

Abstract
The SH2-containing inositol phosphatase-1 (SHIP-1) is a 5′ inositol phosphatase known to negatively regulate the product of phosphoinositide-3 kinase (PI3K), phosphatidylinositol-3.4,5-trisphosphate. SHIP-1 can be recruited to a large number of inhibitory receptors expressed on natural killer (NK) cells. However, its role in NK cell development, maturation, and functions is not well defined. In this study, we found that the absence of SHIP-1 results in a loss of peripheral NK cells. However, using chimeric mice we demonstrated that SHIP-1 expression is not required intrinsically for NK cell lineage development. In contrast, SHIP-1 is required cell autonomously for NK cell terminal differentiation. These findings reveal both a direct and indirect role for SHIP-1 at different NK cell development checkpoints. Notably, SHIP-1–deficient NK cells display an impaired ability to secrete IFN-γ during cytokine receptor–mediated responses, whereas immunoreceptor tyrosine–based activation motif containing receptor-mediated responses is not affected. Taken together, our results provide novel insights on how SHIP-1 participates in the development, maturation, and effector functions of NK cells.
Introduction
Natural killer (NK) cells are large granular lymphocytes with a critical role in innate immunity and are important in orchestrating the adaptive immune system.1,2 Their ability to produce immunoregulatory cytokines, such as interferon (IFN)–γ, and release perforin and granzymes is crucial for tumor immunosurveillance and elimination of pathogens.3 These NK effector functions are dependent on the integration of signals delivered from a variety of activating and inhibitory receptors on interaction with neighboring cells.4 Significant progress has been made on the regulation of receptor and target recognition as well as identification of signals transduced for NK cell action. However, evidence for how these signaling molecules, specifically phosphatases, might govern NK cell development and thereby influence their effector function has been lacking. A better understanding of the mechanisms that govern NK cell development into functional effector cells is crucial to demystifying disease processes and fully utilizing NK cells as therapeutic agents.
The bone marrow (BM) is the main site for NK cell development. An intact BM microenvironment provides NK cells with both cellular substrates and signals required from several stromal factors to sustain cell proliferation and differentiation.1 NK cell precursors (NKPs) in the BM are derived from hematopoietic stem cells that give rise to immature NK (iNK) cells and mature NK (mNK) cells.5 mNK cells egress from the BM and represent the main NK cell population in the peripheral lymphoid organs, such as the spleen. The NK cell maturation process in the BM has been well characterized based on the differential acquisition of NK cell receptors and the achievement of their full effector functions.1,6,7
The acquisition of both activating and inhibitory receptors results in the ability of NK cells to recognize and kill target cells with minimal damage to the host. On NK cell synapse formation, appropriate downstream signaling molecules are recruited.8 Activating receptors are known to bind DAP10 or 12 containing an immunoreceptor tyrosine–based activation motif (ITAM) or YxxM motifs9 and subsequently recruit Src- and Syk-related protein tyrosine kinases, leading to the activation of PI3K and MAP/ERK pathways.10 Inhibitory receptors are known to oppose the activation of these pathways through the recruitment of various phosphatases to their immunoreceptor tyrosine–based inhibitory motif.11
Among these phosphatases, the SH2-containing inositol phosphatase-1 (SHIP-1) is known to be recruited to the immunoreceptor tyrosine–based inhibitory motif and suppress immune responses, including NK cells, by degrading PI3K's product phosphatidylinositol-3,4,5-trisphosphate to phosphatidylinositol-3,4-bisphosphate.12–14 SHIP-1 is primarily expressed in hematopoietic cells, but it can be found in osteoblasts and stromal cells.15–18 Although the role of SHIP-1 has been intensely studied in the development of myeloid cells, B cells, and T cells, its exact function in NK cell development remains poorly understood.
In this study, we found that the absence of SHIP-1 results in a decrease of peripheral NK cell number. In addition, NK cells lacking SHIP-1 exhibited fewer mature attributes, as characterized by the expression of certain receptors. However, the loss of peripheral NK cells is partly restored in bone marrow chimeras and mixed bone marrow chimeras, indicating that SHIP-1 intrinsic expression is not essential for NK cell cellularity. In contrast, SHIP-1 intrinsic expression is required for the transition from immature to mature NK cells. Thus, SHIP-1 expression on stromal cells is sufficient to rescue NK cell frequency and number but is required in NK cells for final stages of differentiation. In addition, we found that NK cells deficient for SHIP-1 display an impaired ability to secrete IFN-γ during cytokine receptor–mediated responses.
Methods
Reagents and antibodies
The following anti–mouse monoclonal antibodies (mAbs) were used for flow cytometry: NK1.1 (PK136), CD19 (eBio1D3), CD3 (145-2C11 or 17A2), CD8 (17A2), CD4 (RM4-5), NKp46 (29A1.4), CD11c (N418), CD45.1 (A20), H2Db (KH95), H2Kb (AF6-88.5), CD122 (TMβ1), CD27 (LG.7F9), CD11b (M1/72), KLRG1 (2F1), CD43 (eBioR2/60), CD51 (RMV-7), cKit (2B8), CD94 (18d3), Ly49F/I/C/H (14B11), Ly49G2 (4D11), Ly49H (3D10), CD49b (DX5), CD107α (1D4B), Lin antibody cocktail, CD127 (A7R34), CD135 (A2F10), Sca-1 (D7), and IFN-γ (XMG1.2). Single-cell suspensions were preincubated with Fc receptor–blocking 2.4G2 mAb and stained for 30 minutes on ice with the indicated surface staining antibodies (all purchased from BD Biosciences and eBioscience). Washed cells were evaluated on an FACSAria cell sorter (BD Biosciences) and analyzed with FlowJo Version 8.8.7 software (TreeStar).
Mice
C57BL/6J (B6) mice were purchased from The Jackson Laboratory. SHIP-1−/− mice (on a B6.SJL background) were generated and kindly provided by W.G.K.14 Five- to 8-week-old SHIP-1−/− mice were used for experiments. Wild-type and SHIP-1+/− littermates were used as controls. Heterozygote siblings were bred. Rag2−/− × Il2rγ−/− mice were purchased from Taconic Laboratory Animals and Services. All mice were maintained at Brown University in accordance with institutional guidelines for animal care and use.
Bone marrow chimeras and mixed chimeras
To generate bone marrow chimeras, 6- to 8-week old B6 mice were lethally irradiated with 1000 to 1050 rads. Donor bone marrow was harvested from SHIP-1−/− or SHIP-1+/− (CD45.1-congenic B6) mice. Bone marrow cells were depleted from mature T and NK cells using anti-CD5 and anti-DX5 magnetic beads, respectively, via the AutoMACS separator (Miltenyi Biotec) and injected intravenously into irradiated recipients. All animals were allowed to reconstitute for 4 to 10 weeks before sacrifice and the NK cell compartment of liver, spleen, bone marrow, and blood was analyzed by flow cytometry. For mixed chimeras, donor bone marrow was harvested from SHIP-1−/− or SHIP-1+/−(CD45.1-congenic B6) mice and mixed 1:1 with bone marrow harvested from competitor, CD45.2-congenic B6 mice; depleted of mature T and NK cells; and injected intravenously into irradiated recipients as described above. Mixed chimeras were allowed to reconstitute 8 to 10 weeks before sacrifice.
