

Abstract
Stimulation of peripheral cranial nerves has been shown to exert anticonvulsant effects in animal models as well as in human patients. Specifically, stimulation of both the trigeminal and vagus nerves has been shown in multiple clinical trials to be anticonvulsant, and stimulation of these nerves at therapeutic levels does not cause pain or negatively affect brain function. However, the neuronal mechanisms by which such stimulation exerts therapeutic effects are not well understood. In this review, the possible locations of action for trigeminal nerve stimulation (TNS) and vagus nerve stimulation (VNS) are explored. Additionally, the multiple time scales on which TNS and VNS function are discussed.
Keywords: Anticonvulsant mechanisms, epilepsy, trigeminal nerve stimulation, vagus nerve stimulation
INTRODUCTION
There exist multiple types of direct or indirect neuro-stimulation methods for therapeutically altering brain activity. Such techniques include stimulating the brain indirectly, as with transcranial magnetic stimulation, stimulating the brain directly, as with deep brain stimulation, or affecting the brain indirectly via stimulation of peripheral nerves. These techniques have been used to treat a range of disorders, including epilepsy. However, little is understood about the neurobiological mechanisms by which any of these stimulation techniques affect brain function, and why or how they can act in a therapeutic manner.
One technique utilizing therapeutic neurostimulation for epilepsy is activation of peripheral cranial nerves. A relatively new therapeutic technique involves stimulating the trigeminal nerve. The trigeminal is the fifth cranial nerve, whose branches can be accessed for stimulation both surgically and transcutaneously. Trigeminal nerve stimulation (TNS) has been shown to reduce seizures in both animal models [27] and in a series of clinical trials with epilepsy patients. [23,24,60] Additionally, TNS has been shown in a preliminary clinical trial to be effective against symptoms of depression. [67]
A similar treatment, vagus nerve stimulation (VNS), was approved by the Food and Drug Administration (FDA) for the treatment of epilepsy in 1997, and has been utilized in tens of thousands of patients. [15] The vagus nerve is the tenth cranial nerve and is stimulated in VNS patients via a nerve cuff electrode surgically implanted in the neck, with leads to a stimulation device implanted near the clavicle (Beekwilder and Beems [7] and Lulic et al. [48] for review). VNS has been applied not only in epilepsy patients, but is also efficacious against depression and is being tested for migraine, Alzheimer's disease, and eating disorders. [7] In addition, it has also been demonstrated that stimulation of the ninth cranial nerve, the glossophyrangeal, can reduce seizures. [59]
The studies of and clinical experience with TNS and VNS demonstrate that these neurostimulation paradigms can exert robust therapeutic effects without deleterious effects on normal brain function. Furthermore, TNS and VNS have been demonstrated to control multiple seizure types across multiple species (e.g., rat, dog, non-human primates, humans), suggesting that these anticonvulsant effects involve widespread neuromodulatory mechanisms that are well-conserved phylogenetically.
However, the mechanism by which stimulation of peripheral cranial nerves can exert central therapeutic effects, including affecting the propensity for seizures, is essentially an open question. This is due in part to a lack of knowledge about the feedforward and feedback effects of such stimulation throughout the central nervous system, and in part to a lack of understanding of the pathological mechanisms that underlie disorders such as epilepsy. This is complicated by the fact that epilepsy is not a disorder with homogenous etiology or symptoms.
This review will discuss what is known about mechanisms that might underlie the anticonvulsant effects of cranial nerve stimulation. First, the anatomy of the trigeminal and vagus nerves will be described briefly, and then hypotheses of mechanisms of action will be addressed by moving systematically up the anatomical axis. In addition to discussing potential anatomical locations of action, this article will address the multiple timeframes on which therapeutic cranial nerve stimulation can affect the central nervous system.
It should be noted that the putative mechanisms discussed here are not mutually exclusive; they may, in fact, interact with and complement one another during treatment. Furthermore, different mechanisms might be optimally evoked by specific stimulus parameters and may be uniquely suited for treating specific types of seizures or other disorders.
Overview of trigeminal and vagus nerve anatomy
The trigeminal nerve is responsible for sending afferent sensory information from the face to the central nervous system [Figure 1]. Aptly named, it has three branches that project from different regions of the face to the trigeminal ganglion near the base of the skull. Afferent projections from the trigeminal ganglion ascend to the trigeminal nuclei in the brainstem. Significantly, these nuclei, in addition to sending tactile and pain information to the thalamus, also project to multiple nearby brainstem nuclei, including the locus coeruleus and the nucleus of the solitary tract (nucleus tractus solitarii, NTS). Both of these brainstem regions profoundly affect functioning throughout the central nervous system and may play a role in the therapeutic effects of TNS.
Figure 1.
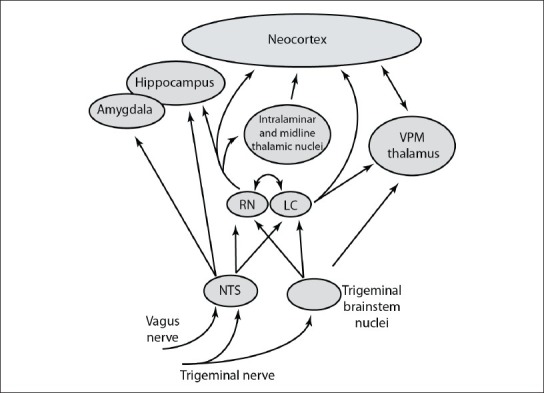
Schematic diagram of ascending projections within the central nervous system that are influenced by input from the trigeminal and vagus nerves. Note that this diagram only depicts a subset of the many divisions of and projections between these highly interconnected brain regions
The vagus nerve innervates multiple structures and organs throughout the neck and torso. Its functions include parasympathetic influence of the heart, lungs, and digestive tract, along with visceral and somatic sensory input from multiple areas and motor control of the pharynx and larynx. Afferent sensory fibers from this complex nerve project to the nodose ganglion, where their cell bodies are located, and subsequently terminate in the NTS. The NTS, in turn, projects mono- or polysynaptically to a range of areas [Figure 1].
Efferent projections are not required for the anticonvulsant effects of vagus nerve stimulation
Whereas the branches of the trigeminal nerve used thus far for therapeutic TNS (V1 and V2) consist entirely of afferent fibers, the vagus nerve carries both afferent and efferent information. However, the number of afferent fibers in the vagus nerve outweighs the number of efferent fibers by a ratio of 4:1 (Ruffoli et al. [65] for a review). Because of these efferent vagus fibers, it was suggested that the mechanism for the effects of VNS was mediated by peripheral responses to activation of efferent axons. However, multiple studies have demonstrated that this is not the case. For example, Zabara [82] showed that the anticonvulsant effects of VNS were not altered by transection of the vagus nerve distal to the stimulating electrode. Similarly, Osharina et al. [57] cut the vagus nerve distal to the site of stimulation and showed that this had no effect on their findings. Additionally, Hassert and colleagues [34] showed that chemically blocking efferent activity during VNS did not affect norepinephrine output in the amygdala.
