

Abstract
Penetration of 3H-labeled water (3H2O) and the 14C-labeled organic acids benzoic acid ([14C]BA), salicylic acid ([14C]SA), and 2,4-dichlorophenoxyacetic acid ([14C]2,4-D) were measured simultaneously in isolated cuticular membranes of Prunus laurocerasus L., Ginkgo biloba L., and Juglans regia L. For each of the three pairs of compounds (3H2O/[14C]BA, 3H2O/[14C]SA, and 3H2O/[14C]2,4-D) rates of cuticular water penetration were highly correlated with the rates of penetration of the organic acids. Therefore, water and organic acids penetrated the cuticles by the same routes. With the combination 3H2O/[14C]BA, co-permeability was measured with isolated cuticles of nine other plant species. Permeances of 3H2O of all 12 investigated species were highly correlated with the permeances of [14C]BA (r2 = 0.95). Thus, cuticular transpiration can be predicted from BA permeance. The application of this experimental method, together with the established prediction equation, offers the opportunity to answer several important questions about cuticular transport physiology in future investigations.
Leaves of higher plants are covered by a cuticle (Martin and Juniper, 1970), which forms the interface between the leaf interior and the atmosphere (Kerstiens, 1996a). Plant cuticles have multiple functions: they reduce leaching and thus prevent leaves from an excessive loss of ions and nutrients (Scherbatskoy and Tyree, 1990; Tyree et al., 1992), and they also form the first effective barrier to infection by pathogens (Kolattukudy, 1985). The most important function is probably the protection of higher, land-living plants from uncontrolled water loss (Schönherr, 1982). When stomata are closed, the remaining water loss from the leaf is determined by the rates of cuticular transpiration. In situations of severe water stress, desiccation and survival of plants depend to a large extent on cuticular water permeability. In adaptation to this important function, the plant cuticle is a hydrophobic extracellular biopolymer composed of esterified cutin acids (Holloway, 1993) and monomeric cuticular waxes deposited to the cutin polymer (Walton, 1990; Bianchi, 1995). Because of their solid-crystalline aggregation (Sitte and Rennier, 1963; Reynhardt and Riederer, 1991, 1994), cuticular waxes form the main barrier to the penetration of molecules such as water, ions, and organic chemicals across the cuticle (Schönherr and Riederer, 1989; Riederer and Schreiber, 1995; Schreiber et al., 1996).
Diffusion is the mechanism by which a molecule penetrates the cuticle (Bukovac and Petracek, 1993). A compound enters the cuticle as single molecules on one side, diffuses through the cuticle, and exits on the other side. Because plant cuticles are heterogeneous membranes composed of cutin, associated waxes, and carbohydrates, it is often speculated that molecules penetrating across plant cuticles might follow two different paths of diffusion. Small, polar molecules such as water should penetrate the cuticle via polar pores (Franke, 1967; Sekse, 1995), whereas larger, more lipophilic molecules should diffuse through the more lipophilic domain of the hydrophobic cuticle. However, this view of cuticular transport properties is not generally accepted. There is substantial experimental evidence that the plant cuticle behaves toward penetrating molecules like a homogeneous membrane (Schönherr and Riederer, 1989; Kerstiens, 1996b; Schreiber and Riederer, 1996a).
We investigated the simultaneous penetration of 3H2O, representing a small, polar molecule, together with 14C-labeled organic compounds ([14C]BA, [14C]SA, and [14C]2,4-D), representing larger, more lipophilic chemicals, across isolated plant cuticles. It was our aim to determine whether plant cuticles behave like homogeneous membranes. Furthermore, we wanted to establish a method to predict cuticular transpiration from cuticular permeances of organic chemicals.
MATERIALS AND METHODS
Fully expanded leaves of 11 tree species and fruits of tomato (Lycopersicon esculentum Mill.) were sampled from fully grown plants (Table I). Astomatous CMs were isolated enzymatically from the fruits and from the adaxial surfaces of the leaves according to the procedure described by Schönherr and Riederer (1986). CMs were stored at room temperature for more than 3 months before being used for measuring cuticular transport, because Geyer and Schönherr (1990) reported that permeances of isolated cuticles decreased by a factor of about 2 within the first weeks after isolation.
