

Abstract
Endocannabinoids are key modulators of synaptic function. By activating cannabinoid receptors expressed in the central nervous system, these lipid messengers can regulate several neural functions and behaviors. As experimental tools advance, the repertoire of known endocannabinoid-mediated effects at the synapse, and their underlying mechanism, continues to expand. Retrograde signaling is the principal mode by which endocannabinoids mediate short- and long-term forms of plasticity at both excitatory and inhibitory synapses. However, growing evidence suggests that endocannabinoids can also signal in a non-retrograde manner. In addition to mediating synaptic plasticity, the endocannabinoid system is itself subject to plastic changes. Multiple points of interaction with other neuromodulatory and signaling systems have now been identified. Synaptic endocannabinoid signaling is thus mechanistically more complex and diverse than originally thought. In this review, we focus on new advances in endocannabinoid signaling and highlight their role as potent regulators of synaptic function in the mammalian brain.
Introduction
Since the discovery of Δ9-tetrahydrocannabinol (THC) as the main psychoactive ingredient in marijuana, and the cloning of cannabinoid receptors and the identification of their endogenous ligands (endocannabinoids, eCBs), our understanding of the molecular basis and functions of the eCB signaling system has evolved considerably. Extensive research in the last 15 years has consolidated our view on eCBs as powerful regulators of synaptic function throughout the central nervous system (CNS). Their role as retrograde messengers suppressing transmitter release in a transient or long-lasting manner, at both excitatory and inhibitory synapses, is now well-established (Alger, 2012; Chevaleyre et al., 2006; Freund et al., 2003; Kano et al., 2009; Katona and Freund, 2012). Apart from signaling in more mature systems, the eCB system has been implicated in synapse formation and neurogenesis (Harkany et al., 2008). It is also widely believed that by modulating synaptic strength, eCBs can regulate a wide range of neural functions, including cognition, motor control, feeding behaviors and pain. Moreover, dysregulation of the eCB system is implicated in neuropsychiatric conditions such as depression and anxiety (Hillard et al., 2012; Mechoulam and Parker, 2012). As such, the eCB system provides an excellent opportunity for therapeutic interventions (Ligresti et al., 2009; Piomelli, 2005). Their prevalence throughout the brain suggests eCBs are fundamental modulators of synaptic function. This review focuses on recent advances in eCB signaling at central synapses.
The eCB signaling system comprises: (1) at least two G-protein-coupled receptors (GPCRs), known as the cannabinoid type-1 and type-2 receptors (CB1R and CB2R); (2) the endogenous ligands (eCBs), of which anandamide (AEA) and 2-arachidonoylglycerol (2-AG) are the best characterized; and (3) synthetic and degradative enzymes and transporters that regulate eCB levels and action at receptors. An enormous amount of information on the general properties of the eCB system has accumulated over the last two decades (for general reviews on the eCB system, see Ahn et al., 2008; Di Marzo, 2009; Howlett et al., 2002; Pertwee et al., 2010; Piomelli, 2003). We discuss essential features of this system in the context of synaptic function.
The principal mechanism by which eCBs regulate synaptic function is through retrograde signaling (for a thorough review, see Kano et al., 2009). Here, postsynaptic activity leads to the production of an eCB that moves backward across the synapse, binds presynaptic CB1Rs, and suppresses neurotransmitter release (Fig. 1A). However, there is also evidence suggesting that eCBs signal in a non-retrograde or autocrine manner, where they can modulate neural function and synaptic transmission by engaging transient receptor potential TRPV1 (vanilloid receptor type 1) and also CB1Rs located on or within the postsynaptic cell (Fig. 1B). Finally, recent studies indicate that eCBs can signal via astrocytes to indirectly modulate presynaptic or postsynaptic function (Fig. 1C). This review aims to highlight the emerging mechanistic diversity of synaptic eCB signaling.
Figure 1. Endocannabinoid signaling at the synapse.

A, Retrograde endocannabinoid (eCB) signaling. eCBs are mobilized from postsynaptic neurons and target presynaptic cannabinoid type-1 receptors (CB1Rs) to suppress neurotransmitter release. B, Non-retrograde eCB signaling. eCBs produced in postsynaptic neurons activate postsynaptic CB1Rs or transient receptor potential vanilloid-type 1 (TRPV1) channels. C, Neuron-astrocyte eCB signaling. eCBs released from postsynaptic neurons stimulate astrocytic CB1Rs, thereby triggering gliotransmission. Glu, glutamate.
Retrograde Endocannabinoid Signaling
The first demonstration of retrograde eCB signaling came from the discovery that eCBs mediate forms of short-term synaptic plasticity known as depolarization-induced suppression of inhibition (DSI) (Ohno-Shosaku et al., 2001; Wilson and Nicoll, 2001) and depolarization-induced suppression of excitation (DSE) (Kreitzer and Regehr, 2001). Shortly after it was shown that eCBs also mediate presynaptic forms of long-term depression (eCB-LTD) at both excitatory (Gerdeman et al., 2002; Robbe et al., 2002) and inhibitory synapses (Chevaleyre and Castillo, 2003; Marsicano et al., 2002). eCBs have since emerged as the best characterized retrograde messengers (Regehr et al., 2009), with numerous examples of short- and long-term forms of synaptic plasticity reported throughout the brain (Heifets and Castillo, 2009; Kano et al., 2009).
CB1/CB2 receptors are Gi/o protein-coupled receptors that mediate almost all effects of exogenous and endogenous cannabinoids. CB1Rs are one of the most widely expressed GPCRs in the brain (Herkenham et al., 1990). Their localization to neuronal terminals (Katona et al., 1999; Katona et al., 2006) strongly suggests CB1Rs play important roles in regulating synaptic function. Indeed, CB1R activation inhibits neurotransmitter release at synapses through two main mechanisms (Fig. 2) (Chevaleyre et al., 2006; Freund et al., 2003; Kano et al., 2009). For short-term plasticity, in which CB1Rs are activated for a few seconds, the mechanism involves direct G protein-dependent (likely via the βγ subunits) inhibition of presynaptic Ca2+ influx through voltage-gated Ca2+ channels (VGCCs) (Brown et al., 2003; Kreitzer and Regehr, 2001; Wilson et al., 2001). For long-term plasticity, the predominant mechanism requires inhibition of adenylyl cyclase and downregulation of the cAMP/PKA pathway via the αi/o limb (Chevaleyre et al., 2006; Heifets and Castillo, 2009). Moreover, CB1Rs only need to be engaged during the induction, but not expression phase of eCB-LTD. Induction also requires combined presynaptic firing with CB1R activation, thereby providing a mechanism for input-specificity; that is, only active synapses detecting eCBs express long-term plasticity (Heifets et al., 2008; Singla et al., 2007). The expression mechanism for eCB-LTD may involve presynaptic proteins Rab3B/RIM1α, (Chevaleyre et al., 2007; Tsetsenis et al., 2011) or a reduction of P/Q-type VGCCs (Mato et al., 2008). While other effectors downstream of CB1Rs have been described, mainly in cultured cells and expression systems (Howlett, 2005; Pertwee et al., 2010), their role in regulating synaptic function is presently less clear. In contrast to CB1Rs, which are widely expressed in the brain, CB2Rs are typically found in the immune system and are poorly expressed in the CNS. Although recent studies support a role for these receptors in the CNS (den Boon et al., 2012; Van Sickle et al., 2005; Xi et al., 2011), when compared with CB1Rs, much less is known about the precise cellular mechanism(s) and contributions of CB2Rs to brain function.
Figure 2. Molecular mechanisms underlying endocannabinoid-mediated short- and long-term synaptic plasticity.

A, Short-term depression. Postsynaptic activity triggers Ca2+ influx via voltage-gated Ca2+ channels (VGCCs). Other Ca2+ sources, like NMDARs and internal stores, may contribute. Ca2+ promotes diacylglycerol lipase (DGLα)-mediated eCB production by an unknown mechanism. Presynaptic activity can also lead to eCB mobilization by activating postsynaptic group-I metabotropic glutamate receptors (I-mGluRs). Phospholipase-Cβ (PLCβ) can now act as a coincidence detector integrating pre- and postsynaptic activity. DGLα promotes 2-arachidonoylglycerol (2-AG) release which retrogradely targets presynaptic CB1Rs, and the βγ subunits likely couple to presynaptic VGCCs to reduce neurotransmitter release. B, eCB-mediated excitatory long-term depression (LTD) and inhibitory LTD (iLTD). Patterned presynaptic stimulation releases Glu which activates postsynaptic mGluRs coupled to PLCβ and DGLα. 2-AG homosynaptically targets CB1Rs localized to excitatory terminals and heterosynaptically engages CB1Rs at inhibitory terminals. A Gαi/o-dependent reduction in adenylyl cyclase (AC) and protein kinase A (PKA) activity suppresses transmitter release. At inhibitory synapses, decreased PKA activity, in conjunction with activation of the Ca2+-sensitive phosphatase calcineurin (CaN), shifts the phosphorylation status of an unidentified presynaptic target (T) required for iLTD. The active zone protein RIM1α and the vesicle-associated protein Rab3B are also necessary for iLTD. Induction of eCB-LTD may require presynaptic Ca2+ rise through VGCCs, NMDARs, or internal stores (not shown). Dashed lines indicate putative pathways.
Although several eCBs have been identified, just two, AEA and 2-AG, emerged as the most relevant and prevalent regulators of synaptic function. While 2-AG seems to be the principal eCB required for activity-dependent retrograde signaling, the relative contribution of 2-AG and AEA to synaptic transmission is still debated. Functional cross-talk between 2-AG and AEA signaling was reported (Maccarrone et al., 2008), and recent findings suggest that 2-AG and AEA can be recruited differentially from the same postsynaptic neuron depending on the type of presynaptic activity (Lerner and Kreitzer, 2012; Puente et al., 2011). A more complete signaling profile for 2-AG and AEA –including production, target identification, and degradation– is indispensable for better understanding their short- and long-term impact on synaptic function.
Synthesis and degradation of eCBs help shape their spatiotemporal signaling profile. For retrograde eCB signaling, postsynaptic neuronal depolarization elevates intracellular Ca2+ via VGCCs and elicits 2-AG production presumably by activating Ca2+-sensitive enzymes. In addition, glutamate release onto postsynaptic group I metabotropic glutamate receptors (I-mGluRs) (Maejima et al., 2001; Varma et al., 2001) can generate 2-AG by activating the enzyme phospholipase Cβ (PLCβ) (for a review, see Hashimotodani et al., 2007a). Most likely Ca2+ influx through VGCCs and downstream signaling from I-mGluR activation converge on the same metabolic pathway to mobilize 2-AG (Fig. 2A). PLCβ is thought to act as a coincidence detector for postsynaptic Ca2+ and GPCR signaling (Hashimotodani et al., 2005; Maejima et al., 2005). This interaction might be important for integrating synaptic activity (Brenowitz and Regehr, 2005). On the other hand, it is worth noting that activation of I-mGluRs is sufficient to mobilize eCBs to trigger short- and long-term forms of plasticity (Chevaleyre et al., 2006). For long-term plasticity, a few minutes of CB1R stimulation is needed, which can result from a brief postsynaptic I-mGluR activation triggering a relatively longer-lasting 2-AG mobilization (Chevaleyre and Castillo, 2003). Of general physiological relevance, many other Gq/11-GPCRs are known to promote eCB synthesis (Katona and Freund, 2012). Upon activation, PLCβ hydrolyzes phosphatidylinositol to generate diacylglycerol, which is converted to 2-AG by diacylglycerol lipase α (DGLα). DGLα is specifically localized to postsynaptic compartments (Katona et al., 2006; Lafourcade et al., 2007; Nomura et al., 2007; Yoshida et al., 2006). Whereas pharmacological studies inconsistently implicated DGLα in short-term synaptic plasticity, genetic deletion of DGLα indicates this enzyme is required for Ca2+-dependent 2-AG production and short- and long-term eCB-dependent synaptic plasticity (Gao et al., 2010; Tanimura et al., 2010; Yoshino et al., 2011). Once synthesized, 2-AG travels backward across the synapse; however, the precise mechanism by which this occurs is still unresolved.