Adoptive transfer
Splenocytes (107) isolated from mixed bone marrow chimeras were adoptively transferred into Rag2−/− × Il2rγ−/− mice. At day 4 after transfer, splenocytes were isolated from Rag2−/− × Il2rγ−/− mice and stained with NK1.1, CD3, CD45.1, CD11b, KLRG1, and CD27.
Functional assays
For measurement of IFN-γ production and CD107α induction, splenocytes from mice either unprimed or primed with Poly(I:C) were stimulated in tissue culture plates coated with anti-NK1.1, anti-NKG2D, anti-Ly49H, or control mAb for 6 hours at 37°C in the presence of GolgiStop (BD Biosciences) and CD107α or isotype-matched control antibody (eBioscience). For control experiments, splenocytes were stimulated with IL-12 and IL-18 (10 ng/mL each) or phorbol myristate acetate (PMA; 200 ng/mL) and ionomycin (5 μg/mL). After stimulation, NK cells were then defined by CD3−NK1.1+ or CD3−DX5+ (for NK1.1 stimulation) and then were fixed and made permeable with Cytofix/Cytoperm buffer (BD Biosciences). Cells were then stained with IFN-γ or isotype-matched control antibody (eBioscience).
Statistical analysis
Statistical analyses were performed using Prism Version 5.0 (GraphPad Software).
Results
Reduced number of NK cells in SHIP-1−/− mice
SHIP-1 has been reported to associate with several inhibitory NK cell receptors including certain Ly49 receptors and the SLAM family member 2B4.14,19 SHIP-1 absence has been shown to differentially influence B- and T-cell development.20–23 In addition, previous studies demonstrate the importance of SHIP-1 in the generation of the NK cell receptor repertoire.14,24,25 We therefore hypothesized that SHIP-1 may play a role in NK cell development and maturation. To test this hypothesis, we examined lymphocyte populations in the spleen, blood, liver, and bone marrow of SHIP-1+/+, SHIP-1+/−, and SHIP-1−/− mice. NK cell number and frequency were significantly reduced in the spleen and blood, but not in the bone marrow of SHIP-1−/− mice compared with SHIP-1+/+ and SHIP-1+/− mice (Figure 1A-B; data not shown). A similar trend was also observed in the spleen and bone marrow when further defining the NK cell population as NKp46+CD3− (supplemental Figure 1A, available on the Blood Web site; see the Supplemental Materials link at the top of the online article). As reported by others, the absence of SHIP-1 was also found to significantly reduce splenic B cells (CD19+; supplemental Figure 1B), and T cells (CD4+ or CD8+; supplemental Figure 1C-E). Notably, iNKT cell frequency also was affected (data not shown). Interestingly, SHIP-1 expression is haplosufficient, as NK cell frequency and numbers in SHIP-1+/− mice were comparable to SHIP-1+/+ mice (Figure 1 and supplemental Figure 1).
Figure 1.
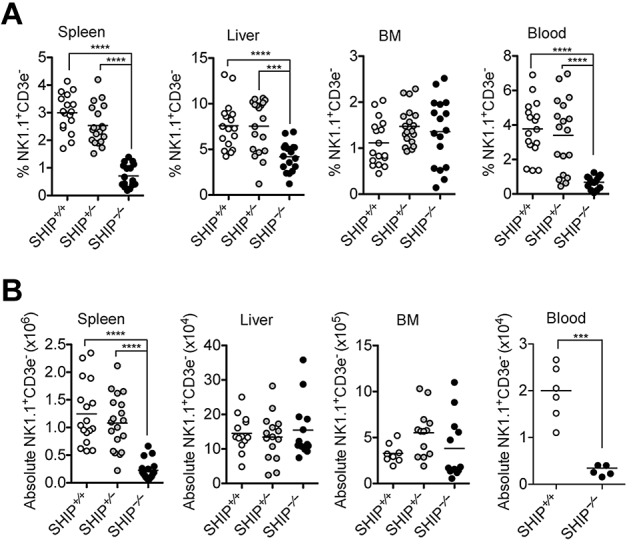
Loss of peripheral NK cells in SHIP−/− mice. (A) Percentages of the indicated organs containing NK cell (NK1.1+CD3− in the lymphocyte gate) populations in the indicated organs from SHIP+/+ (white circles), SHIP+/− (gray circles), and SHIP−/− (black circles) mice. Data are pooled from at least 8 independent experiments, and each dot represents data obtained from 1 mouse; horizontal lines indicate the mean. (B) Total NK cell numbers of the indicated organs. Data are pooled from at least 3 independent experiments, and each dot represents data obtained from one mouse; horizontal lines indicate the mean.
Immature NK cell subsets are overrepresented in SHIP-1−/− mice
NK cells in the periphery are mainly mNK cells that developed from bone marrow hematopoietic stem cells through NKP and iNK cell populations.26 The much lower NK cell numbers in the periphery compared with the bone marrow suggested that SHIP-1−/− mice might have a defect in NK cell development. To examine this possibility, we compared the different NK cell subsets, NKP, iNK, and mNK cells, in the bone marrow6 (Figure 2A and supplemental Figure 2A-B). We found significantly less NKP and mNK cells, but no substantial differences in iNK cells in SHIP-1−/− bone marrow compared with controls.
Figure 2.
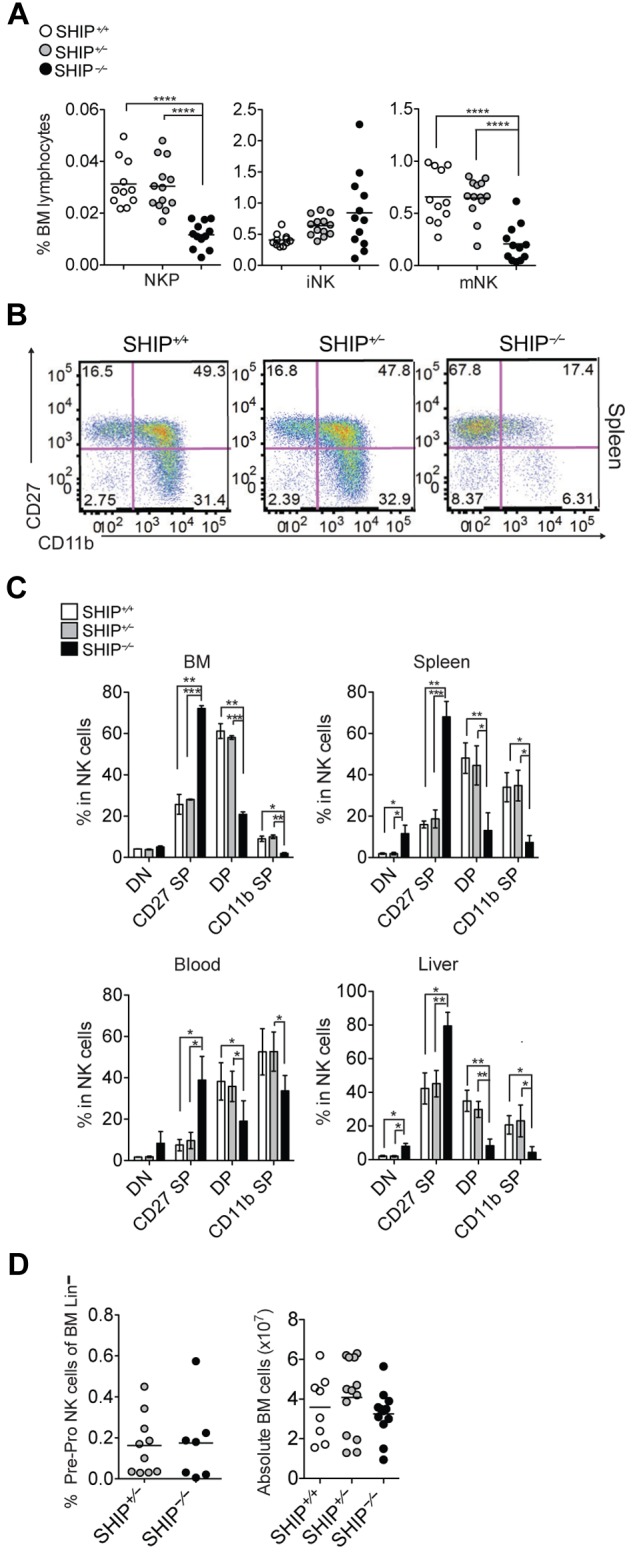
Impaired NK cell development in BM and periphery of SHIP−/− mice. (A) Percentages of NKPs (CD122+NK1.1−CD3−CD11b−), iNK cells (CD122+NK1.1+CD3−CD11b−), and mNK cells (CD122+NK1.1+CD3−CD11b+) among bone marrow lymphocytes derived from littermate SHIP+/+ (white circles), SHIP+/− (gray circles), and SHIP−/− (black circles) mice. Data are pooled from at least 5 independent experiments, and each dot represents data obtained from one mouse; horizontal lines indicate the mean. (B) Representative staining in NK cells (NK1.1+CD3−) expressing DN (CD27−CD11b−), CD27 SP (CD11b−CD27+), DP (CD27+CD11b+), and CD11b SP (CD27−CD11b+) populations in spleens from SHIP+/+, SHIP+/−, and SHIP−/− mice. (C) Percentages in NK cells (NK1.1+CD3−) being DN, CD27 SP, DP, and CD11b SP populations in the indicated organs. Data are representative of at least 8 independent experiments. Data represents means ± SD. (D left panel) Frequency of bone marrow PrePro NK cells (Lineage−, Sca-1+, CD117−, CD135−, CD127+). Data are pooled from 3 independent experiments. (Right panel) Total number of bone marrow cells.
