

ABSTRACT
A new human coronavirus (hCoV-EMC) has emerged very recently in the Middle East. The clinical presentation resembled that of the severe acute respiratory syndrome (SARS) as encountered during the epidemic in 2002/2003. In both cases, acute renal failure was observed in humans. HCoV-EMC is a member of the same virus genus as SARS-CoV but constitutes a sister species. Here we investigated whether it might utilize angiotensin-converting enzyme 2 (ACE2), the SARS-CoV receptor. Knowledge of the receptor is highly critical because the restriction of the SARS receptor to deep compartments of the human respiratory tract limited the spread of SARS. In baby hamster kidney (BHK) cells, lentiviral transduction of human ACE2 (hACE2) conferred permissiveness and replication for SARS-CoV but not for hCoV-EMC. Monkey and human kidney cells (LLC-MK2, Vero, and 769-P) and swine kidney cells were permissive for both viruses, but only SARS-CoV infection could be blocked by anti-hACE2 antibody and could be neutralized by preincubation of virus with soluble ACE2. Our data show that ACE2 is neither necessary nor sufficient for hCoV-EMC replication. Moreover, hCoV-EMC, but not SARS-CoV, replicated in cell lines from Rousettus, Rhinolophus, Pipistrellus, Myotis, and Carollia bats, representing four major chiropteran families from both suborders. As human CoV normally cannot replicate in bat cells from different families, this suggests that hCoV-EMC might use a receptor molecule that is conserved in bats, pigs, and humans, implicating a low barrier against cross-host transmission.
IMPORTANCE
A new human coronavirus (hCoV) emerged recently in the Middle East. The disease resembled SARS (severe acute respiratory syndrome), causing a fatal epidemic in 2002/2003. Coronaviruses have a reservoir in bats and because this novel virus is related to SARS-CoV, we investigated whether it might replicate in bat cells and use the same receptor (angiotensin-converting enzyme 2 [ACE2]). This knowledge is highly critical, because the SARS-CoV receptor influenced pathology, and its localization in the deep respiratory tract is thought to have restricted the transmissibility of SARS. Our data show that hCoV-EMC does not need the SARS-CoV receptor to infect human cells. Moreover, the virus is capable of infecting human, pig, and bat cells. This is remarkable, as human CoVs normally cannot replicate in bat cells as a consequence of host adaptation. Our results implicate that the new virus might use a receptor that is conserved between bats, pigs and humans suggesting a low barrier against cross-host transmission.
Observation
Coronaviruses (CoVs) are large enveloped plus-strand RNA viruses. Over the last decades, several animal CoVs have been identified, including major pathogens such as the bovine CoV and the avian infectious bronchitis virus. Two human CoVs (hCoVs) termed hCoV-OC43 and hCoV-229E have been known since the 1960s, causing mild respiratory disease (1). In 2002/2003, an outbreak of severe acute respiratory syndrome (SARS) leading to over 700 deaths was caused by a novel hCoV termed SARS-CoV (2, 3). In the aftermath of the SARS epidemic, two additional hCoVs termed hCoV-NL63 and -HKU1 as well as numerous novel bat-associated CoVs were described (4–7). CoVs have been reclassified into four genera designated Alphacoronavirus (former groups 1a and 1b), Betacoronavirus (groups 2a to 2d), Gammacoronavirus (group 3), and as recently suggested, Deltacoronavirus (8). A likely animal reservoir for SARS-CoV has been identified in bats (7, 9–11). Nevertheless, bat-derived CoVs have never been isolated in cell culture. Prototypic CoVs, including the SARS-CoV, are almost unculturable in bat-derived cell lines (11, 12). Double infections of bats with different CoVs are very rare (13), and bat CoVs appear to be restricted to their bat hosts at the genus level (14, 15). These points taken together suggest that there is a tight barrier against CoV host switching, preventing humans from acquiring novel CoVs easily (1, 11).
