

Abstract
The hyperglycemia triggers several chronic diabetic complications mediated by increased oxidative stress that eventually causes diabetic nephropathy. The aim of this study was to examine if the sodium-glucose cotransporter (SGLT2) inhibition prevents the oxidative stress in the kidney of diabetic rats. Methods. The diabetic rat model was established by intraperitoneal injection of streptozotocin (50 mg/kg). The inhibition of SGLT2 was induced by daily subcutaneous administration of phlorizin (0.4 g/kg). Oxidative stress was assessed by catalase (CAT), glutathione peroxidase (GPx), and superoxide dismutase (SOD) activities and by immunohistochemical analysis of 3-nitrotyrosine (3-NT). Results. Streptozotocin-induced diabetes caused hyperglycemia and lower body weight. The CAT activity decreased in cortex and medulla from diabetic rats; in contrast, the GPx activity increased. Furthermore the 3-NT staining of kidney from diabetic rats increased compared to control rats. The inhibition of SGLT2 decreased hyperglycemia. However, significant diuresis and glucosuria remain in diabetic rats. The phlorizin treatment restores the CAT and GPX activities and decreases 3-NT staining. Conclusion. The inhibition of SGLT2 by phlorizin prevents the hyperglycemia and oxidative stress in kidney of diabetic rats, suggesting a prooxidative mechanism related to SGLT2 activity.
1. Introduction
Diabetic nephropathy (DN) is a leading cause of end-stage renal failure, accounting for 35% to 40% of all new cases that require dialysis therapy worldwide. Clinical studies have demonstrated that hyperglycemia is an important causal factor that mediates the development and progression of diabetic kidney disease [1]. Several evidences suggest that the controlling of glucose in the renal proximal tubular could play a major role in the genesis of diabetic systemic complications [2, 3].
On the other hand it has been shown that increases in glucose uptake in the kidney during diabetes could lead to high intracellular glucose levels, inducing an enhanced production of reactive oxygen species (ROS), which probably can be amplified by the decreased capacity of the cellular antioxidant defense system in this condition [4, 5]. Recently, we have reported increased sodium glucose cotransporter (SGLT2) activity and expression in rats with diabetes and salt sensitivity [6, 7]. Also, we showed that the increase in oxidative stress and SGLT2 expression was prevented by ursodeoxycholic acid treatment in diabetic rats [8].
The excessive production of ROS has been suggested as a common outcome from the pathways leading to increased oxidative damage, which culminates in DN [5]. Under thisconditioncompounds which are able to diminish or modulate hyperglycemia and oxidative stress could be a plausible target to delay the onset of DN.
The SGLT2 inhibition by phlorizin exerts a hypoglycemic effect in diabetic rodents via induction of glycosuria [9] and prevents elevated blood glucose levels in Zucker diabetic fatty rats [10]. The renal glucose transport inhibition induced by phlorizin treatment leads to glucose plasma levels close to the normal values without significant alteration in insulin plasma levels. These observations suggest that SGLT2 inhibitors may be effective agents to normalize high blood glucose levels, which are associated with diseases such as type 1 and type 2 diabetes mellitus [11].
In the present study we hypothesized that increased SGLT2 activity may contribute to hyperglycemic chronic state leading to the development of oxidative stress during diabetes. Therefore, the present study was undertaken to assess if phlorizin as a sodium glucose cotransporter inhibitor could be effective reducing the alterations in the oxidative stress in kidney in streptozotocin-induced diabetic rat model.
2. Methods
2.1. Reagents
Streptozotocin (STZ), phlorizin, xanthine, nitroblue tetrazolium (NBT), BSA, xanthine oxidase, NADPH, glutathione reductase (GR), and reduced glutathione (GSH) were purchased from Sigma (St. Louis, MO, USA). Rabbit anti-3-nitrotyrosine polyclonal antibodies were from Upstate, Lake Placid, NY, USA. All other chemicals used were of the highest analytical grade available.
2.2. Experimental Design
Male adult Wistar rats (10–14 weeks of age) weighing 250–300 g were used. Animals were randomly divided into four groups: control (C), diabetic (D), diabetic treated with phlorizin (DP), and diabetic treated with insulin (DI). Diabetes was induced by a single administration of STZ (50 mg/kg i.p.) dissolved in citrate buffer (0.1 M, pH 4.5). Control group received the same volume of citrate buffer.
After 72 h of STZ administration, the blood glucose concentration was determined (Accu-Chek sensor comfort, Roche Diagnostics), and only rats having over 20.0 mmol/L were considered diabetic for further studies.
