

Abstract
NK cells exhibit the highest cytotoxic capacity within the immune system. Alteration of their number or functionality may have a deep impact on overall immunity. This is of particular relevance in aging where the elderly population becomes more susceptible to infection, cancer, autoimmune diseases, and neurodegenerative diseases amongst others. As the fraction of elderly increases worldwide, it becomes urgent to better understand the aging of the immune system to prevent and cure the elderly population. For this, a better understanding of the function and phenotype of the different immune cells and their subsets is necessary. We review here NK cell functions and phenotype in healthy aging as well as in various age-associated diseases.
1. Cellular Senescence
Aging, at the whole organism scale, is a very complex process, involving many different mechanisms. Studying the normal senescence process is also complex as we are permanently surrounded by pathogens, have different lifestyle habits, and exposed to different levels of stress which all influence the senescence process. Senescence biology really started in 1961 with Drs. Hayflick and Moorhead [1]. At this time, cells were supposed to be immortal in vitro and death occurring because of nonoptimal conditions. He shattered this dogma by stating that cells were programmed to divide a certain number of times before entering a replicative senescence state, where cells stop to divide. In 1971, Olovnikov discovered that this phenomenon was due to a DNA shortening, occurring at each division [2, 3]. Then, telomeres were studied first in 1978 in Tetrahymena thermophila [4] and few years later in human [5]. The first hints that telomere length could be a cause of senescence came from the observation that its length was not the same in every tissue [6]. One year before, Drs. Greider and Blackburn discovered the telomerase [7], and Hayflick phenomenon could be explained by the fact that telomerase activity and telomere length could be the main actors of normal senescence [8]. Here, we will focus on the senescence of a very important part of the human body, the immune system, and especially the natural killer (NK) cells subsets.
2. NK Cell Biology
NK cells are a very important population of cytotoxic cells linking innate and cellular immunities. They originate from common lymphoid progenitors, like B and T cells, and mature in lymphoid tissues (spleen, bone marrow, tonsil) before entering the blood circulation [9]. A major difference with lymphocytes is their lack of CD3, BCR, or TCR expression. They can be defined as CD3−CD56+CD16+ cells. These cells can react very quickly upon stimulation, faster than T cells, as they can kill directly “missing-self” cells that lack MHC class-I molecules without any need of previous sensitization, antibody binding, or peptide presentation [10]. These cells are very important in antiviral and antitumoral responses. This very fast and efficient ability to kill is still very strictly regulated. The NK cell takes the decision to kill by measuring the balance between signal received by its inhibitory and activating receptors expressed at its surface, inhibition being dominant [11]. These signals are transmitted by 2 families of receptors, the Ig-like and the C-type lectins. Among Ig-like inhibitory receptors there are KIRs (killer-cell immunoglobulin-like receptors) recognizing HLA molecules and sending a strong inhibitory signal [9] and LIR (leucocyte inhibitory receptor) also binding to class-I HLA. Concerning inhibitory C-type lectins, Ly49 and the heterodimer CD94/NKG2A-B, recognizing HLA-E molecules [9] are able to prevent NK cells to kill. The activating receptors are also part of these 2 families. The NCR (natural cytotoxicity receptors) such as CD16 (FcγIIIA), allowing antibody-mediated toxicity, or NKp46, NKp30, and NKp44 belong to the Ig-like family. CD94/NKG2C-E (recognizing HLA-E) as well as NKG2D (recognizing nonclassical HLA) are activating C-type lectins receptors. A part of their role at the interface of innate and adaptive immunities is influenced by the level of CD56 and CD16 they express [12–14]. CD56dimCD16+ are terminally differentiated cytotoxic cells that act more like innate immunity, although they are also cytokine producers. On the other hand CD56brightCD16− cells are less differentiated, cytokine secreting cells able to sustain innate and adaptative immunity [15]. There is a third NK cell subpopulation, CD56−CD16+, originally described in HIV-1+ patients [16], also described in hepatitis B and C [17], with poor proliferative and cytotoxic capacity low cytokine production, and high chemokine production [18]. CD56brightCD16− cells are considered immature cell precursors of CD56dimCD16+ cells [19, 20]. However, it is not clear the relationship between CD56−CD16+ cells and the other NK cell populations.
3. Aging and Overall Immunity
Immunosenescence is defined as the progressive loss of immune functions through aging, and all types of immune cells are concerned. Hematopoietic stem cells (HSCs) become less and less able to renew the blood cells populations due to the shortening of telomeres and the accumulation of DNA lesions due to free radical created during their metabolism [21]. Macrophages lose their bactericidal capacities and their number decreases [22]. Antibody-producing B cell number decreases and leads to a smaller immunoglobulin diversity and affinity [23]. Dendritic cells antigen presenting function decreases with age causing profound changes in cellular immunity [24]. The lymphocytes homeostasis is modified as less and less naïve immune cells are created, and memory populations start to lose their functions, leading to a greater susceptibility to pathogens and cancers [25]. To estimate the immunosenescence, T cells activity is used as a biomarker as nearly all of their functions are modified by aging. They produce less cytokines [26], the repertoire diversity decreases [27], the homeostasis is modified [26], their proliferation is impaired [26], their intracellular signal transduction capability is deregulated [28], and they are less cytotoxic [29].
4. NK Cells in Healthy Aging
During aging, like for lymphocytes, NK cell number functions and phenotype are modulated and modified. Several studies indicate that in the elderly there is an increase in number and a redistribution of NK cell subpopulations, with a decrease of CD56bright population, more immature, and an increase of CD56dim mature cells, particularly those highly differentiated who express CD57, as well as the CD56−CD16+ cells [30–34]. While the CD56bright cells phenotype does not change during healthy aging, the terminally differentiated CD56dim population displays higher levels of HLA-DR and CD95 (Fas) surface expression and a decrease in CD69 (a C-type lectin and early activation antigen) compared to young individuals [30]. When NK cells cytotoxicity was tested in healthy elderly individuals, it was noted that age does not affect it [35], but the increase of CD56dim population in blood does not correlate with an increase of overall cytotoxicity (Figure 1). This supposes an impairment in NK cells cytotoxic activity at the single-cell level although no default was identified in binding to target or in perforin content [36]. The explanation for this intrinsic reduced cytotoxic activity is still under investigation.