The SARS-CoV utilizes the angiotensin-converting enzyme 2 (ACE2) as its necessary and sufficient cellular receptor (11, 16). The cellular and anatomic localization of ACE2 in the human body had an important meaning for the pathogenesis and epidemiology of SARS. The receptor is expressed on pneumocytes deep in the human lung, but hardly in the upper parts of the respiratory tract (17). The direct targeting of pneumocytes may explain why SARS-CoV infection led to a severe clinical picture with early onset of respiratory distress in affected patients. The focusing of SARS-CoV replication to the deep respiratory tract also determined a major hallmark of SARS epidemiology. Because the virus needed to reach the deep respiratory tract to seed an infection, the infectious dose was rather high. Once infected, primary replication in the deep lung rapidly caused severe clinical symptoms leading to early hospitalization and thus, isolation, before patients could develop high virus concentrations in respiratory secretions. These factors taken together are thought to have limited the spread of SARS-CoV in the human population.
In September 2012, health authorities were notified of cases of severe hCoV infection caused by a novel virus type in the Middle East region (18, 19). The disease involved severe acute respiratory failure but as an additional component affected the kidneys, causing acute renal failure (18, 19). A novel betacoronavirus was isolated from one of two patients in Jeddah, Saudi Arabia, and termed hCoV-EMC (also named hCoV-EMC/2012) (10, 20). Another case originating from Qatar was transferred to London, United Kingdom, for treatment (18, 19). Very recently, a third case was reported from Riyadh, Saudi Arabia, suggesting that more cases are to be expected (21). Availability of a virus isolate enables rapid provisional assessments of virus properties in order to guess at virus origins and project epidemic risks. Because of the taxonomic placement of hCoV-EMC and SARS-CoV as sister clades within the same virus genus, we were interested in rapidly assessing any possible overlaps in receptor usage (18, 20).
For the purpose of receptor usage studies, we have established a baby hamster kidney (BHK) cell line carrying human ACE2 (hACE2) following lentiviral transduction. BHK cells normally do not express ACE2 and are not permissive for SARS-CoV. As shown in Fig. 1A, the transduced cells termed BHK-hACE2 expressed hACE2 as demonstrable by antigen staining using an immunofluorescence assay (IFA). Surface expression of hACE2 was detected by fluorescence-activated cell sorting (FACS). SARS-CoV applied at a multiplicity of infection (MOI) of 0.5 replicated more than 10,000-fold within 40 h in BHK-hACE2 but not in the parental BHK cell line (Fig. 1B). Infection with hCoV-EMC under the same conditions showed no replication irrespective of the presence of hACE2. We concluded that hACE2 was sufficient to permit infection by SARS-CoV, but not hCoV-EMC, in BHK cells.
FIG 1.

Human ACE2-independent entry of hCoV-EMC. (A) Baby hamster kidney (BHK) cells were transduced with hACE2-carrying lentiviruses and selected by puromycin treatment. (Left) hACE2 expression was controlled by immunofluorescence assay (IFA) by using goat anti-hACE2 immunoglobulin (Ig) (R&D Systems) followed by cyanin 3-labeled donkey anti-goat Ig (Dianova). For controls, ACE2-expressing primate cell lines (kidney cells from rhesus monkey [LLC-MK2] and African green monkey [MA104]) were stained in parallel. DAPI, 4',6'-diamidino-2-phenylindole. Bar, 50 µm. (Right) ACE2 protein surface expression was determined by FACS analysis using a mouse anti-hACE2 Ig antibody (R&D Systems) in combination with an Alexa Fluor 647-labeled goat anti-mouse Ig (Life Technologies). Data were acquired using FACS Canto and analyzed with FlowJo software. The cells were incubated with the secondary antibody only (shown in gray) for a control. mAB, monoclonal antibody. (B) The interferon-deficient BHK cells and the generated hACE2-expressing transgenic BHK cells were applied in infection studies. LLC-MK2 and MA104 primate cells known to be susceptible for SARS-CoV were infected in parallel with SARS-CoV strain Frankfurt-1 and hCoV-EMC. The cells were infected with an MOI of 0.5 for 1 h, washed