The primary degradative enzyme for 2-AG is monoacylglycerol lipase (MGL) (Blankman et al., 2007). MGL is found presynaptically (Gulyas et al., 2004; Ludanyi et al., 2011) but its expression seems to be heterogeneous across synapses (Tanimura et al., 2012; Uchigashima et al., 2011; Yoshida et al., 2011). The postsynaptically localized serine hydrolase ABHD6 also catabolizes a small fraction of 2-AG (Marrs et al., 2010), suggesting functional redundancy that could help fine-tune 2-AG signaling. Nevertheless, it seems clear that MGL controls the duration and magnitude of 2-AG-mediated synaptic plasticity (Hashimotodani et al., 2007b; Pan et al., 2011; Schlosburg et al., 2010; Szabo et al., 2006). While 2-AG likely signals within 20 μm of its site of origin (Chevaleyre and Castillo, 2004; Wilson and Nicoll, 2001), it would be useful to examine the relative contribution of MGL and ABHD6 to 2-AG diffusion.
In contrast to synaptic 2-AG signaling, AEA synthesis and degradation seems more complex. Postsynaptic depolarization and intracellular Ca2+ influx support AEA production, but how this occurs is not fully understood (Di Marzo, 2011). AEA is in part synthesized by N-acyl-phosphatidylethanolamine-hydrolyzing phospholipase-D (NAPE-PLD). However, alternative synthetic pathways exist (Okamoto et al., 2007). NAPE-PLD can be expressed postsynaptically (Cristino et al., 2008), but was also observed on axonal membranes, in particular at CA3 mossy fiber terminals (Egertova et al., 2008; Nyilas et al., 2008), where AEA could locally modulate presynaptic function. AEA transport across membranes might be facilitated by a lipophilic carrier protein (Beltramo et al., 1997; Fu et al., 2012; Hillard et al., 1997). This protein presumably supports AEA delivery to intracellular compartments where fatty-acid amide hydrolase (FAAH), the enzyme primarily responsible for AEA degradation, is localized (Gulyas et al., 2004). While 2-AG and AEA are hydrolyzed by MGL and FAAH, respectively, oxidizing enzymes like cyclooxygenase and lipoxygenase can also utilize these substrates (Vandevoorde and Lambert, 2007). Of interest, some of these eCB metabolites are biologically active (Nomura et al., 2011) and probably modulate synaptic function, a possibility that needs to be further investigated. Continued exploration of the mechanisms underlying eCB synthesis and degradation will advance our understanding of how lipids shape synaptic function.
Non-retrograde Endocannabinoid Signaling
Besides the classical cannabinoid receptors (CB1R/CB2R), there is growing evidence that TRPV1 channels also participate in eCB signaling (De Petrocellis and Di Marzo, 2010; Pertwee et al., 2010). TRPV1, originally VR1 for vanilloid receptor type-1 (Caterina et al., 1997), is a polymodal transient receptor potential (TRP) ion channel largely expressed in afferent peripheral sensory neurons where its activation regulates synaptic transmission associated with pain sensation (Caterina and Julius, 2001). Interestingly, TRPV1 can bind lipophilic substances, such as AEA (Di Marzo et al., 2002). Of note, AEA is a partial agonist at the CB1R but a full agonist at TRPV1 channels (Smart et al., 2000; Zygmunt et al., 1999). In addition to their expression in the periphery, TRPV1 channels have been found in the CNS (Cristino et al., 2006; Cristino et al., 2008; Mezey et al., 2000; Puente et al., 2011; Roberts et al., 2004; Toth et al., 2005) (but see Cavanaugh et al., 2011) where they appear to regulate synaptic function.
Recent studies revealed that AEA acting on TRPV1 mediates a postsynaptic form of LTD (Fig. 3A). This TRPV1-LTD has been observed in dopamine receptor-2 (D2)-positive medium spiny neurons of the nucleus accumbens (Grueter et al., 2010), in dentate granule cells (Chavez et al., 2010), and in the bed nucleus of the stria terminalis (Puente et al., 2011). In each case, activation of mGluR5, presumably via PLC (Liu et al., 2008) and Ca2+ release from intracellular stores, promotes the synthesis of AEA that activates TRPV1 channels. In addition, TRPV1-LTD relies on AMPAR-endocytosis. These findings are consistent with the notion that AEA can act as an intracellular messenger (van der Stelt and Di Marzo, 2005), but differs from a presynaptic, TRPV1-dependent LTD at glutamatergic synapses onto CA1 hippocampal interneurons (Gibson et al., 2008). While CB1Rs mediate excitatory and inhibitory synaptic plasticity, whether brain TRPV1 channels mediate inhibitory synaptic plasticity is unknown. There is also evidence that TRPV1 localizes to neuronal intracellular compartments like the endoplasmic reticulum, trans-Golgi network, and perhaps even vesicles (Dong et al., 2010). The functional significance of such receptors warrants further investigation.
Figure 3. Non-retrograde eCB signaling.
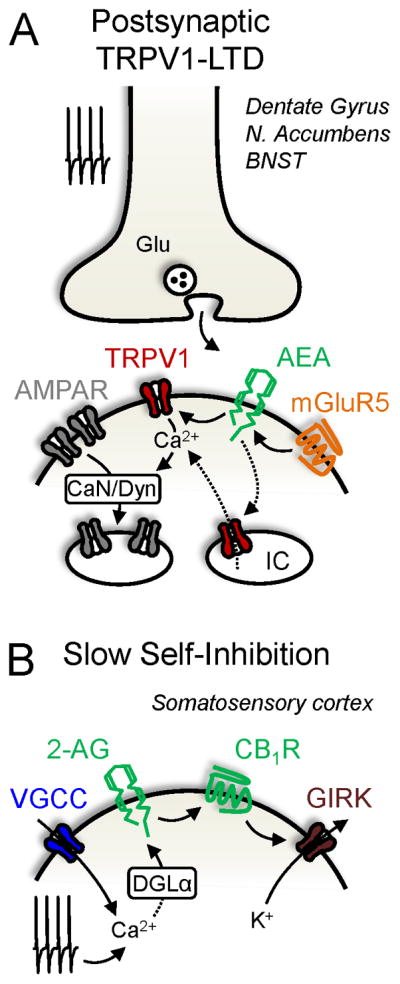
A, Mechanism underlying postsynaptic TRPV1-LTD. Presynaptic activity releases glutamate that stimulates mGluR5. Postsynaptic depolarization may also be required. mGluR5 couples to anandamide (AEA) production which activates TRPV1, leading to enhanced Ca2+ signaling. Ca2+ engages calcineurin/dynamin (CaN/Dyn), causing AMPA receptor (AMPAR) endocytosis and LTD. IC, intracellular compartment. N. Accumbens, nucleus accumbens; BNST, bed nucleus of the stria terminalis. B, Mechanism responsible for slow-self inhibition (autocrine signaling). Postsynaptic activity-induced Ca2+ elevation facilitates 2-AG production. 2-AG activates postsynaptic CB1Rs that signal to a G protein-coupled inwardly rectifying K+ (GIRK) channel to hyperpolarize the membrane potential and inhibit neuronal firing. Dashed lines indicate putative pathways.
Non-retrograde eCB signaling has been observed in other contexts. Repetitive activation of a subtype of neocortical GABAergic interneuron triggers a CB1R-dependent postsynaptic hyperpolarization, which reduced its excitability (Fig. 3B) (Bacci et al., 2004). This slow self-inhibition resulted from activity-dependent rises in intracellular Ca2+, mobilization of 2-AG, and activation of CB1Rs that couple to a G protein-coupled inwardly rectifying K+ channel (Bacci et al., 2004; Marinelli et al., 2008). This form of autocrine signaling was also observed in a fraction of layer 2/3 neocortical pyramidal neurons (Marinelli et al., 2009). Unexpectedly, CB2Rs were recently shown to mediate an activity-induced self-inhibition in medial prefrontal cortical pyramidal neurons (den Boon et al., 2012). CB2Rs were localized to intracellular compartments and coupled to calcium-activated chloride channels to decrease neuronal firing. The generalizability of autocrine eCB signaling to other brain regions should be examined.
Endocannabinoid-mediated Communication between Neurons and Glia
Growing evidence indicates that glia participate in eCB signaling (Stella, 2010). The synthetic machinery for eCB production was observed in oligodendrocytes (Gomez et al., 2010), astrocytes, and microglial cells (Hegyi et al., 2012). Likewise, cultured astrocytes and microglial cells can produce 2-AG or AEA (Stella, 2009). It is not yet clear if eCBs produced by glial cells modulate synaptic transmission. On the other hand, several recent findings support a role for eCBs signaling to astrocytes and their ability to indirectly mediate synaptic function.
At Schaffer collateral excitatory synapses onto hippocampal CA1 pyramidal neurons, postsynaptic activity-dependent release of eCBs was shown to target not only presynaptic CB1Rs but also astrocytic CB1Rs (Fig. 4A). Astrocytic CB1Rs unexpectedly coupled to PLC via Gq/11, which increased intracellular Ca2+ and triggered glutamate release (Navarrete and Araque, 2008). In support of these functional observations, CB1Rs in hippocampal astrocytes have recently been observed using immunoelectron microscopy (Han et al., 2012). Glutamate activated NMDARs on CA1 pyramidal neurons and, at some synapses, triggered short-term facilitation of transmitter release presumably by stimulating presynaptic mGluR1s (Navarrete and Araque, 2008, 2010). Interestingly, this short-term facilitation was not spatially restricted, being observed over 70 μm away from the active pyramidal cell. Thus, eCBs could concomitantly suppress synaptic transmitter release by triggering DSE and indirectly potentiate synaptic transmission through astrocytes, both in a CB1R-dependent manner. While the functional significance of such plasticity is not yet clear, astrocytes may have long-distance neuromodulatory effects that are mediated by eCB signaling.
Figure 4. Astrocytic CB1Rs modulation of synaptic transmission.
