

Abstract
The aim of this study was to investigate the interactions between cytokinin, sugar repression, and light in the senescence-related decline in photosynthetic enzymes of leaves. In transgenic tobacco (Nicotiana tabacum) plants that induce the production of cytokinin in senescing tissue, the age-dependent decline in NADH-dependent hydroxypyruvate reductase (HPR), ribulose-1,5-bisphosphate carboxylase/oxygenase, and other enzymes involved in photosynthetic metabolism was delayed but not prevented. Glucose (Glc) and fructose contents increased with leaf age in wild-type tobacco and, to a greater extent, in transgenic tobacco. To study whether sugar accumulation in senescing leaves can counteract the effect of cytokinin on senescence, discs of wild-type leaves were incubated with Glc and cytokinin solutions. The photorespiratory enzyme HPR declined rapidly in the presence of 20 mm Glc, especially at very low photon flux density. Although HPR protein was increased in the presence of cytokinin, cytokinin did not prevent the Glc-dependent decline. Illumination at moderate photon flux density resulted in the rapid synthesis of HPR and partially prevented the negative effect of Glc. Similar results were obtained for the photosynthetic enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase. It is concluded that sugars, cytokinin, and light interact during senescence by influencing the decline in proteins involved in photosynthetic metabolism.
During the process of leaf senescence, chlorophyll and photosynthetic proteins are degraded (Humbeck et al., 1996). There are several factors that can accelerate or delay this breakdown of the photosynthetic apparatus.
Whereas the plant growth regulators ABA and ethylene accelerate the symptoms of senescence (Smart, 1994), exogenous application of cytokinins inhibits the degradation of chlorophyll and photosynthetic proteins (Richmond and Lang, 1957; Badenoch-Jones et al., 1996). Senescence is also delayed in transgenic plants producing cytokinin by expression of a bacterial gene encoding IPT, the enzyme catalyzing the first step of cytokinin synthesis (Smart et al., 1991; Gan and Amasino, 1995).
Low light intensities or darkness results in the reduced expression of light-dependent genes and the disappearance of photosynthetic proteins and chlorophyll (Thomas, 1978). Since phytochrome acts as the light receptor for the expression of many photosynthetic genes, a lower red/far-red ratio reaching the lower leaves of a plant can also accelerate the senescence of these leaves (Rousscaux et al., 1996).
In nonsenescent leaves sugar accumulation can lead to a decline in chlorophyll and photosynthetic proteins (Stitt et al., 1990; von Schaewen et al., 1990; Krapp et al., 1991; Krapp and Stitt, 1994). Glc and Suc repress the transcription of photosynthetic genes (Sheen, 1990), probably acting via hexokinase as a sugar sensor (Jang and Sheen, 1994; Jang et al., 1997). The involvement of sugar-mediated repression of genes in the regulation of natural senescence is less clear (Feller and Fischer, 1994). The concentration of leaf sugars can increase during leaf senescence (Crafts-Brandner et al., 1984), and accumulation of sugars, induced by removal of sinks or phloem interruption, can both accelerate and delay senescence (e.g. Crafts-Brandner et al., 1984; Fröhlich and Feller, 1991). The response of leaves to the accumulation of sugars must therefore also depend on other factors, such as the C:N status of the leaf (Paul and Driscoll, 1997), light (Dijkwel et al., 1997) and plant growth regulators (Koch, 1996). For example, it has been suggested that cytokinin, in addition to delaying senescence, could block some of the responses to sugars (Jang et al., 1997).