Table I.
Scientific names and origin of plant species used in the permeation experiments
| Leaves
|
Fruits | |
|---|---|---|
| Evergreen species | Deciduous species | |
| Hedera helix L.a | Liriodendron tulipifera L.b | L. esculentum Millc |
| Philodendron selloum (K.) Kochb | Juglans regia L.a | |
| Camellia sinensis (L.) O. Kuntzeb | Ginkgo biloba L.a | |
| Euonymus japonica Thunb.b | Pyrus cf. communis L.a | |
| Monstera deliciosa Liebm.b | ||
| Prunus laurocerasus L.a | ||
| Citrus limon (L.) Burm.b | ||
Botanical garden, Würzburg, Germany.
Botanical garden, Kaiserslautern, Germany.
Local market, Würzburg, Germany.
Radiolabeled Compounds
3H2O and the 14C-labeled organic acids [14C]BA, [14C]SA, and [14C]2,4-D were measured simultaneously in the transport experiments (Table II). The radiochemical purity of the compounds was greater than 98% in all cases. Radiolabeled compounds were dissolved in citric acid (10−2 m) buffered at pH 3.0. Because BA, SA, and 2,4-D are still partially ionized at pH 3.0 (Table II), the driving force used in the transport experiments was corrected for the degree of dissociation of these compounds according to Equation 1 (Fujita et al., 1964):
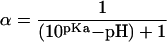 |
1 |
where the degree of dissociation is given by α, pKa represents the acid dissociation constant of the respective compound, and pH is the actual pH value of the donor solution. Methylated SA was synthesized from [14C]SA using diazomethane, which was prepared according to the method of Levitt (1973). Average aqueous concentrations of the 14C-labeled chemicals used in the transport experiments were around 0.1 m.
Table II.
Specific radioactivities (SR) and dissociation constants (DC) of the radiolabeled compounds used in the transport experiments
Co-Permeability Experiments Using Isolated Cuticular Membranes
Co-permeability experiments with isolated cuticular membranes were carried out using the transport chambers described in detail by Schreiber et al. (1995). Isolated cuticles were mounted between the two halves of transport chambers made of stainless steel. The physiological outer surface of the isolated cuticle was oriented toward the donor chamber (1 mL) and the inner surface of the cuticle faced the receiver chamber (1 mL). The interface between the cuticle and the stainless-steel chamber was sealed using high-vacuum silicone grease (Wacker Chemie, Burghausen, Germany).
The buffered, aqueous solution of 3H2O, together with one of the three 14C-labeled organic acids, was added to the donor chamber. Preliminary experiments using 3H- and 14C-labeled standards had shown that isotopic ratios varying between 1:5 and 10:1 were easily separated by dual-label counting. Thus, the ratios of the two differently labeled compounds in the donor solutions were chosen so that the receiver solutions contained the two isotopes in similar amounts. The receiver compartment contained PLS (1% in water; soybean lecithin, Roth, Karlsruhe, Germany; purity 98%). The micelles of the PLS completely solubilized the 14C-labeled organic acids that had diffused across the cuticle, thus keeping the concentration of the penetrated compounds in the receiver at 0 (Bauer and Schönherr, 1992).
Transport chambers prepared in this way were kept in the dark on a rotating bench (60 rpm) at 25°C. After distinct periods of time (1, 2, 3, 4, 5, and 6 h), receiver solutions were completely replaced by fresh PLS. Sampled PLS solutions were mixed with scintillation cocktail (Ultima Gold XR, Canberra Packard, Zurich, Switzerland) and the amounts of both differentially labeled compounds, 3H2O as well as the 14C-labeled organic acids, were simultaneously determined by liquid-scintillation counting (model 1409, Wallac counter, Turku, Finland). The amounts of radioactivity that had penetrated across each single cuticle were added for each sampling point. A plot of the penetrated amounts of radioactivity versus the respective sampling time resulted in linear transport kinetics. The slopes of the regression lines fitted to the transport kinetics corresponded to the flow (in mol s−1 m−2) of the respective radioactive compounds (3H2O and 14C-labeled organic acids) across the isolated cuticle. Permeances (m s−1) relating flow to the respective concentrations (Δc) (mol m−3) used in the experiments were calculated according to Equation 2:
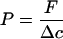 |
2 |
where P is permeance (m s−1), a measure of the barrier properties of a plant cuticle toward a certain compound. It can be used to compare barrier properties of different cuticles toward the same compound or of the same cuticle toward different compounds (Kerler et al., 1984). p, the permeability coefficient (m2 s−1), which is often used in transport physiology, can be calculated by multiplying P by the thickness of the transport-limiting barrier (Becker et al., 1986). However, the thickness of the transport-limiting barrier of plant cuticles, which is made up of cuticular waxes, is not known and can only be estimated. Thus, to analyze cuticular transport, the use of the permeance is preferred, because it does not contain an assumption about the thickness of the transport-limiting barrier of the cuticle.