twice with phosphate-buffered saline (PBS), and supplemented with medium. At time points 0 and 40 h after infection, samples from supernatants were taken in order to extract viral RNA. Real-time reverse transcription-PCR (RT-PCR) was used for absolute quantification of genome equivalents (GE) per ml of supernatant (2, 10). All experiments were performed in triplicate. hpi, hours postinfection. (C) In order to block the ACE2 receptor, Vero cells were preincubated with 10 µg/ml of polyclonal goat anti-hACE2 for 1 h at 37°C. Half of the antibody solution was stored, and cells were subsequently infected by adding viruses at an MOI of 0.01 for 30 min at 4°C. The supernatant was discarded, and medium with antibody solution was added. At time points 0 and 20 h postinfection, samples from supernatants (triplicate samples) were taken for quantification by real-time RT-PCR. pAb, polyclonal antibody. (D) Virus-neutralizing activity of soluble recombinant hACE2 (rhACE2) was tested by preincubating in triplicate different concentrations of rhACE2 (a kind gift from J. M. Penninger, Vienna, Austria) with 104 50% tissue culture infective dose (TCID50)/ml hCoV-EMC (triangles) or 103 TCID50/ml SARS-CoV (squares) for 1 h. The mixture was added to Vero E6 cells for 1 h, after which cells were washed and fresh medium was added. The infection was stopped after 8 h, and cells were visualized with a cross-reactive SARS-CoV serum followed by a goat anti-rabbit IgG antibody conjugated to peroxidase and stained with 3-amino-9-ethylcarbazole (AEC) substrate.
To exclude technical confounders due to transgenic expression of hACE2, infection studies were repeated in kidney cells naturally permissive for SARS-CoV. Next to Vero cells in which the original isolate of hCoV-EMC had been obtained, LLC-MK2 and MA104 cells are commonly used monkey kidney cell lines known to be permissive for SARS-CoV (22). As shown in Fig. 1A, LLC-MK2 and MA104 cells expressed ACE2 as demonstrated by IFA and FACS. SARS-CoV infected cells under conditions as described above replicated more than 1,000-fold within 40 h in both cell lines. In contrast, while hCoV-EMC showed strong replication in LLC-MK2 cells, it did not replicate at all in MA104 cells.
While these findings confirmed that ACE2 did not suffice for hCoV-EMC permissibility, it might still be necessary in addition to another receptor. To clarify its usage by hCoV-EMC in primate cells like LLC-MK2 and Vero cells, a polyclonal goat serum directed against hACE2 was used for receptor-blocking studies. As shown in Fig. 1C, infection of Vero cells by SARS-CoV, but not hCoV-EMC, was completely blocked by the antibody. The blocking of SARS-CoV infection occurred in a dose-dependent manner (see Fig. S1 in the supplemental material).
To exclude the remaining possibility that our antibody blocked an ACE2 epitope relevant for binding of SARS-CoV but irrelevant for hCoV-EMC, infection neutralization assays were carried out using soluble ACE2 protein in Vero cells. As shown in Fig. 1D, infection with SARS-CoV, but not with hCoV-EMC, could be competed in a dose-dependent manner by the presence of soluble ACE2. Overall, these observations led us to conclude that SARS-CoV, but not hCoV-EMC, utilizes hACE2 as a receptor for virus entry.
While it is known that SARS-related CoVs occur widely in bats, the human SARS-CoV is adapted to the human ACE2 receptor and has lost its ability to infect bat cells (11, 12). For reference, we generated kidney cells from Rousettus aegyptiacus bats, termed RoNi/7. As shown in Fig. 2A, lentiviral transduction and selection yielded a variant of these cells, termed RoNi/7-hACE2, which expressed hACE2. Infection of these cells with SARS-CoV resulted in high levels of virus replication, while the parental RoNi/7 cells were not susceptible (Fig. 2B; see Fig. S2 in the supplemental material). Surprisingly, these same parental RoNi/7 cells were highly permissive for hCoV-EMC. The derived RoNi/7-hACE2 cells appeared slightly more permissive for hCoV-EMC, but by performing antibody competition experiments, the possibility that this difference was due to the presence of hACE2 in RoNi/7-hACE2 cells was excluded, as SARS-CoV infection was blocked by anti-hACE2 antibody, and hCoV-EMC infection was not (Fig. 2C).
