

Abstract
Root cortical and stelar protoplasts were isolated from maize (Zea mays L.) plants that were either well watered or water stressed, and the patch-clamp technique was used to investigate their plasma membrane K+ channel activity. In the root cortex water stress did not significantly affect inward- or outward-rectifying K+ conductances relative to those observed in well-watered plants. In contrast, water stress significantly reduced the magnitude of the outward-rectifying K+ current in the root stele but had little effect on the inward-rectifying K+ current. Pretreating well-watered plants with abscisic acid also significantly affected K+ currents in a way that was consistent with abscisic acid mediating, at least in part, the response of roots to water stress. It is proposed that the K+ channels underlying the K+ currents in the root stelar cells represent pathways that allow K+ exchange between the root symplasm and xylem apoplast. It is suggested that the regulation of K+ channel activity in the root in response to water stress could be part of an important adaptation of the plant to survive drying soils.
Water availability is one of the most important determinants of plant growth globally (Alscher and Cummings, 1990). In particular, it is the single most important factor limiting maize (Zea mays L.) production in most areas of the world (Edmeades et al., 1989). Higher plants exhibit a range of biochemical, physiological, and morphological adaptations in response to water stress, including accumulation of ABA (Davies et al., 1986). ABA is known to influence a variety of processes characteristic of water stress (e.g. stomatal closure [MacRobbie, 1992] and solute accumulation in the roots [Sharp and Davies, 1979]), and thus it is thought to act as a signal for the initiation of regulatory processes involved in the adaptation of plants to drying soils. A number of studies have shown that ABA regulates ion transport across the root with no effect on ion uptake. In most studies ion release from the stelar symplasm into the xylem apoplast was inhibited by ABA (Cram and Pitman, 1972; Schaefer et al., 1975; Behl and Jeschke, 1981). However, in other studies ABA stimulated ion transport from the root stelar symplasm (Collins and Kerrigan, 1974). This contradiction has been explained by Pitman et al. (1974), who showed that ABA inhibits ion transport in “low-salt” roots (i.e. plants grown in nutrient-free solutions), whereas in “high-salt” roots (i.e. plants grown in high-nutrient solutions) ABA promoted ion transport.
The molecular mechanisms involved in the regulation of ion transport in roots are unknown. Most ion transport in higher plant roots is catalyzed by transport proteins; plasma membrane ion channels are central components of this ion-transport pathway. Root epidermal and cortical cells mediate the net uptake of ions into the root symplasm. A range of plasma membrane ion channels has been identified in these cells, which are likely to catalyze the uptake of K+ (Findlay et al., 1994; Gassmann and Schroeder, 1994; White and Lemtiri-Chlieh, 1995; Maathuis and Sanders, 1995; Roberts and Tester, 1995), Cl− (Skerret and Tyerman, 1993), and Ca2+ (White, 1993; Piñeros and Tester, 1995). In contrast, the stelar cells of the root mediate the net loss of ions from the root symplasm into the xylem vessels for transport to the shoot. Plasma membrane ion channels have been identified in these cells, which probably catalyze the release of ions into the stelar apoplast (Wegner and Raschke, 1994; Roberts and Tester, 1995).
In previous experiments (Roberts and Tester, 1995) the patch-clamp technique was applied to protoplasts from the cortex and stele of maize roots, and K+-selective channels in these different cell types were characterized. A clear pattern emerged of the types of plasma membrane K+ channels observed in these different cell types, the activities of which paralleled the anticipated transport of K+ across the root (Roberts and Tester, 1995). In most cortical protoplasts a K+-selective inward channel was observed that would allow K+ uptake from the soil solution and would thus likely represent the pathway for low-affinity K+ uptake found in intact maize roots (Kochian et al., 1985). In maize root stelar cells a K+-selective outward channel was observed in most of these cells, which probably mediates the passive release of K+ from the root symplasm into the stelar apoplast (for subsequent transport to the shoot via the xylem vessels).
In the present study the patch-clamp technique was used to investigate: (a) previously uncharacterized TD outward and inward plasma membrane currents in maize root cortical and stelar cells, respectively, and (b) the effects of water stress and exogenous ABA on the plasma membrane currents in maize roots. A novel regulation of K+ channel activity by ABA is described that could be an important response of roots, enabling the plant to survive in drying soils.