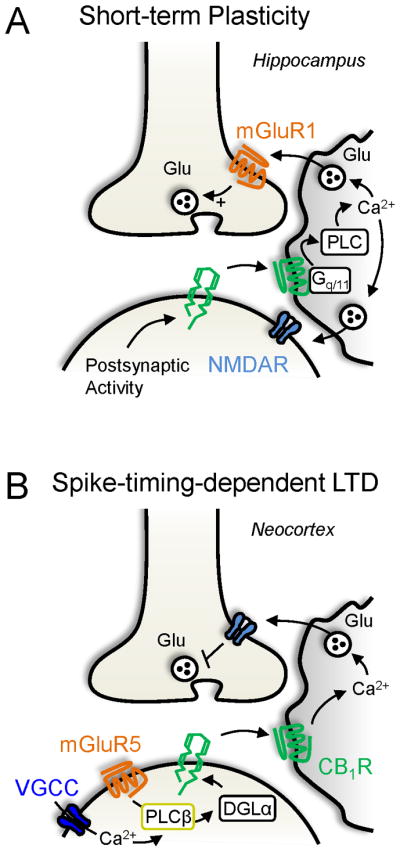
A, Short-term plasticity. Postsynaptic neuronal activity leads to eCB release. eCBs target Gq/11-coupled CB1Rs localized to astrocytes. As a result, PLC activity facilitates astrocytic Ca2+ signaling. Glu released from the astrocyte activates presynaptic mGluR1s to potentiate release and postsynaptic NMDARs to trigger a slow inward current. B, Spike-timing-dependent LTD. Repetitive pairings of post-before-pre synaptic activity mobilizes eCBs through the neuronal PLCβ-coincidence detection mechanism. Released eCB stimulates astrocytic CB1Rs, leading to Ca2+ signaling. Astrocyte-mediated Glu release activates presynaptic NMDARs to depress release.
eCB-mediated neuron-astrocyte communication has also been implicated in long-term plasticity. Spike timing-dependent LTD (tLTD) between neocortical pyramidal neurons is known to require activation of presynaptic NMDARs and CB1Rs (Bender et al., 2006; Nevian and Sakmann, 2006; Sjostrom et al., 2003). Surprisingly, a recent study found that astrocytic CB1Rs were necessary and sufficient to mediate tLTD (Min and Nevian, 2012). eCBs originating from layer 2/3 pyramidal neurons activated astrocytic CB1Rs, which increased intracellular Ca2+, thereby releasing glutamate and stimulating presynaptic NMDARs (Fig. 4B). Given the anatomical and functional evidence for presynaptic CB1Rs in neocortex (Domenici et al., 2006; Hill et al., 2007; Lafourcade et al., 2007), future studies could use astrocyte- and neuron-specific CB1R knockout mice to identify the exact conditions required to activate neuronal and/or astrocytic CB1Rs.
Attesting to the possible physiological relevance of astrocytic CB1Rs, a recent in vivo study showed that intraperitoneal injection of THC induced long-lasting suppression of excitatory synaptic transmission in hippocampal area CA1, an effect that required astrocytic CB1Rs (Han et al., 2012). Previous work in acute hippocampal slices from global CB1R knockout mice suggested that agonist-mediated suppression of excitatory transmission in CA1 depends solely on CB1Rs expressed at Schaffer collateral terminals (Katona et al., 2006; Kawamura et al., 2006; Takahashi and Castillo, 2006). Unexpectedly, however, THC-mediated suppression of synaptic transmission in vivo was intact in glutamatergic and GABAergic specific CB1R knockout mice, whereas it was abolished in glia-specific CB1R knockout mice (Han et al., 2012). Mechanistically, glutamate, presumably released from astrocytes, activated postsynaptic NMDARs, triggering AMPAR endocytosis and subsequent synaptic depression. These results contrast with those observed in vitro in which eCBs indirectly facilitated synaptic transmission via astrocytic CB1Rs (Navarrete and Araque, 2008, 2010). A thorough examination of the conditions necessary for activating synaptic and astrocytic CB1Rs is clearly needed.
Tonic Endocannabinoid Signaling
In addition to the classical, activity-dependent phasic mode of eCB mobilization, tonic eCB signaling has been reported. Tonic signaling can be observed as an increase in basal synaptic transmission following pharmacological blockade of CB1Rs (Auclair et al., 2000; Hentges et al., 2005; Losonczy et al., 2004; Neu et al., 2007; Oliet et al., 2007; Slanina and Schweitzer, 2005; Zhu and Lovinger, 2010). However, CB1R blockade in this manner does not always reveal an eCB tone (Chevaleyre and Castillo, 2003; Pan et al., 2011; van Beugen et al., 2006; Wilson and Nicoll, 2001; Zhong et al., 2011). Build-up of an eCB tone can occur when inhibiting eCB uptake (Wilson and Nicoll, 2001) or genetic deletion of MGL (Pan et al., 2011; Zhong et al., 2011). The fact that most 2-AG is hydrolyzed by MGL (Blankman et al., 2007; Chanda et al., 2010; Nomura et al., 2011) suggests 2-AG mediates tonic eCB signaling, which is consistent with a constitutive release of 2-AG in cultured neurons (Hashimotodani et al., 2007b). On the other hand, AEA can also contribute to tonic eCB signaling. Chronic inactivity in hippocampal slice cultures reduced an AEA tone presumably by augmenting AEA uptake and degradation (Kim and Alger, 2010). Together, these studies suggest tonic eCB signaling can control, under some conditions, basal synaptic neurotransmitter release. It is currently unclear if regional differences in the expression pattern of enzymes responsible for eCB metabolism can fully account for synapse specificity. Moreover, most of these studies were performed in vitro, and therefore, the impact of an eCB tone on synaptic function in vivo should be further examined.
Interaction between Endocannabinoids and other Signaling Systems
The eCB system allows for multiple points of interaction with other signaling and neuromodulatory systems. In addition to regulating release of classical neurotransmitters like glutamate and GABA, CB1Rs can also control the release of several neuromodulators including serotonin, acetylcholine, dopamine, opioids, norepinephrine, and cholecystokinin (Alger, 2002; Kano et al., 2009; Schlicker and Kathmann, 2001). On the other hand, many of these neuromodulators actually couple to eCB synthesis by activating their respective Gq/11 protein-coupled receptors (for a comprehensive list, see Katona and Freund, 2012). Additionally, regulators of G protein signaling were recently shown to control Gq/11 coupled receptors and eCB mobilization (Lerner and Kreitzer, 2012), indicating how GPCRs themselves can fine tune eCB release. Together, these studies not only support a general theme by which Gq/11-coupled GPCRs mobilize eCBs, but demonstrate the existence of multiple routes for eliciting and regulating eCB release.
On the other side of the synapse, functional interactions between CB1Rs and other receptors have been identified. For example, at inhibitory terminals in the prefrontal cortex, D2-like receptors colocalize with CB1Rs where they appear to facilitate CB1R-mediated suppression of transmitter release (Chiu et al., 2010). This is probably due to a cooperative lowering of PKA activity, consistent with similar observations in the ventral tegmental area (Pan et al., 2008). In addition, work in visual cortical slices from young mice suggests that BDNF interferes with CB1R downstream signaling, thereby disrupting eCB-mediated suppression of neurotransmitter release (Huang et al., 2008). This might result from, at least in part, BDNF inhibiting CB1R function through a mechanism requiring cholesterol metabolism and altered membrane lipid raft function (De Chiara et al., 2010). At Schaffer collaterals, adenosine A1 receptors (A1Rs) colocalize with CB1Rs. Tonic activation of A1Rs can reduce the efficacy of CB1R-mediated inhibition of glutamate release (Hoffman et al., 2010). Also in the hippocampus, stimulating GluK1-containing kainate receptors at inhibitory terminals appears to actually facilitate CB1R signaling (Lourenco et al., 2010). The mechanism by which this occurs is not yet clear. Adding to the complexity of eCB signaling, evidence suggests CB1Rs can associate with other GPCRs to form heteromeric complexes. Such interactions have been detected for CB1-D2, CB1-opioid, CB1-A2A, and CB1-orexin-1 receptor pairs (Hudson et al., 2010; Mackie, 2005; Pertwee et al., 2010). Strikingly, higher order heteromeric complexes consisting of CB1, D2, and A2ARs have also been observed (Carriba et al., 2008). Intriguingly, these macromolecular interactions can significantly change the downstream G-proteins recruited during receptor activation. Much more work is needed to determine the physiological impact of these heteromeric complexes in the brain, and in particular, at the synapse.
Plasticity of the Endocannabinoid System
In addition to triggering various forms of synaptic plasticity like DSI/DSE, eCB-LTD, and TRPV1-LTD, the eCB system itself undergoes plastic changes. Mechanistically, plasticity of the eCB system can arise by modifications to any of its components; for example, CB1R number/function or eCB production/degradation. These changes have been observed both in vivo and in vitro, and can be triggered by several natural and experimental conditions including neural activity and agonist-induced CB1R activation. Of clinical relevance, changes in eCB signaling are also associated with several brain disorders. Here, we illustrate how plasticity of the eCB system can profoundly affect synaptic physiology and ultimately, brain function.
An interesting example of agonist-induced plasticity of eCB signaling comes from the observation that a single in vivo exposure to THC abolished for a few days eCB-mediated retrograde signaling in the hippocampus and nucleus accumbens of mice (Mato et al., 2004). This effect was associated with a reduction in CB1R maximal efficacy without modifications in total binding or coupling. Prolonged exposure to agonists in humans and animal models results in behavioral tolerance, which is classically attributed to receptor desensitization and internalization (Coutts et al., 2001; Jin et al., 1999; Wu et al., 2008). However, a reduction in CB1R lateral mobility may also contribute (Mikasova et al., 2008). Understanding the impact of synaptic CB1R signaling and trafficking in vivo should further reveal how eCBs control physiological responses to drugs-of-abuse.
The eCB system also undergoes developmental changes (Harkany et al., 2008). In the hippocampus, both the magnitude of eCB-mediated iLTD and the ability of a CB1R agonist to suppress inhibitory transmission were greater in juvenile than in adult rats (Kang-Park et al., 2007; see also Zhu and Lovinger, 2010). In addition, a form of eCB-mediated heterosynaptic LTD at excitatory synapses was observed in young animals, attenuated across development, and disappeared in the mature brain (Yasuda et al., 2008). Lower expression levels of CB1Rs at excitatory synapses in the adult brain may underlie these changes (Kawamura et al., 2006). Along these lines, developmentally expressed CB1Rs at mossy fiber terminals in the CA3 region of the hippocampus mediate eCB-LTD at immature but not mature synapses (Caiati et al., 2012). Postsynaptic eCB production is also modulated over time. A developmental shift from LTP to eCB-LTD was reported in the striatum (Ade and Lovinger, 2007). Whereas CB1R sensitivity to its agonist was not changed, the shift in plasticity was associated with developmental increases in AEA levels, suggesting that AEA determines the direction of synaptic plasticity. Similarly, it was shown that the magnitude of hippocampal DSI is developmentally regulated, such that DSI is modest in early postnatal days and becomes more robust at two weeks postnatal (Zhu and Lovinger, 2010). The mechanism is not entirely clear but it might rely on a postsynaptic change in eCB release. In addition, tonic eCB release suppresses GABAergic transmission in the mature but not the neonatal hippocampus (Kang-Park et al., 2007; Zhu and Lovinger, 2010). While these studies argue that synaptic eCB signaling is developmentally regulated, the exact mechanisms underlying these changes remain unclear.