FIG 2.
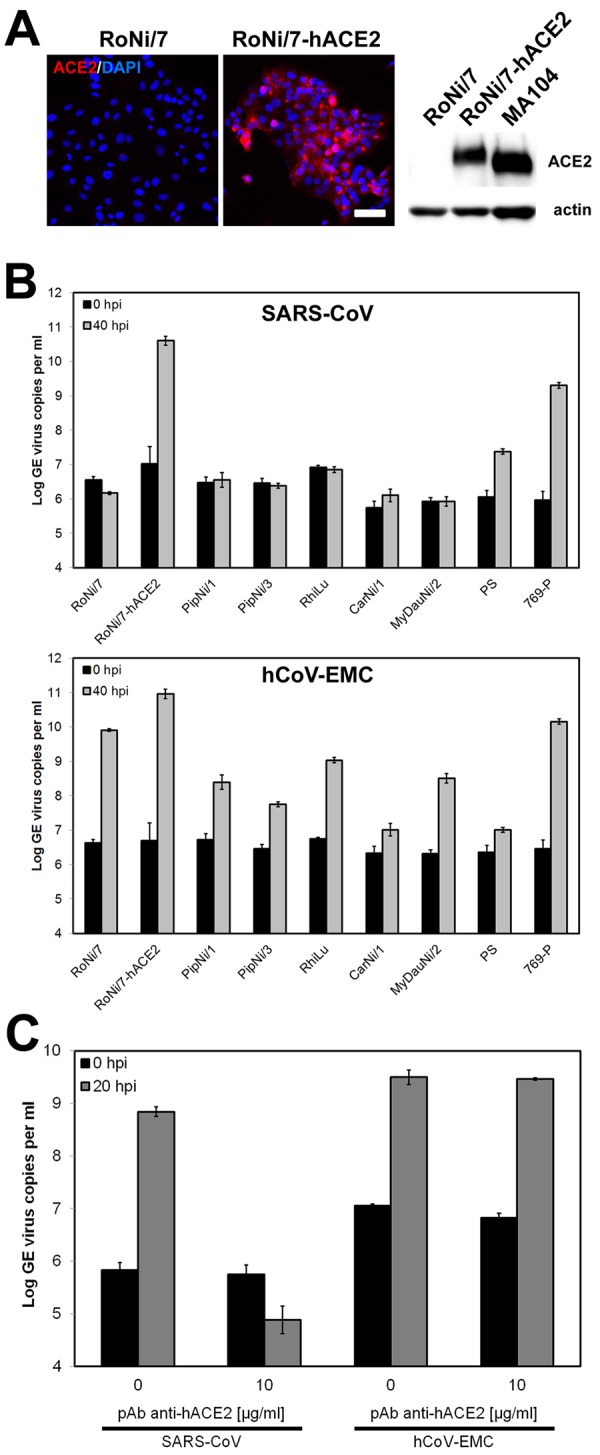
Bat cells allow ACE2-independent entry of hCoV-EMC. (A) Immortalized Rousettus aegyptiacus cells were provided with hACE2. (Left) Expression of hACE2 was controlled by immunofluorescence assay using goat anti-hACE2 Ig and cyanin 3-labeled donkey anti-goat Ig. Bar, 50 µm. (Right) Western blot analysis was done using goat anti-hACE2 Ig followed by horseradish peroxidase-labeled donkey anti-goat Ig (Dianova). For a control, a protein lysate of MA104 cells was applied to the blot. A loading control was done with a rabbit anti-pan-actin Ig (Cell Signaling). (B) Representative bat cell cultures from Yinpterochiroptera (Rousettus aegyptiacus kidney [RoNi/7] and Rhinolophus landeri lung [RhiLu]) and Yangochiroptera (Pipistrellus pipistrellus kidney [PipNi/1 and PipNi/3], Carollia perspicillata kidney [CarNi/1], and Myotis daubentonii kidney [MyDauNi/2]) and porcine (PS) and human kidney (769-P) cell lines were used for infection studies with SARS-CoV (top panel) and hCoV-EMC (bottom panel). For a control and for comparison, hACE2-expressing transgenic Rousettus bat cell cultures (RoNi/7-hACE2) were included. The cells were infected at an MOI of 0.5 for 1 h, washed twice with PBS, and supplemented with medium. At time points 0 and 40 h postinfection, samples from supernatants were taken to extract viral RNA. Real-time RT-PCR was performed as described for absolute quantification of GE. All experiments were performed in triplicate. (C) A hACE2-blocking experiment was done with RoNi/7-hACE2 cells. The cells were preincubated with 10 µg/ml of polyclonal goat anti-hACE2 for 1 h at 37°C. The cells were subsequently infected by adding viruses at an MOI of 0.01 for 30 min at 4°C. The supernatant was discarded, and medium supplemented with hACE2 antibody was added. At time points 0 and 20 h postinfection, samples from supernatants were taken to quantify GE by real-time RT-PCR. The experiment was performed in triplicate.
