

Abstract

Class II diterpene cyclases catalyze bicyclization of geranylgeranyl diphosphate. While this reaction typically is terminated via methyl deprotonation to yield copalyl diphosphate, in rare cases hydroxylated bicycles are produced instead. Abietadiene synthase is a bifunctional diterpene cyclase that usually produces a copalyl diphosphate intermediate. Here it is shown that substitution of aspartate for a conserved histidine in the class II active site of abietadiene synthase leads to selective production of 8α-hydroxy-CPP instead, demonstrating striking plasticity.
The labane-related diterpenoids form a natural products superfamily, consisting of ~7,000 known compounds, whose biosynthesis is characterized by (bi)cyclization of (E,E,E)-geranylgeranyl diphosphate (GGPP; 1), catalyzed via a protonation-initiated carbocationic cascade reaction by class II diterpene cyclases.1 The diterpene cyclases involved in conifer resin acid biosynthesis are bifunctional enzymes that catalyze such a class II cyclization reaction, producing CPP (2) from 1 (Scheme 1).2
Scheme 1.

Class II reactions mediated by wild-type and the H348D mutant of AgAS.
Class II diterpene cyclases are characterized by a DxDD motif, wherein the central aspartic acid residue acts as the catalytic acid.3 However, the catalytic base for this reaction remains unknown despite previously reported extensive mutagenesis.4 Given our recent determination of a crystal structure for the abietadiene synthase from Abies grandis (AgAS),5 we set out to identify the catalytic base in its class II active site, resulting in the surprising finding that substitutions for one potential candidate, histidine 348, led to mutant AgAS that catalyze class II cyclization reactions leading to an unusual hydroxylated product.
While the DxDD motif that cooperatively acts as the catalytic acid is conserved in class II diterpene cyclases, no conserved residue that might act as the catalytic base has yet been identified. This may be due to the variation in bound substrate configuration implied by the production of distinct stereoisomers of CPP by different such enzymes.1 Accordingly, we were intrigued by the observation that the class II active site of AgAS contains a histidine (at position 348) that is not only appropriately positioned to act as the base – i.e., across the substrate binding cavity from the DxDD motif (Figure 1) – but is further conserved among the homologous diterpene synthases involved in gymnosperm resin acid biosynthesis, all of which produce the normal (9S,10S) stereoisomer of CPP (2).
Figure 1.
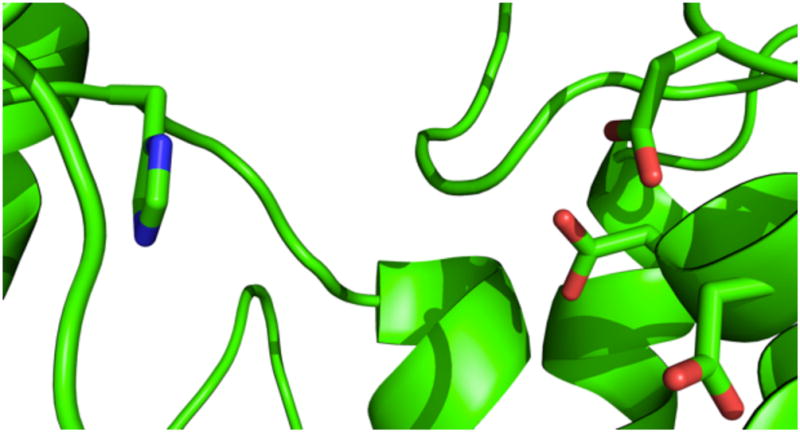
Class II active site of AgAS (backbone shown in cartoon format), with the side chains of the aspartates of the DxDD motif and histidine targeted here shown in stick representation.
Hypothesizing that this might be the catalytic base, we substituted alanine for His348, and tested the activity of the resulting AgAS:H348A mutant in the context of the D621A substitution that eliminates the subsequently acting class I activity.6 Intriguingly, rather than abolishing activity, this mutation led to novel products. In particular, AgAS:H348A/D621A reacted with 1, both in vitro and in our modular metabolic engineering system (whereby Escherichia coli is engineered to produce 1 for use by co-expressed diterpene synthases such as AgAS7), to yield a hydroxylated product, observed as a dephosphorylated diol by GC-MS analysis (Figure S1).
The mass spectra of this compound resembled that previously reported for labd-13E-en-8α,15-diol,8 which results from dephosphorylation of 8α-hydroxy-CPP (3), a known class II diterpene cyclase product.8–13 Comparison with the 3 produced by the relevant enzyme from Nicotiana glutinosa (NgCLS)9 indicated that AgAS:H348A/D621A also produces 3 (Figure S1). Moreover, by increasing metabolic flux to isoprenoids (i.e. 1) in our metabolic engineering system,14 it was possible to isolate sufficient amounts of labd-13E-en-8α,15-diol for verification by NMR spectral analysis (Table S1).
Strikingly, a recently reported cis-abienol synthase from Abies balsamea, homologous to AgAS, similarly proceeds through 3 as the stable intermediate product of its class II activity and contains an aspartate in place of the histidine targeted here, which is otherwise conserved in the known gymnosperm bifunctional diterpene cyclases.15 Thus, we constructed the corresponding AgAS:H348D/D621A mutant, which largely produced 3 (Figure 2), rather than the more evenly distributed mixture of products observed with AgAS:H348A/D621A. These products included not only 2 and 3, but also a double bond isomer of 2, labda-7Z,13E-dienyl diphosphate (4), as identified by comparison of the dephosphorylated alcohol to a previously identified enzymatic product (Figure S1).16
Figure 2.