Because we found no differences in the iNK cell subset but a reduction in the mNK cell subset in the bone marrow, we hypothesized that the NK cells that egress to the periphery are mainly of the iNK cell subset in SHIP-1−/− mice. To test this hypothesis, we stained the bone marrow and spleen with NK maturation markers c-kit, CD51, CD43, CD27, DX5, CD11b, and KLRG1 (Table 1 and supplemental Figure 2E). The most mature NK cells express the markers DX5, CD11b, KLRG1, and CD43, but not the immature markers CD27, ckit, and CD51.2,5,6,27,28 We found that SHIP-1−/− bone marrow and splenic NK cells have a reduced expression of mature NK cell markers, such as DX5, CD11b, and KLRG1 compared with littermate controls (Table 1 and supplemental Figure 2E). Notably, SHIP-1−/− splenic NK cells express mostly immature NK cell markers. Taken together, this suggests that the few SHIP-1−/− NK cells found in the periphery are mainly immature NK cells.
Table 1.
Expression of maturation markers on NK cells from SHIP-1 deficient mice and littermate controls*
| BM | SHIP+/+ | SHIP+/− | SHIP−/− | Spleen | SHIP+/+ | SHIP+/− | SHIP−/− |
|---|---|---|---|---|---|---|---|
| cKit | 5.3 ± 0.5 | 5.8 ± 1.5 | 6.5 ± 3.4 | cKit | 6.5 ± 1.0 | 6.6 ± 0.4 | 21.4 ± 6.9 |
| CD51 | 5.5 ± 0.4 | 8.2 ± 2.0 | 8.5 ± 0.7 | CD51 | 3.7 ± 0.1 | 9.3 ± 1.9 | 42.2 ± 5.9 |
| CD94 | 45.1 ± 0.4 | 48.3 ± 4.1 | 56.2 ± 8.4 | CD94 | 45 ± 1.3 | 56.0 ± 0.9 | 58.3 ± 7.3 |
| Ly49s* | 69.27 ± 0.9 | 69.1 ± 5.5 | 67.0 ± 13.21 | Ly49s* | 69.7 ± 0.4 | 64.5 ± 2.0 | 35.5 ± 17.8 |
| CD43 | 97.4 ± 2.1 | 96.2 ± 1.6 | 97.4 ± 2.6 | CD43 | 99.3 ± 0.1 | 98.4 ± 0.5 | 92.8 ± 3.5 |
| CD27 | 70.0 ± 5.3 | 76.7 ± 3.2 | 79.3 ± 4.4 | CD27 | 64.2 ± 2.1 | 69.4 ± 2.8 | 85.1 ± 2.4 |
| DX5 | 94.9 ± 0.4 | 94.4 ± 1.1 | 84.9 ± 1.5 | DX5 | 92.2 ± 0.3 | 91.5 ± 0.9 | 68.5 ± 11.7 |
| CD11b | 61.6 ± 2.8 | 47.7 ± 2.1 | 12.2 ± 2.2 | CD11b | 84.3 ± 1.7 | 73.2 ± 0.7 | 21.7 ± 8.4 |
| KLRG1 | 12.6 ± 2.4 | 10.1 ± 1.6 | 4.6 ± 0.5 | KLRG1 | 21.5 ± 2.4 | 21.8 ± 5.3 | 10.8 ± 3.3 |
Results are expressed as mean frequency ± SD of 3 mice per genotype and are representative of at least five independent experiments. Values in bold represent P < .05 versus controls (SHIP+/+ and SHIP+/−). % marker-positive NK cells.
Results from a pool of antibodies specific for Ly49C/I/F/H and Ly49G2.
For NK cells to become fully mature effector cells, self-MHC class I molecule recognition by inhibitory receptors is required. This NK cell education process occurs during the iNK to mNK differentiation stage when NK cells acquire inhibitory receptors, such as Ly49s and CD94/NKG2A.29,30 We examined NK cells expressing CD94 and Ly49s (C/I/F/H/G2) in the bone marrow and in the spleen across all SHIP-1 genotypes (supplemental Figure 2C-D). No difference in bone marrow expression of CD94 and Ly49s, as well as splenic expression of CD94 was observed, but there was a significant decrease in splenic expression of Ly49s in SHIP-1−/− mice compared with controls (Table 1 and supplemental Figure 2C-E) as reported previously.24 We also looked at H2-Kb and H2-Db expression in total splenocytes as well as splenic NK cells (supplemental Figure 2F). SHIP-1−/− total splenocytes and splenic NK cells (as well as B, T, and dendritic cells; data not shown) express equivalent levels of MHC class I molecules as controls. Taken together, these data indicate that the immature phenotype of the SHIP-1−/− NK cells was unlikely to be because of an MHC/Ly49 defective educational environment. Notably, KLRG1 expression was not impaired because of the absence of SHIP-1 as KLRG1 expression can be induced on SHIP-1–deficient NK cells when cultured in vitro with IL-2 (supplemental Figure 2G).