Sample Size and Statistics
Permeance of isolated cuticular membranes was measured for at least 15 replicates. Results are given as means with 95% confidence intervals. Before carrying out parametric statistical procedures, permeances were subjected to a logarithmic transformation. It was shown recently (Baur, 1997) and also observed in this study (Fig. 1) that permeances measured with isolated cuticles follow a log-normal distribution. Statistical calculations were performed using SPSS for Windows (version 5.0).
Figure 1.

Distribution of untransformed (A) and log-transformed (B) cuticular water permeances of isolated cuticular membranes obtained from P. laurocerasus. Bars represent the measured permeance (P; in m s−1) for water, whereas the solid lines give the Gaussian distributions fitted to the measured permeance values. The insets show the probability plots calculated from the measured permeance values.
RESULTS
The distribution of water permeances measured with Prunus laurocerasus CMs showed a tailing on the right side (Fig. 1A), and the coefficient of determination of the probability plot was low (r2 = 0.84; Fig. 1A, inset). Plotting the frequency of the water permeances as a function of the logarithms of the permeance, a log-normal distribution was obtained (Fig. 1B) with a highly significant correlation of the probability plot (r2 = 0.99; Fig. 1B, inset). The significance of the log-normal distribution of water permeances was confirmed by statistical analysis (Kolmogoroff-Smirnow test; Sokal and Rohlf, 1995).
In P. laurocerasus the permeance of [14C]SA (when calculated without correction for dissociation) was strongly dependent on the pH of the donor solution. It decreased from 1.6 × 10−9 m s−1 at pH 1.0 to 0.069 × 10−9 m s−1 at pH 7.0 (Fig. 2). The permeance of methylated SA, which was independent at pH values ranging from 3.0 to 7.0, was about 2.0 × 10−9 m s−1 and constantly decreased at pH values less than 3.0 (Fig. 2). At pH 3.0, the amounts of 3H2O, which had penetrated across all 15 investigated cuticles of J. regia at all six sampling times (1, 2, 3, 4, 5, and 6 h), were highly correlated with the amounts of the respective 14C-labeled organic acid measured simultaneously in co-permeability experiments (Fig. 3).
Figure 2.

Permeances (P; in m s−1) of [14C]SA and methylated [14C]SA as a function of the pH of the donor solution. Error bars represent 95% confidence intervals.
Figure 3.

Correlations obtained in co-permeability experiments across isolated cuticles of J. regia investigating the simultaneous penetration of 3H2O together with one of the three 14C-labeled organic acids ([14C]BA, [14C]SA, and [14C]2,4-D). Data points represent the amounts of labeled compounds that had diffused across each of the 15 investigated cuticles at each sampling time (1, 2, 3, 4, 5, and 6 h). Coefficients of determination were 0.99 with the substance pairs 3H2O/[14C]BA and 3H2O/[14C]SA and 0.95 with the substance pair 3H2O/[14C]2,4-D.
Co-permeability of 3H2O and the three 14C-labeled organic acids was measured with the three species P. laurocerasus, G. biloba, and J. regia (Table III). Cuticular permeances of the three 14C-labeled organic acids increased in the order BA, SA, and 2,4-D. The lowest permeances were always observed with P. laurocerasus, whereas J. regia and G. biloba had significantly higher values. Using 3H2O and [14C]BA, co-permeability was measured for isolated cuticular membranes of nine other plant species (Table III). Permeances of 3H2O ranged from 0.13 × 10−9 m s−1 for H. helix to 9.31 × 10−9 m s−1 for C. limon. Permeances of [14C]BA ranged from 0.18 × 10−9 m s−1 for H. helix to 13.7 × 10−9 m s−1 for L. esculentum.