The ability of hCoV-EMC to grow in bat cells was highly remarkable, as most other prototype coronaviruses, including the human viruses and bovine, feline, or porcine CoVs, are normally unable to grow in bat cells (9, 12; our unpublished observations). Because hCoV-EMC was most closely related to bat CoV HKU5 detected in bats of the genus Pipistrellus (18, 20, 23), we generated two different kidney cell lines from the European bat species Pipistrellus pipistrellus as well as a kidney cell line from another European vespertilionid bat species, Myotis daubentonii (MyDauNi/2). A New World bat cell line from the species Carollia perspicillata (CarNi/1) was made, and a permanent cell line from Rhinolophus landeri lung cells was included to represent the bat genus associated with SARS-related CoV. Porcine kidney cells (PS) were tested to represent a larger range of laurasiatherian mammals, and a human kidney cancer cell line (769-P) was included to complement the tested range of primate cells. As shown in Fig. 2B, all of the bat cell lines were readily infected by hCoV-EMC (lower panel) but not by SARS-CoV (upper panel). Porcine and human kidney cell lines were infected by both coronaviruses (Fig. 2B). In summary, hCoV-EMC was able to infect cells from four major families of bats covering the whole diversity of this large mammalian order (Pteropodidae [Rousettus], Rhinolophidae [Rhinolophus], Vespertilionidae [Pipistrellus, Myotis], Phyllostomidae [Carollia]). Replication in porcine and primate cells suggests an even broader tropism across mammals.
Large parts of the epidemiology and pathology of SARS were related to its receptor tropism in humans (3, 11, 16). In a scenario resembling that for avian influenza viruses, the anatomical position of the receptor in the deep respiratory tract and the downregulation of its expression were postulated to aggravate the course of disease in individuals while limiting the spread of disease on the population level (24). While we are still uncertain of the epidemiology of hCoV-EMC-related infections, the importance of knowing its receptor in human cells is unquestionable (11). Our data taken together suggest that hCoV-EMC does not rely on the same receptor as SARS-CoV. Urgent further research is needed in order to identify the cellular receptor for hCoV-EMC and determine its anatomical focus of replication. Our observations regarding the cell culture tropism of this novel human virus raise an intriguing perspective. We have shown here that cells from primates, pigs, as well as bats representing four families from both chiropteran suborders, Yangochiroptera and Yinpterochiroptera, retain susceptibility for the virus. This breadth of tropism is absolutely unique among CoVs (1, 11, 12). For instance, only once have researchers succeeded in culturing any CoV in bat-derived cells, and these experiments required specifically generated bat cell cultures combined with engineered, reporter gene-expressing virus (12). The broad replicative capability of hCoV-EMC suggests that this new virus might utilize a receptor structure that bats, primates, and pigs have in common. If that receptor were expressed in mucosal surfaces in those hosts, repeated acquisition by humans would be conceivable, putting our current idea of a “tight” molecular barrier against coronaviral cross-host transmission into perspective (11, 12). Interestingly, these data provide strong support for the existence of “generalist” CoV, as recently projected based on cell culture studies (12).