It has been demonstrated that mouse NK cell maturation is a 4-stage developmental program: CD11b−CD27− (DN), CD11b−CD27+ (CD27 SP), CD11b+CD27+ (DP), and CD11b+CD27− (CD11b SP), following the pathway CD27 SP → DP → CD11b SP2. To further investigate NK cell subsets present in the periphery, we compared CD27 versus CD11b and KLRG1 expression from various organs across all SHIP-1 genotypes. Flow cytometric analysis revealed that DN and CD27 SP NK cells were the predominant NK cell subset present in SHIP-1−/− bone marrow and peripheral sites (spleen, blood, and liver), whereas SHIP-1+/+ and SHIP-1+/− NK cells were mostly DP and CD11b SP (Figure 2B-C and supplemental Figure 3A). KLRG1, a marker for the most mature NK cells, was also weakly expressed on SHIP-1−/− NK cells (supplemental Figure 3B-C) and observed previously in germ line SHIP-1−/− mice on a C57BL6 background.24 Collectively, these data suggest that SHIP-1 regulates NK cell transition from immature to mature NK cells. NK precursors were also reduced in SHIP-1−/− mice (Figure 2). However, when phenotyping the earliest NK cell precursors as recently described,31,32 we found no significant difference in frequency (Figure 2D and supplemental Figure 3D) and number, as there is no difference in the total number of bone marrow cells (Figure 2D). In addition, in chimeric mice no difference in NK cell precursor number was observed (see Figure 4C), suggesting that SHIP-1 absence is not affecting NK cell precursor formation.
Figure 4.
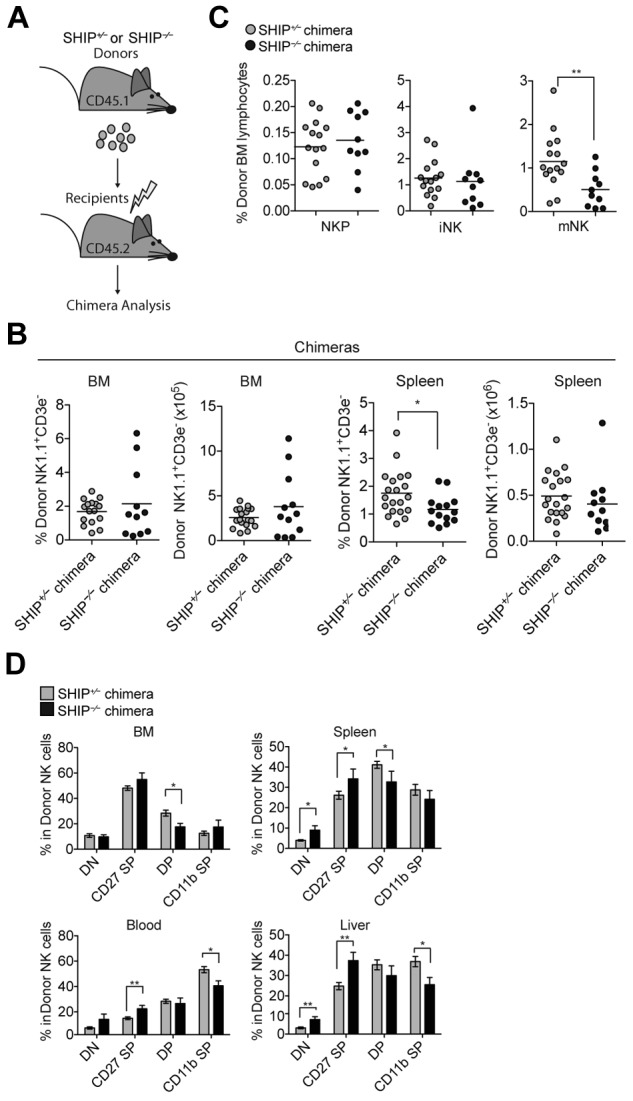
NK cell number is rescued in chimeric mice. (A) Diagram of chimera generation. SHIP+/− or SHIP−/− (CD45.1+) bone marrow cells were transplanted into lethally irradiated WT (CD45.2+) recipients and analyzed 4 to 9 weeks after reconstitution. (B) Percentage and total NK cells (NK1.1+CD3− in the lymphocyte gate) present in the indicated organs from SHIP+/− chimera (gray circles) and SHIP−/− chimera (black circles) mice. Data are pooled from at least 5 independent experiments, and each symbol indicates an individual mouse; horizontal lines indicate the mean. Data are pooled from at least 5 independent experiments, and each symbol indicates an individual mouse; horizontal lines indicate the mean. (C) Percentage of donor contributions of NKPs (CD122+NK1.1−CD3−CD11b−), iNK cells (CD122+NK1.1+CD3−CD11b−), and mNK cells (CD122+NK1.1+CD3−CD11b+) among bone marrow lymphocytes derived from SHIP+/− chimeras (gray circles) and SHIP−/− chimeras (black circles) mice. Data are pooled from at least 5 independent experiments, and each symbol indicates an individual mouse; horizontal lines indicate the mean. (D) Percentages in NK cells (NK1.1+CD3− in the lymphocyte gate) expressing DN (CD27−CD11b−), CD27 SP (CD11b−CD27+), DP (CD27+CD11b+), and CD11b SP (CD27−CD11b+) populations in the indicated organs derived from SHIP+/− chimera (gray bars) and SHIP−/− chimera (black bars) mice. Data are pooled from at least 5 independent experiments. Error bars indicate SEM.
Cytokine-mediated response is impaired in SHIP-1–deficient mice
To address the role of SHIP-1 in NK cell function, we measured both NK cell IFN-γ production and degranulation (monitored by CD107α surface expression) by flow cytometry. Because of the lower expression levels of Ly49H and NK1.1 in SHIP-1−/− NK cells (supplemental Figure 4A), cross-linking of these receptors will be inconclusive. We addressed this issue using mixed bone marrow chimeras, as the expression of these activating receptors on bone marrow–derived SHIP-1−/− and wild-type NK cells are comparable (see supplemental Figure 5B). However, when stimulated with the cytokines IL-12 and IL-18 or PMA and ionomycin, SHIP-1−/− NK cells are producing IFN-γ at a significantly lower level than control NK cells (Figure 3 and supplemental Figure 4B). Thus, the absence of SHIP-1 seems to be associated with impaired NK cell effector functions at least when SHIP-1 is systematically deleted.
Figure 3.
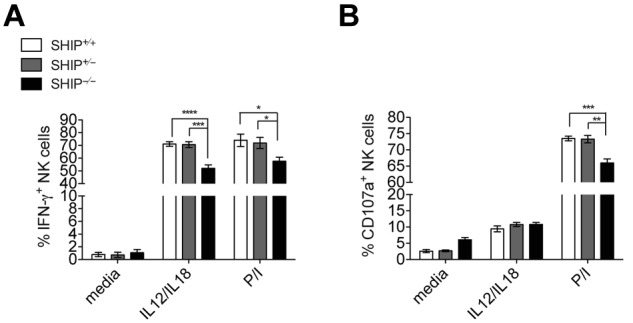
Ex vivo hyporesponsiveness of NK cells from SHIP−/− mice. Intracellular IFN-γ (A) and CD107α (B) in NK cell populations (NK1.1+CD3− in the lymphocyte gate) from total splenocytes incubated with a mix of IL-12 and IL-18 or PMA and ionomycin (P/I) for 6 hours. Data are pooled from at least 3 independent experiments. Error bars indicate SEM.