Figure 1.
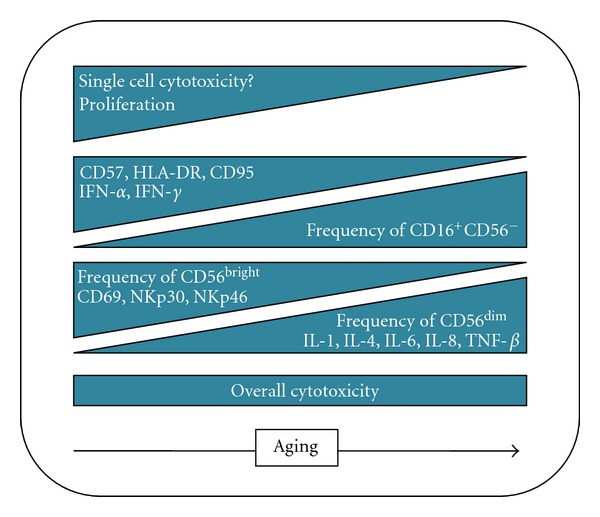
NK cell phenotype and functions in healthy aging. Increase and decrease in the NK-related parameters during aging are depicted. For example, NK cell proliferation is reduced.
One of the most important cytokines for NK cells is IL-2 as it binds adaptive immune response and NK cells. Treating NK cells with cytokines such as IL-2, IL-12, IFN-γ, and IFN-α increases their killing aptitudes and allows them to kill cells usually “NK resistant.” In healthy elderly people, if cytokine stimulation is not impaired, the ability to kill “NK resistant” cells still decreases [35]. IL-2 can also induce NK cells proliferation, but, in elderly people, the intensity of the response varies from a very slight decrease to nearly no proliferation [30]. IL-2 also modifies the NK profile for cytokine secretion. In elderly, compared to young people, IL-2 induction of IFN-γ and IFN-α is decreased whereas IL-1, IL-4, IL-6, IL-8, IL-10, and TNF-α increase [37]. NK cells from elderly also produce less IFN-γ upon IL-2 stimulation whereas perforin and TNF-α were not modified [36]. Almeida-Oliveira et al. recently did a very interesting study about the modulation of NK markers throughout life, from childhood to death [38]. They noticed an expansion of the CD56dim population and shrinkage (in frequency and number) of CD56bright in elderly people, increasing cytotoxic cells while diminishing the NK CD56bright amount of cytokine like IFN-γ, TNF-α, GM-SCF, or IL-10 and IL-13 (Figure 1). Moreover, activated NK cells secrete less IFN-γ. CD56bright cells from children and elderly subjects express more KIR receptors, and in their cohort, most of them express only inhibitory KIR or both inhibitory and activating at the same time. Concerning NCR, they find a decline in the expression of NKp30 and NKp46 in elderly people. NKp30 is known to participate in the crosstalk with dendritic cells leading to the link between innate and adaptive response [39]. They also found a decline in CD94 expression in elderly people in both NK subsets and only in CD56dim cells in children. Interestingly, they found no decline in NKG2D expression neither in children nor in elderly. This could be a form of adaptation to a deficient adaptive immune system as during aging it becomes less effective while during childhood it is mostly naïve regarding antigen encounter.
5. Age-Associated Diseases
Countless diseases are correlated with aging, so in this present paper, we will talk about the most common and immune-related diseases to link up with NK cells. Emerging at alarming speed is certainly Alzheimer's disease (AD) that is associated with aging, with the exception of the early onset congenital form that can occur at any time. Usually this disease is diagnosed after 65 years and is becoming a real problem worldwide, as in 2006, there were 26.6 million patients, and it is predicted to affect 1 in 85 people globally by 2050 [40]. The causes of AD are still unknown, except for the congenital form. Several hypotheses have been proposed to explain the disease. Among them, the amyloid hypothesis postulates that amyloid β (Aβ) accumulation, forming plaques in the brain, is the causative agent of AD [41]. This is supported by several facts: Down syndrome patients got an extra chromosome 21, bearing the Aβ-related gene APP, and develop AD before 40 [42]. Finally, apolipoprotein E 4 gene (APOE4) is a known AD-associated marker as different genotypes in the APOE gene lead to differential accumulation of Aβ in the brain [43]. Nonplaque Aβ oligomer may also be very important as they can bind neuron surface receptor and disrupt synapses [44]. Moreover, one of these receptors may be the prion, responsible for Creutzfeldt-Jakob disease [45]. In 2009, this theory was modified, suggesting that a neighbor of Aβ, and not necessarily the protein itself, could be a major causative agent of the disease. N-APP, a fragment of APP, is adjacent to Aβ and is cleaved from APP by one of the same enzymes. N-APP triggers the self-destruct pathway by binding to a neuronal receptor called DR6 (Death Receptor 6) [46]. DR6 is highly expressed in the part of the brain that is the most affected by AD. It is possible that the N-APP/DR6 pathway might be hijacked in the ageing brain to cause damage. In this model, Aβ only plays a side role. One of the other hypotheses is the Tau hypothesis [47]. Tau protein is a factor that stabilizes microtubules abundant in central nervous system and neurons. It has been shown that hyperphosphorylated tau proteins can cluster together and form neurofibrillary tangles shattering neuron transport system by disintegrating microtubules and leading to the death of the cell [48–50].
Several cancers have been linked to aging so, far and we will discuss here those occurring frequently and in the majority of elderly people. According to the US National Cancer Institute (NIC), after 65 years, there are 10 times more cancer cases than before, and the probability to develop a cancer rises after 40 years of age [51]. The most common cancers in elderly people are prostate, breast, colon, pancreas, bladder, stomach, lung, and rectum. The reasons why it happens mainly in elderly are not clear, but some explanations can be provided. First, some of the cancers develop slowly due to their intrinsic aggressive potential or by its control by the immune system that for some reasons becomes deficient after years or decades of constant immune surveillance. Secondly, with advancing age the immune system starts to weaken, lowering our natural shield against cancer [52]. Thirdly, a longer lifespan is logically associated to higher risks of exposition to carcinogens like those due to pollution, smoking, chemicals, or UV. Finally, the mechanisms involved in cellular detoxification are impaired in aged cells rendering their capacity to prevent (antioxidant levels) and repair DNA damage or protein modifications (e.g., chaperones such as heat shock proteins) that ultimately lead to faulty functions, cell death, or cell transformation [53, 54].