MATERIALS AND METHODS
Plant Growth
Seeds of maize (Zea mays L.) (Pioneer 3578, Pioneer Hi-Bred International, Johnston, IA) were allowed to imbibe overnight in distilled water before being transferred to trays of vermiculite. For well-watered plants, deionized water was used to saturate the vermiculite to field capacity every 12 h (6 am and 6 pm). Water-stressed plants were treated as well-watered plants except that deionized water was withheld for 60 h before harvesting. ABA-pretreated plants were treated as well-watered plants, except that 12 h before harvesting 20 μm ABA was used to saturate the vermiculite to field capacity. This treatment elevates the ABA content of maize roots (Zhang et al., 1995). Maize seedlings were grown in a 12-/12-h day/night cycle at 24/15°C and an irradiance of 150 μmol m−2 s−1. Plants were harvested after 6 to 7 d, when roots were typically 5 to 8 cm long. To calculate the relative water content of the shoot from 7-d-old maize plants, shoot fresh weight was determined (from eight plants) immediately after harvesting. The shoots were then rehydrated by leaving them standing in deionized water in a sealed vial for 12 h and weighed (fresh weighthyd). Shoot dry weight was determined after drying the shoots for 48 h at 70°C. The relative water content of the shoots was calculated as (1 − [{fresh weighthyd − fresh weight}/{fresh weighthyd − dry weight}]) × 100. Water stress was recorded as a 1.5 ± 0.17% (n = 3) reduction in the relative water content of the shoots compared with that recorded in well-watered plants.
Protoplast Isolation
Roots were briefly washed in running tap water before being removed from the plant. After removing the tips, the cortex was stripped from the stele by hand. The following protocol was used to isolate protoplasts from the stele and the cortex. The tissue was finely chopped in a solution (500 mm sorbitol, 1 mm CaCl2, 5 mm Mes/KOH, pH 6.0) that contained (w/v) 0.5% PVP (Mr 10,000), 0.5% BSA, 0.8% cellulase (Onozuka RS, Yakult Honsha Co. Ltd., Tokyo), and 0.08% pectolyase (Sigma). The chopped tissue was agitated at 28°C in the dark for 3 h. The digest was filtered using 50-μm nylon mesh and centrifuged at 60g for 5 min. The pellet was resuspended in 5 mL of ice-cold 500 mm Suc, 1 mm CaCl2, and 5 mm Mes/KOH, pH 6.0. On top of this was layered 2 mL of ice-cold 400 mm Suc, 100 mm sorbitol, 1 mm CaCl2, 5 mm Mes/KOH, pH 6.0, followed by 1 mL of ice-cold 500 mm sorbitol, 1 mm CaCl2, 5 mm Mes/KOH, pH 6.0. This Suc step gradient was centrifuged at 200g for 5 min, and clean protoplasts were collected at the interface between the top two layers. Protoplasts were washed in 5 mL of the same solution that was used in the top layer of the gradient and centrifuged at 60g for 5 min. Protoplasts were resuspended in 1 mL of the same solution and stored on ice.
Electrophysiology
Whole-cell currents from protoplasts were recorded at room temperature (approximately 20°C) with an Axopatch 200A amplifier (Axon Instruments, Foster City, CA) using conventional patch-clamp techniques (Hamill et al., 1981). Cells were held in a chamber that was perfused. The chamber had a thin glass base to which protoplasts adhered loosely. Electrodes were pulled from borosilicate glass capillaries (Kimax 51, Kimax Products, Vineland, NJ) and fire polished using a Zeitz-Instrumente Universal puller (Augsburg, Germany) to give resistances of 10 to 11 MΩ in sealing solutions (see below). An Ag/AgCl reference electrode was connected to the bath via a 3 m KCl/agar salt bridge. After GΩ seals were formed, strong suction was applied to the interior of the pipette to obtain the whole-cell configuration. Whole-cell capacitance and series resistance were partially compensated for by the amplifier. Access resistance was monitored during experiments and was usually less than 20 MΩ. Before analog-to-digital conversion, the voltage signals representing clamp currents were low-pass filtered at 2 kHz.
The generation of voltage test pulses, recording of whole-cell currents, and storage of data were controlled by the software package pClamp6.0 (Axon Instruments) and a 486 personal computer. Analysis of data was done by pClamp software and FigP (version 2.2, Biosoft, Cambridge, UK). Outside-out patches were obtained from the whole-cell configuration by pulling away the pipette from the protoplast. Single-channel currents were measured and analyzed using the same equipment and software that was used for whole-cell experiments. Data were digitized at 1 kHz and filtered at 200 or 300 Hz for analysis. Liquid-junction potentials were corrected for in all experiments as described by Neher (1992). Tip potentials were measured at the end of an experiment by measuring the potential change when the pipette tip was broken. Only experiments in which the tip potential was less than 2 mV were used. Ion equilibrium potentials were calculated after correction for ionic activities (as calculated by GEOCHEM-PC; Parker et al., 1994). Variations in data are presented as the se.
Solutions
All solutions were filtered (0.22 μm, Millipore) before use. GΩ seals were formed in a sealing solution that contained 15 mm KCl, 10 mm CaCl2, and 5 mm Mes, adjusted to pH 6.0 with 2 mm Tris base, and adjusted to 700 milliosmole kg−1 using sorbitol. After obtaining a whole-cell configuration this solution was replaced by a standard bath solution, which was the same as the sealing solution but with only 0.1 mm CaCl2; all currents were recorded in this solution unless otherwise stated. A standard intracellular (pipette-filling) solution (100 mm potassium gluconate, 3 mm MgCl2, 10 mm Hepes, 3 mm K2ATP, and 4 mm EGTA adjusted to pH 7.2 with 17 mm KOH and adjusted to 720 milliosmole kg−1 using sorbitol) was used in all experiments. ABA was added to the standard bath solution as required.