SHIP-1 acts at 2 different levels during NK cell development
SHIP-1 is expressed primarily in hematopoietic cells but also in osteoblasts and bone marrow stromal cells.15–18 Notably, SHIP-1−/− mice have a short life span (average 5-9 weeks) because of SHIP-1 pleiotropic effects. Therefore, we investigated whether the defect in NK cell development, maturation, and effector functions observed in SHIP-1−/− mice was because of the absence of SHIP-1 in NK cells (cell intrinsic) or in their environment (cell extrinsic). To determine whether nonhematopoietic cells play a role in the NK cell development defect observed, we transferred bone marrow cells from SHIP-1−/− or SHIP-1+/− (CD45.1+) mice into lethally irradiated wild-type (CD45.2+) mice (Figure 4A). In contrast to SHIP-1–deficient mice (Figure 1), no difference in NK cell numbers was observed in the bone marrows and spleens of the SHIP-1−/− chimera or the SHIP+/− chimera (Figure 4B). Moreover, no difference was seen in NK precursors and immature NK cell frequency in the bone marrow of the SHIP-1−/− chimera and the SHIP+/− chimera (Figure 4C). These results indicate that SHIP-1 expression is not required in cis and that expression of SHIP-1 on nonhematopoietic cells is sufficient to rescue normal NK cell number. However, we observed a decrease in mature NK cells derived from SHIP-1−/− bone marrows compared with the SHIP-1+/− bone marrow–derived NK cells (Figure 4C). Consistent with its least mature phenotype, the SHIP-1−/− NK cells display a decrease in CD11b SP NK cells and an increase in CD27 SP NK cells compared with controls in the chimeric mice (Figure 4D).
To compare the development of wild-type NK cells and SHIP-1−/− NK cells within the same environment, we performed a series of mixed bone marrow experiments and adoptively transferred SHIP-1−/− or SHIP-1+/− (CD45.1+) bone marrow at a 1:1 ratio with wild-type competitor (CD45.2+) bone marrow into lethally irradiated wild-type (CD45.2+) recipients (Figure 5A). In agreement with the chimeric mice, we found that the frequency and total number of NK cells were similar between SHIP-1−/− and its competitor as well as between SHIP-1+/− and its competitor (Figure 5B). Similarly to the chimeric mice, the SHIP-1−/− NK cells also display a decrease in CD11b SP NK cells and exhibit an accumulation of CD27 SP NK cells compared with wild-type competitors in mixed bone marrow experiments (Figure 5C). In agreement with these findings, KLRG1 expression is reduced on NK cells derived from SHIP-1−/− bone marrow (supplemental Figure 5A). Therefore, SHIP-1 is critical for NK cell transition to the final stages of NK cell differentiation (stage 5 and 6 as recently categorized33). Notably, when splenocytes derived from mixed bone marrow chimeras were transferred in Rag2−/− × Il2rγ−/− mice, the phenotype of the SHIP-1−/− NK cells parallels the phenotype of the donors and remains impaired in maturation (supplemental Figure 6A). Taken together, these results demonstrate that SHIP-1 acts at 2 levels. First, SHIP-1 expression is required in trans for normal NK cell number. Second, SHIP-1 expression is required in cis for NK cells to reach the final stage of differentiation.
Figure 5.
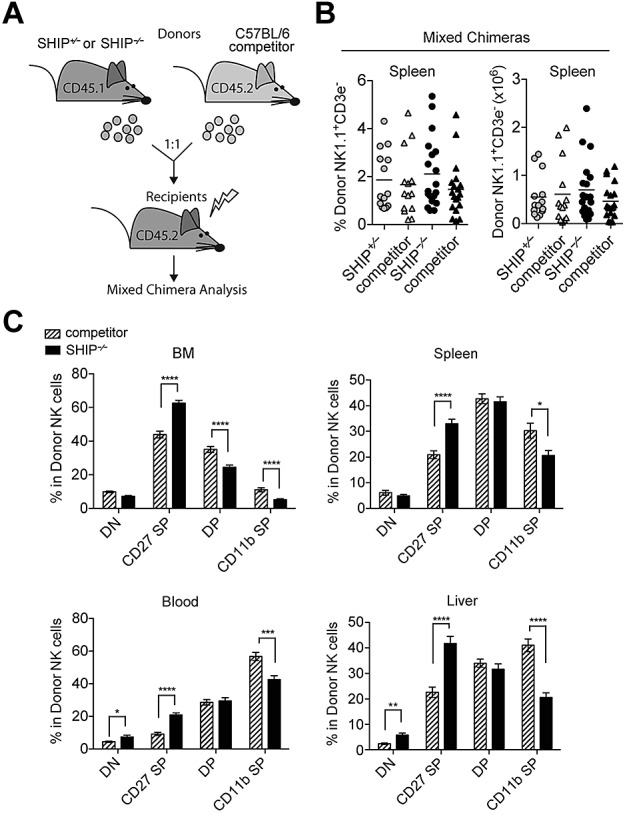
SHIP-1 regulates NK cell final stage of maturation intrinsically. (A) Diagram of mixed chimera generation. SHIP+/− or SHIP−/− (CD45.1+) bone marrow cells were isolated and mixed at an equal ratio with wild-type (WT) competitor (CD45.2+) bone marrow cells before transferring into lethally irradiated WT (CD45.2+) recipients and analyzed 8 to 10 weeks after reconstitution. (B) Percentages and total NK cells (NK1.1+CD3− in the lymphocyte gate) present in the indicated organs from SHIP+/− mixed chimeras (gray circles for SHIP+/− and gray triangles for competitor) and SHIP−/− mixed chimeras (black circles for SHIP−/− and black triangles for competitor). (C) Percentages in NK cells (NK1.1+CD3− in the lymphocyte gate) expressing DN (CD27−CD11b−), CD27 SP (CD11b−CD27+), DP (CD27+CD11b+), and CD11b SP (CD27−CD11b+) populations in the indicated organs derived from SHIP−/− mixed chimeras (black bars for SHIP−/− and striped bars for competitor). Data are pooled from at least 5 independent experiments. Error bars indicate SEM.
To examine whether the impaired functional properties of SHIP-1−/− NK cells are intrinsic, splenic NK cells from bone marrow chimera (not shown) and mixed bone marrow chimeras were stimulated with various conditions and levels of IFN-γ and CD107α were measured. In contrast to NK cells from SHIP-1−/− mice, in the mixed bone marrow chimeric mice cell surface expression of NK1.1, NKG2D, and Ly49H on SHIP-1−/− NK cells is comparable with controls (supplemental Figure 5B). We found no difference in degranulation (monitored by CD107α surface expression) in all the conditions tested (Figure 6A). Levels of IFN-γ activated by NK receptor cross-linking were not significantly affected by the SHIP-1 deficiency (Figure 6A-B left). However, IL-12 and IL-18 stimulation resulted in reduced levels of IFN-γ for SHIP-1−/− NK cells compared with its control in both chimera (not shown) and mixed chimera mice (Figure 6A). Similarly, when chimeric mice were primed in vivo for 24 hours with poly(I:C) only IL-12 and IL-18 stimulation resulted in reduced levels of IFN-γ for SHIP-1−/− NK cells compared with wild-type NK cells (Figure 6B). Interestingly, the reduced IFN-γ was reproducibly decreased at all stages of maturation and significantly at the DP and CD11b SP stages (supplemental Figure 6B). Altogether, the data suggest SHIP-1−/− NK cell cytokine receptor–mediated responses are impaired, whereas ITAM containing receptor–mediated responses are not affected.
Figure 6.