Bone and joint-associated diseases reduce the quality of life of the elderly population rendering them more dependent. One of the most painful age-related diseases is arthritis, which is a generic term covering more than 100 pathologies. The most common form of arthritis is osteoarthritis (or degenerative arthritis) and is predominant in 65+ people (70% of cases in USA). It consists in a loss of cartilage in joints leading to an increased friction between bones inducing pain and a progressing disability [55]. Inflammation of the area is often noted, and the epiphyses damages force a compensatory bone growth that can prevent natural movements. It is related but not caused by aging. The most concerned body parts are hands and knees. Osteoporosis is an age-related condition where bones become porous, leading to an increased risk for fractures [56]. During the disease, bone mineral density is reduced, and the inner architecture is impaired making the bone much more fragile. It occurs in women, after menopause for type 1 osteoporosis while type 2 osteoporosis is prevalent for both genders after 75 (2 : 1 ratio for female). The main problem of this disease is the increased risk of fall plus the fact that fracture can severely disable elderly people [57]. The most common fracture related to osteoporosis is the hip fracture often leading to hip replacement.
Cardiovascular issues like atherosclerosis or high blood pressure are also correlated with age, and the risk of stroke doubles every decade after 55 years of age [58]. It could be partly explained by the fact that the vascular wall loses part of its elasticity, which plays a great role in the control of blood pressure. Moreover, cholesterol levels increase with age, and its accumulation hinders blood flow in small arteries like coronaries. Atherosclerosis is a much more complicated disease as it involves an immunological component [59]. It consists in an accumulation of lipids in the vascular wall generally close to area where there are turbulences in blood flow. It induces an inflammation due to monocyte accumulation and transformation in “foam cells” that make the plaque, or atheroma, growing thicker and thicker. Disruption of this complex may lead to release of thrombogenic molecules in the blood causing clotting. It is usually asymptomatic but often associated with heart attack or sudden death.
To conclude our presentation of age-related diseases, we will discuss about destructive eye diseases like glaucoma, cataract, and age-related macular degeneration (AMD) which are causing severe disruption in daily life. Glaucoma is induced by an increase in pressure leading to the optic nerve compression and destructing of retinal cells and, if left untreated, to blindness [60]. It affects 1 in 200 people aged 50 and younger and 1 in 10 over the age of 80. It is the second cause of blindness worldwide after cataract. Cataract consists in a clouding that develops in the crystalline lens or the lens capsule of the eye and induces a loss of vision that can be complete. The senile cataract, occurring in the elderly, begins with a clouding of the lens followed by its swelling, and finally it shrinks with a complete loss of transparency. Moreover, with time, the cataract cortex liquefies to form a milky white fluid which can cause severe inflammation if the lens capsule leaks [61]. Age is an important risk factor for senile cataract and is often present with AMD which manifests by damages to the retina, in the macula area, leading to a progressive loss of (central) vision. Around 10% of 66–74 people will develop AMD. The prevalence increases to 30% in 75–85 elderly [62].
6. NK Cells in Age-Related Diseases
Among the samples of age-related diseases described in this present paper, NK cells play a role but with differences in their implication in the corresponding diseases (Figure 2). In Alzheimer's disease, NK cells have an increased IL-2-mediated cytotoxic activity that is negatively correlated with cognitive status [63]. Another study from the same team showed that it may be due to a deregulation of protein kinase C (PKC), a regulatory enzyme playing a role in NK exocytosis and cytotoxic response after induction by IL-2 and IFN-β [64]. Cytotoxicity increased by 102% after IL-2 stimulation and 132% after IFN-γ in AD patients compared to controls (healthy elderly and younger people). After IL-2 and IFN-γ stimulation, a physiological decrease in cytosolic PKC concentration was observed in controls but not in AD patients, and cortisol (as immunosuppressor) did not decrease PKC activation in the AD cohort. Finally, IL-2 was shown to induce a greater release of IFN-γ and TNF-α in AD patients compared to controls (healthy elderly), and here also these releases were negatively correlated with cognitive status [65]. Altogether, these data suggest that NK cell cytotoxic activity and overall functionality participate actively in neuroinflammation related to the neurodegeneration observed during AD. It was even suggested to use NK activity as a biomarker of AD progression [66].
Figure 2.
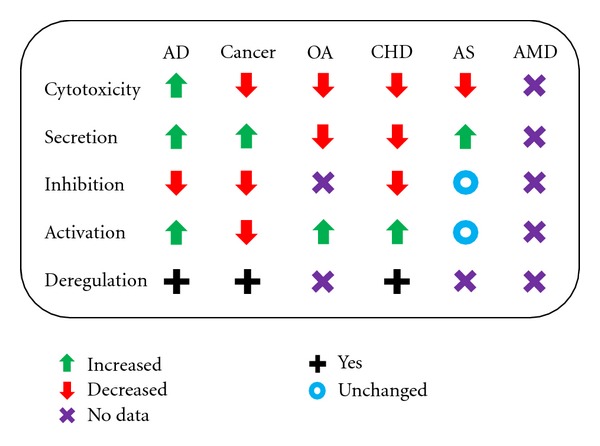
NK cell phenotype and functions in age-associated diseases. The functionality of NK cells in the different diseases is depicted. AD: Alzheimer's disease; OA: osteoarthritis; CHD: cardiovascular/heart diseases; AS: atherosclerosis; AMD: age-related macular degeneration.