Table III.
Permeances (P) of 14C-labeled organic acids and 3H2O in isolated cuticular membranes of the 12 investigated plant species
| Species | 14C-Labeled Acid | 3H2O |
|---|---|---|
| P × 10−9 m s−1 | ||
| Co-permeability of 3H2O and [14C]2,4-D | ||
| P. laurocerasus | 2.71 (1.60–4.60) | 0.51 (0.32–0.81) |
| G. biloba | 5.30 (3.19–8.80) | 0.56 (0.31–1.00) |
| J. regia | 8.19 (5.93–11.3) | 3.85 (2.72–5.44) |
| Co-permeability of 3H2O and [14C]SA | ||
| P. laurocerasus | 1.53 (0.88–2.66) | 0.95 (0.57–1.57) |
| G. biloba | 4.91 (2.64–9.14) | 0.58 (0.33–1.03) |
| J. regia | 5.31 (3.43–8.22) | 7.15 (4.02–12.7) |
| Co-permeability of 3H2O and [14C]BA | ||
| H. helix | 0.18 (0.10–0.31) | 0.13 (0.07–0.23) |
| P. selloum | 0.22 (0.14–0.34) | 0.24 (0.14–0.42) |
| C. sinensis | 0.24 (0.13–0.43) | 0.20 (0.12–0.32) |
| E. japonica | 0.55 (0.36–0.83) | 0.50 (0.28–0.91) |
| M. deliciosa | 0.60 (0.38–0.95) | 0.55 (0.35–0.87) |
| P. laurocerasus | 0.78 (0.34–1.39) | 0.78 (0.60–1.51) |
| L. tulipifera | 2.04 (1.26–3.31) | 1.88 (1.07–3.34) |
| J. regia | 4.31 (2.78–6.68) | 4.02 (2.47–6.23) |
| G. biloba | 4.67 (2.88–7.57) | 1.55 (0.82–2.59) |
| P. cf. communis | 9.26 (6.46–13.4) | 3.55 (2.14–5.89) |
| C. limon | 10.40 (6.45–16.9) | 9.31 (5.37–16.2) |
| L. esculentum | 13.70 (8.91–21.2) | 6.60 (3.89–11.21) |
Permeances are given as means with 95% confidence intervals in parentheses.
DISCUSSION
In several previous studies it was shown that permeances of isolated cuticular membranes were not usually normally distributed but instead possessed a pronounced tailing on the right side of the distribution (Geyer and Schönherr, 1990; Schreiber and Riederer, 1996b). Recently, it was shown that cuticular permeances of water and organic acids follow a log-normal instead of a simple-normal distribution (Baur, 1997). Similar results were observed in this study. A plot of the frequencies of measured water permeances with P. laurocerasus cuticles exhibited a pronounced tailing on the right side, and the probability plot indicated that the distribution was not properly described by a Gaussian distribution (Fig. 1A). However, the logarithms of permeance were very well described by a Gaussian distribution, and the probability plot was highly significant (Fig. 1B), indicating the existence of a log-normal distribution as expected from the recent investigation by Baur (1997). Similar results were obtained for the distribution of BA, SA, and 2,4-D permeances across isolated P. laurocerasus cuticles (data not shown). Thus, further processing of measured permeances was carried out on the basis of a log-normal distribution. Calculation of means and further statistical processing (calculation of confidence intervals) of the permeances was done with the logarithms of permeance (Table III).