Analysis of K+ Channel Activity in Maize Roots
The effects of water stress and ABA on the TD currents in both the root cortex and stele were investigated using the patch-clamp technique and standard bath solution. In the analysis, the densities of the TD inward and outward currents were calculated at −175 and +45 mV, respectively. A second type of outward current, which activated and deactivated rapidly (“instantaneously”), was also observed upon membrane depolarization in most protoplasts. This current was not characterized in the present study; only the TD currents were investigated. The magnitudes of the TD outward currents were calculated by subtraction of instantaneous currents from steady-state currents. The TD inward currents were calculated after subtraction of the linear “leak” current (i.e. the current resulting from the voltage-independent whole-cell membrane and seal resistances calculated by applying a small voltage to the pipette) from the steady-state current. Zero-current values were recorded for cells that did not exhibit a TD current; these were incorporated into the calculation of the mean current density. Magnitudes of current density could be calculated to an accuracy of approximately ± 1 mA m−2. Access resistances during the patch-clamp experiment were monitored, and thus it was possible to conclude that the absence of a TD current was not caused by pipette blockage. The mean current density was taken to reflect ion channel activity. Because whole-cell TD currents are carried mainly by K+ (in the conditions used in the present study), it is assumed that the TD currents reflect mainly K+ channel activity. Student's t tests were performed on the mean current density values to establish whether K+ channel activity was significantly affected by a particular growth regime or treatment.
RESULTS
In a previous report TD K+-selective inward and outward currents were characterized in the plasma membrane of maize root cortical and stelar cells, respectively (Roberts and Tester, 1995). In that study small TD outward currents in cortical cells and TD inward currents in stelar cells were also observed but were not characterized. In the present study these previously uncharacterized TD currents are investigated further.
Outward Current in Maize Root Cortical Protoplasts
Figure 1A shows the TD outward current from a maize root cortical protoplast. Tail-current protocols were used to determine the major ion responsible for this TD outward current (for method, see Roberts and Tester, 1995). The reversal potential (Erev) of the outward current in a standard bath solution (EK = −48 mV and ECl = −23 mV) was −36 ± 2.8 mV (n = 4). Increasing external K+ (EK = −32 mV and ECl = −40 mV) shifted Erev of the outward current to −19 ± 2.7 mV (n = 3), which was closer to the equilibrium potential for K+ (EK) than that for Cl− (ECl), and consistent with this current being primarily carried by K+. However, Erev was approximately 12 to 13 mV positive of EK, indicating that some other ion with a more positive equilibrium potential than EK was contributing to this conductance. This is similar to what was reported for the TD outward current in the stele of maize roots (Roberts and Tester, 1995, 1997) in which Ca2+ influx was shown to be responsible for the deviation of Erev from EK (Roberts and Tester, 1997). In the present study the equilibrium potential for Ca2+ (ECa) remained positive of EK, suggesting that Ca2+ also contributed to the outward current in cortical protoplasts.
Figure 1.

A, Current-voltage relationship of the TD (•) component of the currents shown in the inset. Inset, Plasma membrane currents resulting from voltage pulses ranging from +65 to −95 mV (in 20-mV steps at intervals of 8 s). Holding potential was −75 mV. B, Voltage dependence of the time constants of activation of whole-cell outward currents from maize root cortical protoplasts (error bars denote the se; n = 3). Inset, Examples of least-squares fits of Equation 1 to the activation of whole-cell outward currents from root cortical protoplasts from well-watered plants. Holding potential was −75 mV. C, Recordings of outward K+ channel activity in an outside-out patch from a cortical protoplast. Standard bath solution was used. The holding voltage for each recording is shown on the right. Upward deflections represent channel openings. D, Single-channel current as a function of voltage. Data shown are from two patches in standard bath solution (EK = −48 mV) with different symbols for each cell. The unitary conductance (G) and reversal potential (Erev) were determined by fitting a linear regression to the data; G = 30 pS, Erev = −38 mV.
The outward current in cortical protoplasts activated with a sigmoidal time course and could be fitted by a Hogkin-Huxley-type model (Hodgkin and Huxley, 1952), according to the equation:
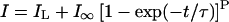 |
1 |
where p is the number of independent membrane-bound gating particles that control the opening of the channel, IL is a constant leak, and I∞ is the steady-state current after activation. Currents were best fitted (using the least-squares fit method) when p was set to 2 (Fig. 1B, inset). The time constant (τ) decreased slightly as voltage increased from +5 to +65 mV (Fig. 1B). Equation 1 also best fitted the activation kinetics of the outward current in maize root stelar protoplasts with a similar relationship between voltage and the activation time constant (Roberts and Tester, 1995).