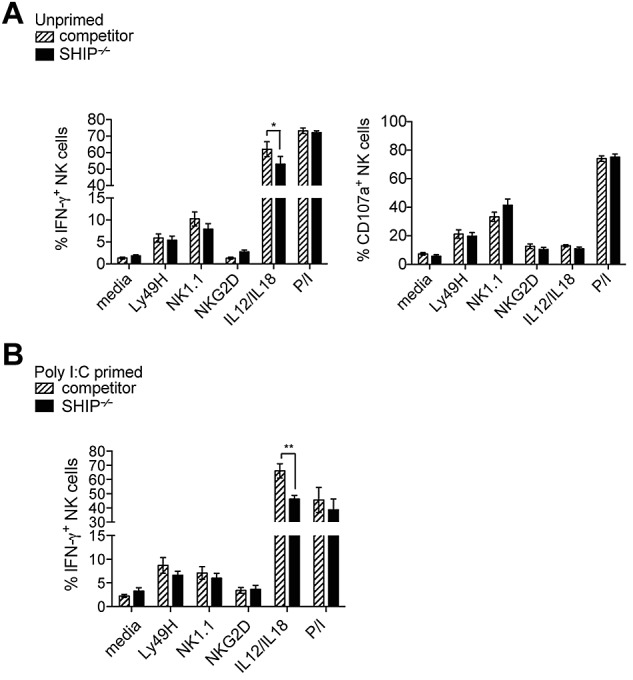
Impaired cytokine receptor–mediated response from SHIP-1–deficient NK cells is cell intrinsic. (A-B) Intracellular IFN-γ and CD107α in NK cell populations (NK1.1+CD3− or DX5+CD3− in the lymphocyte gate) from total splenocytes incubated with antibody-coated plates using anti-Ly49H, anti-NK1.1, and anti-NKG2D or with a mix of IL-12/IL-18 and PMA and ionomycin (P/I) for 6 hours are accessed after gating on donor cells (CD45.1+ for SHIP−/−; CD45.1− for competitor). (A) Mixed chimeras are pooled from at least 4 experiments. (B) Poly(I:C) primed mixed chimeras are pooled from at least 3 experiments. Error bars indicate SEM.
Discussion
Development and maturation processes are integral in defining NK cell effector functions. The process of NK cell development and the acquisition of cell surface receptors is dependent on multiple factors, including cell intrinsic signals and environmental cues.1,34,35 An intact bone marrow microenvironment is required for the development of fully functional mature NK cells.36,37 In addition, cell intrinsic signals, including transcription factors, such as E4BP4 program the development and differentiation of NK cells.38,39
Lipid phosphatases such as SHIP-1 have been recognized as potent negative regulators of immune cells.40,41 The effect of SHIP-1 on NK cell development and functions has not been defined. Using mice with germ line deletion of SHIP-1,14 we found a decrease in bone marrow NK cell precursors and a loss of peripheral NK cells. The reduced cellularity in the NK compartment is reminiscent of the reduced cellularity observed in B and T cells in SHIP-1−/− mice.20 The absence of B cells in these mice has been explained not as a primary defect, but as a consequence of macrophage dysregulation.21 Similarly, SHIP-1−/− mice display reduced T-cell frequency20,22; however, conditional deletion of SHIP-1 in T cells showed normal T-cell numbers and development.23 Interestingly, Collazo et al have shown that SHIP-1 deficiency promotes increased Treg numbers and FoxP3 acquisition by naive T cells in both a T-lineage intrinsic and extrinsic manner, with the extrinsic effect on Treg formation mediated by SHIP-1–deficient myeloid cells.22 Using chimeric mice we demonstrated that the reduced NK cellularity is cell extrinsic and is likely dependent on the expression of SHIP-1 on stromal cells. However, we found that cell intrinsic SHIP-1 expression is critical for NK cells to reach the final stage of maturation. In mixed bone marrow chimeric mice, SHIP-1−/−–deficient NK cells are less mature than wild-type NK cells, demonstrating that SHIP-1 regulates NK cell maturation intrinsically. Peripheral SHIP-1–deficient NK cells are impaired in their transition to CD11b-positive cells. This coupled with a normal Ly49 repertoire and MHC class I environment in mixed bone marrow chimeric mice suggests that NK cell education is not a factor in this case. However, it is possible that other factors besides MHC-Ly49 interactions are influenced by the absence of SHIP-1. For instance, 2B4/CD48 interaction has been shown to influence NK cell homeostasis.24 Regardless of the mechanism, our data reveal both an indirect role and direct role for SHIP-1 at different checkpoints of NK cell development.
Our results also reveal that the decreased functionality of SHIP-1−/− NK cells can be rescued in chimeric mice except when stimulated with IL-12 and IL-18. In this context, our data show that SHIP-1−/− NK cell IFN-γ production is decreased at all stages of maturation, suggesting it is because of an intrinsic defect and not uniquely to a maturation phenotype.5,28 However, SHIP-1 has been shown to be a negative regulator of IFN-γ in human NK cell subsets when stimulated with IL-12 and IL-1842 due presumably to the induction of the microRNA miR-155, which leads to SHIP-1 down regulation.43 Therefore, it is likely that the SHIP-1 inhibitory functions are dissociated from the SHIP-1 developmental function in NK cells and that impaired NK cell development is dominant over the inhibitory functions. Alternatively in the absence of SHIP-1, miR-155 may target other molecules that regulate cytokine receptor signaling. This will be addressed using inducible SHIP-1–deficient mice. SAP adaptors could also play a role as it has been shown that IL-12 induces SAP up-regulation.44 In the absence of SAP adaptors, it has been shown that SLAM-family receptors become inhibitory possibly through an association with SHIP-1, SHP-1, and SHP-2.45 It is therefore possible that in the absence of SHIP-1, SHP-1, or SHP-2 association with SLAM-family receptors lead to a decreased activation.
Interestingly, the presence of the widely expressed SHIP-2 and other SHIP-1 isoforms, such as s-SHIP46 on hematopoietic cells of SHIP-1−/− mice may also participate in the function of these immune cells. Although encoded by a separate gene, SHIP-2 has the same structure as SHIP-1 and like SHIP-1, hydrolyzes the 5-phosphatase from phosphatidylinositol-3,4,5-trisphosphate.47,48 The role of SHIP-2 in B and T cells has not been well defined and whether SHIP-2 is expressed in NK cells has yet to be determined and warrants future investigation. In addition, it was recently reported that ENU-induced mutant SHIP-1 mice have a more drastic phenotype than that observed for SHIP-1−/− mice. This observation was attributed to the mutation affecting both SHIP-1 and its isoform s-SHIP.49 However, s-SHIP protein expression was not detected on FACS sorted NK cells.41 Thus, further experiments evaluating the roles of s-SHIP and SHIP-2 in regulating NK cells are necessary.
Understanding the mechanisms used by NK cells to secrete cytokines as well as induce cytotoxicity is important in NK cell therapy against diseased cells. Indeed, further insights on NK cell phosphatases and how they regulate NK cell development, maturation, and thereby effector functions are needed. Other phosphatases such as CD45 seem to regulate cytokine secretion downstream of ITAM receptors but not cytokine receptors suggesting that regulation of NK cell effector functions is mediated by different phosphatases.50 Phosphatases such as PTEN, SHP-1, and SHP-2 are also ubiquitously expressed in NK cells and their specific role in NK cell development and differentiation has been overlooked. Therefore, the effect of these phosphatases on NK cell development and maturation warrants further investigation.