In this paper we have studied the interactions of cytokinins, light, and sugars during senescence in transgenic tobacco (Nicotiana tabacum L.) plants with autoregulated synthesis of cytokinin (Gan and Amasino, 1995). The transgenic tobacco plants express the gene for IPT under control of the senescence-specific SAG 12 promoter (Lohman et al., 1994). This promoter is activated at the onset of senescence, leading to the synthesis of cytokinin. Because of the inhibition of senescence by cytokinin, the promoter is actively attenuated. This results in an autoregulatory loop, preventing the overproduction of cytokinin and confining expression solely to those tissues that have initiated senescence. Apart from a delay in senescence, these plants therefore develop normally (Gan and Amasino, 1995). To obtain more detailed information on how sugars and cytokinin interact, we incubated leaf discs of wild-type tobacco with Glc and cytokinin solutions. We focused on the effect on NADH-dependent HPR, an enzyme of the photorespiratory C cycle that catalyzes the reduction of hydroxypyruvate to glycerate. Because recycling of C in the photorespiratory cycle is essential for photosynthetic metabolism, photosynthesis depends on the activity of HPR (Murray et al., 1989). In cotyledons of curcurbits the synthesis of HPR is induced by cytokinin (Chen and Leisner, 1985; Andersen et al., 1996) and light (Bertoni and Becker, 1993), and the activity of HPR decreases during senescence (De Bellis and Nishimura, 1991). In this study we show that HPR protein declines during incubation with Glc and provide evidence that this decline is modulated by cytokinin and light.
MATERIALS AND METHODS
Wild-type tobacco (Nicotiana tabacum L. cv Wisconsin) and transgenic tobacco homozygous for the chimeric gene PSAG12-IPT (TTI) (Gan and Amasino, 1995) were grown in high-nutrient compost (M3; Fisons, Ipswich, UK) in a greenhouse in natural daylight supplemented with tungsten/halogen lamps (200 μmol m−2 s−1 PFD). The plants were fertilized weekly with a Hoagland solution containing 12 mm nitrate. All experiments were carried out on leaves taken from plants that had initiated flowering.
Gas Exchange
H2O and CO2 were measured using an IR gas analyzer (LCA3, Analytical Development Company, Ltd., Hoddesdon, UK) equipped with a modified Parkinson leaf chamber to allow temperature control and illumination. Gas composition was controlled by mixing O2, N2, and air containing 5% CO2 (British Oxygen Company, Guildford, UK), using mass flow controllers (Brooks Instruments B.V., Veenendaal, The Netherlands). Air humidity was controlled using a temperature trap and was maintained at 70 ± 5% RH within the leaf chamber. PFD was maintained at 250 μmol m−2 s−1, and leaf temperature was maintained at 25 ± 0.5°C.
Incubation of Leaf Discs
Leaf discs were floated in Petri dishes on solutions containing Glc, sorbitol, iPR, or combinations thereof. All solutions were adjusted to pH 7.0. The Petri dishes were kept in controlled conditions at 23°C and cycles of 16 h of light (20 or 250 μmol m−2 s−1) and 8 h of darkness.
Quantification of Protein and Chlorophyll
Protein was extracted in 50 mm Hepes-KOH (pH 7.4), 5 mm MgCl2, 1 mm EDTA, 1 mm EGTA, 10% (v/v) glycerol, 0.1% (v/v) Triton X-100, and 5 mm DTT, and determined with the Bio-Rad protein assay, according to Bradford (1976). Chlorophyll was extracted in 80% acetone and quantified according to Lichtenthaler and Wellburn (1983).
Quantification of Sugars
Samples for sugar determinations were harvested in the morning, between 3 and 4 h into the photoperiod. Sugars were extracted in 80% ethanol and determined enzymatically as described by Scholes et al. (1994).
Western Blotting
Proteins were extracted in 200 mm Bicine-KOH (pH 9.0), 4.5 mm DTT, and 1% (w/v) SDS. Extracts were boiled for 90 s with equal volumes of solubilization buffer (62.5 mm Tris, 20% [v/v] glycerol, 2.5% [w/v] SDS, and 5% [v/v] 2-mercaptoethanol, pH 6.8). For SDS-PAGE, equal leaf areas (3.3 mm2) were loaded and separated onto gels containing 10% acrylamide. Proteins were transferred onto a PVDF membrane (Immobilon-P, Millipore) and probed with antisera raised to NADH-dependent HPR of spinach (Spinacia oleracea L.) (Kleczkowski et al., 1990), Rubisco of pea (Pisum sativum L.), plastidic Fru-1,6-bisphosphatase of spinach, NADP-dependent glyceraldehyde-3-phosphate dehydrogenase of spinach, NADP-dependent malate-dehydrogenase of pea, and aldolase. A peroxidase-conjugated secondary antibody was used, and immunoreactive bands were visualized with an enhanced chemoluminescence kit (Amersham). For quantification of proteins, the peak volumes of bands were determined by video imaging.