Activation of peripheral C-fibers: Necessary or not?
One hypothesis put forth several decades ago for the central effects of VNS was that therapeutic levels of nerve stimulation required recruitment of small diameter, unmyelinated C-fibers, which have a higher stimulus threshold (e.g., require higher current or longer pulses) than A or B-fibers. C-fibers constitute a majority of the cervical vagus nerve fibers and ~40% of the trigeminal nerve fibers. [81] It was shown by Chase et al. [13] that the electroencephalogram (EEG), a readout of widespread neuronal activity, was synchronized when the faster-conducting, low threshold A- and B-fibers were activated by cervical VNS. However, when stimulus parameters were adjusted to activate C-fibers, the EEG became desynchronized, which would presumably be therapeutic against seizures. But, the issue appears to be more complicated, as there are subsequent contradictory studies on this point. Woodbury and Woodbury [80] performed a study in anesthetized rats showing that levels of VNS sufficient for C-fiber activation were required for the anticonvulsant effect of VNS, as Chase and colleagues concluded from their study. In contrast, Krahl et al. [43] selectively ablated C-fibers in the vagus nerve via subcutaneous capsaicin injection and showed that C-fiber activation was not required for VNS to terminate seizures in awake rats. These authors hypothesized that the difference in the state of vigilance of the animal (anesthetized vs. awake) might explain the discrepancy between their findings and previous studies. However, the strongest evidence against C-fiber recruitment being a necessity for the anticonvulsant effects of VNS is that patients do not report pain or autonomic side effects that would be predicted if these fibers were activated by VNS. [4]
Tests of C-fiber recruitment by therapeutic levels of TNS have not been published. However, animals do not show signs of distress during such stimulation, [27] and human patients do not report pain in conjunction with therapeutic levels of transcutaneous TNS, [23,24,60] suggesting that activation of these axons is not required.
Brainstem participation in the therapeutic effects of cranial nerve stimulation
The brainstem is a complex network of diverse neurons that has profound effects on neuronal activity throughout the neuroaxis. One set of brainstem components that is particularly relevant to the discussion of therapeutic nerve stimulation is the group of nuclei that disseminate neuromodulatory compounds. These include areas such as the locus coeruleus, the main source of noradrenergic input to the brain, and the raphe nuclei, the primary source of serotonin in the brain. In addition, as described above, the NTS is a key brainstem region for the sake of our discussion here.
Nucleus of the solitary tract
The NTS is innervated by both the trigeminal and vagus nerves, [16] among others. NTS efferents, in turn, project widely throughout the brain. Importantly, the NTS projects to the locus coeruleus [75] and the raphe nuclei. Because norepinephrine and serotonin can both have anticonvulsant effects (see below), it is significant that the trigeminal and vagus nerves have access to these brainstem regions via the NTS. In addition, the NTS projects directly to the amygdala, [30,58] hippocampus, [30] and thalamus, except for the midline and medial nuclei. [6,30]
Locus coeruleus/noradrenergic influence
The locus coeruleus is a nucleus in the brainstem that sends noradrenergic input to many levels of the central nervous system, including the raphe nuclei, the thalamus, the hippocampus, and the neocortex. [30,47,52] It receives direct or indirect afferent input from a wide range of brain regions, including the principal sensory trigeminal nuclei of the brainstem [20] and the NTS, [2,75] indicating that the trigeminal and vagus nerves can alter the activity of the locus coeruleus.
There is ample evidence showing that both norepinephrine and activity of the locus coeruleus can exert anticonvulsant effects. First, lesioning the locus coeruleus can lead to increased kindling in rats. [19] Second, Jimenez-Rivera et al., [39] demonstrated that stimulation of the locus coeruleus can delay the onset of severe seizures. Additionally, it is clear that VNS affects norepinephrine levels in multiple regions of the brain via activation of the locus coeruleus. VNS has been shown to activate c-fos, a transcription factor that indicates recent neuronal activity, [55] in locus coeruleus neurons as well as to increase norepinephrine concentrations in the hippocampus and neocortex. [63]
Because the locus coeruleus projects widely throughout the neocortex, [40] and because norepinephrine has been shown to have anticonvulsant effects, [31,45,74,79] it is plausible that the locus coeruleus plays a role in the mechanism(s) of TNS and VNS. One key piece of evidence for such a mechanism comes from Krahl and colleagues, [42] who demonstrated that lesions of the locus coeruleus in rats prevented the anticonvulsant effects of VNS. Additionally, it was later shown that VNS activates neurons in the locus coeruleus. Specifically, VNS increased the basal firing rates [26,32,50] of these neurons as well as their propensity to fire in bursts of action potentials (Manta et al., [50] though see Groves [32] ), which would presumably lead them to exert a stronger influence on their downstream targets. [28] Finally, studies have shown that norepinephrine concentration increased in the amygdala, [34] hippocampus, and neocortex [29,61,63] during VNS and the degree of norepinephrine increase was proportional to the therapeutic effect. [61] It is not yet known whether TNS also affects the locus coeruleus, though the trigeminal brainstem nuclei do influence this region, [20] so the anatomical substrate is in place for such an effect.
Raphe nuclei/serotonergic input
The raphe nuclei consist of multiple groups of cells, many of which contain serotonin. Neurons in these nuclei, project to a number of downstream targets relevant here, including the midline and intralaminar nuclei of the thalamus, the hippocampus, and the neocortex. [78] Application of serotonin agonists or reuptake inhibitors to the brain has been shown to cause an increase in seizure threshold and depletion of brain serotonin has been shown to reduce seizure threshold. [3]
Dorr and Debonnel [26] described an increase in the firing of neurons in the dorsal raphe nucleus during VNS. The increased firing in this region was only present after at least 14 days of VNS application, and was double the baseline level after 90 days of VNS. Another electrophysiological study of dorsal raphe neurons indicated that multiple VNS stimulus regimes successfully increased neuronal firing in this region after 14 days of treatment. [51] Note that Raedt et al. [61] did not observe a change in serotonin concentration in the hippocampus during VNS in rats. This may be because the increase in dorsal raphe activity does not translate into increases in serotonin in the hippocampus during VNS, or, more likely, is due to the fact that Raedt et al. [61] investigated the effects of acute VNS, whereas Dorr and Debonnel [26] looked at longer-term changes in serotonin levels.