Concerning age-related cancers and NK cell function, some evolved an NK escaping process. The perfect example is prostate cancer. Tumor cells can secrete soluble NKG2D that will induce a fake NK response by competing with the true NKG2D for binding on the receptor site and at the same time evade CD8 recognition, as it is expressed on 80% of NK cells, by inhibiting MHC-I expression [10]. This is a fact that shows how cancers evolve to escape from an important population, the NK cells. Concerning pancreatic cancer the major problem is that tumor is surrounded by a fibrotic shield that allows very few cells to reach the core. Among these tumor-infiltrating cells, only a very small number of NK cells were observed [67]. But treating this disease locally with autologous NK cells may be relevant to support for the cure as it was showed that apoptotic pancreatic tumor cells are a very good activator of NK and T cells [68]. Moreover, NK cells stimulated by dendritic cells pulsed with tumor-derived RNA can reverse the resistance of pancreatic carcinoma cells [69]. For colon cancer, NK cells reduced activity was showed to be related to colon cancer metastasis as patient with normal NK response remained free of metastasis whereas low NK response patient showed relapse [70]. NK activity could be used as a marker of colorectal progression and help to identify patients with higher risks of metastasis. In colorectal carcinoma tissue, despite high levels of chemokines and cytokines, tumor infiltrating NK cells are very rare [71]. The tumor escaping mechanism is not yet enlightened but it also promotes the important role of NK cells and seems to be present even in the early stages of the disease. Moreover, NK cells presence in colorectal carcinoma tissue has been negatively correlated with the age of the patients maybe due to an age-related decrease in adherence molecule expression [72]. The decreased expression of activating receptors NCR and DNAM-1 is not only seen in healthy individuals >65 but also in young acute myeloid leukemia patients [31, 73]. This is attributed to increased expression of CD122 and CD155 (DNAM-1 ligands) in leukaemic blasts [34]. Considering the relevance of DNAM-1 in NK recognition/killing of cancer cells, its reduced expression on NK cells from AML patients may represent another mechanism of tumor escape. For stomach cancer, NK cells activity has been correlated to clinic-pathological parameters including tumor size, lymphatic and vascular involvement, and lymph node metastases. The 5-year survival was higher in responding NK group (~95%) compared to nonresponding NK group (72%) [74, 75]. Here also, NK activity could be a good marker for tumor volume and dissemination and prognosis. In lung cancer, a study showed that tumor-infiltrating NK cells are mainly CD56bright and able to secrete cytokines but are unable to kill tumor cells [76]. Cells were CD56bright and CD16−, highly enriched in the tumor, but their cytotoxicity was lower than those from NK cells in peripheral blood. They were also found in the tumor stroma, not in direct contact with tumor cells. Intratumoral NK cells display great phenotypic alterations such as reduced NK cell receptor expression. These defects lead to an impaired degranulation and secretion of cytokines, like IFN-γ. As tumor expresses activating and inhibiting NK cells ligands, it seems that it is a tumoral NK escaping mechanism, and because of this, NK cells are not correlated to the clinical outcome of patients [77].
In osteoarthritis and periprosthetic inflammation, synovial tissue was removed and studied to analyze its immune cell composition [78]. It has been showed that the main infiltrating population was NK cells and that synovial fluid was very rich in NK attractants like CCL-4, CCL-5, CXCL-9, CXCL-10, and chemerin. These NK cells express receptor consistent with an exclusive CD56bright phenotype (Figure 2). They also produce less IFN-γ than peripheral NK cells, which does not prevent further development of the disease as IFN-γ can induce osteoclast differentiation and thus bone repair. This can also have significance in osteoporosis, as elderly possess less IFN-γ secreting NK cells, but this has not been investigated so far.
NK cells have also been linked to coronary heart disease (CHD) [79]. CHD patients had lower NK cytotoxic activity, less CD56dim cells, less CD56bright regulatory cells, and less IFN-γ secreting NK cells than age-matched controls. In idiopathic pulmonary hypertension (PAH), NK cells impairments have also been identified [80]. They revealed that PAH patients' NK cell phenotype was modified. They displayed decreased levels of the activating receptor NKp46 and KIRs, reduced secretion of the cytokine MIP-1β, and a significant impairment in cytolytic function associated with decreased KIR3DL1 expression. These NK cells were more responsive to TGF-β, known to decrease KIR expression. Recent hypotheses suggest the links between innate immunity, TLR, and cardiovascular diseases [81]. During cardiac injuries, some TLR ligands may activate innate immune cells, like NK cells through TLR-2, and thus creating a potentially critical inflammation of the heart.
As discussed, NK cell distribution in tumors is fairly low suggesting mechanisms preventing their recruitment or the possibility that these cells are not the best chemoattractable cells compared to others, at least for certain tissue. In a site like the eye there have been very few reports but the existing ones also suggest a poor presence of NK cells [82]. This rare study also identified large amounts of IgG, IgA, and IgE as well as C1q, C3c, and C3d complement components in the connective stroma and within the new blood vessel walls in AMD patients. A common treatment for cancer and age-related macular degeneration is photodynamic therapy. Other cancers may be treated similarly as suggested by a study showing synergy between photodynamic therapy and other proapoptotic treatments such as FasL and TRAIL [83]. AMD can be subdivided in wet or dry AMD. The wet AMD refers to consequences of choroidal neovascularization. Together with increased levels of cytokines related to innate immunity in the vitreous fluid experiments using KO mice (NK T cell deficient and Jα18 deficient) show significant reduction of the effect of experimentally induced choroidal neovascularization-related diseases [84]. In vitro experiments confirmed the fact NK-like cells could produce VEGF in cocultures with retinal pigment epithelial cells [85]. This suggests that NK-like family may be involved in such diseases (Figure 2). This was partly confirmed by the fact HLA-Cw*0701 allele in combination with the inhibitory KIR AA haplotype was significantly associated with AMD (P = 0.006, OR = 4.35). This genotype combination suggests that NK cells are indeed involved in the pathogenesis of AMD [86].
7. Conclusion
NK cells are important immune cells as they provide a rapid and intense response to challengers. The exact link between NK cell phenotype and their function is still poorly understood and should be pursued to enable a better understanding of diseases, especially in the elderly as this population is showing slow but continuous immune erosion. Immunosenescence of NK cells is recognized more and more as a major player in age-related pathologies and hyporesponsiveness. While the role of NK cells is clearly established in certain pathologies (cancer), their role in other such as autoimmune diseases or immunosurveillance of chronic infectious diseases is less established. As innate cells, NK also participates in the interplay with adaptive immunity by leaving the host with reasonable immune surveillance and cytotoxic activity performed by CD8+ T cells. Thus, altering NK cell functionality naturally, that is, with aging or during diseases, will irreversibly impact on immunity. When both factors are present (aging and diseases), the patients are probably even more at risk. Before NK cells to be used as biomarkers for certain pathologies, as suggested by others, one should first identify NK cell aging as many diseases where NK is involved are seen in the elderly population.
