbio.2001.0502 [DOI] [PubMed] [Google Scholar]
- Reed B. H., Wilk R., Lipshitz H. D. (2001). Downregulation of Jun kinase signaling in the amnioserosa is essential for dorsal closure of the Drosophila embryo. Curr. Biol. 11, 1098–1108 10.1016/S0960-9822(01)00318-9 [DOI] [PubMed] [Google Scholar]
- Ricos M. G., Harden N., Sem K. P., Lim L., Chia W. (1999). Dcdc42 acts in TGF-beta signaling during Drosophila morphogenesis: distinct roles for the Drac1/JNK and Dcdc42/TGF-beta cascades in cytoskeletal regulation. J. Cell Sci. 112, 1225–1235. [DOI] [PubMed] [Google Scholar]
- Riesgo-Escovar J. R., Hafen E. (1997). Drosophila Jun kinase regulates expression of decapentaplegic via the ETS-domain protein Aop and the AP-1 transcription factor DJun during dorsal closure. Genes Dev. 11, 1717–1727 10.1101/gad.11.13.1717 [DOI] [PubMed] [Google Scholar]
- Ring J. M., Martinez Arias A. (1993). puckered, a gene involved in position-specific cell differentiation in the dorsal epidermis of the Drosophila larva. Dev. Suppl, 251–259. [PubMed] [Google Scholar]
- Schock F., Perrimon N. (2002). Cellular processes associated with germ band retraction in Drosophila. Dev. Biol. 248, 29–39 10.1006/dbio.2002.0698 [DOI] [PubMed] [Google Scholar]
- Scuderi A., Letsou A. (2005). Amnioserosa is required for dorsal closure in Drosophila. Dev. Dyn. 232, 791–800 10.1002/dvdy.20306 [DOI] [PubMed] [Google Scholar]
- Sluss H. K., Davis R. J. (1997). Embryonic morphogenesis signaling pathway mediated by JNK targets the transcription factor JUN and the TGF-beta homologue decapentaplegic. J. Cell. Biochem. 67, 1–12 [DOI] [PubMed] [Google Scholar]
- Sluss H. K., Han Z., Barrett T., Goberdhan D. C., Wilson C., Davis R. J., Ip Y. T. (1996). A JNK signal transduction pathway that mediates morphogenesis and an immune response in Drosophila. Genes Dev. 10, 2745–2758 10.1101/gad.10.21.2745 [DOI] [PubMed] [Google Scholar]
- Solon J., Kaya-Copur A., Colombelli J., Brunner D. (2009). Pulsed forces timed by a ratchet-like mechanism drive directed tissue movement during dorsal closure. Cell 137, 1331–1342 10.1016/j.cell.2009.03.050 [DOI] [PubMed] [Google Scholar]
- Stronach B. E., Perrimon N. (2001). Investigation of leading edge formation at the interface of amnioserosa and dorsal ectoderm in the Drosophila embryo. Development 128, 2905–2913. [DOI] [PubMed] [Google Scholar]
- Stronach B., Perrimon N. (2002). Activation of the JNK pathway during dorsal closure in Drosophila requires the mixed lineage kinase, slipper. Genes Dev. 16, 377–387 10.1101/gad.953002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su Y. C., Treisman J. E., Skolnik E. Y. (1998). The Drosophila Ste20-related kinase misshapen is required for embryonic dorsal closure and acts through a JNK MAPK module on an evolutionarily conserved signaling pathway. Genes Dev. 12, 2371–2380 10.1101/gad.12.15.2371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taya Y., O'Kane S., Ferguson M. W. (1999). Pathogenesis of cleft palate in TGF-beta3 knockout mice. Development 126, 3869–3879. [DOI] [PubMed] [Google Scholar]
- Tran P. O., Hinman L. E., Unger G. M., Sammak P. J. (1999). A wound-induced [Ca2+]i increase and its transcriptional activation of immediate early genes is important in the regulation of motility. Exp. Cell Res. 246, 319–326 10.1006/excr.1998.4239 [DOI] [PubMed] [Google Scholar]
- VanHook A., Letsou A. (2008). Head involution in Drosophila: genetic and morphogenetic connections to dorsal closure. Dev. Dyn. 237, 28–38 10.1002/dvdy.21405 [DOI] [PubMed] [Google Scholar]
- Wada A., Kato K., Uwo M. F., Yonemura S., Hayashi S. (2007). Specialized extraembryonic cells connect embryonic and extraembryonic epidermis in response to Dpp during dorsal closure in Drosophila. Dev. Biol. 301, 340–349 10.1016/j.ydbio.2006.09.020 [DOI] [PubMed] [Google Scholar]
- Williams-Masson E. M., Malik A. N., Hardin J. (1997). An actin-mediated two-step mechanism is required for ventral enclosure of the C. elegans hypodermis. Development 124, 2889–2901. [DOI] [PubMed] [Google Scholar]
- Yip M. L., Lamka M. L., Lipshitz H. D. (1997). Control of germ-band retraction in Drosophila by the zinc-finger protein HINDSIGHT. Development 124, 2129–2141. [DOI] [PubMed] [Google Scholar]
- Zahedi B., Shen W., Xu X., Chen X., Mahey M., Harden N. (2008). Leading edge-secreted Dpp cooperates with ACK-dependent signaling from the amnioserosa to regulate myosin levels during dorsal closure. Dev. Dyn. 237, 2936–2946 10.1002/dvdy.21722 [DOI] [PubMed] [Google Scholar]
- Zeitlinger J., Kockel L., Peverali F. A., Jackson D. B., Mlodzik M., Bohmann D. (1997). Defective dorsal closure and loss of epidermal decapentaplegic expression in Drosophila fos mutants. EMBO J. 16, 7393–7401 10.1093/emboj/16.24.7393 [DOI] [PMC free article] [PubMed] [Google Scholar]