RESULTS
Effects of Endogenous Production of Cytokinin on the Senescence-Related Decline in Enzymes Involved in Photosynthetic Metabolism
The effect of cytokinin on the senescence-related decline in photosynthetic enzymes was studied with transgenic tobacco (TTI) expressing a bacterial gene encoding IPT under control of the senescence-specific SAG12 promoter (Gan and Amasino, 1995). As reported previously, the transgenic plants developed normally until the early stages of flowering; at this point the wild-type plants showed senescence-related yellowing of the lower leaves, whereas these leaves remained largely green in the transgenic plants. These differences were reflected in chlorophyll and protein contents. In the wild type, chlorophyll and protein decreased strongly as the leaves aged (Table I). In transgenic tobacco plants this decrease was delayed considerably, although chlorophyll and protein still declined in older leaves. Rates of photosynthetic CO2 assimilation were similar in young leaves of the wild type and of the transgenic tobacco plants (Fig. 1). In mature leaves (leaves four to eight from the bottom), however, the transgenic tobacco plants maintained considerably higher rates of photosynthesis than the wild type.
Table I.
Contents of chlorophyll and protein in leaves of different age from wild-type tobacco (WT) and tobacco with autoregulated production of cytokinin (TTI)
| Leaf Position from Bottom | Chlorophyll
|
Protein
|
||
|---|---|---|---|---|
| WT | TTI | WT | TTI | |
| mg m−2 | g m−2 | |||
| 11 (Young) | 314 ± 17 | 335 ± 9 | 3.16 ± 0.08 | 3.66 ± 0.21 |
| 6 (Mature) | 227 ± 8 | 253 ± 13 | 1.35 ± 0.08 | 1.61 ± 0.06 |
| 2 (Senescing) | 70 ± 5 | 177 ± 6 | 0.50 ± 0.02 | 1.13 ± 0.05 |
Data are means ± se of three to four samples.
Figure 1.

Rates of photosynthesis in leaves of different age from wild-type tobacco plants (○) and tobacco plants with autoregulated production of cytokinin (•). Data are means ± se of three measurements.
Whereas HPR and Rubisco proteins in the wild type declined drastically with leaf age, transgenic tobacco plants maintained higher contents of these enzymes (Fig. 2, A and B). For Rubisco, the effect of the autoregulated production of cytokinin was most apparent in mature leaves taken from the middle of the plant. HPR protein almost disappeared in the lower leaves of the wild type, but not in the transgenic tobacco plants. Other enzymes involved in photosynthesis (plastidic Fru-1,6-bisphosphatase, plastidic aldolase, NADP-dependent glyceraldehyde-3-phosphate dehydrogenase, and NADP-dependent malate dehydrogenase) showed effects similar to HPR (Fig. 2A).
Figure 2.

A, Western blots for NADH-dependent HPR, the large subunit of Rubisco, plastidic Fru-1,6-bisphosphatase, plastidic aldolase, NADP-dependent glyceraldehyde-3-phosphate dehydrogenase, and NADP-dependent malate dehydrogenase in leaves of different age from wild-type tobacco plants (WT) and tobacco plants with autoregulated production of cytokinin (TTI). B, Relative content of HPR and Rubisco as determined from western blots. Data are means ± se of three samples.