The organic acids investigated had dissociation constants varying from 2.73 for 2,4-D to 4.2 for BA (Table II). Thus, aqueous donor solutions had to be buffered at pH 3.0 to ensure that substantial amounts of the 14C-labeled acids were in the uncharged, nondissociated form, which preferentially penetrates across the isolated cuticle. This is shown with the pH-dependent permeance of SA across P. laurocerasus cuticles, which was measured in the pH range of 1.0 to 7.0 (Fig. 2). The decrease of the apparent permeance with increasing pH closely resembles a titration curve (Fig. 2). With the pH rising, the number of ionized SA molecules increases and the apparent permeance decreases. The inflection point of the curve is at pH 3.0, corresponding to the pKa value of SA (Table II). Therefore, apparent permeances measured with the three organic acids investigated were corrected for their degree of dissociation at pH 3.0 (Table III).
Plant cuticles are polyelectrolytes, such as carboxyl groups, carrying fixed charges (Schönherr and Bukovac, 1973). For this reason it cannot be ruled out a priori that different external pH values could indirectly influence cuticular transport properties by changing the charge density within the cutin polymer. Therefore, experiments measuring cuticular permeance at different pH values were repeated using methylated SA, which is no longer ionizable (Fig. 2). At pH values varying from 3.0 to 7.0, cuticular permeance of methylated SA was independent of the pH. This allows the conclusion that cuticular transport properties are not significantly altered in the pH range 3.0 to 7.0, although the charge density of the cutin polymer itself is changed in this pH range. At pH values less than 3.0, the apparent permeance of methylated SA significantly decreased (Fig. 2). However, this may be interpreted as an artifact, because preliminary experiments showed (data not shown) that the methylester of SA was not very stable at acidic pH values. At pH values significantly less than 3.0, methylated SA was probably hydrolyzed again, leading to decreasing apparent permeances (Fig. 2).
The permeance of 3H2O across isolated cuticular membranes of J. regia was highly correlated with simultaneously measured permeances of [14C]BA, [14C]SA, and [14C]2,4-D (Fig. 3), and similar results were obtained with G. biloba and P. laurocerasus (Table III). From these results conclusions can be drawn about possible paths of diffusion across the cuticle, if certain assumptions are taken into account. Because permeances of cuticles sampled from the same species always showed a high degree of variability (Geyer and Schönherr, 1990; Schreiber et al., 1995; Schreiber and Riederer, 1996b; Baur, 1997; Kirsch et al., 1997), it is surprising that with each of the three substance combinations, all of the 15 isolated cuticles fell into the same correlation (Fig. 3). If there had been two different routes of diffusion, a more polar route for water and a more lipophilic route for the acids, as is postulated in the literature (Franke, 1967; Sekse, 1995), one would expect to obtain 15 individual correlations corresponding to the 15 investigated cuticles. Each correlation should represent the respective ratio of polar versus apolar diffusional routes across each investigated cuticle. However, this was not the case (Fig. 3). Therefore, we conclude from our data that plant cuticles behave like homogeneous membranes toward penetrating molecules of different physicochemical properties. There is no indication of the existence of polar pores transversing the plant cuticle and serving as a preferential penetration site for water but not for the investigated organic acids.
The conclusions drawn from this study are in accordance with those from a series of other investigations on cuticular permeability, which correlated the transport of water with the transport of organic molecules (Schönherr and Riederer, 1989; Schreiber and Riederer, 1996a). However, it must be added that under certain conditions our results can also be interpreted in favor of two different sites of cuticular penetration. If the ratios between putative apolar and polar transport routes across the cuticles were identical for each cuticle, the same result depicted in Figure 3 would have been obtained. However, in view of the large variability of cuticular permeances this assumption is not very realistic.
To avoid confusion it must be mentioned at this point that permeances measured recently with cuticles of the same species were all lower (Schreiber and Riederer, 1996a) compared with the permeances measured in this study (Table III). We interpret this difference to be an effect of humidity on cuticular permeance. Recently, water permeances (Schreiber and Riederer, 1996a) were measured by mounting isolated cuticles on transpiration chambers. The physiological inner side of the cuticle faced the water-filled compartment of the transpiration chamber, representing the leaf interior. The physiological outer side of the cuticle faced the atmosphere with 0% humidity, because transpiration chambers were stored over silica gel. Water permeance was obtained by measuring the loss of weight from the transpiration chambers with increasing time.