Additionally, there are reciprocal connections between the locus coeruleus and the raphe nuclei, [30,49,66] so these neuromodulatory regions likely interact when activated directly or indirectly by TNS or VNS.
Collectively, these three brainstem regions are poised anatomically to directly or indirectly influence the neurochemistry of wide regions of the central nervous system. Their influence on seizure activity is generally inhibitory, though this depends in part on the receptor subtype that is activated in a given brain region by a given neuromodulator. It is thus likely that specific brainstem nuclei participate in the anticonvulsant effects of TNS and VNS.
Anticonvulsant effects of thalamic activity
The thalamus is made up of relatively discrete nuclei that subserve several functions. One of these functions is to transfer sensory information from the brainstem to the cortex. For example, tactile information from the face is relayed via the trigeminal nerve and the trigeminal ganglion to a group of brainstem nuclei, which project to the ventral posterior medial (VPM) thalamus. From VPM, the tactile information is sent to the primary somatosensory cortex (SI). In this capacity, the thalamus acts as a ‘relay’ [69] for sensory input. Importantly, sensory input can trigger intra-thalamic inhibition within a given nucleus or activate the inhibitory cells that comprise the reticular nucleus of the thalamus. It is thus possible that afferent trigeminal input arising from TNS could evoke sufficient inhibition to interrupt any pathological oscillatory activity within the thalamus that could contribute to seizure activity.
Neurons in the relay nuclei can function in two modes. [25,38,46,64] The first is a ‘tonic’ mode, during which neurons fire single action potentials, respond readily to the incoming input, and thus represent this input faithfully. The second is an ‘oscillatory’ or ‘burst’ mode, during which neurons fire in short bursts that occur rhythmically and do not respond reliably to external input. The tonic mode occurs when neurons are relatively depolarized, whereas, burst firing occurs when thalamic neurons are hyperpolarized.
Another function of the thalamus involves the midline and intralaminar nuclei, which appear to be related to ‘arousal’ or behavioural state. [33,41,76] These nuclei project to many divisions of the neocortex [72] and are thus positioned to have widespread influence on neocortical function. These two thalamic regions receive afferent input from over 50 nuclei in the brainstem, including the NTS and areas that deliver neuromodulators, such as the locus coeruleus and the raphe nuclei. [44]
With these two general functions, the thalamus is able to affect neuronal activity in large parts of the thalamocortical system. This is relevant to seizure activity in at least three ways. First, if thalamic nuclei are in the ‘burst’ mode described above, their neurons can participate readily in spike-and-wave discharges (SWD), which have been associated with absence epilepsy. Activation of neurons in thalamic nuclei by brainstem regions such as the NTS, trigeminal nuclei, locus coeruleus, and raphe nuclei, could push neurons into the ‘tonic’ mode, which would presumably prevent SWDs. Thus, stimulation of the trigeminal or vagus nerves might, by way of brainstem, influence on the thalamus, lead to a reduction in seizures that involve these thalamocortical rhythms.
A second method by which changes in thalamic activity might reduce seizures is by affecting overall levels of neuronal firing in the neocortex and limbic system. The midline and intralaminar thalamic nuclei have widespread, diffuse projections to the neocortex that are thought to be ‘activating′, causing heightened attention and behavioral arousal. They also send similar projections to the limbic system. Neurons in the midline and intralaminar thalamic regions can fire in burst and tonic modes, as with the relay nuclei. When in burst mode, they appear to participate in SWDs and in this manner could contribute to specific types of seizure activity. However, these thalamic nuclei receive noradrenergic and serotonergic input from the brainstem, [56] which, if depolarizing, would serve to keep the thalamus in tonic mode. This may aid in the desynchronization of cortical and limbic activity.
Finally, if the thalamus is involved in coordinating the oscillatory activity of multiple cortical regions, any manipulation that interrupts the thalamic activity would, theoretically, also interrupt the spread of the seizure activity as it travels across the neocortex. Thus, even if a cortically-based seizure were initiated, its spread could be limited by an interruption in thalamic activity.
Effects of cranial nerve stimulation on the limbic system
Because the limbic system is heavily implicated in many epilepsies, it is important to consider the effects of TNS and VNS on this region. A main route by which stimulation of the trigeminal or vagus nerves could influence the limbic system is via the neuromodulatory effects of brainstem activation. Generally, noradrenergic input from the locus coeruleus to the limbic system appears to be anticonvulsant, [10,18] and norepinephrine concentrations are increased in the limbic system by VNS. [61,63] The effects of TNS on the limbic system have not been reported to date. However, given that the trigeminal nerve projects to the NTS, which sends indirect projections to the locus coeruleus, and that there are projections from the trigeminal sensory nuclei to the locus coeruleus, it is likely that activation of the trigeminal nerve could have effects on the limbic system that are similar to those of VNS.
Changes in neocortical activity as a result of cranial nerve stimulation
Morruzi and Magoun [54] observed that stimulation of the brainstem reticular activating system caused cortical activity to shift from a ‘synchronized’ state, as might be observed during slow-wave sleep or administration of certain anesthetics, to a ‘desynchronized’ state, as is typical of rapid eye movement (REM) sleep and wakefulness. Because seizures are characterized by highly synchronous activity, it was hypothesized that desynchronization of the neocortex would have anticonvulsant effects. Subsequently, Chase and colleagues [12,13,14] demonstrated that stimulation of the vagus nerve at frequencies above 70 Hz caused synchronization of the EEG, whereas VNS presented at lower than 70 Hz caused desynchronization of the EEG. This work led to the use of low frequency (20 - 30 Hz) vagus nerve stimulation to reduce seizures, first in animals [82] and then in humans.
One method for investigating cortical excitability in response to an antiepileptic treatment is by measuring the current threshold at which motor seizures are evoked by electrical stimulation of the motor cortex. De Herdt and colleagues [21] used this technique to demonstrate that VNS significantly increased the threshold current required to elicit motor seizures. This suggested that VNS was capable of influencing cortical activity, presumably decreasing it, resulting in an increased seizure threshold. Because the NTS does not provide direct projections to the neocortex, this effect was likely due to indirect pathways, such as via the locus coeruleus.