Single-channel activity such as that shown in Figure 1C was observed in outside-out patches from cortical cell protoplasts. These single-channel currents also reversed approximately 10 mV positive of EK, consistent with this channel activity underlying the macroscopic outward currents in cortical protoplasts (although ion channel activity other than that shown in Fig. 1C may also contribute to the macroscopic TD outward current). This channel had a unitary conductance of 30 pS (15:123 mm K+, outside:cytosol) (Fig. 1D), similar to that reported for the outward K+ channel in the stele of maize when recorded in similar solutions (Roberts and Tester, 1995).
Inward Current in Maize Root Stelar Protoplasts
Figure 2A shows the TD inward current from a maize root stelar protoplast. Tail currents were also used to determine the major ion responsible for the whole-cell inward current in stelar cell protoplasts. In standard bath solution Erev was −50 ± 2.5 mV (n = 6). This is much closer to the reversal potential of K+ than any other ion in solution, and suggests that K+ is the major ion responsible for this current. The activation kinetics of the inward current were best fitted (using the least-squares fit method) by the sum of two exponential components using the following equation (Fig. 2B, inset):
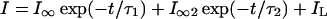 |
2 |
where IL is a constant leak and I∞ and I∞2 are the steady-state currents of the two components after activation. The time constants (τ1 and τ2) showed no apparent voltage dependence between −195 and −115 mV (Fig. 2B). Equation 2 was also used to describe the activation kinetics of the inward current in maize root cortical protoplasts, and a similar relationship between voltage and the time constants was observed (Roberts and Tester, 1995).
Figure 2.

A, Current-voltage relationship of the TD (•) component of the currents shown in the inset. Inset, Plasma membrane currents resulting from voltage pulses ranging from +65 to −175 mV (in 20-mV steps at intervals of 8 s). Holding potential was −75 mV. The displayed currents have had the leak current subtracted (leak conductance was 200 pS). B, Voltage dependence of the time constants of activation of the whole-cell inward current from maize root stelar protoplasts (the number of replicates is indicated in the parentheses for each point; error bars denote the se). Inset, Examples of least-squares fits of Equation 2 to the activation of whole-cell inward currents from root stelar protoplasts from water-stressed plants. Holding potential was −75 mV. C, Recordings of inward K+ channel activity in an outside-out patch from a stelar protoplast. A standard bath solution was used. The holding voltage for each recording is shown on the right. Downward deflections represent channel openings. D, Single-channel current as a function of voltage. Data are shown from three patches in standard bath solution (EK = −48 mV), with different symbols for each cell. The unitary conductance (G) and reversal potential (Erev) were determined by fitting a linear regression to the data; G = 10 pS, Erev = −53 mV.
Single-channel activity, such as that shown in Figure 2C, was observed in outside-out patches from stelar cell protoplasts. These single-channel currents reversed close to EK, which is consistent with this channel activity underlying the macroscopic inward currents in stelar protoplasts (although ion channel activity other than that shown in Figure 2C may also contribute to the macroscopic TD inward current). The channel had a unitary conductance of 10 pS (15:123 mm K+, outside:cytosol) (Fig. 2D), similar to that reported for the inward K+ channel in the cortex of maize when recorded in similar solutions (Roberts and Tester, 1995).
Effects of Water Stress and ABA on K+ Channel Activity in Maize Roots
Comparing well-watered and water-stressed roots, a similar mean current density was observed for the TD inward current (Fig. 3) in the root cortex; thus, K+ channel activity underlying the inward current was unaffected by water stress (t = 0.432). Similarly, water stress only slightly reduced the mean density of the outward current (Fig. 3), but this was not statistically significant (t = 0.09). The effects of water stress were more conspicuous on the K+ channel activity of the stelar cell. Water stress significantly reduced the magnitude of the TD outward K+ current (Fig. 3; t < 0.001), but did not significantly affect the magnitude of the TD inward current (t = 0.44).
Figure 3.

Mean current density of the whole-cell TD inward and outward currents in protoplasts from the cortex and stele of maize roots that were either well watered (WW), water stressed (WS), or pretreated with ABA. Open and hatched bars represent values from cortical and stelar protoplasts, respectively. Error bars denote the se. Positive values represent outward current density and negative values represent inward current density. The number of replicates is shown in parentheses for each determination of mean current density. Inset, As for the main figure except that the mean current density of whole-cell instantaneously activating outward current is shown from cortical and stelar protoplasts. Current densities were calculated at +45 and −175 mV (for outward and inward current, respectively).
ABA pretreatment of maize roots also had no significant effect on K+ channel activity in the cortex. Student's t tests indicated that the TD inward K+ current in cortical protoplasts from ABA-pretreated roots was not significantly different from that observed in well-watered (t = 0.269) and water-stressed roots (t = 0.21) (Fig. 3). Likewise, the TD outward current in cortical protoplasts was unaffected by ABA pretreatment compared with that in well-watered (t = 0.36) and water-stressed roots (t = 0.23) (Fig. 3).