Phlorizin was administered (0.4 g/kg/day/4 weeks, s.c. in a 20% of propylene glycol solution) subcutaneous injection in two equally divided doses that were given at 12 h intervals to ensure continuous inhibition of renal tubular glucose reabsorption.
Insulin treatment was administered i.p. (Humulin; Eli Lilly and Company, Indianapolis, IN). Insulin was given in initial dose of 6 IU followed by 8 to 10 IU depending on morning blood glucose values.
After diabetes confirmation, the treatments started and were maintained during 30 days. All experimental groups were maintained on laboratory diet and water ad libitum.
Thirty days after STZ administration, urine samples (24 h) were collected in metabolic cages; the urinary variables measures were diuresis and glucose (IL 300 plus, clinical chemistry analyzer). Later, all the animals were anesthetized with sodium pentobarbital (50 mg/kg). The kidneys were perfused and rapidly removed. The cortex and medulla were isolated and submerged in liquid nitrogen before enzyme activity measurements. Some kidneys were placed in paraformaldehyde 4% for nitrosative stress analysis, measured by immunohistochemical analysis of 3-nitrotyrosine modified proteins.
All animal procedures were performed in accordance with the Mexican Federal Regulation for Animal Experimentation and Care (NOM-062-ZOO-2001) and were approved by the Bioethics and Investigation Committees of the Instituto Nacional de Cardiología “Ignacio Chávez.”
2.3. Evaluation of Oxidative Stress
2.3.1. Preparation of Renal Tissue
Cortex and medulla were washed thoroughly with ice sold saline, 10% (w/v); each tissue was homogenized separately in a Potter Elvehjem homogenizer in ice-cold 50 mM phosphate buffer pH 7.4 containing mammalian protease inhibitor cocktail. The homogenates were centrifuged at 10,000 ×g for 30 min at 4°C. The supernatants were used for measuring the enzyme activities. A portion of the supernatant was used for the determination of total protein concentration by Bradford method [12] using bovine serum albumin as standard.
2.3.2. Catalase Assay
Renal catalase (CAT) activity was assayed at 25°C by Aebi method which is based on the disappearance of H2O2 from a solution containing 30 mmol/L H2O2 in 10 mmol/L potassium phosphate buffer (pH 7) at 240 nm [13]. The results were expressed as U/mL/min.
2.3.3. Glutathione Peroxidase Assay
Renal glutathione peroxidase (GPx) activity was assessed by a method previously described [14]. The results were expressed as U/mL/min.
2.3.4. Superoxide Dismutase Assay
Superoxide dismutase (SOD) activity in kidney homogenates was measured by a competitive inhibition assay using xanthine-xanthine oxidase system to reduce NBT, a previously reported method [15]. Results were expressed as U/mg protein.
2.3.5. Immunohistochemical Localization of 3-Nitrotyrosine (3-NT)
For immunohistochemistry, paraffin-embedded kidney tissues were sectioned (5 μm) and transferred to positively charged slides. Samples were treated with H2O2 (4.5%) to quench/inhibit endogenous peroxidase. After blocking, the sections were reacted with anti-3-NT antibody (Upstate Biotechnology Inc., Lake Placid, NY) for 1 h at room temperature. After extensive washing with PBS, the sections were incubated with antibody peroxidase conjugated for 1 h and finally incubated with diaminobenzidine for 30 min. Quantitative image analysis was performed with image analysis software (Image-Pro Plus 6.0, Media Cybernetics Inc, Bethesda, Maryland, MD, USA). The software determines densitometry mean values of selected tissue regions. Thus, 10 fields/rats were randomly selected, and the intensity of the 3-NT immunostaining was determined.
2.4. Statistics
The data were expressed as the mean ± SEM. Data were analyzed with a nonpaired t-test or with ANOVA followed by multiple comparisons by Bonferroni t-test, when applicable (Prism 3.0; GraphPad Software, San Diego, CA, USA). Statistical significance was assumed at P ≤ 0.05.
3. Results
3.1. Animal Characteristics
STZ-induced diabetes caused significant increases in blood glucose concentration, diuresis and glucosuria and a significant decrease in body weight (Table 1).
Table 1.
Physiological parameters in experimental groups.
| Body weight (g) |
Blood glucose (mg dL−1) |
Diuresis (mL 24 hrs) |
Urinary glucose (mg dL−1 24 hrs) |
|
|---|---|---|---|---|
| Control | 408.8 ± 1.750 | 88.00 ± 5.119 | 16.81 ± 2.113 | 20.00 ± 7.638 |
| Diabetic | 271.2 ± 4.964* | 527.4 ± 8.834* | 44.88 ± 4.569* | 1748 ± 176.6* |
| Diabetic phlorizin | 316.3 ± 5.609∗† | 133.2 ± 7.929∗† | 55.33 ± 6.015* | 1920 ± 179.7* |
| Diabetic insulin | 408.5 ± 9.192∗† | 118.0 ± 10.18∗† | 16.10 ± 2.578† | 71.67 ± 36.71∗† |
Data are mean ± SEM of 8 animals in each group. *P < 0.05 versus control, † P < 0.05 versus D.