In contrast to the vagus nerve, the trigeminal nerve passes afferent tactile sensations to the brainstem, which transmits them to the ventral posterior medial (VPM) thalamus (a relay nucleus of the thalamus), and then on to the primary somatosensory cortex (SI). The signal is propagated through this pathway very rapidly (generally less than 10 ms from the periphery to SI) and evokes an excitatory-inhibitory sequence in both VPM and SI. It is possible that neocortical activity resulting from this afferent input has a disrupting effect on neuronal firing in the neocortex, which could be involved in preventing the spread of neocortical seizures. Additionally, this effect could occur quickly and might be responsible for the rapid anticonvulsant effects of TNS. [27]
Imaging studies of cranial nerve stimulation
Schrader and colleagues investigated which brain regions were activated or deactivated by TNS, using positron emission tomography (PET) scanning. [17,68,70] Regions of activation during TNS included the inferior frontal gyrus, middle frontal gyrus, bilateral parietotemporal cortex, and bilateral anterior cingulate gyri. In contrast, deactivation was observed in the left parahippocampal gyrus, right sensorimotor cortex, right superior parietal area, bilateral temporo-occipital cortex, and bilateral visual cortex. Further studies will elucidate whether and how these changes underlie the anticonvulsant and other therapeutic effects of TNS.
Positron emission tomography scanning has also been employed in studies designed to determine which areas of the brain are involved in the therapeutic effects of VNS. Henry and colleagues [37] showed that in response to VNS there was an increase in thalamic blood flow, which was correlated with the anticonvulsant effects of VNS. Henry et al. [35] demonstrated that with both short- and long-term VNS there was an increase in cerebral blood flow in the thalamus and SI cortex (among other regions), whereas with short-term stimulation there were decreases in blood flow in the limbic system. Likewise, Chae et al. [11] described a decrease in blood flow in the limbic system, also using PET scanning.
Note, however, that other imaging studies have contradicted these results. Generally, single-photon emission computed tomography (SPECT) studies tend to show decreases in blood flow in areas that show increases with PET scans. [62,77] However, this discrepancy may have to do with the different timescales on which these methods record data. SPECT data tend to require a longer collection period. [5] Additionally, some imaging studies are conducted during VNS stimulation, whereas others are collected in the periods following stimulation, which may lead to different results. It should also be noted that it can be difficult to interpret the neuronal correlates of increased blood flow. For example, the thalamus is not a homogeneous structure in terms of either neuron type (excitatory vs. inhibitory) or function (local relay vs. modulatory). It would be critical to know which thalamic nuclei and/or what neuronal subtype within a given nucleus were activated in order to interpret these results.
Multiple studies have shown that there are increases in cerebral blood flow in the SI cortex during VNS. [36] As with the thalamus, it is not clear which layer(s) of neocortex was affected, and it is not clear whether excitatory or inhibitory neurons were preferentially affected. To complicate the matter, it was demonstrated by Snow and colleagues [71] that locus coeruleus stimulation largely decreased the firing of spontaneously active SI neurons. Note that the cell types recorded in SI were not specified in this study.
Time scales of trigeminal nerve stimulation and vagus nerve stimulation efficacy
An interesting finding about TNS and VNS is that there is evidence for both short- and long-term effects for each type of stimulation. On the short-term end of the spectrum, both TNS [27] and VNS [82] have been shown to stop ongoing seizures. For example, Fanselow and colleagues [27] demonstrated in a closed-loop paradigm, wherein stimulation was triggered by the start of a seizure, that seizures could be stopped within 0.5 seconds of seizure/stimulus onset. Similarly, Zabara [82] demonstrated that VNS in canines could stop pentylenetetrazole seizures within 0.5 - 5 seconds. In addition, for human patients, VNS devices are equipped with an external magnet that can be used to activate the VNS stimulator if a patient or caregiver detects an impending seizure. Boon et al. [9] reported successful use of this technique, with a 46% responder rate (responders showed >50% seizure reduction). In many cases seizures could be interrupted using magnet stimulation. Finally, McLachlan et al. [53] showed that seizure durations were reduced from 30 to 5 seconds with VNS in rats in a penicillin seizure model. Thus, there appears to be a fast anticonvulsant effect of cranial nerve stimulation.
Another observed phenomenon is that the effects of VNS have been shown to outlast the duration of the stimulus-ON periods. This may be one reason that the common ~30 seconds ON, ~5 minutes OFF stimulus paradigm used in VNS has therapeutic value. Most of the studies demonstrating this have shown that the anticonvulsant effects last on the order of tens of seconds to tens of minutes. For example, Zabara [82] showed that there was a period of ‘protection’ following VNS delivery that lasted approximately four times the duration of the stimulation period. Groves et al. [32] saw an increase in locus coeruleus signaling with VNS that reached statistical significance between 90 and 120 seconds after stimulus offset. McLachlan and colleagues [53] used a penicillin model to show that the reduction in interictal spike frequency they observed during 20 seconds of VNS lasted < = 3 minutes. Van Laere et al. [77] used SPECT to monitor blood flow in the thalamus following VNS, and reported that there was a period after the stimulus during which there was hypoperfusion of the thalamus that lasted for several minutes. These authors suggested that this finding might be correlated with the fact that the acute effect of VNS lasts after the stimulus is terminated. In addition, Takaya et al. [73] quantified the duration of the anticonvulsant effect of VNS and showed that it decayed with time to baseline levels over the course of 10 minutes following the end of the stimulus-ON period.
On a longer time scale, Van Bockstaele and colleagues [75] showed that in the amygdala VNS-triggered increases in norepinephrine lasted for at least 80 minutes. Likewise, Hassert et al. [34] observed a two-hour increase in norepinephrine in the amygdala with VNS. [63]
Finally, there are reports of very long-term increases in the anticonvulsant effects of VNS. [1,8] For example, DeGiorgio et al. [22] reported that VNS efficacy improved over the course of 12 months. This long time frame suggests that neuroplastic mechanisms might be involved in the therapeutic effects of cranial nerve stimulation. Ruffoli et al. [65] pointed out in a review that VNS might affect the plasticity of the epileptic circuitry itself, leading to a long-term anticonvulsant effect.
Advantages to understanding the mechanisms of therapeutic peripheral nerve stimulation
Why is it important to understand the mechanisms by which TNS or VNS cause anticonvulsant effects in the central nervous system? First, the current methods for setting stimulus parameters for these treatments are somewhat empirical, starting with recommended standards and modifying these on a patient-by-patient basis. [57] An understanding of why TNS or VNS stops seizures may allow for evidence-based modification of stimulus parameters. In a related way, it is possible that different stimulation patterns ideally activate or inactivate different regions of the brain (cf. [28,51] ). Therefore, different stimulus paradigms may be ideal for specific types of epilepsy or for different seizure loci. Currently, little is known about how to ‘tune’ stimulus characteristics to target a specific brain region or alleviate a specific seizure type, but this may be possible if the mechanism(s) for TNS and VNS are revealed.