Supplementary Material
Acknowledgments
The authors thank S. Terrizzi for cell sorting and Céline Fugère for intravenous injections.
This work was supported by National Institutes of Health grant AI58181 (L.B.) and National Institutes of Health National Research Service Award F31 0AI080230 (C.B.).
Footnotes
The online version of the article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Authorship
Contribution: C.B. conceived, performed, and analyzed the experiments and wrote the paper; S.M.S.M. conceived, performed, and analyzed the experiments; W.G.K. provided reagents and wrote the paper; and L.B. conceived, and analyzed the experiments and wrote the paper.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Laurent Brossay, Department of Molecular Microbiology and Immunology, Division of Biology and Medicine, Box G-B579, Brown University, Providence, RI 02912; e-mail: laurent_brossay@brown.edu.
References
- 1.Di Santo JP. Natural killer cell developmental pathways: a question of balance. Annu Rev Immunol. 2006;24:257–286. doi: 10.1146/annurev.immunol.24.021605.090700. [DOI] [PubMed] [Google Scholar]
- 2.Chiossone L, Chaix J, Fuseri N, Roth C, Vivier E, Walzer T. Maturation of mouse NK cells is a 4-stage developmental program. Blood. 2009;113(22):5488–5496. doi: 10.1182/blood-2008-10-187179. [DOI] [PubMed] [Google Scholar]
- 3.Lodoen MB, Lanier LL. Natural killer cells as an initial defense against pathogens. Curr Opin Immunol. 2006;18(4):391–398. doi: 10.1016/j.coi.2006.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bottino C, Moretta L, Moretta A. NK cell activating receptors and tumor recognition in humans. Curr Top Microbiol Immunol. 2006;298:175–182. doi: 10.1007/3-540-27743-9_9. [DOI] [PubMed] [Google Scholar]
- 5.Huntington ND, Tabarias H, Fairfax K, et al. NK cell maturation and peripheral homeostasis is associated with KLRG1 up-regulation. J Immunol. 2007;178(8):4764–4770. doi: 10.4049/jimmunol.178.8.4764. [DOI] [PubMed] [Google Scholar]
- 6.Kim S, Iizuka K, Kang HS, et al. In vivo developmental stages in murine natural killer cell maturation. Nat Immunol. 2002;3(6):523–528. doi: 10.1038/ni796. [DOI] [PubMed] [Google Scholar]
- 7.Rosmaraki EE, Douagi I, Roth C, Colucci F, Cumano A, Di Santo JP. Identification of committed NK cell progenitors in adult murine bone marrow. Eur J Immunol. 2001;31(6):1900–1909. doi: 10.1002/1521-4141(200106)31:6<1900::aid-immu1900>3.0.co;2-m. [DOI] [PubMed] [Google Scholar]
- 8.Dustin ML, Long EO. Cytotoxic immunological synapses. Immunol Rev. 2010;235(1):24–34. doi: 10.1111/j.0105-2896.2010.00904.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lanier LL. DAP10- and DAP12-associated receptors in innate immunity. Immunol Rev. 2009;227(1):150–160. doi: 10.1111/j.1600-065X.2008.00720.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lanier LL. Natural killer cell receptor signaling. Curr Opin Immunol. 2003;15(3):308–314. doi: 10.1016/s0952-7915(03)00039-6. [DOI] [PubMed] [Google Scholar]
- 11.Ravetch JV, Lanier LL. Immune inhibitory receptors. Science. 2000;290(5489):84–89. doi: 10.1126/science.290.5489.84. [DOI] [PubMed] [Google Scholar]
- 12.Eissmann P, Beauchamp L, Wooters J, Tilton JC, Long EO, Watzl C. Molecular basis for positive and negative signaling by the natural killer cell receptor 2B4 (CD244). Blood. 2005;105(12):4722–4729. doi: 10.1182/blood-2004-09-3796. [DOI] [PubMed] [Google Scholar]
- 13.Tessmer MS, Fugere C, Stevenaert F, et al. KLRG1 binds cadherins and preferentially associates with SHIP-1. Int Immunol. 2007;19(4):391–400. doi: 10.1093/intimm/dxm004. [DOI] [PubMed] [Google Scholar]
- 14.Wang JW, Howson JM, Ghansah T, et al. Influence of SHIP on the NK repertoire and allogeneic bone marrow transplantation. Science. 2002;295(5562):2094–2097. doi: 10.1126/science.1068438. [DOI] [PubMed] [Google Scholar]
- 15.Ware MD, Rosten P, Damen JE, Liu L, Humphries RK, Krystal G. Cloning and characterization of human SHIP, the 145-kD inositol 5-phosphatase that associates with SHC after cytokine stimulation. Blood. 1996;88(8):2833–2840. [PubMed] [Google Scholar]
- 16.Liu Q, Shalaby F, Jones J, Bouchard D, Dumont DJ. The SH2-containing inositol polyphosphate 5-phosphatase, ship, is expressed during hematopoiesis and spermatogenesis. Blood. 1998;91(8):2753–2759. [PubMed] [Google Scholar]
- 17.Geier SJ, Algate PA, Carlberg K, et al. The human SHIP gene is differentially expressed in cell lineages of the bone marrow and blood. Blood. 1997;89(6):1876–1885. [PubMed] [Google Scholar]
- 18.Hazen AL, Smith MJ, Desponts C, Winter O, Moser K, Kerr WG. SHIP is required for a functional hematopoietic stem cell niche. Blood. 2009;113(13):2924–2933. doi: 10.1182/blood-2008-02-138008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wahle JA, Paraiso KH, Kendig RD, et al. Inappropriate recruitment and activity by the Src homology region 2 domain-containing phosphatase 1 (SHP1) is responsible for receptor dominance in the SHIP-deficient NK cell. J Immunol. 2007;179(12):8009–8015. doi: 10.4049/jimmunol.179.12.8009. [DOI] [PubMed] [Google Scholar]
- 20.Helgason CD, Damen JE, Rosten P, et al. Targeted disruption of SHIP leads to hemopoietic perturbations, lung pathology, and a shortened life span. Genes Dev. 1998;12(11):1610–1620. doi: 10.1101/gad.12.11.1610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Karlsson MC, Guinamard R, Bolland S, Sankala M, Steinman RM, Ravetch JV. Macrophages control the retention and trafficking of B lymphocytes in the splenic marginal zone. J Exp Med. 2003;198(2):333–340. doi: 10.1084/jem.20030684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Collazo MM, Wood D, Paraiso KH, et al. SHIP limits immunoregulatory capacity in the T-cell compartment. Blood. 2009;113(13):2934–2944. doi: 10.1182/blood-2008-09-181164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tarasenko T, Kole HK, Chi AW, Mentink-Kane MM, Wynn TA, Bolland S. T cell-specific deletion of the inositol phosphatase SHIP reveals its role in regulating Th1/Th2 and cytotoxic responses. Proc Natl Acad Sci U S A. 2007;104(27):11382–11387. doi: 10.1073/pnas.0704853104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fortenbery NR, Paraiso KH, Taniguchi M, Brooks C, Ibrahim L, Kerr WG. SHIP influences signals from CD48 and MHC class I ligands that regulate NK cell homeostasis, effector function, and repertoire formation. J Immunol. 2010;184(9):5065–5074. doi: 10.4049/jimmunol.0901862. [DOI] [PubMed] [Google Scholar]