As a senescence-related loss of chlorophyll or protein can be induced by increased sugar contents, soluble sugars were measured in the leaves of wild-type and transgenic tobacco plants. Leaf contents of Glc and Fru increased with leaf age (Fig. 3), and this increase was accompanied by lower starch contents in old leaves (data not shown). Both hexoses accumulated to a higher extent in the transgenic tobacco than in the wild-type plants. Suc, on the other hand, was not affected by the autoregulated synthesis of cytokinin and was lower in mature leaves than in young and senescing leaves. We tested the hypothesis that the accumulation of hexoses in old leaves caused the repression of photosynthetic genes, and that cytokinin and sugars interact in the regulation of senescence.
Figure 3.

Sugar contents in leaves of different age from wild-type tobacco (WT, □) and tobacco with autoregulated production of cytokinin (TTI, ▨). Data are means ± se of three samples.
Effects of Exogenous Supply of Cytokinin and Glc on HPR
The effects of cytokinin and Glc were studied by incubating leaf discs of wild-type tobacco with the cytokinin iPR and Glc. The discs were routinely illuminated for 16 h per day at a very low PFD (20 μmol m−2 s−1). At this PFD, sugar repression by internally accumulating sugars that could occur at higher PFDs can be prevented (Krapp et al., 1991). Complete darkness, on the other hand, would have caused starvation of the tissue. After 10 d of incubation with 50 mm Glc, discs from mature and senescing leaves showed a strong loss of chlorophyll (Table II), whereas discs from young leaves were affected to a lesser extent. Incubation with sorbitol as a control did not result in significantly decreased chlorophyll contents in discs from young and mature leaves. However, sorbitol did induce the death of discs from senescing leaves. All further experiments were performed with mature (fully expanded) leaves.
Table II.
Contents of chlorophyll in leaf discs from wild-type leaves of different age
| Leaf Position from Bottom | Chlorophyll
|
||
|---|---|---|---|
| Water | Glc | Sorbitol | |
| mg m−2 | |||
| 11 (Young) | 323 ± 9 | 220 ± 2 (68%) | 296 ± 8 (92%) |
| 6 (Mature) | 165 ± 1 | 94 ± 3 (57%) | 151 ± 8 (92%) |
| 2 (Senescing) | 38 ± 5 | 21 ± 1 (55%) | n.d.a |
The discs were incubated for 10 d at very low PFD (20 μmol m−2 s−1) with water, 50 mm Glc, or 50 mm sorbitol. Data are means ± se of three to four samples. The relative contents compared with the water controls are given in parentheses.
n.d., Not determined.
Incubation of leaf discs with 50 mm Glc, but not with 50 mm sorbitol (data not shown), caused a strong reduction in HPR protein compared with the water control (Fig. 4). We tested whether this decline could be overcome by the inclusion of cytokinin in the incubation medium. Figure 4 shows that the content of HPR protein increased with concentrations of iPR between 10−7 and 10−5 m, but decreased when the concentration was as high as 10−4 m. Despite the positive effect of iPR on HPR protein, iPR did not prevent the Glc-dependent (20 or 50 mm Glc) decline in HPR. Similar results were obtained with the cytokinins N6-[Δ2]isopentenyladenine and zeatin riboside (data not shown). Although cytokinin clearly did not affect the final content of HPR in the presence of Glc, we tested whether it may have altered the time course of the Glc-dependent decrease. Leaf discs were harvested over a period of 10 d. In the very low PFD necessary for this experiment, the content of HPR protein decreased even in the absence of Glc (Fig. 5); however, incubation with 50 mm Glc clearly accelerated the drop in HPR, which occurred before a visible loss of chlorophyll. In the absence of Glc, iPR delayed the decline in HPR and even caused a slight increase in HPR between d 2 and 4, but it did not delay the Glc-dependent drop in HPR.
Figure 4.

Relative content of NADH-dependent HPR in leaf discs of wild-type tobacco plants after incubation for 10 d at very low PFD (20 μmol m−2 s−1) with different concentrations of Glc and iPR.
Figure 5.