CONCLUSIONS
In summary, it is not yet clear how or where TNS or VNS exert their anticonvulsant effects. Given the above-mentioned findings, there are several possible loci for influence, which are not mutually exclusive. First, for neocortex and hippocampus, it is possible that neuromodulatory activity (e.g., noradrenergic input) leads to desynchronization in these regions, which prevents the correlated activity of neurons that may otherwise develop into seizures or pre-seizure conditions. Second, the thalamus is known to be involved in the oscillatory components of certain types of seizures (e.g., absence seizures) and may be responsible, in part, for the spread of seizures, due to its widespread connections with diverse regions of the neocortex. Thus, changing thalamic activity, even if it involves a putative increase in activity, as suggested by multiple imaging studies, may interrupt this process, limiting the spread of seizures. Third, on a longer time scale, the excitability of epileptogenic tissue may decrease due to neuroplastic changes in relevant structures, potentially changing the epileptogenic tissue in such a way as to make it less prone to seizures. As studies of TNS and VNS mechanisms progress, these techniques can potentially be improved, so they are efficacious in a larger number of patients and can yield even higher rates of seizure reduction.
Footnotes
Disclaimer: The authors of this paper have received no outside funding, and have nothing to disclose.
Available FREE in open access from: http://www.surgicalneurologyint.com/text.asp?2012/3/5/247/103014
REFERENCES
- 1.Amar AP, DeGiorgio CM, Tarver WB, Apuzzo ML. Long-term multicenter experience with vagus nerve stimulation for intractable partial seizures: Results of the XE5 trial. Stereotact Funct Neurosurg. 1999;73:104–8. doi: 10.1159/000029764. [DOI] [PubMed] [Google Scholar]
- 2.Aston-Jones G, Shipley MT, Chouvet G, Ennis M, van Bockstaele E, Pieribone V, et al. Afferent regulation of locus coeruleus neurons: Anatomy, physiology and pharmacology. Prog Brain Res. 1991;88:47–75. doi: 10.1016/s0079-6123(08)63799-1. [DOI] [PubMed] [Google Scholar]
- 3.Bagdy G, Kecskemeti V, Riba P, Jakus R. Serotonin and epilepsy. J Neurochem. 2007;100:857–73. doi: 10.1111/j.1471-4159.2006.04277.x. [DOI] [PubMed] [Google Scholar]
- 4.Banzett RB, Guz A, Paydarfar D, Shea SA, Schachter SC, Lansing RW. Cardiorespiratory variables and sensation during stimulation of the left vagus in patients with epilepsy. Epilepsy Res. 1999;35:1–11. doi: 10.1016/s0920-1211(98)00126-0. [DOI] [PubMed] [Google Scholar]
- 5.Barnes A, Duncan R, Chisholm JA, Lindsay K, Patterson J, Wyper D. Investigation into the mechanisms of vagus nerve stimulation for the treatment of intractable epilepsy, using 99mTc-HMPAO SPET brain images. Eur J Nucl Med Mol Imaging. 2003;30:301–5. doi: 10.1007/s00259-002-1026-8. [DOI] [PubMed] [Google Scholar]
- 6.Beckstead RM, Morse JR, Norgren R. The nucleus of the solitary tract in the monkey: Projections to the thalamus and brain stem nuclei. J Comp Neurol. 1980;190:259–82. doi: 10.1002/cne.901900205. [DOI] [PubMed] [Google Scholar]
- 7.Beekwilder JP, Beems T. Overview of the clinical applications of vagus nerve stimulation. J Clin Neurophysiol. 2010;27:130–8. doi: 10.1097/WNP.0b013e3181d64d8a. [DOI] [PubMed] [Google Scholar]
- 8.Boon P, Vonck K, de Reuck J, Caemaert J. Vagus nerve stimulation for refractory epilepsy. Seizure. 2002;11(Suppl A):456. [PubMed] [Google Scholar]
- 9.Boon P, Vonck K, Van Walleghem P, D’Have M, Goossens L, Vandekerckhove T, et al. Programmed and magnet-induced vagus nerve stimulation for refractory epilepsy. J Clin Neurophysiol. 2001;18:402–7. doi: 10.1097/00004691-200109000-00003. [DOI] [PubMed] [Google Scholar]
- 10.Bortolotto ZA, Cavalheiro EA. Effect of DSP4 on hippocampal kindling in rats. Pharmacol Biochem Behav. 1986;24:777–9. doi: 10.1016/0091-3057(86)90591-5. [DOI] [PubMed] [Google Scholar]
- 11.Chae JH, Nahas Z, Lomarev M, Denslow S, Lorberbaum JP, Bohning DE, et al. A review of functional neuroimaging studies of vagus nerve stimulation (VNS) J Psychiatr Res. 2003;37:443–55. doi: 10.1016/s0022-3956(03)00074-8. [DOI] [PubMed] [Google Scholar]
- 12.Chase MH, Nakamura Y. EEG response to afferent abdominal vagal stimulation. Electroencephalogr Clin Neurophysiol. 1968;24:396. [PubMed] [Google Scholar]
- 13.Chase MH, Nakamura Y, Clemente CD, Sterman MB. Afferent vagal stimulation: Neurographic correlates of induced EEG synchronization and desynchronization. Brain Res. 1967;5:236–49. doi: 10.1016/0006-8993(67)90089-3. [DOI] [PubMed] [Google Scholar]
- 14.Chase MH, Sterman MB, Clemente CD. Cortical and subcortical patterns of response to afferent vagal stimulation. Exp Neurol. 1966;16:36–49. doi: 10.1016/0014-4886(66)90084-7. [DOI] [PubMed] [Google Scholar]
- 15.Connor DE, Jr, Nixon M, Nanda A, Guthikonda B. Vagal nerve stimulation for the treatment of medically refractory epilepsy: A review of the current literature. Neurosurg Focus. 2012;32:E12. doi: 10.3171/2011.12.FOCUS11328. [DOI] [PubMed] [Google Scholar]
- 16.Contreras RJ, Beckstead RM, Norgren R. The central projections of the trigeminal, facial, glossopharyngeal and vagus nerves: An autoradiographic study in the rat. J Auton Nerv Syst. 1982;6:303–22. doi: 10.1016/0165-1838(82)90003-0. [DOI] [PubMed] [Google Scholar]
