

Abstract
AIM: To investigate the effect of moxibustion on intestinal flora and release of interleukin-12 (IL-12) and tumor necrosis factor-α (TNF-α) from the colon in rat with ulcerative colitis (UC).
METHODS: A rat model of UC was established by local stimulation of the intestine with supernatant from colonic contents harvested from human UC patients. A total of 40 male Sprague-Dawley rats were randomly divided into the following groups: normal (sham), model (UC), herb-partition moxibustion (HPM-treated), and positive control sulfasalazine (SA-treated). Rats treated with HPM received HPM at acupuncture points ST25 and RN6, once a day for 15 min, for a total of 8 d. Rats in the SA group were perfused with SA twice a day for 8 d. The colonic histopathology was observed by hematoxylin-eosin. The levels of intestinal flora, including Bifidobacterium, Lactobacillus, Escherichia coli (E. coli), and Bacteroides fragilis (B. fragilis), were tested by real-time quantitative polymerase chain reaction to detect bacterial 16S rRNA/DNA in order to determine DNA copy numbers of each specific species. Immunohistochemical assays were used to observe the expression of TNF-α and IL-12 in the rat colons.
RESULTS: HPM treatment inhibited immunopathology in colonic tissues of UC rats; the general morphological score and the immunopathological score were significantly decreased in the HPM and SA groups compared with the model group [3.5 (2.0-4.0), 3.0 (1.5-3.5) vs 6.0 (5.5-7.0), P < 0.05 for the general morphological score, and 3.00 (2.00-3.50), 3.00 (2.50-3.50) vs 5.00 (4.50-5.50), P < 0.01 for the immunopathological score]. As measured by DNA copy number, we found that Bifidobacterium and Lactobacillus, which are associated with a healthy colon, were significantly higher in the HPM and SA groups than in the model group (1.395 ± 1.339, 1.461 ± 1.152 vs 0.045 ± 0.036, P < 0.01 for Bifidobacterium, and 0.395 ± 0.325, 0.851 ± 0.651 vs 0.0015 ± 0.0014, P < 0.01 for Lactobacillus). On the other hand, E. coli and B. fragilis, which are associated with an inflamed colon, were significantly lower in the HPM and SA groups than in the model group (0.244 ± 0.107, 0.628 ± 0.257 vs 1.691 ± 0.683, P < 0.01 for E. coli, and 0.351 ± 0.181, 0.416 ± 0.329 vs 1.285 ± 1.039, P < 0.01 for B. fragilis). The expression of TNF-α and IL-12 was decreased after HPM and SA treatment as compared to UC model alone (4970.81 ± 959.78, 6635.45 ± 1135.16 vs 12333.81 ± 680.79, P < 0.01 for TNF-α, and 5528.75 ± 1245.72, 7477.38 ± 1259.16 vs 12550.29 ± 1973.30, P < 0.01 for IL-12).
CONCLUSION: HPM treatment can regulate intestinal flora and inhibit the expression of TNF-α and IL-12 in the colon tissues of UC rats, indicating that HPM can improve colonic immune response.
Keywords: Ulcerative colitis, Herb-partition moxibustion, Intestinal flora, Immune regulation
INTRODUCTION
Ulcerative colitis (UC) is a chronic inflammatory disease of the gastrointestinal tract characterized by increased stool frequency, bleeding and abdominal pain, and ulceration limited to the colon mucosa. The role of immunological pathogenesis in UC has been widely recognized[1,2]. In recent years, the involvement of pathogen- mediated immune dysfunction has received extensive attention. Studies have shown that UC patients have altered intestinal flora species. For example, Bifidobacterium, Lactobacillus and fusobacterium Escherichia coli (E. coli) desulfovibrio were significantly decreased, while Bacteroides fragilis (B. fragilis), were significantly increased[3-6]. In hosts with altered intestinal flora, the immune system recognizes intestinal flora and its metabolites as pathogenic antigens that can cause an abnormal immune response and stimulate bowel epithelial cells in UC patients with genetic susceptibility. This activity then destroys the structure and function of colonic mucosa, leading to the ongoing pathology associated with UC[7]. Alteration of the intestinal flora has been considered to be the initiating and continuous factor in the onset of UC[7-10]. Therefore, the modulation of intestinal flora has been suggested as a potential approach to treat UC.
Previous studies have shown that herb-partition moxibustion (HPM) can be anti-inflammatory[11]. It can also improve symptoms in mild-to-moderate UC patients, such as relieving abdominal pain and decreasing bloody diarrhea[12-14]. Experiments have also demonstrated that HPM can down-regulate the protein and mRNA expression of interleukin (IL)-8 and intercellular adhesion molecule-1 in colon tissues of UC patients, indicating that HPM can inhibit chemotaxis and cell migration into inflamed tissues[14]. HPM can also promote neutrophil apoptosis and down-regulate cytokines, such as IL-1 beta, IL-6, and tumor necrosis factor-alpha (TNF-α), further suggesting an anti-inflammatory role for HPM treatment[15]. Moreover, a recent report shows that intestinal injury was decreased and TNF-α was also found to be decreased after HPM treatment in a Crohn’s disease model[16]. Taken together, these results indicate that HPM can regulate immune function in UC. However, whether HPM can also alter the intestinal flora composition is still unknown.