SUPPLEMENTAL MATERIAL
Dose-dependent inhibition of SARS-CoV growth by polyclonal anti-hACE2 immunoglobulin. Primate cells (LLC-MK2) were preincubated with different concentrations of polyclonal goat-anti hACE2 immunoglobulin for 1 h at 37°C. The cells were washed once with PBS and subsequently infected with SARS-CoV Frankfurt-1 at an MOI of 0.5 for 1 h, washed twice with PBS, and supplemented with medium. At time points 0 and 40 h postinfection, samples from supernatants were taken in order to extract viral RNA and perform real-time RT-PCR. The percent growth at 40 h postinfection in one representative experiment is shown. All 40-h values were calibrated by subtracting respective time point 0-h values. The negative control was set at 100%. All experiments were performed twice in triplicate samples. Download Figure S1, TIF file, 0.1 MB.
Plaque titration of supernatants of infected bat cell cultures. Kidney bat cell cultures generated from Rousettus aegyptiacus (RoNi/7 and RoNi/7-hACE2 carrying the SARS-CoV receptor hACE2), Pipistrellus pipistrellus (PipNi/1 and PipNi/3), and a Rhinolophus landeri lung cell culture (RhiLu) were infected in triplicate with SARS-CoV and hCoV-EMC at an MOI of 0.5. After 40 h postinfection, supernatants were harvested and titrated in serial dilutions (duplicates) on Vero cells. nd, not detected. Download Figure S2, TIF file, 0.2 MB.
ACKNOWLEDGMENTS
This study was supported by the European Union FP7 project EMPERIE (contract number 223498), ANTIGONE (contract number 278976), the German Research Foundation (DFG grants DR 772/3-1 and KA1241/18-1), as well as the German Ministry of Education and Research (BMBF SARS II).
We are grateful to Florian Gloza-Rausch (Noctalis, Bad Segeberg, Germany), Matthias Habetha, Artem Siemens, and Andrea Rasche (Institute of Virology, Bonn, Germany), and Kim Schmidt (Institute of Medical Microbiology, Immunology and Parasitology) and Elmar Endl (FACS core facility of the University of Bonn Medical Center) for technical assistance. We also thank Frank Westermann (German Cancer Research Center, Heidelberg, Germany) for providing 769-P cells.
Footnotes
Citation Müller MA, et al. 2012. Human coronavirus EMC does not require the SARS-coronavirus receptor and maintains broad replicative capability in mammalian cell lines. mBio 3(6):e00515-12. doi:10.1128/mBio.00515-12.
REFERENCES
- 1. Saif LJ. 2004. Animal coronaviruses: what can they teach us about the severe acute respiratory syndrome? Rev. Sci. Tech. 23:643–660 [DOI] [PubMed] [Google Scholar]
- 2. Drosten C, et al. 2003. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 348:1967–1976 [DOI] [PubMed] [Google Scholar]
- 3. Peiris JS, Yuen KY, Osterhaus AD, Stöhr K. 2003. The severe acute respiratory syndrome. N. Engl. J. Med. 349:2431–2441 [DOI] [PubMed] [Google Scholar]
- 4. Fouchier RA, et al. 2004. A previously undescribed coronavirus associated with respiratory disease in humans. Proc. Natl. Acad. Sci. U. S. A. 101:6212–6216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. van der Hoek L, et al. 2004. Identification of a new human coronavirus. Nat. Med. 10:368–373 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Woo PC, et al. 2005. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J. Virol. 79:884–895 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Woo PC, Lau SK, Huang Y, Yuen KY. 2009. Coronavirus diversity, phylogeny and interspecies jumping. Exp. Biol. Med. (Maywood) 234:1117–1127 [DOI] [PubMed] [Google Scholar]
- 8. Woo PC, et al. 2012. Discovery of seven novel mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J. Virol. 86:3995–4008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Biesold SE, et al. 2011. Type I interferon reaction to viral infection in interferon-competent, immortalized cell lines from the African fruit bat Eidolon helvum. PLoS One 6:e28131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Corman V, et al. 2012. Detection of a novel human coronavirus by real-time reverse-transcription polymerase chain reaction. Euro Surveill. 17(39):pii =20285 [DOI] [PubMed] [Google Scholar]