- 17.Cook IA, Schrader LM, Geist CL, Silverman DH, Miller PR, Susselman MK, et al. Regional PET Activation with Trigeminal Nerve Stimulation in Major Depression. Boca Raton FL: NCDEU Annual Meeting. 2011 [Google Scholar]
- 18.Corcoran ME. Characteristics of accelerated kindling after depletion of noradrenaline in adult rats. Neuropharmacology. 1988;27:1081–4. doi: 10.1016/0028-3908(88)90072-x. [DOI] [PubMed] [Google Scholar]
- 19.Corcoran ME, Mason ST. Role of forebrain catecholamines in amygdaloid kindling. Brain Res. 1980;190:473–84. doi: 10.1016/0006-8993(80)90289-9. [DOI] [PubMed] [Google Scholar]
- 20.Couto LB, Moroni CR, dos Reis Ferreira CM, Elias-Filho DH, Parada CA, Pela IR, et al. Descriptive and functional neuroanatomy of locus coeruleus-noradrenaline-containing neurons involvement in bradykinin-induced antinociception on principal sensory trigeminal nucleus. J Chem Neuroanat. 2006;32:28–45. doi: 10.1016/j.jchemneu.2006.03.003. [DOI] [PubMed] [Google Scholar]
- 21.De Herdt V, De Waele J, Raedt R, Wyckhuys T, El Tahry R, Vonck K, et al. Modulation of seizure threshold by vagus nerve stimulation in an animal model for motor seizures. Acta Neurol Scand. 2010;121:271–6. doi: 10.1111/j.1600-0404.2009.01223.x. [DOI] [PubMed] [Google Scholar]
- 22.De Giorgio CM, Schachter SC, Handforth A, Salinsky M, Thompson J, Uthman B, et al. Prospective long-term study of vagus nerve stimulation for the treatment of refractory seizures. Epilepsia. 2000;41:1195–1200. doi: 10.1111/j.1528-1157.2000.tb00325.x. [DOI] [PubMed] [Google Scholar]
- 23.DeGiorgio CM, Shewmon A, Murray D, Whitehurst T. Pilot study of trigeminal nerve stimulation (TNS) for epilepsy: A proof-of-concept trial. Epilepsia. 2006;47:1213–5. doi: 10.1111/j.1528-1167.2006.00594.x. [DOI] [PubMed] [Google Scholar]
- 24.De Giorgio CM, Shewmon DA, Whitehurst T. Trigeminal nerve stimulation for epilepsy. Neurology. 2003;61:421–2. doi: 10.1212/01.wnl.0000073982.42650.57. [DOI] [PubMed] [Google Scholar]
- 25.Deschenes M, Paradis M, Roy JP, Steriade M. Electrophysiology of neurons of lateral thalamic nuclei in cat: Resting properties and burst discharges. J Neurophysiol. 1984;51:1196–219. doi: 10.1152/jn.1984.51.6.1196. [DOI] [PubMed] [Google Scholar]
- 26.Dorr AE, Debonnel G. Effect of vagus nerve stimulation on serotonergic and noradrenergic transmission. J Pharmacol Exp Ther. 2006;318:890–8. doi: 10.1124/jpet.106.104166. [DOI] [PubMed] [Google Scholar]
- 27.Fanselow EE, Reid AP, Nicolelis MA. Reduction of pentylenetetrazole-induced seizure activity in awake rats by seizure-triggered trigeminal nerve stimulation. J Neurosci. 2000;20:8160–8. doi: 10.1523/JNEUROSCI.20-21-08160.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Florin-Lechner SM, Druhan JP, Aston-Jones G, Valentino RJ. Enhanced norepinephrine release in prefrontal cortex with burst stimulation of the locus coeruleus. Brain Res. 1996;742:89–97. doi: 10.1016/s0006-8993(96)00967-5. [DOI] [PubMed] [Google Scholar]
- 29.Follesa P, Biggio F, Gorini G, Caria S, Talani G, Dazzi L, et al. Vagus nerve stimulation increases norepinephrine concentration and the gene expression of BDNF and bFGF in the rat brain. Brain Res. 2007;1179:28–34. doi: 10.1016/j.brainres.2007.08.045. [DOI] [PubMed] [Google Scholar]
- 30.Foote SL, Bloom FE, Aston-Jones G. Nucleus locus ceruleus: New evidence of anatomical and physiological specificity. Physiol Rev. 1983;63:844–914. doi: 10.1152/physrev.1983.63.3.844. [DOI] [PubMed] [Google Scholar]
- 31.Giorgi FS, Pizzanelli C, Biagioni F, Murri L, Fornai F. The role of norepinephrine in epilepsy: From the bench to the bedside. Neurosci Biobehav Rev. 2004;28:507–524. doi: 10.1016/j.neubiorev.2004.06.008. [DOI] [PubMed] [Google Scholar]
- 32.Groves DA, Bowman EM, Brown VJ. Recordings from the rat locus coeruleus during acute vagal nerve stimulation in the anaesthetised rat. Neurosci Lett. 2005;379:174–9. doi: 10.1016/j.neulet.2004.12.055. [DOI] [PubMed] [Google Scholar]
- 33.Hanbery J, Jasper H. The non-specific thalamocortical projection system. J Neurosurg. 1954;11:24–5. doi: 10.3171/jns.1954.11.1.0024. [DOI] [PubMed] [Google Scholar]
- 34.Hassert DL, Miyashita T, Williams CL. The effects of peripheral vagal nerve stimulation at a memory-modulating intensity on norepinephrine output in the basolateral amygdala. Behav Neurosci. 2004;118:79–88. doi: 10.1037/0735-7044.118.1.79. [DOI] [PubMed] [Google Scholar]
- 35.Henry TR, Bakay RA, Pennell PB, Epstein CM, Votaw JR. Brain blood-flow alterations induced by therapeutic vagus nerve stimulation in partial epilepsy: II.prolonged effects at high and low levels of stimulation. Epilepsia. 2004;45:1064–70. doi: 10.1111/j.0013-9580.2004.03104.x. [DOI] [PubMed] [Google Scholar]
- 36.Henry TR, Bakay RA, Votaw JR, Pennell PB, Epstein CM, Faber TL, et al. Brain blood flow alterations induced by therapeutic vagus nerve stimulation in partial epilepsy: I. Acute effects at high and low levels of stimulation. Epilepsia. 1998;39:983–90. doi: 10.1111/j.1528-1157.1998.tb01448.x. [DOI] [PubMed] [Google Scholar]
- 37.Henry TR, Votaw JR, Pennell PB, Epstein CM, Bakay RA, Faber TL, et al. Acute blood flow changes and efficacy of vagus nerve stimulation in partial epilepsy. Neurology. 1999;52:1166–73. doi: 10.1212/wnl.52.6.1166. [DOI] [PubMed] [Google Scholar]