Acknowledgments
This work was supported by the Singapore Agency for Science Technology and Research (A*STAR). This work was partly supported by Grants FIS PI061320 and PS09/00723 from Spanish Ministry of Health (to R. Solana). A. Larbi is an ISAC Scholar.
References
- 1.Hayflick L, Moorhead PS. The serial cultivation of human diploid cell strains. Experimental Cell Research. 1961;25(3):585–621. doi: 10.1016/0014-4827(61)90192-6. [DOI] [PubMed] [Google Scholar]
- 2.Olovnikov AM. Principle of marginotomy in template synthesis of polynucleotides. Doklady Akademii Nauk SSSR. 1971;201(6):1496–1499. [PubMed] [Google Scholar]
- 3.Olovnikov AM. A theory of marginotomy: the incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. Journal of Theoretical Biology. 1973;41(1):181–190. doi: 10.1016/0022-5193(73)90198-7. [DOI] [PubMed] [Google Scholar]
- 4.Blackburn EH, Gall JG. A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena. Journal of Molecular Biology. 1978;120(1):33–53. doi: 10.1016/0022-2836(78)90294-2. [DOI] [PubMed] [Google Scholar]
- 5.Moyzis RK, Buckingham JM, Cram LS, et al. A highly conserved repetitive DNA sequence, (TTAGGG)(n), present at the telomeres of human chromosomes. Proceedings of the National Academy of Sciences of the United States of America. 1988;85(18):6622–6626. doi: 10.1073/pnas.85.18.6622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cooke HJ, Smith BA. Variability at the telomeres of the human X/Y pseudoautosomal region. Cold Spring Harbor Symposia on Quantitative Biology. 1986;51(1):213–219. doi: 10.1101/sqb.1986.051.01.026. [DOI] [PubMed] [Google Scholar]
- 7.Greider CW, Blackburn EH. Identification of a specific telomere terminal transferase activity in tetrahymena extracts. Cell. 1985;43(2 I):405–413. doi: 10.1016/0092-8674(85)90170-9. [DOI] [PubMed] [Google Scholar]
- 8.Harley CB, Futcher AB, Greider CW. Telomeres shorten during ageing of human fibroblasts. Nature. 1990;345(6274):458–460. doi: 10.1038/345458a0. [DOI] [PubMed] [Google Scholar]
- 9.Iannello A, Debbeche O, Samarani S, Ahmad A. Antiviral NK cell responses in HIV infection—I. NK cell receptor genes as determinants of HIV resistance and progression to AIDS. Journal of Leukocyte Biology. 2008;84(1):1–26. doi: 10.1189/jlb.0907650. [DOI] [PubMed] [Google Scholar]
- 10.Vivier E, Raulet DH, Moretta A, et al. Innate or adaptive immunity? The example of natural killer cells. Science. 2011;331(6013):44–49. doi: 10.1126/science.1198687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Farag SS, Fehniger TA, Ruggeri L, Velardi A, Caligiuri MA. Natural killer cell receptors: new biology and insights into the graft-versus-leukemia effect. Blood. 2002;100(6):1935–1947. doi: 10.1182/blood-2002-02-0350. [DOI] [PubMed] [Google Scholar]
- 12.Cooper MA, Fehniger TA, Caligiuri MA. The biology of human natural killer-cell subsets. Trends in Immunology. 2001;22(11):633–640. doi: 10.1016/s1471-4906(01)02060-9. [DOI] [PubMed] [Google Scholar]
- 13.Farag SS, VanDeusen JB, Fehniger TA, Caligiuri MA. Biology and clinical impact of human natural killer cells. International Journal of Hematology. 2003;78(1):7–17. doi: 10.1007/BF02983234. [DOI] [PubMed] [Google Scholar]
- 14.Wendt K, Wilk E, Buyny S, Buer J, Schmidt RE, Jacobs R. Gene and protein characteristics reflect functional diversity of CD56dim and CD56bright NK cells. Journal of Leukocyte Biology. 2006;80(6):1529–1541. doi: 10.1189/jlb.0306191. [DOI] [PubMed] [Google Scholar]
- 15.Fauriat C, Long EO, Ljunggren HG, Bryceson YT. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood. 2010;115(11):2167–2176. doi: 10.1182/blood-2009-08-238469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tarazona R, Casado JG, Delarosa O, et al. Selective depletion of CD56dim NK cell subsets and maintenance of CD56bright NK cells in treatment-naive HIV-1-seropositive individuals. Journal of Clinical Immunology. 2002;22(3):176–183. doi: 10.1023/a:1015476114409. [DOI] [PubMed] [Google Scholar]
- 17.Gonzalez VD, Falconer K, Björkström NK, et al. Expansion of functionally skewed CD56-negative NK cells in chronic hepatitis C virus infection: correlation with outcome of pegylated IFN-α and ribavirin treatment. Journal of Immunology. 2009;183(10):6612–6618. doi: 10.4049/jimmunol.0901437. [DOI] [PubMed] [Google Scholar]
- 18.Björkström NK, Ljunggren HG, Sandberg JK. CD56 negative NK cells: origin, function, and role in chronic viral disease. Trends in Immunology. 2010;31(11):401–406. doi: 10.1016/j.it.2010.08.003. [DOI] [PubMed] [Google Scholar]
- 19.Ouyang Q, Baerlocher G, Vulto I, Lansdorp PM. Telomere length in human natural killer cell subsets. Annals of the New York Academy of Sciences. 2007;1106:240–252. doi: 10.1196/annals.1392.001. [DOI] [PubMed] [Google Scholar]
- 20.Chan A, Hong DL, Atzberger A, et al. CD56bright human NK cells differentiate into CD56dim cells: role of contact with peripheral fibroblasts. Journal of Immunology. 2007;179(1):89–94. doi: 10.4049/jimmunol.179.1.89. [DOI] [PubMed] [Google Scholar]
- 21.Ito K, Hirao A, Arai F, et al. Regulation of oxidative stress by ATM is required for self-renewal of haematopoietic stem cells. Nature. 2004;431(7011):997–1002. doi: 10.1038/nature02989. [DOI] [PubMed] [Google Scholar]