A large number of bacteria comprise the human intestinal flora, outnumbering host cells by a ratio of 10:1. Various types of bacteria constitute the gut flora, including symbiotic and pathogenic bacteria whose pathogenicity is hidden from the immune system from various barrier and immune-mediated mechanisms. For this study, we observed the relative proportions of both the symbiotic - and most prevalent - bacteria, such as bifidobacteria and lactobacilli, and the conditioned pathogenic flora, such as E. coli and B. fragilis, as readouts to explore the effects of HPM on modulating the intestinal flora of rats in a model of UC. We also investigated whether HPM regulates the expression of pro-inflammatory cytokines, such as TNF-α and IL-12, in colon tissues of UC rats. This study would aid in understanding the mechanisms of how HPM can alleviate symptoms of UC.
MATERIALS AND METHODS
Animals
Sprague-Dawley rats were obtained from the Department of Experimental Animal Science of Shanghai Medical College at Fudan University [No. SCXK(SH)2009-0019]. All protocols were performed in strict accordance with the Guidance Suggestions for the Care and Use of Laboratory Animals, formulated by the Ministry of Science and Technology of the People’s Republic of China[17]. This study received permission from the Ethics Committee in Yueyang Hospital of Integrated Traditional Chinese and Western Medicine, which is affiliated with Shanghai University of Traditional Chinese Medicine, China.
Establishment of the UC rat model
Forty neonatal, male Sprague Dawley rats weighing 183 ± 10 g were randomly divided into the following experimental groups: normal, model, HPM, and sulfasalazine (SA). The UC rat model was established according to an immunological method and local stimulation[15,18-20]. In brief, fresh human colonic mucosa was obtained from surgical colonic specimens, homogenized in normal saline and centrifuged for 30 min at 3000 rpm in order to remove cells and bacteria. The supernatant, containing antigens released from UC colon patients, was diluted into an appropriate protein concentration and mixed with Freund’s adjuvant (Shanghai Chemical Reagent Company, Shanghai, China). One mL of the antigen plus adjuvant mixture, containing a total of 6 mg protein, was injected into the front footpad at day 0. Following this initial dose, the same mixture was then injected into the back footpad, dorsa, inguina, and abdominal cavities on days 10, 17, 24 and 31, respectively. On day 38, rats were anesthetized intraperitoneally with 2% pentobarbital sodium (30 mg/kg) and 3 mL enema of 3% formalin were administered to the rats - in order to stimulate the colon immune response - for 1 h following a saline wash after a 2 mL enema of antigen fluid (without Freund’s adjuvant) for 2 h following a saline wash.
Treatment
After the UC model was established in the rats, HPM was performed during days 39-46 of UC model in the treatment groups. Moxa cones (0.5 cm in diameter and 0.6 cm high) (Nanyang Hanyi Moxa Co., Ltd. China) were placed vertically on a medicinal formula composed of radix aconite, cortex, radix, carthami and salviae miltiorrhizae. The medicinal formula was then placed on acupoints ST25 (located on a horizontal line 2 cun laterally to the midline and 5 cun above the symphysis pubis, a point of hand Yangming, the mu point of the large intestine meridian, and it regulates the function of the intestine and stomach)[21,22] and RN6 (located on ventral midline 3.5 cun above the symphysis pubis and 1.5 cun below the navel, a point of the conception meridian, and it strengthens original qi and improves immune function); the moxa cones were then ignited and each acupoint was treated twice in 15 min increments. This treatment was repeated once daily for a total of 8 d[23]. As a positive control for inhibition of UC symptoms, the immunosuppressant SA was used. For the SA treatment group, salicylazosulfapyridine solution (0.25 g/tablet) was intragastrically administered to rats (Sanwei Pharmaceutical, Shanghai, China; Batch No. 200807C30) twice daily for a total of 8 d. The salicylazosulfapyridine solution concentration was 20 mg/mL with a daily dose of 100 mg/kg, which is equivalent to 0.1 g/kg in a human patient[24].
Morphological observation of fixed colon samples
Following treatment and sacrifice by cervical dislocation, samples were collected from the descending colon (5 cm above the anus), cleaned with normal saline, and general morphology was then scored (Table 1). The samples were fixed in 10% formalin, dehydrated, embedded in paraffin, and sectioned into 4 μm thick slices. These sections were then stained by hematoxylin-eosin for pathological observation and the histological grade was scored with the bland method[25,26] (Table 2).
Table 1.
General morphological scoring
| General morphological manifestation | Score | |
| Colon adhesion | No adhesion | 0 |
| Mild adhesion | 1 | |
| Severe adhesion | 2 | |
| Ulcer and | No ulcer and no inflammation | 0 |
| inflammation | Local congestion, no ulcer | 1 |
| 1 ulcer without congestion or bowel wall thickening | 2 | |
| 1 ulcer with inflammation | 3 | |
| > 2 ulcer and inflammation | 4 | |
| > 2 ulcer and/or inflammation area > 1 cm | 5 | |
| > 2 ulcer and/or inflammation area > 2 cm, one more damage, plus 1 | 6-8 | |
Table 2.