After 30 days of treatment in diabetic rats, phlorizin reduced blood glucose levels; however, significant diuresis, glucosuria, and low body weight remain (Table 1). The insulin treatment in diabetic rats induced a significant increase in body weight compared to nontreated diabetic rats. Moreover, insulin treatment completely reversed hyperglycemia, diuresis, and glucosuria (Table 1).
3.2. Evaluation of Oxidative Stress
3.2.1. Renal Activity of Antioxidant Enzymes
The antioxidant enzyme CAT, GPx, and total SOD activities were determined in the cortex and medulla of diabetic, phlorizin-treated, insulin-treated rats and control group.
Figures 1 to 3 showed that in the cortex and medulla of diabetic rats, CAT activity was decreased (Figures 1(a) and 1(b)), whereas GPx level was increased when compared with the control group (Figures 2(a) and 2(b)). Interestingly the treatment with phlorizin or insulin restores to normal levels CAT and GPX enzyme activities in cortex and medulla (Figures 1 and 2). SOD activity in cortex and medulla was similar in the four groups studied (Figures 3(a) and 3(b)).
Figure 1.
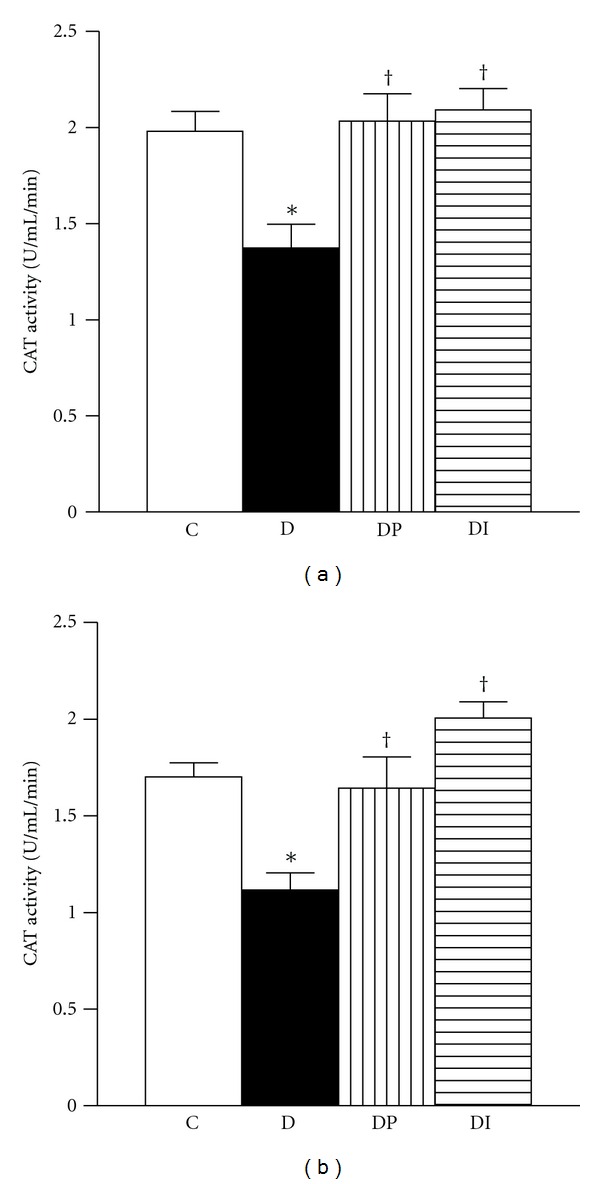
Catalase activity was measured in kidney from control (C), diabetic (D), diabetic treated with phlorizin (DP), and diabetic treated with insulin (DI). (a) cortex and (b) medulla. Data are mean ± SEM of eight animals in each group; *P < 0.05 versus C, † P < 0.05 versus D.
Figure 3.