- 22.Stout RD, Suttles J. Immunosenescence and macrophage functional plasticity: dysregulation of macrophage function by age-associated microenvironmental changes. Immunological Reviews. 2005;205:60–71. doi: 10.1111/j.0105-2896.2005.00260.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Han S, Yang K, Ozen Z, et al. Enhanced differentiation of splenic plasma cells but diminished long-lived high-affinity bone marrow plasma cells in aged mice. Journal of Immunology. 2003;170(3):1267–1273. doi: 10.4049/jimmunol.170.3.1267. [DOI] [PubMed] [Google Scholar]
- 24.Uyemura K, Castle SC, Makinodan T. The frail elderly: role of dendritic cells in the susceptibility of infection. Mechanisms of Ageing and Development. 2002;123(8):955–962. doi: 10.1016/s0047-6374(02)00033-7. [DOI] [PubMed] [Google Scholar]
- 25.Hakim FT, Gress RE. Immunosenescence: deficits in adaptive immunity in the elderly. Tissue Antigens. 2007;70(3):179–189. doi: 10.1111/j.1399-0039.2007.00891.x. [DOI] [PubMed] [Google Scholar]
- 26.Voehringer D, Koschella M, Pircher H. Lack of proliferative capacity of human effector and memory T cells expressing killer cell lectinlike receptor G1 (KLRG1) Blood. 2002;100(10):3698–3702. doi: 10.1182/blood-2002-02-0657. [DOI] [PubMed] [Google Scholar]
- 27.Naylor K, Li G, Vallelo AN, et al. The influence of age on T cell generation and TCR diversity. Journal of Immunology. 2005;174(11):7446–7452. doi: 10.4049/jimmunol.174.11.7446. [DOI] [PubMed] [Google Scholar]
- 28.Fülöp T, Jr., Gagné D, Goulet AC, et al. Age-related impairment of p56(lck) and ZAP-70 activities in human T lymphocytes activated through the TcR/CD3 complex. Experimental Gerontology. 1999;34(2):197–216. doi: 10.1016/s0531-5565(98)00061-8. [DOI] [PubMed] [Google Scholar]
- 29.Ouyang Q, Wagner WM, Voehringer D, et al. Age-associated accumulation of CMV-specific CD8+ T cells expressing the inhibitory killer cell lectin-like receptor G1 (KLRG1) Experimental Gerontology. 2003;38(8):911–920. doi: 10.1016/s0531-5565(03)00134-7. [DOI] [PubMed] [Google Scholar]
- 30.Borrego F, Alonso MC, Galiani MD, et al. NK phenotypic markers and IL2 response in NK cells from elderly people. Experimental Gerontology. 1999;34(2):253–265. doi: 10.1016/s0531-5565(98)00076-x. [DOI] [PubMed] [Google Scholar]
- 31.Sanchez-Correa B, Gayoso I, Bergua JM, et al. Decreased expression of DNAM-1 on NK cells from acute myeloid leukemia patients. Immunology and Cell Biology. 2012;90(1):109–115. doi: 10.1038/icb.2011.15. [DOI] [PubMed] [Google Scholar]
- 32.Solana R, Mariani E. NK and NK/T cells in human senescence. Vaccine. 2000;18(16):1613–1620. doi: 10.1016/s0264-410x(99)00495-8. [DOI] [PubMed] [Google Scholar]
- 33.Solana R, Pawelec G, Tarazona R. Aging and Innate Immunity. Immunity. 2006;24(5):491–494. doi: 10.1016/j.immuni.2006.05.003. [DOI] [PubMed] [Google Scholar]
- 34.Gayoso I, Sanchez-Correa B, Campos C, et al. Immunosenescence of human natural killer cells. Journal of Innate Immunity. 2011;3(4):337–343. doi: 10.1159/000328005. [DOI] [PubMed] [Google Scholar]
- 35.Kutza J, Muraskoz DM. Age-associated decline in IL-2 and IL-12 induction of LAK cell activity of human PBMC samples. Mechanisms of Ageing and Development. 1996;90(3):209–222. doi: 10.1016/0047-6374(96)01772-1. [DOI] [PubMed] [Google Scholar]
- 36.Mariani E, Sgobbi S, Meneghetti A, et al. Perforins in human cytolytic cells: the effect of age. Mechanisms of Ageing and Development. 1996;92(2-3):195–209. doi: 10.1016/s0047-6374(96)01829-5. [DOI] [PubMed] [Google Scholar]
- 37.Rink L, Cakman I, Kirchner H. Altered cytokine production in the elderly. Mechanisms of Ageing and Development. 1998;102(2-3):199–209. doi: 10.1016/s0047-6374(97)00153-x. [DOI] [PubMed] [Google Scholar]
- 38.Almeida-Oliveira A, Smith-Carvalho M, Porto LC, et al. Age-related changes in natural killer cell receptors from childhood through old age. Human Immunology. 2011;72(4):319–329. doi: 10.1016/j.humimm.2011.01.009. [DOI] [PubMed] [Google Scholar]
- 39.Walzer T, Dalod M, Robbins SH, Zitvogel L, Vivier E. Natural-killer cells and dendritic cells: “L’union fait la force”. Blood. 2005;106(7):2252–2258. doi: 10.1182/blood-2005-03-1154. [DOI] [PubMed] [Google Scholar]
- 40.Brookmeyer R, Johnson E, Ziegler-Graham K, Arrighi HM. Forecasting the global burden of Alzheimer’s disease. Alzheimer’s and Dementia. 2007;3(3):186–191. doi: 10.1016/j.jalz.2007.04.381. [DOI] [PubMed] [Google Scholar]
- 41.Hardy J, Allsop D. Amyloid deposition as the central event in the aetiology of Alzheimer’s disease. Trends in Pharmacological Sciences. 1991;12(10):383–388. doi: 10.1016/0165-6147(91)90609-v. [DOI] [PubMed] [Google Scholar]
- 42.Nistor M, Don M, Parekh M, et al. Alpha- and beta-secretase activity as a function of age and beta-amyloid in Down syndrome and normal brain. Neurobiology of Aging. 2007;28(10):1493–1506. doi: 10.1016/j.neurobiolaging.2006.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Games D, Adams D, Alessandrini R, et al. Alzheimer-type neuropathology in transgenic mice overexpressing V717F β-amyloid precursor protein. Nature. 1995;373(6514):523–527. doi: 10.1038/373523a0. [DOI] [PubMed] [Google Scholar]