This experimental design represents the extreme case for cuticular transpiration, because maximum driving forces for water are present. In the study presented here, self-diffusion of 3H2O across an isolated cuticle facing aqueous solutions on both sides was measured. Cuticles were exposed to 100% humidity on both sides and minimum driving forces were present. Because all permeances measured with 3H2O were higher compared with those measured with the gravimetric system, we suggest that cuticular permeability might increase with increasing air humidity, a theory currently under investigation in our laboratory.
Further co-permeability experiments using the substance pair 3H2O/[14C]BA and isolated cuticles of nine additional plant species also showed strong correlations between cuticular water permeances and cuticular permeances of BA (Table III). This provides further substantial evidence for the conclusions drawn above concerning the homogeneous structure of plant cuticles as transport barriers. Furthermore, a double-logarithmic plot of 3H2O permeances as a function of [14C]BA permeances measured with isolated cuticles of the same species resulted in a linear correlation (r2 = 0.95; Fig. 4). Therefore, on the basis of this correlation it is possible to establish a predictive equation that allows estimation of cuticular transpiration of other species from the cuticular permeance of BA of the respective species with high accuracy:
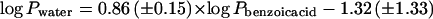 |
3 |
This predictive equation will be of great advantage for investigating a series of different questions concerning cuticular transport physiology. Until now it was not possible to obtain reliable values of cuticular water permeance of stomatous leaf surfaces. Furthermore, in many plant species cuticles cannot be isolated or stomata do occur on both sides of the leaves, making the accurate determination of cuticular water permeance impossible. However, by the method described here cuticular transpiration can be estimated. Because of their high surface tension, aqueous solutions will not enter stomatal pores (Schönherr and Bukovac, 1972). Cuticular permeance of BA, which is not volatile and therefore cannot enter the leaf through the gas phase via open stomata, can be measured and cuticular transpiration can be calculated by applying Equation 3.
Figure 4.

Double-logarithmic plot showing simultaneous penetration of the substance pair 3H2O/[14C]BA through isolated cuticular membranes from 12 different plant species. Error bars represent 95% confidence intervals. Parameters of the linear regression equation are: log Pwater = 0.86 (± 0.15) × log Pbenzoic acid − 1.32 (± 1.33); r2 = 0.95.
One of the most prominent questions concerns the residual transpiration of intact leaves when stomata are closed. Usually, the remaining amount of water loss from the leaf surface is thought to represent cuticular transpiration (Kerstiens, 1996b). However, there are several observations indicating that residual transpiration is still determined to a large extent by water loss of incompletely closed stomata (Grace, 1990). Other interesting results supporting this conclusion come from the comparison of cuticular transpiration of astomatous, isolated cuticles with residual transpiration of intact leaves when stomata are assumed to be completely closed.
Usually, permeances measured with isolated, astomatous cuticular membranes are significantly lower compared with permeances obtained from experiments investigating intact leaves with closed stomata (Kerstiens, 1996b). Nevertheless, this problem still awaits a convincing solution. This should now be possible by applying the experimental procedures described here in combination with recently published data, which demonstrated that BA permeances measured with intact leaves and isolated cuticles are identical (Kirsch et al., 1997). Consequently, BA permeances measured with intact leaf surfaces, even with leaf surfaces carrying stomata, will allow the estimation of cuticular transpiration from Equation 3. Even if it turns out that cuticular permeance depends on air humidity, as indicated above, the application of this method still represents an important step toward an improved experimental analysis of cuticular permeability by comparing stomatous versus astomatous leaf surfaces under identical experimental conditions (100% air humidity).
Co-permeability experiments, as they are described here, give valuable insight into cuticular transport mechanisms and offer the possibility of estimating cuticular water permeance from permeances of organic chemicals. Future experiments using co-permeability can be used to investigate the effects of temperature on cuticular permeability. The occurrence of phase transitions at temperatures above 313 K, leading to substantially increased permeances of water, has been described (Schönherr et al., 1979). On the basis of co-permeability experiments it will be possible to decide whether the observed phase transitions in plant cuticles will also affect permeances of larger organic chemicals. Furthermore, this method might also be used in investigating the effect of decreased leaf water potentials on cuticular water permeability, as was recently described by Boyer et al. (1997).