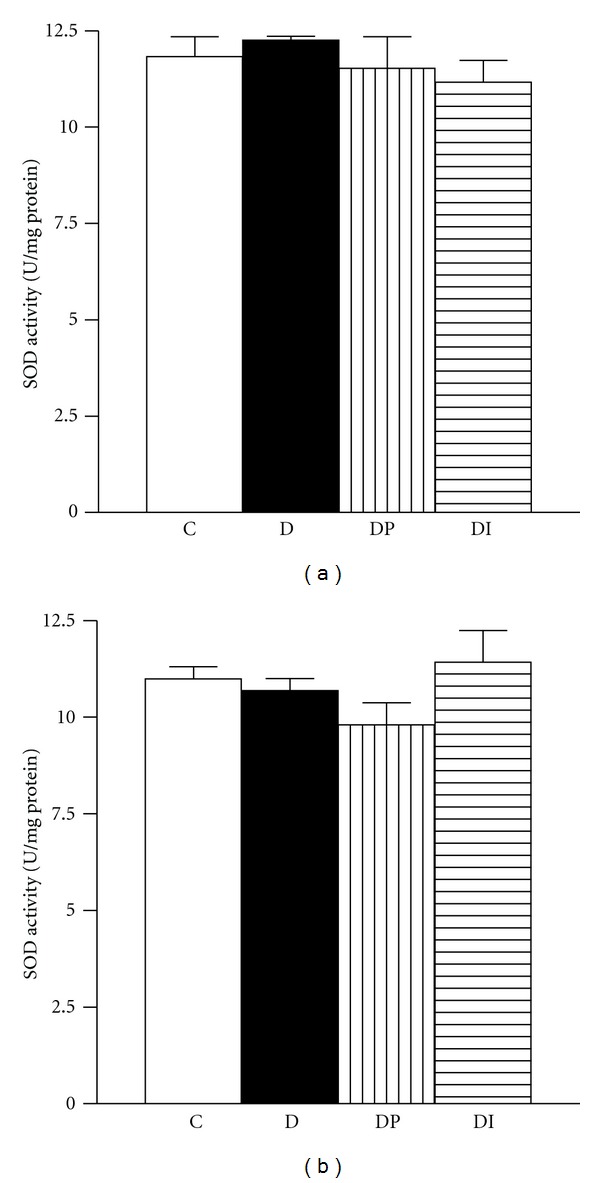
Superoxide dismutase activity was measured in kidney from control (C), diabetic (D), diabetic treated with phlorizin (DP), and diabetic treated with insulin (DI). (a) cortex and (b) medulla. Data are mean ± SEM of eight animals in each group; *P < 0.05 versus C, † P < 0.05 versus D.
Figure 2.
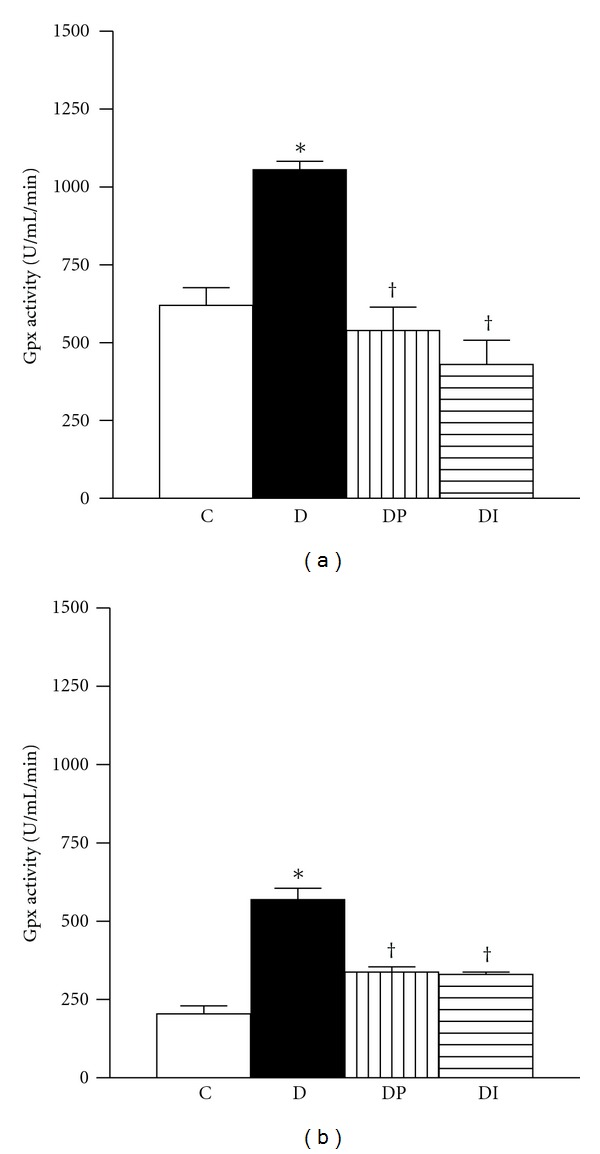
Glutathione peroxidase activity was measured in kidney from control (C), diabetic (D), diabetic treated with phlorizin (DP), and diabetic treated with insulin (DI). (a) cortex and (b) medulla. Data are mean ± SEM of eight animals in each group; *P < 0.05 versus C, † P < 0.05 versus D.