More conspicuous was the effect of ABA pretreatment on K+ channel activity in stelar protoplasts. ABA pretreatment significantly increased the magnitude of the TD inward current in root stelar cells (Fig. 3) compared with well-watered (t = 0.033) and water-stressed (t = 0.038) plants. The magnitude of the TD outward current in the stele was also significantly reduced by ABA pretreatment compared with that observed in well-watered roots (t = 0.002), but it was not significantly different from that observed in water-stressed roots (t = 0.103).
In addition to the TD outward current, an instantaneously activating outward current was also observed in both the cortical and stelar protoplasts. Although the identity of this current is not known, it is noteworthy that this current was not significantly affected by either water stress or ABA pretreatment of roots (Fig. 3, inset). Furthermore, there was no effect of drought or ABA on the voltage dependence of current activation, so the current densities shown in Figure 3 represent effects of treatments on membrane conductance.
In addition to pretreating roots with ABA, the direct effects of ABA on the plasma membrane currents in stelar protoplasts from well-watered roots were also investigated. In 6 of 11 protoplasts addition of 20 μm external ABA irreversibly reduced the magnitude of the outward current by 47.5 ± 8% (n = 6) (Fig. 4). This effect occurred between 2 and 20 min after the addition of ABA; it was not caused by nonspecific “rundown,” because outward currents were stable before addition of ABA (data not shown) and “rundown” of outward currents was not observed in untreated stelar cells. No effect of ABA was observed on the inward current in root stelar (or cortical) protoplasts (data not shown). Furthermore, addition of ABA (for up to 1 h) did not induce an inward current in stelar protoplasts, which did not exhibit the inward current before the addition of ABA (data not shown).
Figure 4.

The effect of external ABA on the outward current from protoplasts from the stele of maize roots grown in well-watered conditions. A standard bath solution was used. Holding potential was −75 mV and voltage pulses ranged from +65 to −75 mV (in 20-mV steps at intervals of 30 s). A, Before addition of ABA. B, Five minutes after addition of 20 μm external ABA. C, Current-voltage relationship of the time-dependent component of outward currents from stelar protoplasts before (•) and after (▪) the addition of 20 μm external ABA. Values shown are the mean currents from six independent experiments and the error bars denote the se. Currents were normalized with respect to the current at +65 mV before the addition of ABA for each independent experiment.
DISCUSSION
Similar inward and outward K+ channels exist in both the cortex and stele of maize roots with respect to their selectivity, activation kinetics, and single-channel conductance. However, although the same channel types appear to be present in both the cortex and stele of maize roots, the channels are regulated differently. This could reflect the fact that either different signal transduction pathways exist in the cortical and stelar cells or that the channels themselves are different with respect to their sensitivity to signaling compounds.
Physiological Significance of the Regulation of K+ Channel Activity by Water Stress and ABA
Cortex
Inward K+ channels in the cortex and epidermis of higher plant roots probably mediate low-affinity K+ uptake from the soil solution (Gassmann and Schroeder, 1994; Maathuis and Sanders, 1995; Roberts and Tester, 1995). In the present study water stress and ABA had no significant effect on the K+ channels underlying the TD inward current in root cortical protoplasts and thus had no effect on the pathway for low-affinity K+ uptake. This is consistent with previous reports, which show that ABA does not regulate low-affinity K+ uptake in intact maize and barley (Hordeum vulgare L.) roots (Cram and Pitman, 1972; Schaefer et al., 1975; Pitman and Wellfare, 1978).
Stele
ABA is thought to act as a signal for the initiation of water-stress-induced processes in plants (Davies et al., 1986). In the present study K+ channel activity underlying the TD outward current in the root stele was significantly reduced by both water stress and ABA. However, ABA pretreatment also increased K+ channel activity responsible for the TD inward current. This shows that the elevation of ABA content cannot account for the water-stress response. This may reflect that (a) the ABA content of roots from water-stressed plants may be different from that in ABA-pretreated roots, and that the K+ channel activity in maize roots may be dependent on the concentration of ABA; or (b) other signaling compounds that also accumulate in roots in response to water stress (e.g. cytokinins [Collins and Kerrigan, 1974] and auxins [Ribaut and Pilet, 1994]) regulate K+ channel activity in the stele (Pitman et al., 1974). It will be necessary to investigate the effects of differing ABA concentrations and other potential signaling compounds on ion channel activity in the root stele.