In mature animals, eCB signaling can be modified in an activity-dependent manner. High frequency (Chen et al., 2007) or low frequency stimulation (Zhu and Lovinger, 2007) of Schaffer collaterals, as well as brief pharmacological activation of I-mGluRs (Edwards et al., 2008), triggered a long-lasting potentiation in the magnitude of DSI. Remarkably, the transient postsynaptic Ca2+ rise that occurs during a single episode of DSI facilitated subsequent I-mGluR-dependent mobilization of eCBs and the induction of iLTD (Edwards et al., 2008). The molecular components that undergo priming are unknown. A similar DSI potentiation was observed following a single episode of experimentally induced febrile seizures (Chen et al., 2003). This potentiation was due to an increase in the number of CB1Rs associated with perisomatic inhibitory inputs. In contrast, the epileptic human hippocampus showed a reduction in the expression of CB1Rs at glutamatergic terminals (Ludanyi et al., 2008). Nevertheless, both upregulation of CB1Rs at GABAergic terminals and downregulation of CB1Rs at excitatory terminals are potentially epileptogenic, suggesting that dysregulation of the eCB system could play a role in epilepsy. Identifying the molecular basis for these activity-dependent changes in CB1R expression levels is important because it may uncover novel therapeutic targets.
Altered eCB signaling has been reported in experimental models for disorders like Fragile X syndrome. Up-regulation of eCB signaling was found in Fragile X mental retardation protein knockout mice as indicated by facilitation of I-mGluR agonist-induced iLTD. Facilitated iLTD might result from aberrant coupling between I-mGluR activation and eCB mobilization (Zhang and Alger, 2010). Aberrant coupling might be due to changes in Homer 1a protein, which reportedly interacts with mGluRs to regulate eCB release in cultured hippocampal neurons (Roloff et al., 2010). Another possible mediator of aberrant coupling includes the excitatory synapse-specific scaffolding protein SAPAP3, which can modulate postsynaptic mGluRs and eCB-mediated synaptic plasticity in the striatum (Chen et al., 2011). Continued exploration of the mechanisms underlying mGluR-coupled eCB production should provide clues as to how to treat patients with Fragile X syndrome.
Several studies indicate physiological responses to stress modulate the expression levels of key components of the eCB system (Riebe and Wotjak, 2011). In general, how stress modulates eCB signaling is largely dependent on brain regions, stress paradigm, and duration of stress exposure. In the striatum and nucleus accumbens, chronic stress inhibited CB1R mediated suppression of synaptic transmission (Rossi et al., 2008; Wang et al., 2010). Downregulation of CB1R function might underlie this eCB signaling deficiency since stress-induced downregulation of CB1R function was observed in the hypothalamus (Wamsteeker et al., 2010). There is also evidence that stress can enhance eCB signaling. Repeated restraint stress increased 2-AG levels and enhanced DSI in the basolateral amygdala (Patel et al., 2009). Similarly, restraint stress increased 2-AG levels and enhanced DSI in hippocampal CA1 pyramidal neurons (Wang et al., 2012).
Food intake is another physiological process that modulates the eCB system (Banni and Di Marzo, 2010; Dipatrizio and Piomelli, 2012). For example, CB1R agonists increase food intake whereas antagonists reduce food consumption. Providing mechanistic insight as to how this modulation may occur, a recent study showed that diet-induced obesity in mice increased hippocampal DGLα protein, 2-AG and AEA production, as well as CB1R expression (Massa et al., 2010). Levels of DGLβ, MGL, and FAAH were unchanged. Consistently, DSI and eCB-mediated iLTD were augmented in these mice (Massa et al., 2010). Diet restrictions likewise cause significant changes in the eCB system. In hypothalamic feeding circuits, food deprivation downregulated CB1R signaling, converting eCB-mediated LTD expressing synapses into ones that show nitric-oxide-dependent LTP (Crosby et al., 2011). In addition, polyunsaturated fatty acid diet-deficient mice showed impaired eCB-mediated LTD in both prefrontal cortex and nucleus accumbens (Lafourcade et al., 2011). Lack of eCB-LTD was attributed to reduced coupling of the CB1R to its downstream Gi/o protein. Intriguingly, these mice exhibited defects in mood and emotional behavior, implicating synaptic eCB signaling in affective behaviors. Taken together, these studies highlight how behavioral manipulations profoundly regulate eCB signaling and synaptic function.
Conclusions and Future Directions
In this review, we have highlighted essential properties of eCB signaling at the synapse. Research in the last decade has bolstered eCBs as powerful regulators of synaptic function throughout the CNS. Exciting developments have uncovered new mechanisms underlying eCB-mediated regulation of synaptic transmission. Moreover, the dynamics of synaptic eCB signaling displays an intricate, and sometimes reciprocal, set of interactions with other neuromodulatory systems. These emerging levels of complexity clearly indicate that much more work lies ahead in our pursuit to fully understand eCB signaling at the synapse.
While an overwhelming body of evidence strongly suggests eCBs are retrograde synaptic messengers, a major outstanding issue with the model is how a lipid traverses an aqueous synaptic cleft. Moreover, once in the extracellular space, how far do eCBs diffuse? While AEA seems to be transported by a lipid carrier protein, whether 2-AG is also transported by a lipid chaperone is unknown. Alternatively, specialized protein/lipid bridges, akin to synaptic intercellular adhesion molecules, could adopt a structural conformation that exposes lipophilic patches to reduce the retrograde energy barrier. Regardless of the exact mechanism, it is clear that eCB signaling powerfully regulates synaptic function. Developing new technologies to image lipid signaling, in real-time, should dramatically propel the field of eCB research forward.
Apart from their more traditional role in retrograde signaling, eCBs also appear to act in a non-retrograde manner to modulate postsynaptic function as well as trigger gliotransmission. However, the general physiological relevance of non-retrograde signaling mediated by TRPV1 in the CNS is not yet clear. While experimental evidence for eCBs targeting postsynaptic receptors is growing, whether presynaptically produced eCBs activate presynaptic CB1Rs or TRPV1 channels to modulate synaptic function remains unknown. In addition, the role of CB1Rs in regulating gliotransmission and indirectly, synaptic plasticity, warrants further investigation. Given the myriad of evidence supporting synaptic CB1Rs in modulating synaptic transmission, the precise conditions necessary for activating neuronal versus astrocytic CB1Rs must be defined.
Several other fundamental mechanistic questions remain unanswered. What are the rules governing CB1R trafficking into and out of membranes? What are the conditions required for CB1R heteromerization with other neuromodulatory receptors, and what is their impact on synaptic function? As for the two main eCBs, 2-AG and AEA, are there specific patterns of activity that predominantly mobilize one lipid versus the other? Perhaps these eCBs subserve specific functions at the synapse. If so, which ones? What is the precise role of tonic eCB release in the brain? In vitro approaches are unquestionably useful for addressing fundamental mechanisms underlying synaptic eCB signaling, but much more work in vivo is required to determine their contribution to physiological and pathological conditions. While a great deal of progress has been made in our understanding of eCB signaling and synaptic function, the greatest challenges lie ahead.
Acknowledgments
This work was supported by the NIH (R01-MH081935 and R01-DA17392 to P.E.C). A.E.C. was supported by a Ruth L. Kirschstein Award from the US National Institute of Neurological Disorders and Stroke (F32NS071821). Y.H. was supported by the Japan Society for the Promotion of Science Postdoctoral Fellowships for Research Abroad. We apologize to authors whose work we could not cite due to space limitations.