- 44.Lacor PN, Buniel MC, Furlow PW, et al. Aβ oligomer-induced aberrations in synapse composition, shape, and density provide a molecular basis for loss of connectivity in Alzheimer’s disease. Journal of Neuroscience. 2007;27(4):796–807. doi: 10.1523/JNEUROSCI.3501-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Laurén J, Gimbel DA, Nygaard HB, Gilbert JW, Strittmatter SM. Cellular prion protein mediates impairment of synaptic plasticity by amyloid-β oligomers. Nature. 2009;457(7233):1128–1132. doi: 10.1038/nature07761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nikolaev A, McLaughlin T, O’Leary DDM, Tessier-Lavigne M. APP binds DR6 to trigger axon pruning and neuron death via distinct caspases. Nature. 2009;457(7232):981–989. doi: 10.1038/nature07767. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 47.Mudher A, Lovestone S. Alzheimer’s disease—do tauists and baptists finally shake hands? Trends in Neurosciences. 2002;25(1):22–26. doi: 10.1016/s0166-2236(00)02031-2. [DOI] [PubMed] [Google Scholar]
- 48.Goedert M, Spillantini MG, Crowther RA. Tau proteins and neurofibrillary degeneration. Brain Pathology. 1991;1(4):279–286. doi: 10.1111/j.1750-3639.1991.tb00671.x. [DOI] [PubMed] [Google Scholar]
- 49.Iqbal K, Del A, Chen S, et al. Tau pathology in Alzheimer disease and other tauopathies. Biochimica et Biophysica Acta. 2005;1739(2):198–210. doi: 10.1016/j.bbadis.2004.09.008. [DOI] [PubMed] [Google Scholar]
- 50.Chun W, Johnson GVW. The role of tau phosphorylation and cleavage in neuronal cell death. Frontiers in Bioscience. 2007;12(2):733–756. doi: 10.2741/2097. [DOI] [PubMed] [Google Scholar]
- 51.Curado MP, Shin HR, Storm H, Ferlay J, Heanue M, Boyle P. Cancer incidence in five continents. IARC Scientific Publications. 2008;(160):1–837. [PubMed] [Google Scholar]
- 52.Derhovanessian E, Solana R, Larbi A, Pawelec G. Immunity, ageing and cancer. Immunity and Ageing. 2008;5, article 11 doi: 10.1186/1742-4933-5-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Nohl H. Involvement of free radicals in ageing: a consequence or cause of senescence. British Medical Bulletin. 1993;49(3):653–667. doi: 10.1093/oxfordjournals.bmb.a072638. [DOI] [PubMed] [Google Scholar]
- 54.Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chemico-Biological Interactions. 2006;160(1):1–40. doi: 10.1016/j.cbi.2005.12.009. [DOI] [PubMed] [Google Scholar]
- 55.Radin EL, Paul IL. Response of joints to impact loading—I. In vitro wear. Arthritis and Rheumatism. 1971;14(3):356–362. doi: 10.1002/art.1780140306. [DOI] [PubMed] [Google Scholar]
- 56.Pasco JA, Kotowicz MA, Henry MJ, Sanders KM, Nicholson GC. Statin use, bone mineral density, and fracture risk: Geelong osteoporosis study. Archives of Internal Medicine. 2002;162(5):537–540. doi: 10.1001/archinte.162.5.537. [DOI] [PubMed] [Google Scholar]
- 57.Ganz DA, Bao Y, Shekelle PG, Rubenstein LZ. Will my patient fall? Journal of the American Medical Association. 2007;297(1):77–86. doi: 10.1001/jama.297.1.77. [DOI] [PubMed] [Google Scholar]
- 58.Kannel WB, Wolf PA, Benjamin EJ, Levy D. Prevalence, incidence, prognosis, and predisposing conditions for atrial fibrillation: population-based estimates. American Journal of Cardiology. 1998;82(8):2N–9N. doi: 10.1016/s0002-9149(98)00583-9. [DOI] [PubMed] [Google Scholar]
- 59.Hansson GK, Hermansson A. The immune system in atherosclerosis. Nature Immunology. 2011;12(3):204–212. doi: 10.1038/ni.2001. [DOI] [PubMed] [Google Scholar]
- 60.Oliver JE, Hattenhauer MG, Herman D, et al. Blindness and glaucoma: a comparison of patients progressing to blindness from glaucoma with patients maintaining vision. American Journal of Ophthalmology. 2002;133(6):764–772. doi: 10.1016/s0002-9394(02)01403-4. [DOI] [PubMed] [Google Scholar]
- 61.Leske MC, Sperduto RD. The epidemiology of senile cataracts: a review. American Journal of Epidemiology. 1983;118(2):152–165. doi: 10.1093/oxfordjournals.aje.a113625. [DOI] [PubMed] [Google Scholar]
- 62.Feret A, Steinweg S, Griffin HC, Glover S. Macular degeneration: types, causes, and possible interventions. Geriatric Nursing. 2007;28(6):387–392. doi: 10.1016/j.gerinurse.2007.03.003. [DOI] [PubMed] [Google Scholar]
- 63.Solerte SB, Fioravanti M, Severgnini S, et al. Enhanced cytotoxic response of natural killer cells to interleukin-2 in Alzheimer’s disease. Dementia. 1996;7(6):343–348. doi: 10.1159/000106901. [DOI] [PubMed] [Google Scholar]
- 64.Solerte SB, Fioravanti M, Pascale A, Ferrari E, Govoni S, Battaini F. Increased natural killer cell cytotoxicity in Alzheimer’s disease may involve protein kinase C dysregulation. Neurobiology of Aging. 1998;19(3):191–199. doi: 10.1016/s0197-4580(98)00050-5. [DOI] [PubMed] [Google Scholar]
- 65.Solerte SB, Cravello L, Ferrari E, Fioravanti M. Overproduction of IFN-γ and TNF-α from natural killer (NK) cells is associated with abnormal NK reactivity and cognitive derangement in Alzheimer’s disease. Annals of the New York Academy of Sciences. 2000;917:331–340. doi: 10.1111/j.1749-6632.2000.tb05399.x. [DOI] [PubMed] [Google Scholar]