- 11. Graham RL, Baric RS. 2010. Recombination, reservoirs, and the modular spike: mechanisms of coronavirus cross-species transmission. J. Virol. 84:3134–3146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Huynh J, et al. 2012. Evidence supporting a zoonotic origin of human coronavirus strain NL63. J. Virol. 86:12816–12825 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Lau SK, et al. 2010. Coexistence of different genotypes in the same bat and serological characterization of Rousettus bat coronavirus HKU9 belonging to a novel Betacoronavirus subgroup. J. Virol. 84:11385–11394 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 14. Drexler JF, et al. 2010. Genomic characterization of severe acute respiratory syndrome-related coronavirus in European bats and classification of coronaviruses based on partial RNA-dependent RNA polymerase gene sequences. J. Virol. 84:11336–11349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Gloza-Rausch F, et al. 2008. Detection and prevalence patterns of group I coronaviruses in bats, northern Germany. Emerg. Infect. Dis. 14:626–631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Li W, et al. 2003. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426:450–454 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Bertram S, et al. 2012. Influenza and SARS-coronavirus activating proteases TMPRSS2 and HAT are expressed at multiple sites in human respiratory and gastrointestinal tracts. PLoS One 7:e35876 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Bermingham A, et al. 2012. Severe respiratory illness caused by a novel coronavirus, in a patient transferred to the United Kingdom from the Middle East. Euro Surveill. 17(40):pii =20290 [PubMed] [Google Scholar]
- 19. Pebody RG, et al. 2012. The United Kingdom public health response to an imported laboratory confirmed case of a novel coronavirus in September 2012. Euro Surveill. 17(40):pii =20292 [PubMed] [Google Scholar]
- 20. Zaki AM, van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA. 2012. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 367:1814–1820 [DOI] [PubMed] [Google Scholar]
- 21. Memish Z. 2012. A new Saudi novel coronavirus case diagnosed in KSA (Kingdom of Saudi Arabia). Archive no. 20121104.1391285, Program for Monitoring Emerging Diseases; (ProMedMail: ). International Society for Infectious Diseases, Brookline, MA [Google Scholar]
- 22. Kaye M. 2006. SARS-associated coronavirus replication in cell lines. Emerg. Infect. Dis. 12:128–133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Woo PC, et al. 2006. Molecular diversity of coronaviruses in bats. Virology 351:180–187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Kuba K, et al. 2005. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 11:875–879 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Dose-dependent inhibition of SARS-CoV growth by polyclonal anti-hACE2 immunoglobulin. Primate cells (LLC-MK2) were preincubated with different concentrations of polyclonal goat-anti hACE2 immunoglobulin for 1 h at 37°C. The cells were washed once with PBS and subsequently infected with SARS-CoV Frankfurt-1 at an MOI of 0.5 for 1 h, washed twice with PBS, and supplemented with medium. At time points 0 and 40 h postinfection, samples from supernatants were taken in order to extract viral RNA and perform real-time RT-PCR. The percent growth at 40 h postinfection in one representative experiment is shown. All 40-h values were calibrated by subtracting respective time point 0-h values. The negative control was set at 100%. All experiments were performed twice in triplicate samples. Download Figure S1, TIF file, 0.1 MB.
Plaque titration of supernatants of infected bat cell cultures. Kidney bat cell cultures generated from Rousettus aegyptiacus (RoNi/7 and RoNi/7-hACE2 carrying the SARS-CoV receptor hACE2), Pipistrellus pipistrellus (PipNi/1 and PipNi/3), and a Rhinolophus landeri lung cell culture (RhiLu) were infected in triplicate with SARS-CoV and hCoV-EMC at an MOI of 0.5. After 40 h postinfection, supernatants were harvested and titrated in serial dilutions (duplicates) on Vero cells. nd, not detected. Download Figure S2, TIF file, 0.2 MB.