ACKNOWLEDGMENT
The authors are indebted to Dr. Gerhard Kerstiens (Institute of Environmental and Biological Sciences, Lancaster University, UK) for critically reviewing the manuscript.
Abbreviations:
- BA
benzoic acid
- CM
isolated cuticular membrane
- 3H2O
3H-labeled water
- PLS
phospholipid suspension
- SA
salicylic acid
Footnotes
This work was supported by the Deutsche Forschungsgemeinschaft (DFG-Projekt SCHR 506/2-2 and SFB 251).
LITERATURE CITED
- Albert A, Serjeant EP (1962) The Determination of Ionization Constants. Wiley, New York
- Bauer H, Schönherr J. Determination of mobilities of organic compounds in plant cuticles and correlation with molar volumes. Pestic Sci. 1992;35:1–11. [Google Scholar]
- Baur P. Lognormal distribution of water permeability and organic solute mobility in plant cuticles. Plant Cell Environ. 1997;20:167–177. [Google Scholar]
- Becker M, Kerstiens G, Schönherr J. Water permeability of plant cuticles: permeance, diffusion and partition coefficients. Trees. 1986;1:54–60. [Google Scholar]
- Bianchi G (1995) Plant waxes. In RJ Hamilton, ed, Waxes: Chemistry, Molecular Biology and Functions. The Oily Press, Dundee, UK, pp 175–222
- Boyer JS, Wong SC, Farquhar D. CO2 and water vapor exchange across leaf cuticle (epidermis) at various water potentials. Plant Physiol. 1997;114:185–191. doi: 10.1104/pp.114.1.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bukovac MJ, Petracek PD. Characterizing pesticide and surfactant penetration with isolated plant cuticles. Pestic Sci. 1993;37:179–194. [Google Scholar]
- Franke W. Mechanisms of foliar penetration of solutions. Annu Rev Plant Physiol. 1967;18:181–300. [Google Scholar]
- Fujita T, Iwasa J, Hansch C. A new substituent constant π derived from partition coefficients. J Am Chem Soc. 1964;86:5175–5179. [Google Scholar]
- Geyer U, Schönherr J. The effect of the environment on permeability and composition of Citrus leaf cuticles. 1. Water permeability of isolated cuticular membranes. Planta. 1990;180:147–153. doi: 10.1007/BF00193989. [DOI] [PubMed] [Google Scholar]
- Grace J. Cuticular water loss unlikely to explain tree-line in Scotland. Oecologia. 1990;84:64–68. doi: 10.1007/BF00665596. [DOI] [PubMed] [Google Scholar]
- Holloway PJ. Structure and chemistry of plant cuticles. Pestic Sci. 1993;37:203–232. [Google Scholar]
- Kerler F, Riederer M, Schönherr J. Non-electrolyte permeability of plant cuticles: a critical evaluation of experimental methods. Physiol Plant. 1984;62:599–602. [Google Scholar]
- Kerstiens G (1996a) Diffusion of water vapour and gases across cuticles and through stomatal pores presumed closed. In G Kerstiens, ed, Plant Cuticles: An Integrated Functional Approach. BIOS Scientific Publishers, Oxford, UK, pp 212–234
- Kerstiens G. Cuticular water permeability and its physiological significance. J Exp Bot. 1996b;47:1813–1832. [Google Scholar]
- Kirsch T, Kaffarnik F, Riederer M, Schreiber L. Cuticular permeability of the three tree species Prunus lauocerasus L., Ginkgo biloba L. and Juglans regia L.: comparative investigation of the transport properties of intact leaves, isolated cuticles and reconstituted cuticular waxes. J Exp Bot. 1997;48:1035–1045. [Google Scholar]
- Kolattukudy PE. Enzymatic penetration of the plant cuticle by fungal pathogens. Annu Rev Phytopathol. 1985;23:223–250. [Google Scholar]
- Levitt MJ. Rapid methylation of micro amounts of nonvolatile acids. Ann Chem. 1973;45:618–620. [Google Scholar]
- Martin JT, Juniper BE. The Cuticles of Plants. London: Edward Arnold; 1970. [Google Scholar]