Footnotes
There are no conflicts of interest to report.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Ade KK, Lovinger DM. Anandamide regulates postnatal development of long-term synaptic plasticity in the rat dorsolateral striatum. J Neurosci. 2007;27:2403–2409. doi: 10.1523/JNEUROSCI.2916-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahn K, McKinney MK, Cravatt BF. Enzymatic pathways that regulate endocannabinoid signaling in the nervous system. Chemical reviews. 2008;108:1687–1707. doi: 10.1021/cr0782067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alger BE. Retrograde signaling in the regulation of synaptic transmission: focus on endocannabinoids. Prog Neurobiol. 2002;68:247–286. doi: 10.1016/s0301-0082(02)00080-1. [DOI] [PubMed] [Google Scholar]
- Alger BE. Endocannabinoids at the synapse a decade after the Dies Mirabilis (29 March 2001): what we still do not know. J Physiol. 2012 doi: 10.1113/jphysiol.2011.220855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auclair N, Otani S, Soubrie P, Crepel F. Cannabinoids modulate synaptic strength and plasticity at glutamatergic synapses of rat prefrontal cortex pyramidal neurons. J Neurophysiol. 2000;83:3287–3293. doi: 10.1152/jn.2000.83.6.3287. [DOI] [PubMed] [Google Scholar]
- Bacci A, Huguenard JR, Prince DA. Long-lasting self-inhibition of neocortical interneurons mediated by endocannabinoids. Nature. 2004;431:312–316. doi: 10.1038/nature02913. [DOI] [PubMed] [Google Scholar]
- Banni S, Di Marzo V. Effect of dietary fat on endocannabinoids and related mediators: consequences on energy homeostasis, inflammation and mood. Molecular nutrition & food research. 2010;54:82–92. doi: 10.1002/mnfr.200900516. [DOI] [PubMed] [Google Scholar]
- Beltramo M, Stella N, Calignano A, Lin SY, Makriyannis A, Piomelli D. Functional role of high-affinity anandamide transport, as revealed by selective inhibition. Science. 1997;277:1094–1097. doi: 10.1126/science.277.5329.1094. [DOI] [PubMed] [Google Scholar]
- Bender VA, Bender KJ, Brasier DJ, Feldman DE. Two coincidence detectors for spike timing-dependent plasticity in somatosensory cortex. J Neurosci. 2006;26:4166–4177. doi: 10.1523/JNEUROSCI.0176-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blankman JL, Simon GM, Cravatt BF. A Comprehensive Profile of Brain Enzymes that Hydrolyze the Endocannabinoid 2-Arachidonoylglycerol. Chem Biol. 2007;14:1347–1356. doi: 10.1016/j.chembiol.2007.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brenowitz SD, Regehr WG. Associative short-term synaptic plasticity mediated by endocannabinoids. Neuron. 2005;45:419–431. doi: 10.1016/j.neuron.2004.12.045. [DOI] [PubMed] [Google Scholar]
- Brown SP, Brenowitz SD, Regehr WG. Brief presynaptic bursts evoke synapse-specific retrograde inhibition mediated by endogenous cannabinoids. Nat Neurosci. 2003;6:1048–1057. doi: 10.1038/nn1126. [DOI] [PubMed] [Google Scholar]
- Caiati MD, Sivakumaran S, Lanore F, Mulle C, Richard E, Verrier D, Marsicano G, Miles R, Cherubini E. Developmental regulation of CB1-mediated spike-time dependent depression at immature mossy fiber-CA3 synapses. Sci Rep. 2012;2:285. doi: 10.1038/srep00285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carriba P, Navarro G, Ciruela F, Ferre S, Casado V, Agnati L, Cortes A, Mallol J, Fuxe K, Canela EI, et al. Detection of heteromerization of more than two proteins by sequential BRET-FRET. Nat Methods. 2008;5:727–733. doi: 10.1038/nmeth.1229. [DOI] [PubMed] [Google Scholar]
- Caterina MJ, Julius D. The vanilloid receptor: a molecular gateway to the pain pathway. Annu Rev Neurosci. 2001;24:487–517. doi: 10.1146/annurev.neuro.24.1.487. [DOI] [PubMed] [Google Scholar]
- Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389:816–824. doi: 10.1038/39807. [DOI] [PubMed] [Google Scholar]
- Cavanaugh DJ, Chesler AT, Jackson AC, Sigal YM, Yamanaka H, Grant R, O’Donnell D, Nicoll RA, Shah NM, Julius D, Basbaum AI. Trpv1 reporter mice reveal highly restricted brain distribution and functional expression in arteriolar smooth muscle cells. J Neurosci. 2011;31:5067–5077. doi: 10.1523/JNEUROSCI.6451-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chanda PK, Gao Y, Mark L, Btesh J, Strassle BW, Lu P, Piesla MJ, Zhang MY, Bingham B, Uveges A, et al. Monoacylglycerol lipase activity is a critical modulator of the tone and integrity of the endocannabinoid system. Mol Pharmacol. 2010;78:996–1003. doi: 10.1124/mol.110.068304. [DOI] [PubMed] [Google Scholar]
- Chavez AE, Chiu CQ, Castillo PE. TRPV1 activation by endogenous anandamide triggers postsynaptic long-term depression in dentate gyrus. Nat Neurosci. 2010;13:1511–1518. doi: 10.1038/nn.2684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen K, Neu A, Howard AL, Foldy C, Echegoyen J, Hilgenberg L, Smith M, Mackie K, Soltesz I. Prevention of plasticity of endocannabinoid signaling inhibits persistent limbic hyperexcitability caused by developmental seizures. J Neurosci. 2007;27:46–58. doi: 10.1523/JNEUROSCI.3966-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen K, Ratzliff A, Hilgenberg L, Gulyas A, Freund TF, Smith M, Dinh TP, Piomelli D, Mackie K, Soltesz I. Long-term plasticity of endocannabinoid signaling induced by developmental febrile seizures. Neuron. 2003;39:599–611. doi: 10.1016/s0896-6273(03)00499-9. [DOI] [PubMed] [Google Scholar]
- Chen M, Wan Y, Ade K, Ting J, Feng G, Calakos N. Sapap3 deletion anomalously activates short-term endocannabinoid-mediated synaptic plasticity. J Neurosci. 2011;31:9563–9573. doi: 10.1523/JNEUROSCI.1701-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chevaleyre V, Castillo PE. Heterosynaptic LTD of hippocampal GABAergic synapses: a novel role of endocannabinoids in regulating excitability. Neuron. 2003;38:461–472. doi: 10.1016/s0896-6273(03)00235-6. [DOI] [PubMed] [Google Scholar]
- Chevaleyre V, Castillo PE. Endocannabinoid-mediated metaplasticity in the hippocampus. Neuron. 2004;43:871–881. doi: 10.1016/j.neuron.2004.08.036. [DOI] [PubMed] [Google Scholar]
- Chevaleyre V, Heifets BD, Kaeser PS, Sudhof TC, Castillo PE. Endocannabinoid-mediated long-term plasticity requires cAMP/PKA signaling and RIM1alpha. Neuron. 2007;54:801–812. doi: 10.1016/j.neuron.2007.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chevaleyre V, Takahashi KA, Castillo PE. Endocannabinoid-mediated synaptic plasticity in the CNS. Annu Rev Neurosci. 2006;29:37–76. doi: 10.1146/annurev.neuro.29.051605.112834. [DOI] [PubMed] [Google Scholar]
- Chiu CQ, Puente N, Grandes P, Castillo PE. Dopaminergic modulation of endocannabinoid-mediated plasticity at GABAergic synapses in the prefrontal cortex. J Neurosci. 2010;30:7236–7248. doi: 10.1523/JNEUROSCI.0736-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coutts AA, Anavi-Goffer S, Ross RA, MacEwan DJ, Mackie K, Pertwee RG, Irving AJ. Agonist-induced internalization and trafficking of cannabinoid CB1 receptors in hippocampal neurons. J Neurosci. 2001;21:2425–2433. doi: 10.1523/JNEUROSCI.21-07-02425.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cristino L, de Petrocellis L, Pryce G, Baker D, Guglielmotti V, Di Marzo V. Immunohistochemical localization of cannabinoid type 1 and vanilloid transient receptor potential vanilloid type 1 receptors in the mouse brain. Neuroscience. 2006;139:1405–1415. doi: 10.1016/j.neuroscience.2006.02.074. [DOI] [PubMed] [Google Scholar]
- Cristino L, Starowicz K, De Petrocellis L, Morishita J, Ueda N, Guglielmotti V, Di Marzo V. Immunohistochemical localization of anabolic and catabolic enzymes for anandamide and other putative endovanilloids in the hippocampus and cerebellar cortex of the mouse brain. Neuroscience. 2008;151:955–968. doi: 10.1016/j.neuroscience.2007.11.047. [DOI] [PubMed] [Google Scholar]
- Crosby KM, Inoue W, Pittman QJ, Bains JS. Endocannabinoids gate state-dependent plasticity of synaptic inhibition in feeding circuits. Neuron. 2011;71:529–541. doi: 10.1016/j.neuron.2011.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Chiara V, Angelucci F, Rossi S, Musella A, Cavasinni F, Cantarella C, Mataluni G, Sacchetti L, Napolitano F, Castelli M, et al. Brain-derived neurotrophic factor controls cannabinoid CB1 receptor function in the striatum. J Neurosci. 2010;30:8127–8137. doi: 10.1523/JNEUROSCI.1683-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Petrocellis L, Di Marzo V. Non-CB1, non-CB2 receptors for endocannabinoids, plant cannabinoids, and synthetic cannabimimetics: focus on G-protein-coupled receptors and transient receptor potential channels. J Neuroimmune Pharmacol. 2010;5:103–121. doi: 10.1007/s11481-009-9177-z. [DOI] [PubMed] [Google Scholar]
- den Boon FS, Chameau P, Schaafsma-Zhao Q, van Aken W, Bari M, Oddi S, Kruse CG, Maccarrone M, Wadman WJ, Werkman TR. Excitability of prefrontal cortical pyramidal neurons is modulated by activation of intracellular type-2 cannabinoid receptors. Proc Natl Acad Sci U S A. 2012;109:3534–3539. doi: 10.1073/pnas.1118167109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Marzo V. The endocannabinoid system: its general strategy of action, tools for its pharmacological manipulation and potential therapeutic exploitation. Pharmacol Res. 2009;60:77–84. doi: 10.1016/j.phrs.2009.02.010. [DOI] [PubMed] [Google Scholar]
- Di Marzo V. Endocannabinoid signaling in the brain: biosynthetic mechanisms in the limelight. Nat Neurosci. 2011;14:9–15. doi: 10.1038/nn.2720. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, De Petrocellis L, Fezza F, Ligresti A, Bisogno T. Anandamide receptors. Prostaglandins Leukot Essent Fatty Acids. 2002;66:377–391. doi: 10.1054/plef.2001.0349. [DOI] [PubMed] [Google Scholar]
- Dipatrizio NV, Piomelli D. The thrifty lipids: endocannabinoids and the neural control of energy conservation. Trends Neurosci. 2012;35:403–411. doi: 10.1016/j.tins.2012.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domenici MR, Azad SC, Marsicano G, Schierloh A, Wotjak CT, Dodt HU, Zieglgansberger W, Lutz B, Rammes G. Cannabinoid receptor type 1 located on presynaptic terminals of principal neurons in the forebrain controls glutamatergic synaptic transmission. J Neurosci. 2006;26:5794–5799. doi: 10.1523/JNEUROSCI.0372-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong XP, Wang X, Xu H. TRP channels of intracellular membranes. J Neurochem. 2010;113:313–328. doi: 10.1111/j.1471-4159.2010.06626.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards DA, Zhang L, Alger BE. Metaplastic control of the endocannabinoid system at inhibitory synapses in hippocampus. Proc Natl Acad Sci U S A. 2008;105:8142–8147. doi: 10.1073/pnas.0803558105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egertova M, Simon GM, Cravatt BF, Elphick MR. Localization of N-acyl phosphatidylethanolamine phospholipase D (NAPE-PLD) expression in mouse brain: A new perspective on N-acylethanolamines as neural signaling molecules. J Comp Neurol. 2008;506:604–615. doi: 10.1002/cne.21568. [DOI] [PubMed] [Google Scholar]