- 38.Jahnsen H, Llinas R. Electrophysiological properties of guinea-pig thalamic neurones: An in vitro study. J Physiol. 1984;349:205–26. doi: 10.1113/jphysiol.1984.sp015153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Jimenez-Rivera C, Voltura A, Weiss GK. Effect of locus ceruleus stimulation on the development of kindled seizures. Exp Neurol. 1987;95:13–20. doi: 10.1016/0014-4886(87)90002-1. [DOI] [PubMed] [Google Scholar]
- 40.Jones BE, Moore RY. Ascending projections of the locus coeruleus in the rat .II. Autoradiographic study. 1977;127:25–53. [PubMed] [Google Scholar]
- 41.Jones EG. Thalamic organization and function after Cajal. Prog Brain Res. 2002;136:333–57. doi: 10.1016/s0079-6123(02)36029-1. [DOI] [PubMed] [Google Scholar]
- 42.Krahl SE, Clark KB, Smith DC, Browning RA. Locus coeruleus lesions suppress the seizure-attenuating effects of vagus nerve stimulation. Epilepsia. 1998;39:709–14. doi: 10.1111/j.1528-1157.1998.tb01155.x. [DOI] [PubMed] [Google Scholar]
- 43.Krahl SE, Senanayake SS, Handforth A. Destruction of peripheral C-fibers does not alter subsequent vagus nerve stimulation-induced seizure suppression in rats. Epilepsia. 2001;42:586–9. doi: 10.1046/j.1528-1157.2001.09700.x. [DOI] [PubMed] [Google Scholar]
- 44.Krout KE, Belzer RE, Loewy AD. Brainstem projections to midline and intralaminar thalamic nuclei of the rat. J Comp Neurol. 2002;448:53–101. doi: 10.1002/cne.10236. [DOI] [PubMed] [Google Scholar]
- 45.Libet B, Gleason CA, Wright EW, Jr, Feinstein B. Suppression of an eplieptiform type of electrocortical activity in the rat by stimulation in the vicinity of locus coeruleus. Epilepsia. 1977;18:451–62. doi: 10.1111/j.1528-1157.1977.tb04991.x. [DOI] [PubMed] [Google Scholar]
- 46.Llinas R, Jahnsen H. Electrophysiology of mammalian thalamic neurones in vitro. Nature. 1982;297:406–8. doi: 10.1038/297406a0. [DOI] [PubMed] [Google Scholar]
- 47.Loy R, Koziell DA, Lindsey JD, Moore RY. Noradrenergic innervation of the adult rat hippocampal formation. J Comp Neurol. 1980;189:699–710. doi: 10.1002/cne.901890406. [DOI] [PubMed] [Google Scholar]
- 48.Lulic D, Ahmadian A, Baaj AA, Benbadis SR, Vale FL. Vagus nerve stimulation. Neurosurg Focus. 2009;27:E5. doi: 10.3171/2009.6.FOCUS09126. [DOI] [PubMed] [Google Scholar]
- 49.Luppi PH, Aston-Jones G, Akaoka H, Chouvet G, Jouvet M. Afferent projections to the rat locus coeruleus demonstrated by retrograde and anterograde tracing with cholera-toxin B subunit and Phaseolus vulgaris leucoagglutinin. Neuroscience. 1995;65:119–60. doi: 10.1016/0306-4522(94)00481-j. [DOI] [PubMed] [Google Scholar]
- 50.Manta S, Dong J, Debonnel G, Blier P. Optimization of vagus nerve stimulation parameters using the firing activity of serotonin neurons in the rat dorsal raphe. Eur Neuropsychopharmacol. 2009;19:250–5. doi: 10.1016/j.euroneuro.2008.12.001. [DOI] [PubMed] [Google Scholar]
- 51.Manta S, El Mansari M, Blier P. Novel attempts to optimize vagus nerve stimulation parameters on serotonin neuronal firing activity in the rat brain. Brain Stimul. 2012;5:422–9. doi: 10.1016/j.brs.2011.04.005. [DOI] [PubMed] [Google Scholar]
- 52.Mason ST, Fibiger HC. Regional topography within noradrenergic locus coeruleus as revealed by retrograde transport of horseradish peroxidase. J Comp Neurol. 1979;187:703–24. doi: 10.1002/cne.901870405. [DOI] [PubMed] [Google Scholar]
- 53.McLachlan RS. Suppression of interictal spikes and seizures by stimulation of the vagus nerve. Epilepsia. 1993;34:918–23. doi: 10.1111/j.1528-1157.1993.tb02112.x. [DOI] [PubMed] [Google Scholar]
- 54.Moruzzi G, Magoun HW. Brain stem reticular formation and activation of the EEG. Electroencephalogr Clin Neurophysiol. 1949;1:455–73. [PubMed] [Google Scholar]
- 55.Naritoku DK, Terry WJ, Helfert RH. Regional induction of fos immunoreactivity in the brain by anticonvulsant stimulation of the vagus nerve. Epilepsy Res. 1995;22:53–62. doi: 10.1016/0920-1211(95)00035-9. [DOI] [PubMed] [Google Scholar]
- 56.Oke AF, Carver LA, Gouvion CM, Adams RN. Three-dimensional mapping of norepinephrine and serotonin in human thalamus. Brain Res. 1997;763:69–78. doi: 10.1016/s0006-8993(97)00404-6. [DOI] [PubMed] [Google Scholar]
- 57.Osharina V, Bagaev V, Wallois F, Larnicol N. Autonomic response and Fos expression in the NTS following intermittent vagal stimulation: Importance of pulse frequency. Auton Neurosci. 2006;126-127:72–80. doi: 10.1016/j.autneu.2006.03.011. [DOI] [PubMed] [Google Scholar]
- 58.Ottersen OP. Afferent connections to the amygdaloid complex of the rat with some observations in the cat. III Afferents from the lower brain stem. J Comp Neurol. 1981;202:335–56. doi: 10.1002/cne.902020304. [DOI] [PubMed] [Google Scholar]
- 59.Patwardhan RV, Tubbs RS, Killingsworth CR, Rollins DL, Smith WM, Ideker RE. Ninth cranial nerve stimulation for epilepsy control.Part 1: Efficacy in an animal model. Pediatr Neurosurg. 2002;36:236–43. doi: 10.1159/000058426. [DOI] [PubMed] [Google Scholar]
- 60.Pop J, Murray D, Markovic D, DeGiorgio CM. Acute and long-term safety of external trigeminal nerve stimulation for drug-resistant epilepsy. Epilepsy Behav. 2011;22:574–6. doi: 10.1016/j.yebeh.2011.06.024. [DOI] [PubMed] [Google Scholar]