3.2.2. Immunohistochemical Localization of 3-Nitrotyrosine (3-NT)
The 3-NT levels were significantly increased in diabetic rats (Figures 4(a) and 4(b)). The phlorizin or insulin treatments were able to reverse the oxidative damage to normal values. These results indicate that tyrosine nitration of proteins is enhanced in the kidney of diabetic rats, and the blood glucose control was able to prevent these hyperglycemia-induced effects.
Figure 4.
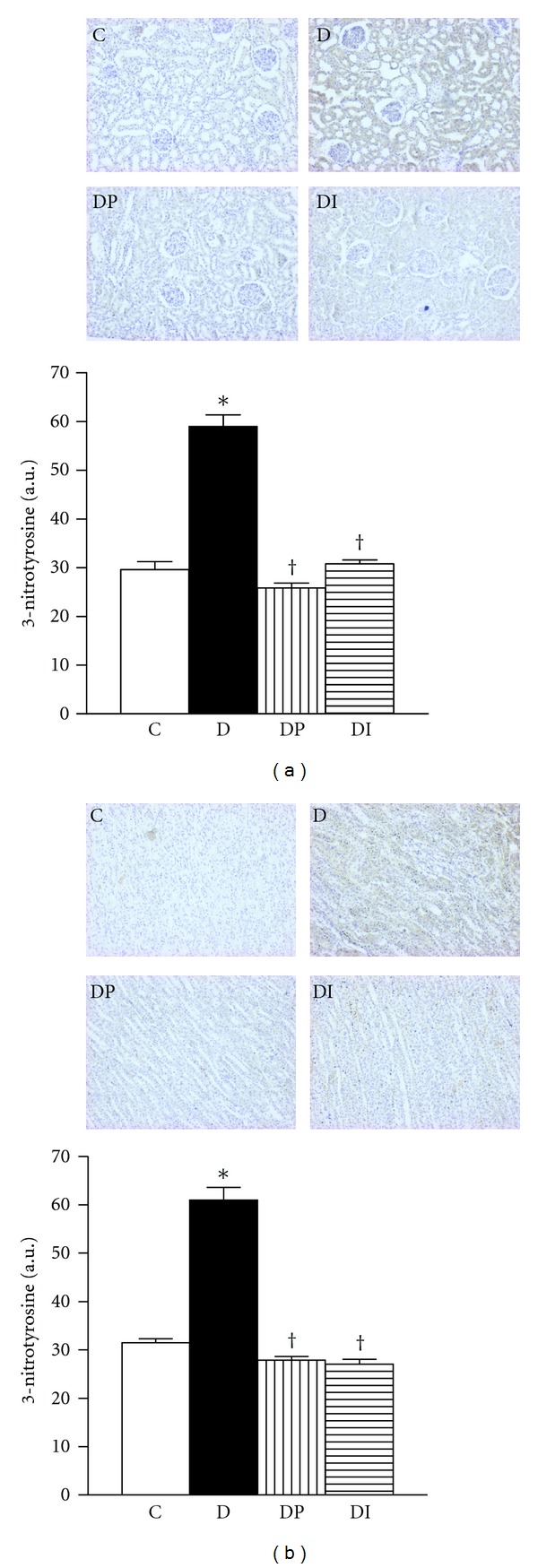
Immunohistochemistry and semiquantitative analysis of nitrotyrosine levels in kidney from control (C), diabetic (D), diabetic treated with phlorizin (DP), and Diabetic treated with insulin (DI). (a) cortex and (b) medulla. Data are mean ± SEM of eight animals in each group; *P < 0.05 versus C, † P < 0.05 versus D.
4. Discussion
In the present study, diabetes induction was followed by significant increases in renal oxidative stress evidenced by low CAT activity, whereas GPx was increased; moreover, diabetic rats showed increase in nitrotyrosine levels in cortex and medulla. Four-week phlorizin treatment restores blood glucose levels, enzymatic activities, and nitrotyrosine levels. These effects were observed without any influence on the metabolic control. Insulin treatment prevents the alterations that are diabetes-induced with normalization of the metabolic derangement unbalance.
Increased formation of ROS and diabetic nephropathy may occur in diabetes possibly associated with increased glucose concentration in plasma, tissues, and renal intracellular glucose levels [2, 16, 17]. High proximal tubular glucose concentration in poorly controlled diabetes may lead to excessive glucose, sodium, and water reabsorption, which could be mediated by enhanced activity of the Na+-glucose cotransporter (SGLT); this might contribute to development of diabetic complications [3, 6–8, 18–20]. The main goal of this study was to investigate the effect of phlorizin (a SGLT2 inhibitor) on oxidative stress in diabetic rats. Therefore, we examined the antioxidant enzymes activity and tyrosine nitration in cortex and medulla from control, diabetic, diabetic treated with phlorizin, and diabetic rats treated with insulin.