- Freund TF, Katona I, Piomelli D. Role of endogenous cannabinoids in synaptic signaling. Physiol Rev. 2003;83:1017–1066. doi: 10.1152/physrev.00004.2003. [DOI] [PubMed] [Google Scholar]
- Fu J, Bottegoni G, Sasso O, Bertorelli R, Rocchia W, Masetti M, Guijarro A, Lodola A, Armirotti A, Garau G, et al. A catalytically silent FAAH-1 variant drives anandamide transport in neurons. Nat Neurosci. 2012;15:64–69. doi: 10.1038/nn.2986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao Y, Vasilyev DV, Goncalves MB, Howell FV, Hobbs C, Reisenberg M, Shen R, Zhang MY, Strassle BW, Lu P, et al. Loss of retrograde endocannabinoid signaling and reduced adult neurogenesis in diacylglycerol lipase knock-out mice. J Neurosci. 2010;30:2017–2024. doi: 10.1523/JNEUROSCI.5693-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerdeman GL, Ronesi J, Lovinger DM. Postsynaptic endocannabinoid release is critical to long-term depression in the striatum. Nat Neurosci. 2002;5:446–451. doi: 10.1038/nn832. [DOI] [PubMed] [Google Scholar]
- Gibson HE, Edwards JG, Page RS, Van Hook MJ, Kauer JA. TRPV1 channels mediate long-term depression at synapses on hippocampal interneurons. Neuron. 2008;57:746–759. doi: 10.1016/j.neuron.2007.12.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez O, Arevalo-Martin A, Garcia-Ovejero D, Ortega-Gutierrez S, Cisneros JA, Almazan G, Sanchez-Rodriguez MA, Molina-Holgado F, Molina-Holgado E. The constitutive production of the endocannabinoid 2-arachidonoylglycerol participates in oligodendrocyte differentiation. Glia. 2010;58:1913–1927. doi: 10.1002/glia.21061. [DOI] [PubMed] [Google Scholar]
- Grueter BA, Brasnjo G, Malenka RC. Postsynaptic TRPV1 triggers cell type-specific long-term depression in the nucleus accumbens. Nat Neurosci. 2010;13:1519–1525. doi: 10.1038/nn.2685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulyas AI, Cravatt BF, Bracey MH, Dinh TP, Piomelli D, Boscia F, Freund TF. Segregation of two endocannabinoid-hydrolyzing enzymes into pre- and postsynaptic compartments in the rat hippocampus, cerebellum and amygdala. Eur J Neurosci. 2004;20:441–458. doi: 10.1111/j.1460-9568.2004.03428.x. [DOI] [PubMed] [Google Scholar]
- Han J, Kesner P, Metna-Laurent M, Duan T, Xu L, Georges F, Koehl M, Abrous DN, Mendizabal-Zubiaga J, Grandes P, et al. Acute cannabinoids impair working memory through astroglial CB1 receptor modulation of hippocampal LTD. Cell. 2012;148:1039–1050. doi: 10.1016/j.cell.2012.01.037. [DOI] [PubMed] [Google Scholar]
- Harkany T, Mackie K, Doherty P. Wiring and firing neuronal networks: endocannabinoids take center stage. Curr Opin Neurobiol. 2008;18:338–345. doi: 10.1016/j.conb.2008.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimotodani Y, Ohno-Shosaku T, Kano M. Ca(2+)-assisted receptor-driven endocannabinoid release: mechanisms that associate presynaptic and postsynaptic activities. Curr Opin Neurobiol. 2007a;17:360–365. doi: 10.1016/j.conb.2007.03.012. [DOI] [PubMed] [Google Scholar]
- Hashimotodani Y, Ohno-Shosaku T, Kano M. Presynaptic monoacylglycerol lipase activity determines basal endocannabinoid tone and terminates retrograde endocannabinoid signaling in the hippocampus. J Neurosci. 2007b;27:1211–1219. doi: 10.1523/JNEUROSCI.4159-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimotodani Y, Ohno-Shosaku T, Tsubokawa H, Ogata H, Emoto K, Maejima T, Araishi K, Shin HS, Kano M. Phospholipase Cbeta serves as a coincidence detector through its Ca2+ dependency for triggering retrograde endocannabinoid signal. Neuron. 2005;45:257–268. doi: 10.1016/j.neuron.2005.01.004. [DOI] [PubMed] [Google Scholar]
- Hegyi Z, Hollo K, Kis G, Mackie K, Antal M. Differential distribution of diacylglycerol lipase-alpha and N-acylphosphatidylethanolamine-specific phospholipase d immunoreactivity in the superficial spinal dorsal horn of rats. Glia. 2012;60:1316–1329. doi: 10.1002/glia.22351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heifets BD, Castillo PE. Endocannabinoid signaling and long-term synaptic plasticity. Annu Rev Physiol. 2009;71:283–306. doi: 10.1146/annurev.physiol.010908.163149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heifets BD, Chevaleyre V, Castillo PE. Interneuron activity controls endocannabinoid-mediated presynaptic plasticity through calcineurin. Proc Natl Acad Sci U S A. 2008;105:10250–10255. doi: 10.1073/pnas.0711880105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hentges ST, Low MJ, Williams JT. Differential regulation of synaptic inputs by constitutively released endocannabinoids and exogenous cannabinoids. J Neurosci. 2005;25:9746–9751. doi: 10.1523/JNEUROSCI.2769-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herkenham M, Lynn AB, Little MD, Johnson MR, Melvin LS, de Costa BR, Rice KC. Cannabinoid receptor localization in brain. Proc Natl Acad Sci U S A. 1990;87:1932–1936. doi: 10.1073/pnas.87.5.1932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill EL, Gallopin T, Ferezou I, Cauli B, Rossier J, Schweitzer P, Lambolez B. Functional CB1 receptors are broadly expressed in neocortical GABAergic and glutamatergic neurons. J Neurophysiol. 2007;97:2580–2589. doi: 10.1152/jn.00603.2006. [DOI] [PubMed] [Google Scholar]
- Hillard CJ, Edgemond WS, Jarrahian A, Campbell WB. Accumulation of N-arachidonoylethanolamine (anandamide) into cerebellar granule cells occurs via facilitated diffusion. J Neurochem. 1997;69:631–638. doi: 10.1046/j.1471-4159.1997.69020631.x. [DOI] [PubMed] [Google Scholar]
- Hillard CJ, Weinlander KM, Stuhr KL. Contributions of endocannabinoid signaling to psychiatric disorders in humans: genetic and biochemical evidence. Neuroscience. 2012;204:207–229. doi: 10.1016/j.neuroscience.2011.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman AF, Laaris N, Kawamura M, Masino SA, Lupica CR. Control of cannabinoid CB1 receptor function on glutamate axon terminals by endogenous adenosine acting at A1 receptors. J Neurosci. 2010;30:545–555. doi: 10.1523/JNEUROSCI.4920-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howlett AC. Cannabinoid receptor signaling. Handb Exp Pharmacol. 2005:53–79. doi: 10.1007/3-540-26573-2_2. [DOI] [PubMed] [Google Scholar]
- Howlett AC, Barth F, Bonner TI, Cabral G, Casellas P, Devane WA, Felder CC, Herkenham M, Mackie K, Martin BR, et al. International Union of Pharmacology. XXVII. Classification of Cannabinoid Receptors. Pharmacol Rev. 2002;54:161–202. doi: 10.1124/pr.54.2.161. [DOI] [PubMed] [Google Scholar]
- Huang Y, Yasuda H, Sarihi A, Tsumoto T. Roles of endocannabinoids in heterosynaptic long-term depression of excitatory synaptic transmission in visual cortex of young mice. J Neurosci. 2008;28:7074–7083. doi: 10.1523/JNEUROSCI.0899-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hudson BD, Hebert TE, Kelly ME. Ligand- and heterodimer-directed signaling of the CB(1) cannabinoid receptor. Mol Pharmacol. 2010;77:1–9. doi: 10.1124/mol.109.060251. [DOI] [PubMed] [Google Scholar]
- Jin W, Brown S, Roche JP, Hsieh C, Celver JP, Kovoor A, Chavkin C, Mackie K. Distinct domains of the CB1 cannabinoid receptor mediate desensitization and internalization. J Neurosci. 1999;19:3773–3780. doi: 10.1523/JNEUROSCI.19-10-03773.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang-Park MH, Wilson WA, Kuhn CM, Moore SD, Swartzwelder HS. Differential sensitivity of GABA A receptor-mediated IPSCs to cannabinoids in hippocampal slices from adolescent and adult rats. J Neurophysiol. 2007;98:1223–1230. doi: 10.1152/jn.00091.2007. [DOI] [PubMed] [Google Scholar]
- Kano M, Ohno-Shosaku T, Hashimotodani Y, Uchigashima M, Watanabe M. Endocannabinoid-mediated control of synaptic transmission. Physiol Rev. 2009;89:309–380. doi: 10.1152/physrev.00019.2008. [DOI] [PubMed] [Google Scholar]
- Katona I, Freund TF. Multiple functions of endocannabinoid signaling in the brain. Annu Rev Neurosci. 2012;35:529–558. doi: 10.1146/annurev-neuro-062111-150420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katona I, Sperlagh B, Sik A, Kafalvi A, Vizi ES, Mackie K, Freund TF. Presynaptically located CB1 cannabinoid receptors regulate GABA release from axon terminals of specific hippocampal interneurons. J Neurosci. 1999;19:4544–4558. doi: 10.1523/JNEUROSCI.19-11-04544.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katona I, Urban GM, Wallace M, Ledent C, Jung KM, Piomelli D, Mackie K, Freund TF. Molecular composition of the endocannabinoid system at glutamatergic synapses. J Neurosci. 2006;26:5628–5637. doi: 10.1523/JNEUROSCI.0309-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawamura Y, Fukaya M, Maejima T, Yoshida T, Miura E, Watanabe M, Ohno-Shosaku T, Kano M. The CB1 cannabinoid receptor is the major cannabinoid receptor at excitatory presynaptic sites in the hippocampus and cerebellum. J Neurosci. 2006;26:2991–3001. doi: 10.1523/JNEUROSCI.4872-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J, Alger BE. Reduction in endocannabinoid tone is a homeostatic mechanism for specific inhibitory synapses. Nat Neurosci. 2010;13:592–600. doi: 10.1038/nn.2517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreitzer AC, Regehr WG. Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto Purkinje cells. Neuron. 2001;29:717–727. doi: 10.1016/s0896-6273(01)00246-x. [DOI] [PubMed] [Google Scholar]
- Lafourcade M, Elezgarai I, Mato S, Bakiri Y, Grandes P, Manzoni OJ. Molecular components and functions of the endocannabinoid system in mouse prefrontal cortex. PLoS ONE. 2007;2:e709. doi: 10.1371/journal.pone.0000709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lafourcade M, Larrieu T, Mato S, Duffaud A, Sepers M, Matias I, De Smedt-Peyrusse V, Labrousse VF, Bretillon L, Matute C, et al. Nutritional omega-3 deficiency abolishes endocannabinoid-mediated neuronal functions. Nat Neurosci. 2011;14:345–350. doi: 10.1038/nn.2736. [DOI] [PubMed] [Google Scholar]
- Lerner TN, Kreitzer AC. RGS4 is required for dopaminergic control of striatal LTD and susceptibility to Parkinsonian motor deficits. Neuron. 2012;73:347–359. doi: 10.1016/j.neuron.2011.11.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ligresti A, Petrosino S, Di Marzo V. From endocannabinoid profiling to ‘endocannabinoid therapeutics’. Curr Opin Chem Biol. 2009;13:321–331. doi: 10.1016/j.cbpa.2009.04.615. [DOI] [PubMed] [Google Scholar]