- Reynhardt EC, Riederer M. Structure and molecular dynamics of the cuticular wax from leaves of Citrus aurantium L. J Phys D Appl Phys. 1991;24:478–486. [Google Scholar]
- Reynhardt EC, Riederer M. Structure and molecular dynamics of plant waxes. II Cuticular waxes from leaves of Fagus sylvatica L. and Hordeum vulgare L. Eur Biophys J. 1994;23:236–247. [Google Scholar]
- Riederer M, Schreiber L (1995) Waxes: the transport barriers of plant cuticles. In RJ Hamilton, ed, Waxes: Chemistry, Molecular Biology and Functions. The Oily Press, Dundee, UK, pp 131–156
- Rippen G. Handbuch der Umweltchemikalien. Germany: Ecomed, Landsberg; 1992. [Google Scholar]
- Scherbatskoy T, Tyree MT. Kinetics of exchange of ions between artificial precipitation and maple leaf surfaces. New Phytol. 1990;114:703–712. [Google Scholar]
- Schönherr J. Resistance of plant surfaces to water loss: transport properties of cutin, suberin and associated lipids. In: Lange OL, Nobel PS, Osmond CB, Ziegler H, editors. Encyclopedia of Plant Physiology, New Series, Vol 12B. Berlin: Springer; 1982. pp. 153–179. [Google Scholar]
- Schönherr J, Bukovac MJ. Penetration of stomata by liquids. Plant Physiol. 1972;49:813–819. doi: 10.1104/pp.49.5.813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schönherr J, Bukovac MJ. Ion exchange properties of isolated tomato fruit cuticular membrane: exchange capacity, nature of fixed charges and cation selectivity. Planta. 1973;109:73–93. doi: 10.1007/BF00385454. [DOI] [PubMed] [Google Scholar]
- Schönherr J, Eckl K, Gruler H. Water permeability of plant cuticles: the effect of temperature on diffusion of water. Planta. 1979;147:21–26. doi: 10.1007/BF00384585. [DOI] [PubMed] [Google Scholar]
- Schönherr J, Riederer M. Plant cuticles sorb lipophilic compounds during enzymatic isolation. Plant Cell Environ. 1986;9:459–466. [Google Scholar]
- Schönherr J, Riederer M. Foliar penetration and accumulation of organic chemicals in plant cuticles. Rev Environ Contam Toxicol. 1989;108:1–70. [Google Scholar]
- Schreiber L, Bach S, Kirsch T, Knoll D, Schalz K, Riederer M. A simple photometric device analysing cuticular transport physiology: surfactant effect on permeability of isolated cuticular membranes of Prunus laurocerasus L. J Exp Bot. 1995;46:1915–1921. [Google Scholar]
- Schreiber L, Kirsch T, Riederer M (1996) Diffusion through cuticles: principles and models. In G Kerstiens, ed, Plant Cuticles: An Integrated Functional Approach. BIOS Scientific Publishers, Oxford, UK, pp 109–120
- Schreiber L, Riederer M. Determination of diffusion coefficients of octadecanoic acid in isolated cuticular waxes and their relationship to cuticular water permeabilities. Plant Cell Environ. 1996a;19:1075–1082. [Google Scholar]
- Schreiber L, Riederer M. Ecophysiology of cuticular transpiration: comparative investigation of cuticular water permeability of plant species from different habitats. Oecologia. 1996b;107:426–432. doi: 10.1007/BF00333931. [DOI] [PubMed] [Google Scholar]
- Sekse L. Fruit cracking in sweet cherries (Prunus avium L). Some physiological aspects: a mini review. Sci Hortic. 1995;63:135–141. [Google Scholar]
- Sitte P, Rennier R. Untersuchungen an cuticularen Zellwandschichten. Planta. 1963;60:19–40. [Google Scholar]
- Sokal RR, Rohlf FJ (1995) Biometry: The Principles and Practice of Statistics in Biological Research. Freeman, New York
- Tyree MT, Wescott CR, Tabor CA, Morse AD. Diffusion and electric mobility of KCl within isolated cuticles of Citrus aurantium. Plant Physiol. 1992;99:1057–1061. doi: 10.1104/pp.99.3.1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walton TJ. Waxes, cutin and suberin. Methods Plant Biochem. 1990;4:105–158. [Google Scholar]