Time course of the relative content of NADH-dependent HPR in leaf discs of wild-type tobacco plants after incubation at very low PFD (20 μmol m−2 s−1) ± 50 mm Glc and ± 30 μm iPR. Data are means ± se of three samples. ○, H2O; □, iPR; •, Glc; and ▪, Glc plus iPR.
The expression of the HPR gene is known to be regulated by light (Bertoni and Becker, 1993). Therefore, we also employed a higher PFD to test whether sugar repression of HPR also occurs when the gene is induced by light. In moderate PFD, approximately the PFD at which the plants were grown (250 μmol m−2 s−1), HPR declined in water or in Glc solution. iPR prevented this loss of HPR when it was included in water, but not when Glc was present (Fig. 6A). All four treatments led to a visible loss of chlorophyll (data not shown) and the accumulation of sugars (Fig. 6B). Compared with d 0, the Glc content rose 78-fold during 6 d in water. After incubation with Glc all sugar contents were approximately twice as high as after incubation without Glc. Apart from a slightly lower Suc content, the presence of iPR had no effect on the accumulation of sugars. To determine if the reduction in HPR in water was due to sugar accumulation or general aging of the tissue, leaf discs were transferred for 2 d into continuous, moderate light, after they had been incubated for 6 d in the low-light conditions described earlier. In this experiment the transfer into a higher PFD resulted in a rapid accumulation of HPR in the absence of Glc (Fig. 7, A and B). This indicates that the leaf discs were still capable of de novo protein synthesis after 6 d of incubation, and that the decline in HPR observed in water (Fig. 6) was not due to aging of the tissue but to accumulation of sugars. Even in the presence of Glc, HPR increased slightly (1.5-fold) after the transfer into moderate PFD, whereas longer incubation in low light led to a further decline in HPR. The large subunit of Rubisco showed a similar response to HPR (Fig. 7A). In both low and moderate PFD, iPR had a positive effect on the content of Rubisco protein in the absence of Glc, but incubation with Glc resulted in a decline in Rubisco that was not prevented by iPR.
Figure 6.

A, Relative content of NADH-dependent HPR in leaf discs of wild-type tobacco plants at d 0 and after incubation for 6 d at moderate PFD (250 μmol m−2 s−1) ± 50 mm Glc and ± 30 μm iPR. B, Sugar contents in the leaf discs. Data are means ± se of three samples.
Figure 7.

Contents of NADH-dependent HPR and the large subunit of Rubisco in leaf discs of wild-type tobacco plants after transfer from very low PFD (20 μmol m−2 s−1) into moderate PFD (250 μmol m−2 s−1). A, Western blots for discs incubated for 6 d at very low PFD (LL) or for 6 d at very low PFD plus 2 d at continuous, moderate PFD (ML) ± 50 mm Glc and ± 30 μm iPR. B, Relative content of HPR as determined from western blots after incubation for 6 d at very low PFD (▨) for 8 d at very low PFD (▪), and for 6 d at very low PFD plus 2 d in continuous moderate PFD (□). Data are means ± se of three samples.
DISCUSSION
Inhibition of the Decline in HPR and Other Enzymes by Autoregulated Production of Cytokinin
Senescence can be retarded considerably by the autoregulated production of cytokinin in tobacco leaves, resulting in a prolonged photosynthetic life span (Gan and Amasino, 1995). The basis for the maintained photosynthetic activity in old leaves (Fig. 1) is the delayed decline in chlorophyll and in enzymes involved in photosynthetic metabolism (Fig. 2). The production of cytokinin in these leaves could directly influence the amount of the enzymes by a variety of mechanisms that influence the rate of protein synthesis or degradation. It has, for example, been demonstrated that cytokinin treatment can increase the activities of Rubisco, Fru-1,6-bisphosphatase, NADP-dependent glyceraldehyde-3-phosphate dehydrogenase, NADP-dependent malate dehydrogenase (Harvey et al., 1974; Feierabend and de Boer, 1978), and HPR (Chen and Leisner, 1985). Although cytokinin accumulates in old leaves of TTI (W. Jordi, personal communication), the senescence-related decline in these proteins could not be prevented completely by the autoregulated production of cytokinin. This could be due to the fact that cytokinin is only produced in TTI when the process of senescence has already started, or to the fact that other factors in addition to cytokinin regulate senescence. For example, the high hexose contents in the old leaves of TTI (Fig. 3) could have counteracted the effect of cytokinin by repressing the genes for photosynthetic enzymes, such as Rubisco, Fru-1,6-bisphosphatase, and NADP-dependent glyceraldehyde-3-phosphate dehydrogenase (Stitt et al., 1990; Krapp et al., 1991).