- Liu J, Wang L, Harvey-White J, Huang BX, Kim HY, Luquet S, Palmiter RD, Krystal G, Rai R, Mahadevan A, et al. Multiple pathways involved in the biosynthesis of anandamide. Neuropharmacology. 2008;54:1–7. doi: 10.1016/j.neuropharm.2007.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Losonczy A, Biro AA, Nusser Z. Persistently active cannabinoid receptors mute a subpopulation of hippocampal interneurons. Proc Natl Acad Sci U S A. 2004;101:1362–1367. doi: 10.1073/pnas.0304752101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lourenco J, Cannich A, Carta M, Coussen F, Mulle C, Marsicano G. Synaptic activation of kainate receptors gates presynaptic CB(1) signaling at GABAergic synapses. Nat Neurosci. 2010;13:197–204. doi: 10.1038/nn.2481. [DOI] [PubMed] [Google Scholar]
- Ludanyi A, Eross L, Czirjak S, Vajda J, Halasz P, Watanabe M, Palkovits M, Magloczky Z, Freund TF, Katona I. Downregulation of the CB1 cannabinoid receptor and related molecular elements of the endocannabinoid system in epileptic human hippocampus. J Neurosci. 2008;28:2976–2990. doi: 10.1523/JNEUROSCI.4465-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludanyi A, Hu SS, Yamazaki M, Tanimura A, Piomelli D, Watanabe M, Kano M, Sakimura K, Magloczky Z, Mackie K, et al. Complementary synaptic distribution of enzymes responsible for synthesis and inactivation of the endocannabinoid 2-arachidonoylglycerol in the human hippocampus. Neuroscience. 2011;174:50–63. doi: 10.1016/j.neuroscience.2010.10.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maccarrone M, Rossi S, Bari M, De Chiara V, Fezza F, Musella A, Gasperi V, Prosperetti C, Bernardi G, Finazzi-Agro A, et al. Anandamide inhibits metabolism and physiological actions of 2-arachidonoylglycerol in the striatum. Nat Neurosci. 2008;11:152–159. doi: 10.1038/nn2042. [DOI] [PubMed] [Google Scholar]
- Mackie K. Cannabinoid receptor homo- and heterodimerization. Life sciences. 2005;77:1667–1673. doi: 10.1016/j.lfs.2005.05.011. [DOI] [PubMed] [Google Scholar]
- Maejima T, Hashimoto K, Yoshida T, Aiba A, Kano M. Presynaptic inhibition caused by retrograde signal from metabotropic glutamate to cannabinoid receptors. Neuron. 2001;31:463–475. doi: 10.1016/s0896-6273(01)00375-0. [DOI] [PubMed] [Google Scholar]
- Maejima T, Oka S, Hashimotodani Y, Ohno-Shosaku T, Aiba A, Wu D, Waku K, Sugiura T, Kano M. Synaptically driven endocannabinoid release requires Ca2+-assisted metabotropic glutamate receptor subtype 1 to phospholipase Cbeta4 signaling cascade in the cerebellum. J Neurosci. 2005;25:6826–6835. doi: 10.1523/JNEUROSCI.0945-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marinelli S, Pacioni S, Bisogno T, Di Marzo V, Prince DA, Huguenard JR, Bacci A. The endocannabinoid 2-arachidonoylglycerol is responsible for the slow self-inhibition in neocortical interneurons. J Neurosci. 2008;28:13532–13541. doi: 10.1523/JNEUROSCI.0847-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marinelli S, Pacioni S, Cannich A, Marsicano G, Bacci A. Self-modulation of neocortical pyramidal neurons by endocannabinoids. Nat Neurosci. 2009;12:1488–1490. doi: 10.1038/nn.2430. [DOI] [PubMed] [Google Scholar]
- Marrs WR, Blankman JL, Horne EA, Thomazeau A, Lin YH, Coy J, Bodor AL, Muccioli GG, Hu SS, Woodruff G, et al. The serine hydrolase ABHD6 controls the accumulation and efficacy of 2-AG at cannabinoid receptors. Nat Neurosci. 2010;13:951–957. doi: 10.1038/nn.2601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsicano G, Wotjak CT, Azad SC, Bisogno T, Rammes G, Cascio MG, Hermann H, Tang J, Hofmann C, Zieglgansberger W, et al. The endogenous cannabinoid system controls extinction of aversive memories. Nature. 2002;418:530–534. doi: 10.1038/nature00839. [DOI] [PubMed] [Google Scholar]
- Massa F, Mancini G, Schmidt H, Steindel F, Mackie K, Angioni C, Oliet SH, Geisslinger G, Lutz B. Alterations in the hippocampal endocannabinoid system in diet-induced obese mice. J Neurosci. 2010;30:6273–6281. doi: 10.1523/JNEUROSCI.2648-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mato S, Chevaleyre V, Robbe D, Pazos A, Castillo PE, Manzoni OJ. A single in-vivo exposure to delta 9THC blocks endocannabinoid-mediated synaptic plasticity. Nat Neurosci. 2004;7:585–586. doi: 10.1038/nn1251. [DOI] [PubMed] [Google Scholar]
- Mato S, Lafourcade M, Robbe D, Bakiri Y, Manzoni OJ. Role of the cyclic-AMP/PKA cascade and of P/Q-type Ca(++) channels in endocannabinoid-mediated long-term depression in the nucleus accumbens. Neuropharmacology. 2008;54:87–94. doi: 10.1016/j.neuropharm.2007.04.014. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Parker LA. The Endocannabinoid System and the Brain. Annu Rev Psychol. 2012 doi: 10.1146/annurev-psych-113011-143739. [DOI] [PubMed] [Google Scholar]
- Mezey E, Toth ZE, Cortright DN, Arzubi MK, Krause JE, Elde R, Guo A, Blumberg PM, Szallasi A. Distribution of mRNA for vanilloid receptor subtype 1 (VR1), and VR1-like immunoreactivity, in the central nervous system of the rat and human. Proc Natl Acad Sci U S A. 2000;97:3655–3660. doi: 10.1073/pnas.060496197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mikasova L, Groc L, Choquet D, Manzoni OJ. Altered surface trafficking of presynaptic cannabinoid type 1 receptor in and out synaptic terminals parallels receptor desensitization. Proc Natl Acad Sci U S A. 2008;105:18596–18601. doi: 10.1073/pnas.0805959105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Min R, Nevian T. Astrocyte signaling controls spike timing-dependent depression at neocortical synapses. Nat Neurosci. 2012;15:746–753. doi: 10.1038/nn.3075. [DOI] [PubMed] [Google Scholar]
- Navarrete M, Araque A. Endocannabinoids mediate neuron-astrocyte communication. Neuron. 2008;57:883–893. doi: 10.1016/j.neuron.2008.01.029. [DOI] [PubMed] [Google Scholar]
- Navarrete M, Araque A. Endocannabinoids potentiate synaptic transmission through stimulation of astrocytes. Neuron. 2010;68:113–126. doi: 10.1016/j.neuron.2010.08.043. [DOI] [PubMed] [Google Scholar]
- Neu A, Foldy C, Soltesz I. Postsynaptic origin of CB1-dependent tonic inhibition of GABA release at cholecystokinin-positive basket cell to pyramidal cell synapses in the CA1 region of the rat hippocampus. J Physiol. 2007;578:233–247. doi: 10.1113/jphysiol.2006.115691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevian T, Sakmann B. Spine Ca2+ signaling in spike-timing-dependent plasticity. J Neurosci. 2006;26:11001–11013. doi: 10.1523/JNEUROSCI.1749-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nomura DK, Morrison BE, Blankman JL, Long JZ, Kinsey SG, Marcondes MC, Ward AM, Hahn YK, Lichtman AH, Conti B, Cravatt BF. Endocannabinoid hydrolysis generates brain prostaglandins that promote neuroinflammation. Science. 2011;334:809–813. doi: 10.1126/science.1209200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nomura S, Fukaya M, Tsujioka T, Wu D, Watanabe M. Phospholipase Cbeta3 is distributed in both somatodendritic and axonal compartments and localized around perisynapse and smooth endoplasmic reticulum in mouse Purkinje cell subsets. Eur J Neurosci. 2007;25:659–672. doi: 10.1111/j.1460-9568.2007.05334.x. [DOI] [PubMed] [Google Scholar]
- Nyilas R, Dudok B, Urban GM, Mackie K, Watanabe M, Cravatt BF, Freund TF, Katona I. Enzymatic machinery for endocannabinoid biosynthesis associated with calcium stores in glutamatergic axon terminals. J Neurosci. 2008;28:1058–1063. doi: 10.1523/JNEUROSCI.5102-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohno-Shosaku T, Maejima T, Kano M. Endogenous cannabinoids mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. Neuron. 2001;29:729–738. doi: 10.1016/s0896-6273(01)00247-1. [DOI] [PubMed] [Google Scholar]
- Okamoto Y, Wang J, Morishita J, Ueda N. Biosynthetic pathways of the endocannabinoid anandamide. Chem Biodivers. 2007;4:1842–1857. doi: 10.1002/cbdv.200790155. [DOI] [PubMed] [Google Scholar]
- Oliet SH, Baimoukhametova DV, Piet R, Bains JS. Retrograde regulation of GABA transmission by the tonic release of oxytocin and endocannabinoids governs postsynaptic firing. J Neurosci. 2007;27:1325–1333. doi: 10.1523/JNEUROSCI.2676-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan B, Hillard CJ, Liu QS. D2 dopamine receptor activation facilitates endocannabinoid-mediated long-term synaptic depression of GABAergic synaptic transmission in midbrain dopamine neurons via cAMP-protein kinase A signaling. J Neurosci. 2008;28:14018–14030. doi: 10.1523/JNEUROSCI.4035-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan B, Wang W, Zhong P, Blankman JL, Cravatt BF, Liu QS. Alterations of endocannabinoid signaling, synaptic plasticity, learning, and memory in monoacylglycerol lipase knock-out mice. J Neurosci. 2011;31:13420–13430. doi: 10.1523/JNEUROSCI.2075-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel S, Kingsley PJ, Mackie K, Marnett LJ, Winder DG. Repeated homotypic stress elevates 2-arachidonoylglycerol levels and enhances short-term endocannabinoid signaling at inhibitory synapses in basolateral amygdala. Neuropsychopharmacology. 2009;34:2699–2709. doi: 10.1038/npp.2009.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee RG, Howlett AC, Abood ME, Alexander SP, Di Marzo V, Elphick MR, Greasley PJ, Hansen HS, Kunos G, Mackie K, et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: beyond CB(1) and CB(2) Pharmacol Rev. 2010;62:588–631. doi: 10.1124/pr.110.003004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piomelli D. The molecular logic of endocannabinoid signalling. Nat Rev Neurosci. 2003;4:873–884. doi: 10.1038/nrn1247. [DOI] [PubMed] [Google Scholar]
- Piomelli D. The endocannabinoid system: a drug discovery perspective. Curr Opin Investig Drugs. 2005;6:672–679. [PubMed] [Google Scholar]
- Puente N, Cui Y, Lassalle O, Lafourcade M, Georges F, Venance L, Grandes P, Manzoni OJ. Polymodal activation of the endocannabinoid system in the extended amygdala. Nat Neurosci. 2011;14:1542–1547. doi: 10.1038/nn.2974. [DOI] [PubMed] [Google Scholar]
- Regehr WG, Carey MR, Best AR. Activity-dependent regulation of synapses by retrograde messengers. Neuron. 2009;63:154–170. doi: 10.1016/j.neuron.2009.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riebe CJ, Wotjak CT. Endocannabinoids and stress. Stress. 2011;14:384–397. doi: 10.3109/10253890.2011.586753. [DOI] [PubMed] [Google Scholar]