Root stelar cells regulate the ionic composition of the transpiration stream (Clarkson, 1993). The outward K+ channels in these cells represent a pathway for the efflux of K+ from the root symplasm into the xylem apoplast (Wegner and Raschke, 1994; Roberts and Tester, 1995). The inward K+ channels may be involved in the reabsorption of K+ from the xylem apoplast (Wegner et al., 1994; Wegner and Raschke, 1994). Thus, water stress and ABA modify the permeability of the plasma membrane of root stelar cells in favor of K+ influx and against K+ efflux. However, the regulation of K+ channels by themselves only affects the capacity for passive transport of K+. K+ transport via ion channels will also depend on membrane potential and cytosolic and extracellular K+ concentrations. Thus, plasma membrane hyperpolarization (to generate an electrochemical gradient favoring K+ influx) is necessary for increased passive K+ uptake via K+ channels in stelar cells. ABA is reported to stimulate a transient increase in proton efflux into the xylem apoplast of onion roots (Clarkson and Hanson, 1986), which could indicate that the proton pump is stimulated by ABA. Activation of the proton pump could mediate a membrane hyperpolarization. However, to date there are no measurements of the effects of ABA on the membrane potential of root stelar cells.
Several studies have shown that ABA and water stress promote K+ accumulation by root stelar cells. Tracer-flux experiments have shown that ABA reduces K+ efflux from the stelar symplasm of maize and barley without any effect on K+ uptake by the root cortex (Cram and Pitman, 1972; Schaefer et al., 1975; Pitman and Wellfare, 1978). Although Collins and Kerrigan (1974) showed that ABA promotes ion release from the stele symplasm in maize roots, this contradiction has been explained by Pitman et al. (1974), who showed that ABA inhibits ion efflux in low-salt roots, whereas in high-salt roots ABA promotes ion efflux. In the present study low-salt roots were used. In addition to the flux experiments, a 2-fold increase in the vacuolar K+ content of root stelar cells has been reported in water-stressed maize plants relative to that in nonstressed plants (Pritchard et al., 1996). The regulation of the K+ channel activity reported in the present study is consistent with both reduced K+ transport to the shoot and increased accumulation of K+ in the root symplasm.
K+ accumulation by the root may be part of an important adaptation of the plant, allowing it to survive drying soils. An early response of plants to water stress is an inhibition of shoot growth while root growth is maintained (Sharp and Davies, 1979; Saab et al., 1990). Thus, ions transported from the root could accumulate in the apoplast of leaves, lowering the extracellular osmotic potential and decreasing the water deficit of leaf cells. Also, ABA promotes the net accumulation of solutes in roots during water stress (Sharp and Davies, 1979; Henson, 1985; Ribaut and Pilet, 1991), which is thought to maintain a water potential favoring the uptake of water and cell turgor pressure necessary for root growth (Morgan, 1984). Although organic compounds are the major constituents of osmoregulation in plant cells during water stress (Morgan, 1984), inorganic ions (e.g. K+) would also contribute to osmoregulation.
Mechanism of Regulation of K+ Channels in Maize Roots
ABA may regulate K+ channels in two ways: (a) via expression and/or incorporation of channel proteins in the plasma membrane (see below) or (b) by regulating the activity of channel proteins after their incorporation into the plasma membrane. Application of ABA to stelar protoplasts decreased the outward current in approximately 55% of stelar cells, consistent with ABA regulation of the activity of the outward K+ channels after their incorporation into the plasma membrane. It is noteworthy that ABA regulation of K+ channel activity in maize root stele is opposite to that observed in guard cells. In guard cells ABA increases the outward K+ current and decreases the inward K+ current (Blatt and Armstrong, 1993; Lemtiri-Chlieh and MacRobbie, 1994). Plasma membrane K+ channel activity in mesophyll cells is also regulated differently from that in guard cells (Li et al., 1994). Future experiments should be able to determine the mechanism of ABA regulation of K+ channel activity in root stelar cells and whether there are similarities with mesophyll cells.
It is not clear why only 55% of maize root stelar cells exhibiting an outward current respond to extracellular ABA. However, in similar patch-clamp experiments on guard cell protoplasts, ABA regulation of K+ currents was observed in only approximately 70% of cells (Lemtiri-Chlieh and MacRobbie, 1994), whereas in intact guard cells (which were impaled with a microelectrode) ABA is reported to regulate the K+ currents in nearly all of the cells (Blatt and Armstrong, 1993). In patch-clamp experiments the cytosol from the protoplast is replaced by the pipette solution. Thus, the absence of the ABA-mediated effect during patch-clamp experiments may be attributable to the loss of soluble cytoplasmic factors, which are an integral part of the ABA signal transduction pathway. In experiments in which protoplasts are isolated from ABA-pretreated roots, the ABA-mediated regulation of K+ channel activity occurs before the patch-clamp experiment. In these experiments current magnitudes remained stable during whole-cell experiments, indicating that the effects of ABA pretreatment on K+ channel activity in intact stelar cells were unaffected by replacement of the cytosol with pipette solution.
Maize root stelar cells from ABA-pretreated plants had increased inward K+ channel activity; however, no effect of ABA was observed on inward K+ channel activity during a patch-clamp experiment. Hence, it is not known if ABA regulates the activity of the inward K+ channel (but is not observed because of the loss of cytoplasmic factors during the patch-clamp experiment; see above) or if ABA regulates the expression of the K+ channel protein in the plasma membrane. Schaefer et al. (1975) estimated that protein turned over by 50% at 23°C in approximately 2 h in the root stele. Thus, ABA could rapidly regulate the permeability of the plasma membrane via the expression of the inward K+ channel. It may be possible to investigate the expression of the inward K+ channel in maize root stelar cells using antibodies raised against the inward K+ channel AKT1 (Lagarde et al., 1996).