- 25.Wahle JA, Paraiso KH, Costello AL, Goll EL, Sentman CL, Kerr WG. Cutting edge: dominance by an MHC-independent inhibitory receptor compromises NK killing of complex targets. J Immunol. 2006;176(12):7165–7169. doi: 10.4049/jimmunol.176.12.7165. [DOI] [PubMed] [Google Scholar]
- 26.Huntington ND, Vosshenrich CA, Di Santo JP. Developmental pathways that generate natural-killer-cell diversity in mice and humans. Nat Rev Immunol. 2007;7(9):703–714. doi: 10.1038/nri2154. [DOI] [PubMed] [Google Scholar]
- 27.Robbins SH, Tessmer MS, Mikayama T, Brossay L. Expansion and contraction of the NK cell compartment in response to murine cytomegalovirus infection. J Immunol. 2004;173(1):259–266. doi: 10.4049/jimmunol.173.1.259. [DOI] [PubMed] [Google Scholar]
- 28.Hayakawa Y, Smyth MJ. CD27 dissects mature NK cells into two subsets with distinct responsiveness and migratory capacity. J Immunol. 2006;176(3):1517–1524. doi: 10.4049/jimmunol.176.3.1517. [DOI] [PubMed] [Google Scholar]
- 29.Fernandez NC, Treiner E, Vance RE, Jamieson AM, Lemieux S, Raulet DH. A subset of natural killer cells achieves self-tolerance without expressing inhibitory receptors specific for self-MHC molecules. Blood. 2005;105(11):4416–4423. doi: 10.1182/blood-2004-08-3156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kim S, Poursine-Laurent J, Truscott SM, et al. Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature. 2005;436(7051):709–713. doi: 10.1038/nature03847. [DOI] [PubMed] [Google Scholar]
- 31.Carotta S, Pang SH, Nutt SL, Belz GT. Identification of the earliest NK-cell precursor in the mouse BM. Blood. 2011;117(20):5449–5452. doi: 10.1182/blood-2010-11-318956. [DOI] [PubMed] [Google Scholar]
- 32.Fathman JW, Bhattacharya D, Inlay MA, Seita J, Karsunky H, Weissman IL. Identification of the earliest natural killer cell-committed progenitor in murine bone marrow. Blood. 2011;118(20):5439–5447. doi: 10.1182/blood-2011-04-348912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Narni-Mancinelli E, Vivier E. NK cell genesis: a trick of the trail. Immunity. 2012;36(1):1–3. doi: 10.1016/j.immuni.2012.01.001. [DOI] [PubMed] [Google Scholar]
- 34.Martín-Fontecha A, Lord GM, Brady HJ. Transcriptional control of natural killer cell differentiation and function. Cell Mol Life Sci. 2011;68(21):3495–3503. doi: 10.1007/s00018-011-0800-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sun JC, Lanier LL. NK cell development, homeostasis and function: parallels with CD8 T cells. Nat Rev Immunol. 2011;11(10):645–657. doi: 10.1038/nri3044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Elliott JM, Wahle JA, Yokoyama WM. MHC class I-deficient natural killer cells acquire a licensed phenotype after transfer into an MHC class I-sufficient environment. J Exp Med. 2010;207(10):2073–2079. doi: 10.1084/jem.20100986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Joncker NT, Shifrin N, Delebecque F, Raulet DH. Mature natural killer cells reset their responsiveness when exposed to an altered MHC environment. J Exp Med. 2010;207(10):2065–2072. doi: 10.1084/jem.20100570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gascoyne DM, Long E, Veiga-Fernandes H, et al. The basic leucine zipper transcription factor E4BP4 is essential for natural killer cell development. Nat Immunol. 2009;10(10):1118–1124. doi: 10.1038/ni.1787. [DOI] [PubMed] [Google Scholar]
- 39.Kamizono S, Duncan GS, Seidel MG, et al. Nfil3/E4bp4 is required for the development and maturation of NK cells in vivo. J Exp Med. 2009;206(13):2977–2986. doi: 10.1084/jem.20092176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Veillette A, Latour S, Davidson D. Negative regulation of immunoreceptor signaling. Annu Rev Immunol. 2002;20:669–707. doi: 10.1146/annurev.immunol.20.081501.130710. [DOI] [PubMed] [Google Scholar]
- 41.Kerr WG, Colucci F. Inositol phospholipid signaling and the biology of natural killer cells. J Innate Immun. 2011;3(3):249–257. doi: 10.1159/000323920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Trotta R, Parihar R, Yu J, et al. Differential expression of SHIP1 in CD56bright and CD56dim NK cells provides a molecular basis for distinct functional responses to monokine costimulation. Blood. 2005;105(8):3011–3018. doi: 10.1182/blood-2004-10-4072. [DOI] [PubMed] [Google Scholar]
- 43.Trotta R, Chen L, Ciarlariello D, et al. MiR-155 regulates IFN-gamma production in natural killer cells. Blood. 2012;119(15):3478–3485. doi: 10.1182/blood-2011-12-398099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Endt J, Eissmann P, Hoffmann SC, Meinke S, Giese T, Watzl C. Modulation of 2B4 (CD244) activity and regulated SAP expression in human NK cells. Eur J Immunol. 2007;37(1):193–198. doi: 10.1002/eji.200636341. [DOI] [PubMed] [Google Scholar]
- 45.Dong Z, Cruz-Munoz ME, Zhong MC, Chen R, Latour S, Veillette A. Essential function for SAP family adaptors in the surveillance of hematopoietic cells by natural killer cells. Nat Immunol. 2009;10(9):973–980. doi: 10.1038/ni.1763. [DOI] [PubMed] [Google Scholar]
- 46.Tu Z, Ninos JM, Ma Z, et al. Embryonic and hematopoietic stem cells express a novel SH2-containing inositol 5′-phosphatase isoform that partners with the Grb2 adapter protein. Blood. 2001;98(7):2028–2038. doi: 10.1182/blood.v98.7.2028. [DOI] [PubMed] [Google Scholar]
- 47.Dyson JM, O'Malley CJ, Becanovic J, et al. The SH2-containing inositol polyphosphate 5-phosphatase, SHIP-2, binds filamin and regulates submembraneous actin. J Cell Biol. 2001;155(6):1065–1079. doi: 10.1083/jcb.200104005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Pesesse X, Moreau C, Drayer AL, Woscholski R, Parker P, Erneux C. The SH2 domain containing inositol 5-phosphatase SHIP2 displays phosphatidylinositol 3,4,5-trisphosphate and inositol 1,3,4,5-tetrakisphosphate 5-phosphatase activity. FEBS Lett. 1998;437(3):301–303. doi: 10.1016/s0014-5793(98)01255-1. [DOI] [PubMed] [Google Scholar]
- 49.Nguyen NY, Maxwell MJ, Ooms LM, et al. An ENU-induced mouse mutant of SHIP1 reveals a critical role of the stem cell isoform for suppression of macrophage activation. Blood. 2011;117(20):5362–5371. doi: 10.1182/blood-2011-01-331041. [DOI] [PubMed] [Google Scholar]
- 50.Huntington ND, Xu Y, Nutt SL, Tarlinton DM. A requirement for CD45 distinguishes Ly49D-mediated cytokine and chemokine production from killing in primary natural killer cells. J Exp Med. 2005;201(9):1421–1433. doi: 10.1084/jem.20042294. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.