Involvement of Cytokinin, Sugar Repression, and Light in the Regulation of HPR
The possibility that an accumulation of sugars can override the positive effect of cytokinin on photosynthetic proteins such as HPR, was further investigated by studying the interactions of sugars and cytokinin in vitro.
HPR has not previously been reported to be affected by sugars. Our results clearly show that the presence of Glc accelerated the decline in HPR when the PFD was very low (Figs. 4 and 5). Since the expression of HPR is induced by light (Bertoni and Becker, 1993), it was important to investigate if sugars could also repress light induction, as shown for genes encoding plastocyanin and chlorophyll a/b-binding protein (Dijkwel et al., 1997). In our experiments we observed that the light-induced increase in HPR was lower in the presence than in the absence of Glc (Fig. 7), and that the accumulation of endogenous sugars in moderate light resulted in a decrease in HPR protein (Fig. 6).
The positive effect of cytokinin on HPR was independent of light (Figs. 4, 5, and 7). Cytokinin can regulate the expression of HPR by acting at the level of transcription and perhaps posttranscriptionally (Andersen et al., 1996). In very low light Glc clearly counteracted the positive effect of cytokinin on HPR. However, when the PFD was increased, cytokinin could prevent the decline in HPR observed in water controls. This decline was probably brought about by the internal accumulation of sugars that occurred in high-light conditions (Fig. 6B). Only when the sugar contents increased drastically in the presence of exogenously supplied sugars did cytokinin have no effect.
Light is a major factor in the regulation of HPR synthesis. It has a stronger effect than cytokinin and can partially prevent the decline in HPR induced by sugars. Consequently, in the light-induced state, sugars do not completely counteract the positive effect of cytokinin. Identification of the level (transcription, mRNA stability, protein synthesis or breakdown) at which sugars act in the reduction in the HPR protein requires further investigation.
Interaction of Cytokinin, Sugar Repression, and Light with Senescence
Leaf senescence requires dismantling of the photosynthetic apparatus and mobilization of the N contained in photosynthetic proteins. Our results show that this process can be regulated by sugars, cytokinin, and light, and that these factors interact. A scheme summarizing these interactions is shown in Figure 8. Cytokinin production slows down the process of senescence. However, accumulation of sugars during senescence, due to the breakdown of accumulated starch or the preferential export of N2 from the leaf, can block the effect of cytokinin, especially in low light. Light also interacts with sugars by partially reducing the effect of sugar repression. The genetic control of senescence-related processes will clearly require modification of several regulatory mechanisms if functional stay-green leaves are to be achieved.
Figure 8.

Interaction of cytokinin, sugar repression, and light in the regulation of senescence. ⊖, Inhibition of senescence; 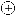 , acceleration of senescence;
, acceleration of senescence; 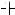 , block of the effect of cytokinin; and
, block of the effect of cytokinin; and 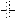 , partial block of the effect of sugars.
, partial block of the effect of sugars.
ACKNOWLEDGMENTS
We are grateful to R.M. Amasino (University of Wisconsin, Madison) for providing the transgenic tobacco and to the laboratory of R. Scheibe (University of Osnabrück) and U. Sonnewald (IPK, Gatersleben, Germany) for providing antisera. We would also like to thank B.C. Jarvis (University of Sheffield) for advice on the application of cytokinins.