Effect of H348D mutation on product outcome of the class II cyclization reaction mediated by AgAS. Chromatograms from GC-MS analysis of the dephosphorylated products (also shown is the structure of 4).
It has been indicated that the addition of water in class II cyclization reactions can only occur when the water molecule is in the optimal position and orientation for addition to the empty 2p orbital of the relevant carbocation intermediate, as the water would otherwise serve as a general base.17 Thus, the ability of AgAS:H348A/D621A to produce a mixture of compounds resulting from either direct deprotonation of the labda-13E-en-8-yl carbocation intermediate (5+) initially formed by bicyclization (i.e., to yield 2 or 4), or after the addition of water (i.e., to yield 3), suggests the use of water as a base, at least in this mutant.
Based on these results, we hypothesize that the histidine residue found in the wild-type enzyme positions a water molecule to act as the catalytic base, while aspartate substitution positions this water for addition, and the presence of an alanine simply allows variable positioning. In those cases where water is added, the resulting oxacarbenium ion must still be deprotonated to form a stable product, and we further hypothesize that an additional water molecule in the class II active site may serve this purpose.
Notably, the observed specific production of the 8α stereoisomer indicates distal attack of water on the empty 2p orbital of C8 following initial (bi)cyclization (i.e., in 5+). Such distal addition of water, as well as the observed products resulting from direct deprotonation (as noted elsewhere for 2 and 416), all are consistent with a concerted cyclization mechanism (Scheme 2). Although it has been suggested elsewhere that distal positioning of water for addition is required for class II cyclization, as a water positioned for proximal attack might interfere sterically with the preceding cyclization,17 it should be noted that NgCLS has been suggested to produce a small amount of the 8β-hydroxy epimer (~10%),9 hinting at some ability to continue beyond the initial (presumably concerted) (bi)cyclization, which would be consistent with the observed production of rearranged products by class II diterpene cyclases (e.g., halimadienyl18 or clerodadienyl19 diphosphates).1
Scheme 2.

Roles for water in class II cyclization reactions catalyzed by AgAS or the mutants reported here.
Regardless of exact mechanism, the ability of simple substitution of an aspartate for histidine, which requires only a single nucleotide change, to selectively change final product outcome is striking. While the ability of single residue changes to substantially alter product outcome in class I diterpene synthases has been previously demonstrated,20–24 the ability of such a change to alter chemical composition of the class II cyclization reaction product was unexpected. Nevertheless, the results reported here demonstrate intriguing plasticity for at least the class II diterpene cyclase active sites of the bifunctional diterpene synthases from gymnosperm resin acid biosynthesis as well. This change further has implications for the derived diterpenoids (Scheme 3), as the exocyclic-8(17)-double bond of 2 is involved in the subsequent cyclization typically catalyzed by the downstream class I diterpene synthases – e.g., to form pimaradienes (e.g., 6) and abietanes (e.g., 7). In addition, while class I terpene synthases also can add hydroxyl groups (e.g., in formation of 725), the presence of a hydroxyl in 3 offers further chemical diversity – e.g., the direct production of cis-abienol (8)15 or a diol such as sclareol (9).11,12
Scheme 3.