- 61.Raedt R, Clinckers R, Mollet L, Vonck K, El Tahry R, Wyckhuys T, et al. Increased hippocampal noradrenaline is a biomarker for efficacy of vagus nerve stimulation in a limbic seizure model. J Neurochem. 2011;117:461–9. doi: 10.1111/j.1471-4159.2011.07214.x. [DOI] [PubMed] [Google Scholar]
- 62.Ring HA, White S, Costa DC, Pottinger R, Dick JP, Koeze T, et al. A SPECT study of the effect of vagal nerve stimulation on thalamic activity in patients with epilepsy. Seizure. 2000;9:380–4. doi: 10.1053/seiz.2000.0438. [DOI] [PubMed] [Google Scholar]
- 63.Roosevelt RW, Smith DC, Clough RW, Jensen RA, Browning RA. Increased extracellular concentrations of norepinephrine in cortex and hippocampus following vagus nerve stimulation in the rat. Brain Res. 2006;1119:124–32. doi: 10.1016/j.brainres.2006.08.04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Roy JP, Clercq M, Steriade M, Deschenes M. Electrophysiology of neurons of lateral thalamic nuclei in cat: Mechanisms of long-lasting hyperpolarizations. J Neurophysiol. 1984;51:1220–35. doi: 10.1152/jn.1984.51.6.1220. [DOI] [PubMed] [Google Scholar]
- 65.Ruffoli R, Giorgi FS, Pizzanelli C, Murri L, Paparelli A, Fornai F. The chemical neuroanatomy of vagus nerve stimulation. J Chem Neuroanat. 2011;42(4):288–296. doi: 10.1016/j.jchemneu.2010.12.002. [DOI] [PubMed] [Google Scholar]
- 66.Samuels ER, Szabadi E. Functional neuroanatomy of the noradrenergic locus coeruleus: Its roles in the regulation of arousal and autonomic function part I: Principles of functional organisation. Curr Neuropharmacol. 2008;6:235–53. doi: 10.2174/157015908785777229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Schrader LM, Cook IA, Miller PR, Maremont ER, DeGiorgio CM. Trigeminal nerve stimulation in major depressive disorder: First proof of concept in an open pilot trial. Epilepsy Behav. 2011;22:475–8. doi: 10.1016/j.yebeh.2011.06.026. [DOI] [PubMed] [Google Scholar]
- 68.Schrader LM, Geist CL, DeGiorgio C, Silverman D, Miller P, Susselman MK, et al. Regional PET activations with trigeminal nerve stimulation (TNS) and correlation with therapeutic response. Epilepsy Curr. 2012;12:235–6. [Google Scholar]
- 69.Sherman SM. A wake-up call from the thalamus. Nat Neurosci. 2001;4:344–6. doi: 10.1038/85973. [DOI] [PubMed] [Google Scholar]
- 70.Silverman DH, Geist CL, Cook I, DeGiorgio C, Susselman M, Schrader L. Effects on regional cerebral blood flow of depressed patients undergoing trigeminal nerve stimulation. J Nucl Med. 2011;52:1288. [Google Scholar]
- 71.Snow PJ, Andre P, Pompeiano O. Effects of locus coeruleus stimulation on the responses of SI neurons of the rat to controlled natural and electrical stimulation of the skin. Arch Ital Biol. 1999;137:1–28. [PubMed] [Google Scholar]
- 72.Steriade M, Glenn LL. Neocortical and caudate projections of intralaminar thalamic neurons and their synaptic excitation from midbrain reticular core. J Neurophysiol. 1982;48:352–71. doi: 10.1152/jn.1982.48.2.352. [DOI] [PubMed] [Google Scholar]
- 73.Takaya M, Terry WJ, Naritoku DK. Vagus nerve stimulation induces a sustained anticonvulsant effect. Epilepsia. 1996;37:1111–6. doi: 10.1111/j.1528-1157.1996.tb01033.x. [DOI] [PubMed] [Google Scholar]
- 74.Trottier S, Lindvall O, Chauvel P, Bjorklund A. Facilitation of focal cobalt-induced epilepsy after lesions of the noradrenergic locus coeruleus system. Brain Res. 1988;454:308–14. doi: 10.1016/0006-8993(88)90831-1. [DOI] [PubMed] [Google Scholar]
- 75.van Bockstaele EJ, Peoples J, Telegan P. Efferent projections of the nucleus of the solitary tract to peri-locus coeruleus dendrites in rat brain: Evidence for a monosynaptic pathway. J Comp Neurol. 1999;412:410–28. doi: 10.1002/(sici)1096-9861(19990927)412:3<410::aid-cne3>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- 76.van der Werf YD, Witter MP, Groenewegen HJ. The intralaminar and midline nuclei of the thalamus.Anatomical and functional evidence for participation in processes of arousal and awareness. Brain Res Brain Res Rev. 2002;39:107–40. doi: 10.1016/s0165-0173(02)00181-9. [DOI] [PubMed] [Google Scholar]
- 77.van Laere K, Vonck K, Boon P, Brans B, Vandekerckhove T, Dierckx R. Vagus nerve stimulation in refractory epilepsy: SPECT activation study. J Nucl Med. 2000;41:1145–54. [PubMed] [Google Scholar]
- 78.Vertes RP, Fortin WJ, Crane AM. Projections of the median raphe nucleus in the rat. J Comp Neurol. 1999;407:555–82. [PubMed] [Google Scholar]
- 79.Weiss GK, Lewis J, Jimenez-Rivera C, Vigil A, Corcoran ME. Antikindling effects of locus coeruleus stimulation: Mediation by ascending noradrenergic projections. Exp Neurol. 1990;108:136–40. doi: 10.1016/0014-4886(90)90020-s. [DOI] [PubMed] [Google Scholar]
- 80.Woodbury DM, Woodbury JW. Effects of vagal stimulation on experimentally induced seizures in rats. Epilepsia. 1990;31(Suppl 2):S7–19. doi: 10.1111/j.1528-1157.1990.tb05852.x. [DOI] [PubMed] [Google Scholar]
- 81.Young RF, King RB. Fiber spectrum of the trigeminal sensory root of the baboon determined by electron microscopy. J Neurosurg. 1973;38:65–72. doi: 10.3171/jns.1973.38.1.0065. [DOI] [PubMed] [Google Scholar]
- 82.Zabara J. Inhibition of experimental seizures in canines by repetitive vagal stimulation. Epilepsia. 1992;33:1005–12. doi: 10.1111/j.1528-1157.1992.tb01751.x. [DOI] [PubMed] [Google Scholar]