Abbreviations:
- HPR
hydroxypyruvate reductase
- iPR
N6-[Δ2-isopentenyl]adenosine
- IPT
isopentenyl transferase
- PFD
photon flux density
LITERATURE CITED
- Andersen BR, Jin G, Chen R, Ertl JR, Chen C-M. Transcriptional regulation of hydroxypyruvate reductase gene expression by cytokinin in etiolated pumpkin cotyledons. Planta. 1996;198:1–5. doi: 10.1007/BF00197579. [DOI] [PubMed] [Google Scholar]
- Badenoch-Jones J, Parker CW, Letham DS, Singh S. Effect of cytokinins supplied via the xylem at multiples of endogenous concentrations on transpiration and senescence in derooted seedlings of oat and wheat. Plant Cell Environ. 1996;19:504–516. [Google Scholar]
- Bertoni GP, Becker WM. Effects of light fluence and wavelength on expression of the gene encoding cucumber hydroxypyruvate reductase. Plant Physiol. 1993;103:933–941. doi: 10.1104/pp.103.3.933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Chen C-M, Leisner SM. Cytokinin-modulated gene expression in excised pumpkin cotyledons. Plant Physiol. 1985;77:99–103. doi: 10.1104/pp.77.1.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crafts-Brandner SJ, Below FE, Wittenbach VA, Harper JE, Hageman RH. Differential senescence of maize hybrids following ear removal. II. Selected leaf. Plant Physiol. 1984;74:368–373. doi: 10.1104/pp.74.2.368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Bellis L, Nishimura M. Development of enzymes of the glyoxylate cycle during senescence of pumpkin cotyledons. Plant Cell Physiol. 1991;32:555–561. [Google Scholar]
- Dijkwel PP, Huijser C, Weisbeck PJ, Chua N-H, Smeekens SCM. Sucrose control of phytochrome A signaling in Arabidopsis. Plant Cell. 1997;9:583–595. doi: 10.1105/tpc.9.4.583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feierabend J, de Boer J. Comparative analysis of the action of cytokinin and light on the formation of ribulose-bisphosphate carboxylase and plastid biogenesis. Planta. 1978;142:75–82. doi: 10.1007/BF00385123. [DOI] [PubMed] [Google Scholar]
- Feller U, Fischer A. Nitrogen metabolism in senescing leaves. Crit Rev Plant Sci. 1994;13:241–273. [Google Scholar]
- Fröhlich V, Feller U. Effect of phloem interruption on senescence and protein remobilization in the flag leaf of field-grown wheat. Biochem Physiol Pflanzen. 1991;187:139–147. [Google Scholar]
- Gan S, Amasino RM. Inhibition of leaf senescence by autoregulated production of cytokinin. Science. 1995;270:1986–1988. doi: 10.1126/science.270.5244.1986. [DOI] [PubMed] [Google Scholar]
- Harvey BMR, Lu BC, Fletcher RA. Benzyladenine accelerates chloroplast differentiation and stimulates photosynthetic enzyme activity in cucumber cotyledons. Can J Bot. 1974;52:2581–2586. [Google Scholar]
- Humbeck K, Quast S, Krupinska K. Functional and molecular changes in the photosynthetic apparatus during senescence of flag leaves from field-grown barley plants. Plant Cell Environ. 1996;19:337–344. [Google Scholar]
- Jang J-C, León P, Zhou L, Sheen J. Hexokinase as a sugar sensor in higher plants. Plant Cell. 1997;9:5–19. doi: 10.1105/tpc.9.1.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang J-C, Sheen J. Sugar sensing in higher plants. Plant Cell. 1994;6:1665–1679. doi: 10.1105/tpc.6.11.1665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleczkowski LA, Edwards GE, Blackwell RD, Lea PJ, Givan CV. Enzymology of the reduction of hydroxypyruvate and glyoxylate in a mutant of barley lacking peroxisomal hydroxypyruvate reductase. Plant Physiol. 1990;94:819–825. doi: 10.1104/pp.94.2.819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch KE. Carbohydrate-modulated gene expression in plants. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:509–540. doi: 10.1146/annurev.arplant.47.1.509. [DOI] [PubMed] [Google Scholar]