One month of STZ-induced diabetes resulted in a reduced CAT activity in cortex and medulla. In contrast, the GPx activity increased in cortex and medulla from diabetic rats to protect cellular and tissue injury. The increase in GPx and the decrease in the CAT activities in the kidney cortex suggest a compensatory mechanism in the different antioxidant enzymes in response to oxidative stress.
Although Cu/Zn SOD mRNA has been shown to be significantly induced in the total kidney of diabetic rats [21], we were not able to find any change in total SOD activity neither cortex or medulla. However, we are not discarding the possibility that diabetes change the activity of Cu-Zn SOD or Mn SOD.
The functional and pathophysiological role of excessive oxidative stress in diabetic kidney disease was indicated by the presence of increased levels of lipid peroxides and 8-hydroxydeoxyguanosine in the kidney of STZ-induced diabetic rats [22, 23]. These observations are lacking in the direct evidence for the presence of oxidative stress in cortex or medulla and might reflect a common consequence of diabetic kidney damage. Here we provide the evidence for the presence of oxidative stress in cortex and medulla by two methods, enzymatic activity measurements and 3-NT immunostaining, which has been used to estimate the formation of nitrogen reactive species.
Peroxynitrite, formed by the reaction between superoxides and nitric oxide (NO), modifies tyrosine in proteins to form nitrotyrosine, and this stable end-product is involved in the inactivation of mitochondrial and cytosolic proteins, resulting in damage of cellular constituents [24]. Our results showed that diabetes is associated with an increased tyrosine nitration in cortex and medulla, which is consistent with the previous studies in patients and experimental models with DM [25–28]. Presently, it is unclear if nitrotyrosine represents only a marker for oxidative stress from NO-induced oxidants or whether it alters protein structure sufficiently to cause abnormal enzyme, receptor, or signaling function. The phlorizin treatment in diabetic animals decreased the nitrotyrosine stain in cortex and medulla, consistent with their effects in preventing hyperglycemia and restoring the antioxidant enzyme activity. These data have also been shown by insulin treatment.
In previous studies, phlorizin treatment of diabetic rats has been shown to normalize the blood glucose levels, blood pressure, proteinuria, and hyperfiltration [7, 11, 29]. Previously it has been reported that phlorizin treatment increases glycosuria [29]; however, in this study we were not able to observe these effects.
In the present study phlorizin and insulin treatments were almost equally effective in their ability to prevent the decrease in CAT activity and increase in GPX activity and nitrotyrosine levels observed in diabetic rats. Our data clearly demonstrate an SGLT2 participation in the development of oxidative stress during diabetes. We hypothesize that phlorizin may inhibit SGLT2 activity in vivo. That means that phlorizin treatment can reduce the glucose reabsorption and therefore hyperglycemia, consequently the potential glycation of enzymes or probably reducing reactive oxygen-free radicals formation improving the activities of antioxidant enzymes.
The results obtained in this approach performed in vitro and in vivo suggest that the SGLT2 inhibition might be used in diabetic patients to alleviate the dysfunctions caused by diabetes and to lower oxidative stress via inhibition of SGLT2 activity.
Previously, we have demonstrated increase in SGLT2 activity and expression in cortex of diabetic and hypertensive rats [6, 7]. It has been suggested that the consequences of the SGLT2 induction not only represent responses designed to adjust the transtubular glucose reabsorption in different filtration rates, but also play an active role in renal injury, such as oxidative stress, diabetic nephropathy, hypertension, and Na+ reabsorption. Thus, SGLT2 inhibitors may be potential antidiabetic agents [9–11, 30–32] with a renoprotective effect by specifically mitigating transcellular epithelial glucose flux [32], which may prevent some of the cellular mechanisms leading to diabetic renal complications [32].
In conclusion, our evidence demonstrated that SGLT2 inhibition prevents oxidative and nitrative stress in kidney of diabetic rats, which was significantly influenced by glycemic control. Furthermore, our data confirm that early continuous aggressive treatment of glycemia is important to avoid future complications.
Conflict of Interests
There are no conflict of interests.
Acknowledgments
This project was supported by the Mexican Council of Science and Technology (CONACYT), Research Grant 155604 to Dr. H. Osorio, and by Fondos del Gasto Directo Autorizado a la Subdirección de Investigación Básica INC “Ignacio Chávez.” Dra. Claudia Rangel-Barajas critically revised the paper.