- 66.Prolo P, Chiappelli F, Angeli A, et al. Physiologic modulation of natural killer cell activity as an index of Alzheimer's disease progression. Bioinformation. 2007;1(9):363–366. doi: 10.6026/97320630001363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Ademmer K, Ebert M, Müller-Ostermeyer F, et al. Effector T lymphocyte subsets in human pancreatic cancer: detection of CD8+ CD18+ cells and CD8+ CD103+ cells by multi-epitope imaging. Clinical and Experimental Immunology. 1998;112(1):21–26. doi: 10.1046/j.1365-2249.1998.00546.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Schnurr M, Scholz C, Rothenfusser S, et al. Apoptotic pancreatic tumor cells are superior to cell lysates in promoting cross-priming of cytotoxic T cells and activate NK and γδT cells. Cancer Research. 2002;62(8):2347–2352. [PubMed] [Google Scholar]
- 69.Ziske C, Märten A, Schöttker B, et al. Resistance of pancreatic carcinoma cells is reversed by coculturing NK-like T cells with dendritic cells pulsed with tumor-derived RNA and CA 19-9. Molecular Therapy. 2001;3(1):54–60. doi: 10.1006/mthe.2000.0230. [DOI] [PubMed] [Google Scholar]
- 70.Nüssler NC, Strange BJ, Petzold M, Nussler AK, Glanemann OGM. Reduced NK-cell activity in patients with metastatic colon cancer. Experimental and Clinical Sciences. 2007;6:1–9. [Google Scholar]
- 71.Halama N, Braun M, Kahlert C, et al. Natural killer cells are scarce in colorectal carcinoma tissue despite high levels of chemokines and cytokines. Clinical Cancer Research. 2011;17(4):678–689. doi: 10.1158/1078-0432.CCR-10-2173. [DOI] [PubMed] [Google Scholar]
- 72.Papanikolaou IS, Lazaris AC, Apostolopoulos P, et al. Tissue detection of natural killer cells in colorectal adenocarcinoma. BMC Gastroenterology. 2004;4, article 20 doi: 10.1186/1471-230X-4-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Sanchez-Correa B, Morgado S, Gayoso I, et al. Human NK cells in acute myeloid leukaemia patients: analysis of NK cell-activating receptors and their ligands. Cancer Immunology, Immunotherapy. 2011;60(8):1195–1205. doi: 10.1007/s00262-011-1050-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ishigami S, Natsugoe S, Tokuda K, et al. Prognostic value of intratumoral natural killer cells in gastric carcinoma. Cancer. 2000;88(3):577–583. [PubMed] [Google Scholar]
- 75.Takeuchi H, Maehara Y, Tokunaga E, Koga T, Kakeji Y, Sugimachi K. Prognostic significance of natural killer cell activity in patients with gastric carcinoma: a multivariate analysis. American Journal of Gastroenterology. 2001;96(2):574–578. doi: 10.1111/j.1572-0241.2001.03535.x. [DOI] [PubMed] [Google Scholar]
- 76.Carrega P, Morandi B, Costa R, et al. Natural killer cells infiltrating human nonsmall-cell lung cancer are enriched in CD56brightCD16- cells and display an impaired capability to kill tumor cells. Cancer. 2008;112(4):863–875. doi: 10.1002/cncr.23239. [DOI] [PubMed] [Google Scholar]
- 77.Platonova S, Cherfils-Vicini J, Damotte D, et al. Profound coordinated alterations of intratumoral NK cell phenotype and function in lung carcinoma. Cancer Research. 2011;71(16):5412–5422. doi: 10.1158/0008-5472.CAN-10-4179. [DOI] [PubMed] [Google Scholar]
- 78.Huss RS, Huddleston JI, Goodman SB, Butcher EC, Zabel BA. Synovial tissue-infiltrating natural killer cells in osteoarthritis and periprosthetic inflammation. Arthritis Care and Research. 2010;62(12):3799–3805. doi: 10.1002/art.27751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hak Ł, Mysłiwska J, Wieckiewicz J, et al. NK cell compartment in patients with coronary heart disease. Immunity and Ageing. 2007;4, article 3 doi: 10.1186/1742-4933-4-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ormiston ML, Chang C, Long LL, et al. Impaired natural killer cell phenotype and function in idiopathic and heritable pulmonary arterial hypertension. Circulation. 2012;126(9):1099–1109. doi: 10.1161/CIRCULATIONAHA.112.110619. [DOI] [PubMed] [Google Scholar]
- 81.Spirig R, Tsui J, Shaw S. The emerging role of TLR and innate immunity in cardiovascular disease. Cardiology Research and Practice. 2012;2012:12 pages. doi: 10.1155/2012/181394.181394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Baudouin C, Peyman GA, Fredj-Reygrobellet D, et al. Immunohistological study of subretinal membranes in age-related macular degeneration. Japanese Journal of Ophthalmology. 1992;36(4):443–451. [PubMed] [Google Scholar]
- 83.Granville DJ, Jiang H, McManus BM, Hunt DWC. Fas ligand and TRAIL augment the effect of photodynamic therapy on the induction of apoptosis in JURKAT cells. International Immunopharmacology. 2001;1(9-10):1831–1840. doi: 10.1016/s1567-5769(01)00107-2. [DOI] [PubMed] [Google Scholar]
- 84.Sonoda KH, Ismbashi T. Association of intraocular neovascular disease and innate immune response. Fukuoka Igaku Zasshi. 2008;99(7):137–143. [PubMed] [Google Scholar]
- 85.Hijioka K, Sonoda KH, Tsutsumi-Miyahara C, et al. Investigation of the role of CD1d-restricted invariant NKT cells in experimental choroidal neovascularization. Biochemical and Biophysical Research Communications. 2008;374(1):38–43. doi: 10.1016/j.bbrc.2008.06.080. [DOI] [PubMed] [Google Scholar]
- 86.Goverdhan SV, Khakoo SI, Gaston H, Chen X, Lotery AJ. Age-related macular degeneration is associated with the HLA-cw*0701 genotype and the natural killer cell receptor AA haplotype. Investigative Ophthalmology and Visual Science. 2008;49(11):5077–5082. doi: 10.1167/iovs.08-1837. [DOI] [PMC free article] [PubMed] [Google Scholar]