- Robbe D, Kopf M, Remaury A, Bockaert J, Manzoni OJ. Endogenous cannabinoids mediate long-term synaptic depression in the nucleus accumbens. Proc Natl Acad Sci U S A. 2002;99:8384–8388. doi: 10.1073/pnas.122149199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts JC, Davis JB, Benham CD. [3H]Resiniferatoxin autoradiography in the CNS of wild-type and TRPV1 null mice defines TRPV1 (VR-1) protein distribution. Brain Res. 2004;995:176–183. doi: 10.1016/j.brainres.2003.10.001. [DOI] [PubMed] [Google Scholar]
- Roloff AM, Anderson GR, Martemyanov KA, Thayer SA. Homer 1a gates the induction mechanism for endocannabinoid-mediated synaptic plasticity. J Neurosci. 2010;30:3072–3081. doi: 10.1523/JNEUROSCI.4603-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi S, De Chiara V, Musella A, Kusayanagi H, Mataluni G, Bernardi G, Usiello A, Centonze D. Chronic psychoemotional stress impairs cannabinoid-receptor-mediated control of GABA transmission in the striatum. J Neurosci. 2008;28:7284–7292. doi: 10.1523/JNEUROSCI.5346-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlicker E, Kathmann M. Modulation of transmitter release via presynaptic cannabinoid receptors. Trends Pharmacol Sci. 2001;22:565–572. doi: 10.1016/s0165-6147(00)01805-8. [DOI] [PubMed] [Google Scholar]
- Schlosburg JE, Blankman JL, Long JZ, Nomura DK, Pan B, Kinsey SG, Nguyen PT, Ramesh D, Booker L, Burston JJ, et al. Chronic monoacylglycerol lipase blockade causes functional antagonism of the endocannabinoid system. Nat Neurosci. 2010;13:1113–1119. doi: 10.1038/nn.2616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singla S, Kreitzer AC, Malenka RC. Mechanisms for synapse specificity during striatal long-term depression. J Neurosci. 2007;27:5260–5264. doi: 10.1523/JNEUROSCI.0018-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sjostrom PJ, Turrigiano GG, Nelson SB. Neocortical LTD via coincident activation of presynaptic NMDA and cannabinoid receptors. Neuron. 2003;39:641–654. doi: 10.1016/s0896-6273(03)00476-8. [DOI] [PubMed] [Google Scholar]
- Slanina KA, Schweitzer P. Inhibition of cyclooxygenase-2 elicits a CB1-mediated decrease of excitatory transmission in rat CA1 hippocampus. Neuropharmacology. 2005;49:653–659. doi: 10.1016/j.neuropharm.2005.04.019. [DOI] [PubMed] [Google Scholar]
- Smart D, Gunthorpe MJ, Jerman JC, Nasir S, Gray J, Muir AI, Chambers JK, Randall AD, Davis JB. The endogenous lipid anandamide is a full agonist at the human vanilloid receptor (hVR1) Br J Pharmacol. 2000;129:227–230. doi: 10.1038/sj.bjp.0703050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stella N. Endocannabinoid signaling in microglial cells. Neuropharmacology. 2009;56(Suppl 1):244–253. doi: 10.1016/j.neuropharm.2008.07.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stella N. Cannabinoid and cannabinoid-like receptors in microglia, astrocytes, and astrocytomas. Glia. 2010;58:1017–1030. doi: 10.1002/glia.20983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szabo B, Urbanski MJ, Bisogno T, Di Marzo V, Mendiguren A, Baer WU, Freiman I. Depolarization-induced retrograde synaptic inhibition in the mouse cerebellar cortex is mediated by 2-arachidonoylglycerol. J Physiol. 2006;577:263–280. doi: 10.1113/jphysiol.2006.119362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi KA, Castillo PE. The CB1 cannabinoid receptor mediates glutamatergic synaptic suppression in the hippocampus. Neuroscience. 2006;139:795–802. doi: 10.1016/j.neuroscience.2006.01.024. [DOI] [PubMed] [Google Scholar]
- Tanimura A, Uchigashima M, Yamazaki M, Uesaka N, Mikuni T, Abe M, Hashimoto K, Watanabe M, Sakimura K, Kano M. Synapse type-independent degradation of the endocannabinoid 2-arachidonoylglycerol after retrograde synaptic suppression. Proc Natl Acad Sci U S A. 2012;109:12195–12200. doi: 10.1073/pnas.1204404109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanimura A, Yamazaki M, Hashimotodani Y, Uchigashima M, Kawata S, Abe M, Kita Y, Hashimoto K, Shimizu T, Watanabe M, et al. The endocannabinoid 2-arachidonoylglycerol produced by diacylglycerol lipase alpha mediates retrograde suppression of synaptic transmission. Neuron. 2010;65:320–327. doi: 10.1016/j.neuron.2010.01.021. [DOI] [PubMed] [Google Scholar]
- Toth A, Boczan J, Kedei N, Lizanecz E, Bagi Z, Papp Z, Edes I, Csiba L, Blumberg PM. Expression and distribution of vanilloid receptor 1 (TRPV1) in the adult rat brain. Brain Res Mol Brain Res. 2005;135:162–168. doi: 10.1016/j.molbrainres.2004.12.003. [DOI] [PubMed] [Google Scholar]
- Tsetsenis T, Younts TJ, Chiu CQ, Kaeser PS, Castillo PE, Sudhof TC. Rab3B protein is required for long-term depression of hippocampal inhibitory synapses and for normal reversal learning. Proc Natl Acad Sci U S A. 2011;108:14300–14305. doi: 10.1073/pnas.1112237108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uchigashima M, Yamazaki M, Yamasaki M, Tanimura A, Sakimura K, Kano M, Watanabe M. Molecular and morphological configuration for 2-arachidonoylglycerol-mediated retrograde signaling at mossy cell-granule cell synapses in the dentate gyrus. J Neurosci. 2011;31:7700–7714. doi: 10.1523/JNEUROSCI.5665-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Beugen BJ, Nagaraja RY, Hansel C. Climbing fiber-evoked endocannabinoid signaling heterosynaptically suppresses presynaptic cerebellar long-term potentiation. J Neurosci. 2006;26:8289–8294. doi: 10.1523/JNEUROSCI.0805-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Stelt M, Di Marzo V. Anandamide as an intracellular messenger regulating ion channel activity. Prostaglandins Other Lipid Mediat. 2005;77:111–122. doi: 10.1016/j.prostaglandins.2004.09.007. [DOI] [PubMed] [Google Scholar]
- Van Sickle MD, Duncan M, Kingsley PJ, Mouihate A, Urbani P, Mackie K, Stella N, Makriyannis A, Piomelli D, Davison JS, et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science. 2005;310:329–332. doi: 10.1126/science.1115740. [DOI] [PubMed] [Google Scholar]
- Vandevoorde S, Lambert DM. The multiple pathways of endocannabinoid metabolism: a zoom out. Chem Biodivers. 2007;4:1858–1881. doi: 10.1002/cbdv.200790156. [DOI] [PubMed] [Google Scholar]
- Varma N, Carlson GC, Ledent C, Alger BE. Metabotropic glutamate receptors drive the endocannabinoid system in hippocampus. J Neurosci. 2001;21:RC188. doi: 10.1523/JNEUROSCI.21-24-j0003.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wamsteeker JI, Kuzmiski JB, Bains JS. Repeated stress impairs endocannabinoid signaling in the paraventricular nucleus of the hypothalamus. J Neurosci. 2010;30:11188–11196. doi: 10.1523/JNEUROSCI.1046-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang M, Hill MN, Zhang L, Gorzalka BB, Hillard CJ, Alger BE. Acute restraint stress enhances hippocampal endocannabinoid function via glucocorticoid receptor activation. Journal of psychopharmacology. 2012;26:56–70. doi: 10.1177/0269881111409606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang W, Sun D, Pan B, Roberts CJ, Sun X, Hillard CJ, Liu QS. Deficiency in endocannabinoid signaling in the nucleus accumbens induced by chronic unpredictable stress. Neuropsychopharmacology. 2010;35:2249–2261. doi: 10.1038/npp.2010.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson RI, Kunos G, Nicoll RA. Presynaptic specificity of endocannabinoid signaling in the hippocampus. Neuron. 2001;31:453–462. doi: 10.1016/s0896-6273(01)00372-5. [DOI] [PubMed] [Google Scholar]
- Wilson RI, Nicoll RA. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature. 2001;410:588–592. doi: 10.1038/35069076. [DOI] [PubMed] [Google Scholar]
- Wu DF, Yang LQ, Goschke A, Stumm R, Brandenburg LO, Liang YJ, Hollt V, Koch T. Role of receptor internalization in the agonist-induced desensitization of cannabinoid type 1 receptors. J Neurochem. 2008;104:1132–1143. doi: 10.1111/j.1471-4159.2007.05063.x. [DOI] [PubMed] [Google Scholar]
- Xi ZX, Peng XQ, Li X, Song R, Zhang HY, Liu QR, Yang HJ, Bi GH, Li J, Gardner EL. Brain cannabinoid CB(2) receptors modulate cocaine’s actions in mice. Nat Neurosci. 2011;14:1160–1166. doi: 10.1038/nn.2874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuda H, Huang Y, Tsumoto T. Regulation of excitability and plasticity by endocannabinoids and PKA in developing hippocampus. Proc Natl Acad Sci U S A. 2008;105:3106–3111. doi: 10.1073/pnas.0708349105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida T, Fukaya M, Uchigashima M, Miura E, Kamiya H, Kano M, Watanabe M. Localization of diacylglycerol lipase-alpha around postsynaptic spine suggests close proximity between production site of an endocannabinoid, 2-arachidonoyl-glycerol, and presynaptic cannabinoid CB1 receptor. J Neurosci. 2006;26:4740–4751. doi: 10.1523/JNEUROSCI.0054-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida T, Uchigashima M, Yamasaki M, Katona I, Yamazaki M, Sakimura K, Kano M, Yoshioka M, Watanabe M. Unique inhibitory synapse with particularly rich endocannabinoid signaling machinery on pyramidal neurons in basal amygdaloid nucleus. Proc Natl Acad Sci U S A. 2011;108:3059–3064. doi: 10.1073/pnas.1012875108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshino H, Miyamae T, Hansen G, Zambrowicz B, Flynn M, Pedicord D, Blat Y, Westphal RS, Zaczek R, Lewis DA, Gonzalez-Burgos G. Postsynaptic diacylglycerol lipase mediates retrograde endocannabinoid suppression of inhibition in mouse prefrontal cortex. J Physiol. 2011;589:4857–4884. doi: 10.1113/jphysiol.2011.212225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang L, Alger BE. Enhanced endocannabinoid signaling elevates neuronal excitability in fragile X syndrome. J Neurosci. 2010;30:5724–5729. doi: 10.1523/JNEUROSCI.0795-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong P, Pan B, Gao XP, Blankman JL, Cravatt BF, Liu QS. Genetic deletion of monoacylglycerol lipase alters endocannabinoid-mediated retrograde synaptic depression in the cerebellum. J Physiol. 2011;589:4847–4855. doi: 10.1113/jphysiol.2011.215509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu PJ, Lovinger DM. Persistent synaptic activity produces long-lasting enhancement of endocannabinoid modulation and alters long-term synaptic plasticity. J Neurophysiol. 2007;97:4386–4389. doi: 10.1152/jn.01228.2006. [DOI] [PubMed] [Google Scholar]
- Zhu PJ, Lovinger DM. Developmental alteration of endocannabinoid retrograde signaling in the hippocampus. J Neurophysiol. 2010;103:1123–1129. doi: 10.1152/jn.00327.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zygmunt PM, Petersson J, Andersson DA, Chuang H, Sorgard M, Di Marzo V, Julius D, Hogestatt ED. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature. 1999;400:452–457. doi: 10.1038/22761. [DOI] [PubMed] [Google Scholar]