To summarize, the present study demonstrates that K+ channel activity in the stele of maize roots is regulated by water availability and ABA. This regulation may at least in part represent the mechanism by which ABA and water stress modulate K+ transport and accumulation in the intact root (Cram and Pitman, 1972; Schaefer et al., 1975; Pitman and Wellfare, 1978; Pritchard et al., 1996). Future investigations will need to focus on determining whether ABA alone is regulating K+ channel activity during water stress or whether other factors are involved (e.g. cytokinins and auxins). It will also be of interest to investigate the mechanism of ABA regulation of these ion channels and to determine whether this represents a novel signaling pathway in plant cells.
ACKNOWLEDGMENTS
I thank Mr. P. Freeman for technical assistance and Mark Tester for useful discussions and comments on the manuscript.
Abbreviations:
- pS
picosiemens
- TD
time-dependent
Footnotes
This work was supported by a grant from the Agricultural and Food Research Council to Mark Tester.
LITERATURE CITED
- Alscher RG, Cummings JR. Stress Responses in Plants: Adaptations and Acclimation Mechanisms. New York: Wiley-Liss; 1990. [Google Scholar]
- Behl R, Jeschke WD. Influence of abscisic acid on unidirectional fluxes and intracellular compartmentation of K+ and Na+ in excised barley root segments. Physiol Plant. 1981;53:95–100. [Google Scholar]
- Blatt MR, Armstrong F. K+ channels of stomatal guard cells: abscisic acid-evoked control of the outward rectifier mediated by cytoplasmic pH. Planta. 1993;191:330–341. [Google Scholar]
- Clarkson DT (1993) Roots and the delivery of solutes to the xylem. Philos Trans R Soc Lond B 341: 5–17
- Clarkson DT, Hanson JB. Proton fluxes and the activity of a stelar proton pump in onion roots. J Exp Bot. 1986;37:1136–1150. [Google Scholar]
- Collins JC, Kerrigan AP. The effect of kinetin and ABA on water and ion transport in isolated guard cells. New Phytol. 1974;73:309–314. [Google Scholar]
- Cram WJ, Pitman MG. The action of abscisic acid on ion uptake and water flow in plant roots. Aust J Biol Sci. 1972;25:1125–1132. [Google Scholar]
- Davies WJ, Metcalfe J, Lodge TA, da Costa AR. Plant growth substances and the regulation of growth under drought. Aust J Plant Physiol. 1986;13:105–125. [Google Scholar]
- Edmeades GO, Bolanos J, Laffite HR, Rajaram S, Pfeiffer W, Fischer RA (1989) Traditional approaches to breeding for drought resistance in cereals. In FWG Baker, ed, Drought Resistance in Cereals. ICSU and CABI, Paris and Wallingford, UK, pp 27–52
- Findlay GP, Tyerman SD, Garrill A, Skerret M. Pump and K+ inward rectifiers in the plasmalemma of wheat root protoplasts. J Membr Biol. 1994;139:103–116. doi: 10.1007/BF00232429. [DOI] [PubMed] [Google Scholar]
- Gassmann W, Schroeder JI. Inward-rectifying K+ channels in root hairs of wheat. A mechanism for aluminum-sensitive low-affinity K+ uptake and membrane potential control. Plant Physiol. 1994;105:1399–1408. doi: 10.1104/pp.105.4.1399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch clamp techniques for high resolution current recording from cells and cell-free membrane patches. Pfluegers Arch. 1981;391:85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- Henson IE. Solute accumulation and growth in plants of pearl millet (Pennisetum americanum [L] Leeke) exposed to abscisic acid or water stress. J Exp Bot. 1985;36:1889–1899. [Google Scholar]
- Hodgkin AL, Huxley AF. A quantitative description of membrane current and its implication to conduction and excitation in nerve. J Physiol. 1952;144:505–524. doi: 10.1113/jphysiol.1952.sp004764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kochian LV, Xin Zhi J, Lucas WJ. Potassium transport in corn roots. IV. Characterization of the linear component. Plant Physiol. 1985;79:771–776. doi: 10.1104/pp.79.3.771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagarde D, Mireille B, Lepetit M, Conejero G, Gaymard F, Astruc S, Grignon C. Tissue-specific expression of Arabidopsis AKT1 gene is consistent with a role in K+ nutrition. Plant J. 1996;9:195–203. doi: 10.1046/j.1365-313x.1996.09020195.x. [DOI] [PubMed] [Google Scholar]
- Lemtiri-Chlieh F, MacRobbie EAC. Role of calcium in the modulation of Vicia guard cell potassium channels by abscisic acid: a patch clamp study. J Membr Biol. 1994;137:99–107. doi: 10.1007/BF00233479. [DOI] [PubMed] [Google Scholar]