Products of class I diterpene synthases catalyzed reactions with 2 or 3.
Supplementary Material
Acknowledgments
This work was supported by a grant from the NIH (GM076324) to R.J.P., while F.S. was supported by a BBSRC CASE studentship to M.H.B. and Quest International. We thank Dr. Jiachen Zi (Iowa State University) for carrying out NMR analysis of labd-13E-en-8α,15-diol.
Footnotes
Supporting Information Available Additional figure illustrating AgAS:H348A/D621A product analysis. Description of experimental methods. NMR chemical shift data for labda-13E-en-8α,15-diol. This material is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.Peters RJ. Nat Prod Rep. 2010;27:1521. doi: 10.1039/c0np00019a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Keeling CI, Bohlmann J. Phytochemistry. 2006;67:2415. doi: 10.1016/j.phytochem.2006.08.019. [DOI] [PubMed] [Google Scholar]
- 3.Prisic S, Xu J, Coates RM, Peters RJ. Chem Bio Chem. 2007;8:869. doi: 10.1002/cbic.200700045. [DOI] [PubMed] [Google Scholar]
- 4.Peters RJ, Croteau RB. Biochemistry. 2002;41:1836. doi: 10.1021/bi011879d. [DOI] [PubMed] [Google Scholar]
- 5.Zhou K, Gao Y, Hoy JA, Mann FM, Honzatko RB, Peters RJ. J Biol Chem. 2012;287:6840. doi: 10.1074/jbc.M111.337592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Peters RJ, Ravn MM, Coates RM, Croteau RB. J Am Chem Soc. 2001;123:8974. doi: 10.1021/ja010670k. [DOI] [PubMed] [Google Scholar]
- 7.Cyr A, Wilderman PR, Determan M, Peters RJ. J Am Chem Soc. 2007;129:6684. doi: 10.1021/ja071158n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Falara V, Pichersky E, Kanellis AK. Plant Physiol. 2010;154:301. doi: 10.1104/pp.110.159566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shephard F. University of Bristol. 2002. [Google Scholar]
- 10.Sallaud C, Giacalone C, Topfer R, Goepfert S, Bakaher N, Rosti S, Tissier A. Plant J. 2012 doi: 10.1111/j.1365-313X.2012.05068.x. epub. [DOI] [PubMed] [Google Scholar]
- 11.Caniard A, Zerbe P, Legrand S, Cohade A, Valot N, Magnard JL, Bohlmann J, Legendre L. BMC Plant Biol. 2012;12:119. doi: 10.1186/1471-2229-12-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schalk M, Pastore L, Mirata MA, Khim S, Schouwey M, Deguerry F, Pineda V, Rocci L, Daviet L. J Am Chem Soc. 2012 doi: 10.1021/ja307404u. [DOI] [PubMed] [Google Scholar]
- 13.Gunnewich N, Higashi Y, Feng X, Choi KB, Schmidt J, Kutchan TM. Phytochemistry. 2012 doi: 10.1016/j.phytochem.2012.07.019. [DOI] [PubMed] [Google Scholar]
- 14.Morrone D, Lowry L, Determan MK, Hershey DM, Xu M, Peters RJ. Appl Microbiol Biotechnol. 2010;85:1893. doi: 10.1007/s00253-009-2219-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zerbe P, Chiang A, Yuen M, Hamberger B, Draper JA, Britton R, Bohlmann J. J Biol Chem. 2012;287:12121. doi: 10.1074/jbc.M111.317669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mafu S, Hillwig ML, Peters RJ. Chem Bio Chem. 2011;12:1984. doi: 10.1002/cbic.201100336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kolesnikova MD, Obermeyer AC, Wilson WK, Lynch DA, Xiong Q, Matsuda SP. Org Lett. 2007;9:2183. doi: 10.1021/ol070709b. [DOI] [PubMed] [Google Scholar]
- 18.Nakano C, Okamura T, Sato T, Dairi T, Hoshino T. Chem Commun (Camb) 2005;2005:1016. doi: 10.1039/b415346d. [DOI] [PubMed] [Google Scholar]
- 19.Hamano Y, Kuzuyama Y, Itoh N, Furihata K, Seto H, Dairi T. J Biol Chem. 2002;277:37098. doi: 10.1074/jbc.M206382200. [DOI] [PubMed] [Google Scholar]
- 20.Xu M, Wilderman PR, Peters RJ. Proc Natl Acad Sci USA. 2007;104:7397. doi: 10.1073/pnas.0611454104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wilderman PR, Peters RJ. J Am Chem Soc. 2007;129:15736. doi: 10.1021/ja074977g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Morrone D, Xu M, Fulton DB, Determan MK, Peters RJ. J Am Chem Soc. 2008;130:5400. doi: 10.1021/ja710524w. [DOI] [PubMed] [Google Scholar]
- 23.Keeling CI, Weisshaar S, Lin RPC, Bohlmann J. Proc Natl Acad Sci USA. 2008;105:1085. doi: 10.1073/pnas.0709466105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhou K, Peters RJ. Chem Comm. 2011;47:4074. doi: 10.1039/c0cc02960b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Keeling CI, Madilao LL, Zerbe P, Dullat HK, Bohlmann J. J Biol Chem. 2011;286:21145. doi: 10.1074/jbc.M111.245951. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.