- Krapp A, Quick WP, Stitt M. Ribulose-1,5-bisphosphate carboxylase-oxygenase, other Calvin-cycle enzymes, and chlorophyll decrease when glucose is supplied to mature spinach leaves via the transpiration stream. Planta. 1991;186:58–69. doi: 10.1007/BF00201498. [DOI] [PubMed] [Google Scholar]
- Krapp A, Stitt M. Influence of high carbohydrate content on the activity of plastidic and cytosolic isoenzyme pairs in photosynthetic tissues. Plant Cell Environ. 1994;17:861–866. [Google Scholar]
- Lichtenthaler HK, Wellburn AR. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc Trans. 1983;603:591–592. [Google Scholar]
- Lohman KN, Gan S, John MC, Amasino RM. Molecular analysis of natural leaf senescence in Arabidopsis thaliana. Physiol Plant. 1994;92:322–328. [Google Scholar]
- Murray AJS, Blackwell RD, Lea PJ. Metabolism of hydroxypyruvate in a mutant of barley lacking NADH-dependent hydroxypyruvate reductase, an important photorespiratory enzyme activity. Plant Physiol. 1989;91:395–400. doi: 10.1104/pp.91.1.395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paul MJ, Driscoll SP. Sugar repression of photosynthesis: the role of carbohydrates in signalling nitrogen deficiency through source:sink imbalance. Plant Cell Environ. 1997;20:110–116. [Google Scholar]
- Richmond AE, Lang A. Effect of kinetin on protein content and survival of detached Xanthium leaves. Science. 1957;125:650–651. [Google Scholar]
- Rousscaux MC, Hall AJ, Sánchez RA. Far-red enrichment and photosynthetically active radiation level influence leaf senescence in field-grown sunflower. Physiol Plant. 1996;96:217–224. [Google Scholar]
- Scholes JD, Lee PJ, Horton P, Lewis DH. Invertase: understanding changes in the photosynthetic and carbohydrate metabolism of barley leaves infected with powdery mildew. New Phytol. 1994;126:213–222. [Google Scholar]
- Sheen J. Metabolic repression of transcription in higher plants. Plant Cell. 1990;2:1027–1038. doi: 10.1105/tpc.2.10.1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smart CM (1994) Gene expression during leaf senescence. New Phytol 126: 419–448 [DOI] [PubMed]
- Smart CM, Scofield SR, Bevan MW, Dyer TA. Delayed leaf senescence in tobacco plants transformed with tmr, a gene for cytokinin production in Agrobacterium. Plant Cell. 1991;3:647–656. doi: 10.1105/tpc.3.7.647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stitt M, von Schaewen A, Willmitzer L. “Sink” regulation of photosynthetic metabolism in transgenic tobacco plants expressing yeast invertase in their cell wall involves a decrease of the Calvin-cycle enzymes and an increase of glycolytic enzymes. Planta. 1990;183:40–50. doi: 10.1007/BF00197565. [DOI] [PubMed] [Google Scholar]
- Thomas H. Enzymes of nitrogen mobilization in detached leaves of Lolium temulentum during senescence. Planta. 1978;142:161–169. doi: 10.1007/BF00388207. [DOI] [PubMed] [Google Scholar]
- Von Schaewen A, Stitt M, Schmidt R, Sonnewald U, Willmitzer L. Expression of a yeast-derived invertase in the cell wall of tobacco and Arabidopsis plants leads to accumulation of carbohydrate and inhibition of photosynthesis and strongly influences growth and phenotype of transgenic tobacco plants. EMBO J. 1990;9:3033–3044. doi: 10.1002/j.1460-2075.1990.tb07499.x. [DOI] [PMC free article] [PubMed] [Google Scholar]