- Li W, Luan S, Schreiber SL, Assmann SM. Evidence for protein phosphatase 1 and 2A regulation of K+ channels in two types of leaf cells. Plant Physiol. 1994;106:963–970. doi: 10.1104/pp.106.3.963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maathuis FJM, Sanders D. Contrasting roles in ion transport of two K+-channel types in root cells of Arabidopsis thaliana. Planta. 1995;197:456–464. doi: 10.1007/BF00196667. [DOI] [PubMed] [Google Scholar]
- MacRobbie EAC. Calcium and ABA-induced stomatal closure. Philos Trans R Soc Lond B. 1992;338:5–18. [Google Scholar]
- Morgan JM. Osmoregulation and water stress in higher plants. Annu Rev Plant Physiol. 1984;35:299–319. [Google Scholar]
- Neher E. Correction for liquid junction potentials in patch clamp experiments. Methods Enzymol. 1992;207:123–131. doi: 10.1016/0076-6879(92)07008-c. [DOI] [PubMed] [Google Scholar]
- Parker DR, Norvell WA, Chaney RL (1994) GEOCHEM-PC: a chemical speciation program for IBM and compatible computers. In RH Leoppert, S Goldberg, AP Schwab, eds, Chemical Equilibrium Reaction Models. American Society of Agronomy, Madison, WI, pp 253–269
- Piñeros M, Tester M. Characterization of a voltage-dependent Ca2+-selective channel from wheat roots. Planta. 1995;195:478–488. [Google Scholar]
- Pitman MG, Luttge U, Lauchli A, Ball E. Action of abscisic acid on ion transport as affected by root temperature and nutritional status. J Exp Bot. 1974;25:147–155. [Google Scholar]
- Pitman MG, Wellfare D. Inhibition of ion transport in excised barley roots by abscisic acid: relation to water permeability of the roots. J Exp Bot. 1978;29:1125–1138. [Google Scholar]
- Pritchard J, Fricke W, Tomos D. Turgor-regulation during extension growth and osmotic stress of maize roots: an example of single cell mapping. Plant Soil. 1996;187:11–21. [Google Scholar]
- Ribaut JM, Pilet PE. Effects of water stress on growth, osmotic potential and abscisic acid content of maize roots. Physiol Plant. 1991;81:156–162. [Google Scholar]
- Ribaut JM, Pilet PE. Water stress and indol-3yl-acetic acid content of maize roots. Planta. 1994;193:502–507. [Google Scholar]
- Roberts SK, Tester M. Inward and outward K+-selective currents in the plasma membrane of protoplasts from maize root cortex and stele. Plant J. 1995;8:811–825. [Google Scholar]
- Roberts SK, Tester M. Permeation of Ca2+ and monovalent cations through an outwardly rectifying channel in maize root stelar cells. J Exp Bot. 1997;48:839–846. [Google Scholar]
- Saab IN, Sharp RE, Pritchard J, Voetberg G. Increased endogenous abscisic acid maintains primary root growth and inhibits shoot growth of maize seedlings at low water potentials. Plant Physiol. 1990;93:1329–1336. doi: 10.1104/pp.93.4.1329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaefer N, Wildes RA, Pitman MG. Inhibition by p-fluorophenylalanine of protein synthesis and of ion transport across the roots in barley seedlings. Aust J Plant Physiol. 1975;2:61–73. [Google Scholar]
- Sharp RE, Davies WJ. Solute regulation and growth by roots and shoots in water-stressed maize plants. Planta. 1979;147:43–49. doi: 10.1007/BF00384589. [DOI] [PubMed] [Google Scholar]
- Skerret M, Tyerman SD. A channel that allows inwardly directed fluxes of anions in protoplasts derived from wheat roots. Planta. 1993;102:295–305. [Google Scholar]
- Wegner LH, de Boer AH, Raschke K. Properties of the inward rectifier in the plasma membrane of xylem parenchyma cells from barley roots: effects of TEA+, Ca2+, Ba2+, and La3+ J Membr Biol. 1994;142:363–379. doi: 10.1007/BF00233442. [DOI] [PubMed] [Google Scholar]
- Wegner LH, Raschke K. Ion channels in the xylem parenchyma of barley roots. Plant Physiol. 1994;105:799–813. doi: 10.1104/pp.105.3.799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White PJ. Characterization of a high-conductance, voltage-dependent cation channel from the plasma membrane of rye roots in planar lipid bilayers. Planta. 1993;191:541–551. [Google Scholar]
- White PJ, Lemtiri-Chlieh F. Potassium currents across the plasma membrane of protoplasts derived from rye roots: a patch clamp study. J Exp Bot. 1995;46:457–511. [Google Scholar]
- Zhang J, Zhang X, Liang J. Exudation rate and hydraulic conductivity of maize roots are enhanced by soil drying and abscisic acid treatment. New Phytol. 1995;131:329–336. [Google Scholar]