References
- 1.Shamoon H, Duffy H, Fleischer N, et al. The effect of intensive treatment of diabetes on the development and progression of long-term complications in insulin-dependent diabetes mellitus. New England Journal of Medicine. 1993;329(14):977–986. doi: 10.1056/NEJM199309303291401. [DOI] [PubMed] [Google Scholar]
- 2.Bagby SP. Diabetic nephropathy and proximal tubule ROS: challenging our glomerulocentricity. Kidney International. 2007;71(12):1199–1202. doi: 10.1038/sj.ki.5002286. [DOI] [PubMed] [Google Scholar]
- 3.Vallon V. The proximal tubule in the pathophysiology of the diabetic kidney. The American Journal of Physiology. 2011;300(5):R1009–R1022. doi: 10.1152/ajpregu.00809.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Garg MC, Ojha S, Bansal DD. Antioxidant status of streptozotocin diabetic rats. Indian Journal of Experimental Biology. 1996;34(3):264–266. [PubMed] [Google Scholar]
- 5.Ha H, Lee HB. Reactive oxygen species amplify glucose signalling in renal cells cultured under high glucose and in diabetic kidney. Nephrology. 2005;10(supplement 2):S7–S10. doi: 10.1111/j.1440-1797.2005.00448.x. [DOI] [PubMed] [Google Scholar]
- 6.Osorio H, Bautista R, Rios A, Franco M, Santamaría J, Escalante B. Effect of treatment with losartan on salt sensitivity and SGLT2 expression in hypertensive diabetic rats. Diabetes Research and Clinical Practice. 2009;86(3):e46–e49. doi: 10.1016/j.diabres.2009.09.006. [DOI] [PubMed] [Google Scholar]
- 7.Osorio H, Bautista R, Rios A, et al. Effect of phlorizin on SGLT2 expression in the kidney of diabetic rats. Journal of Nephrology. 2010;23(5):541–546. [PubMed] [Google Scholar]
- 8.Osorio H, Coronel I, Arellano A, Franco M, Escalante B, Bautista R. Ursodeoxycholic acid decreases sodium-glucose cotransporter (SGLT2) expression and oxidative stress in the kidney of diabetic rats. Diabetes Research and Clinical Practice. 2012;97(2):276–282. doi: 10.1016/j.diabres.2012.02.022. [DOI] [PubMed] [Google Scholar]
- 9.Asano T, Anai M, Sakoda H, et al. SGLT as a therapeutic target. Drugs of the Future. 2004;29(5):461–466. [Google Scholar]
- 10.Handlon AL. Sodium glucose co-transporter 2 (SGLT2) inhibitors as potential antidiabetic agents. Expert Opinion on Therapeutic Patents. 2005;15(11):1531–1540. [Google Scholar]
- 11.Janssen SW, Martens GJ, Sweep CG, Span PN, Verhofstad AAJ, Hermus ARMM. Phlorizin treatment prevents the decrease in plasma insulin levels but not the progressive histopathological changes in the pancreatic islets during aging of Zucker diabetic fatty rats. Journal of Endocrinological Investigation. 2003;26(6):508–515. doi: 10.1007/BF03345212. [DOI] [PubMed] [Google Scholar]
- 12.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Analytical Biochemistry. 1976;72(1-2):248–254. doi: 10.1006/abio.1976.9999. [DOI] [PubMed] [Google Scholar]
- 13.Aebi H. Catalase in vitro. Methods in Enzymology. 1984;105:121–126. doi: 10.1016/s0076-6879(84)05016-3. [DOI] [PubMed] [Google Scholar]
- 14.Lawrence RA, Burk RF. Glutathione peroxidase activity in selenium deficient rat liver. Biochemical and Biophysical Research Communications. 1976;71(4):952–958. doi: 10.1016/0006-291x(76)90747-6. [DOI] [PubMed] [Google Scholar]
- 15.Sun Y, Oberley LW, Li Y. A simple method for clinical assay of superoxide dismutase. Clinical Chemistry. 1988;34(3):497–500. [PubMed] [Google Scholar]
- 16.Brownlee M. Biochemistry and molecular cell biology of diabetic complications. Nature. 2001;414(6865):813–820. doi: 10.1038/414813a. [DOI] [PubMed] [Google Scholar]
- 17.Ha H, Kim KH. Pathogenesis of diabetic nephropathy: the role of oxidative stress and protein kinase C. Diabetes Research and Clinical Practice. 1999;45(2-3):147–151. doi: 10.1016/s0168-8227(99)00044-3. [DOI] [PubMed] [Google Scholar]
- 18.Gilbert RE, Cooper ME. The tubulointerstitium in progressive diabetic kidney disease: more than an aftermath of glomerular injury? Kidney International. 1999;56(5):1627–1637. doi: 10.1046/j.1523-1755.1999.00721.x. [DOI] [PubMed] [Google Scholar]
- 19.Bank N, Aynedjian HS. Progressive increases in luminal glucose stimulate proximal sodium absorption in normal and diabetic rats. Journal of Clinical Investigation. 1990;86(1):309–316. doi: 10.1172/JCI114700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Noonan WT, Shapiro VM, Banks RO. Renal glucose reabsorption during hypertonic glucose infusion in female streptozotocin-induced diabetic rats. Life Sciences. 2001;68(26):2967–2977. doi: 10.1016/s0024-3205(01)01090-6. [DOI] [PubMed] [Google Scholar]
- 21.Sechi LA, Ceriello A, Griffin CA, et al. Renal antioxidant enzyme mRNA levels are increased in rats with experimental diabetes mellitus. Diabetologia. 1997;40(1):23–29. doi: 10.1007/s001250050638. [DOI] [PubMed] [Google Scholar]
- 22.Horie K, Miyata T, Maeda K, et al. Immunohistochemical colocalization of glycoxidation products and lipid peroxidation products in diabetic renal glomerular lesions. Implication for glycoxidative stress in the pathogenesis of diabetic nephropathy. Journal of Clinical Investigation. 1997;100(12):2995–3004. doi: 10.1172/JCI119853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ha H, Kim C, Son Y, Chung MH, Kim KH. DNA damage in the kidneys of diabetic rats exhibiting microalbuminuria. Free Radical Biology and Medicine. 1994;16(2):271–274. doi: 10.1016/0891-5849(94)90152-x. [DOI] [PubMed] [Google Scholar]
- 24.Halliwell B. What nitrates tyrosine? Is nitrotyrosine specific as a biomarker of peroxynitrite formation in vivo. FEBS Letters. 1997;411(2-3):157–160. doi: 10.1016/s0014-5793(97)00469-9. [DOI] [PubMed] [Google Scholar]
- 25.Pacher P, Obrosova I, Mabley J, Szabó C. Role of nitrosative stress and peroxynitrite in the pathogenesis of diabetic complications: emerging new therapeutical strategies. Current Medicinal Chemistry. 2005;12(3):267–275. doi: 10.2174/0929867053363207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ishii N, Patel KP, Lane PH, et al. Nitric oxide synthesis and oxidative stress in the renal cortex of rats with diabetes mellitus. Journal of the American Society of Nephrology. 2001;12(8):1630–1639. doi: 10.1681/ASN.V1281630. [DOI] [PubMed] [Google Scholar]
- 27.Onozato ML, Tojo A, Goto A, Fujita T, Wilcox CS. Oxidative stress and nitric oxide synthase in rat diabetic nephropathy: effects of ACEI and ARB. Kidney International. 2002;61(1):186–194. doi: 10.1046/j.1523-1755.2002.00123.x. [DOI] [PubMed] [Google Scholar]
- 28.Thuraisingham RC, Nott CA, Dodd SM, Yaqoob MM. Increased nitrotyrosine staining in kidneys from patients with diabetic nephropathy. Kidney International. 2000;57(5):1968–1972. doi: 10.1046/j.1523-1755.2000.00046.x. [DOI] [PubMed] [Google Scholar]
- 29.Malatiali S, Francis I, Barac NM. Phlorizin prevents glomerular hyperfiltration but not hypertrophy in diabetic rats. Experimental Diabetes Research. 2008;2008:7 pages. doi: 10.1155/2008/305403.305403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Adachi T, Yasuda K, Okamoto Y, et al. T-1095, A renal Na+-glucose transporter inhibitor, improves hyperglycemia in streptozotocin-induced diabetic rats. Metabolism. 2000;49(8):990–995. doi: 10.1053/meta.2000.7729. [DOI] [PubMed] [Google Scholar]
- 31.Oku A, Ueta K, Arakawa K, et al. T-1095, an inhibitor of renal Na+-glucose cotransporters, may provide a novel approach to treating diabetes. Diabetes. 1999;48(9):1794–1800. doi: 10.2337/diabetes.48.9.1794. [DOI] [PubMed] [Google Scholar]
- 32.Arakawa K, Ishihara T, Oku A, et al. Improved diabetic syndrome in C57BL/KsJ-db/db mice by oral administration of the Na+-glucose cotransporter inhibitor T-1095. British Journal of Pharmacology. 2001;132(2):578–586. doi: 10.1038/sj.bjp.0703829. [DOI] [PMC free article] [PubMed] [Google Scholar]