Immunopathological scoring
| Immunopathological manifestation | Score | |
| Ulcer | No ulcer | 0 |
| Ulcer area < 3 cm | 1 | |
| Ulcer area > 3 cm | 2 | |
| Inflammation | No inflammation | 0 |
| Mild inflammation | 1 | |
| Severe inflammation | 2 | |
| Granuloma | No granuloma | 0 |
| Granuloma | 1 | |
| Lesion depth | No lesion | 0 |
| Submucosa | 1 | |
| Muscular layer | 2 | |
| Serosa layer | 3 | |
| Fibrosis | No fibrosis | 0 |
| Mild fibrosis | 1 | |
| Severe fibrosis | 2 | |
TNF-α and IL-12 immunohistochemistry
Paraformaldehyde-fixed and paraffin-embedded samples were sliced into 4 μm slices. Paraffin slides were deparaffinized in xylene I, xylene II, and xylene III; dehydrated in 95%, 90% and 70% ethanol; and then incubated with primary rabbit antibodies at 4 °C for 18 h (anti-TNF-α was diluted to 1:150; IL-12 was diluted to 1:200) (Abcam Co., United Kingdom). The tissues were then visualized with 0.5 g/L diaminobenzidine and 0.3 g/L H2O2 in distilled water, and rinsed in phosphate buffered saline for 10 min. A known positive sample was used as a positive control for all slices, and phosphate buffered saline was substituted for the primary antibody and isotype controls as negative controls. All the samples were analyzed by a Motic Med 6.0 image analysis system (Motic Group Co., Ltd.). Three fields were randomly selected under an optical microscope (Olympus Co., Ltd.) at 400 × magnification to calculate the positive target value of the integral optical density.
16S rRNA real-time quantitative polymerase chain reaction
Rat feces taken directly from inoculated rats was placed into 5-10 mL of nutrient broth and was shaken and cultivated overnight at 37 °C. To extract DNA, 50 μL of the bacterial culture fluid was placed in a 1.5 ml sterile centrifuge tube, and 50 μL of DNA extract was added. This was mixed well at a constant temperature of 100°C for 10 min. It was then centrifuged at 12 000 rpm for 5 min and saved for later use. For the quantitative reverse transcription-polymerase chain reaction, the 7500 Sequence Detection System was used (ABI Co., USA). The sequence of the forward primer (F) and reverse primer (R) are shown in Table 3. DNA was prepared for PCR amplification in the following way: 10 μL 5× PCR buffer, 0.5 μL forward primer F, 0.5 μL reverse primer R, 0.5 μL dNTPs, 0.5 μL TaqMan fluorescent probe, 1 μL Taq enzyme, 32 μL dHO2 water, and 5 μL cDNA template. We used the following amplification conditions: (1) 50 °C 2 min, (2) 95 °C 5min, (3) 95 °C 15 s, and (4) 60 °C 45 s, for 40 cycles.
Table 3.
Oligonucleotide primers and TaqMan fluorescent probe sequences used for real-time polymerase chain reaction assays
| Primer or probe name | Primer sense | Sequence (5’→3’) | Amplification product (bp) |
| Bifidobacterium | Forward | 5'-ACTGGAATTCCCGGTGTAAC-3' | 85 |
| Reverse | 5'-GTCAGTAACGGCCCAGAGAC-3' | ||
| Lactococcus lactis | Forward | 5'-CAACATTTGGAAACGAATGC-3' | 134 |
| Reverse | 5'-CCTTGGTGAGCCTTTACCTC-3' | ||
| Bacteroides | Forward | 5'-ATTGCAGTGGAATGATGTGG-3' | 106 |
| Reverse | 5'-TATGGCACTTAAGCCGACAC-3' | ||
| Bacillus vallismortis | Forward | 5'-ACCGCATGGTTCAGACATAA-3' | 88 |
| Reverse | 5'-AGCCGTTACCTCACCAACT-3' |
Statistical analysis
All data were analyzed using SPSS 10.0 statistical software (SPSS Inc., United States), and all data were expressed as mean ± SD for normally distributed continuous variables and as median (QL-QU) for abnormal variables. A one-way analysis of variance was used for normal distributions, LSD/SNK-q for homogeneity of variance, and Dunnett’s T3 for heterogeneity of variance. A non-parametric test was used for when the data did not follow Gaussian distribution, and the Spearman method was used for correlation analysis. A value of P < 0.05 was considered statistically significant.
RESULTS
HPM inhibits tissue damage in colonic tissues of UC rats
When the colonic tissues were observed for morphological changes upon treatment, we observed that our UC model group exhibited edematous colonic mucosa and there was serious congestion, erosion, and ulcer formation as compared to the normal group, which exhibited a smooth colonic mucosa surface, clear vascular texture, and no erosion or ulcers. Interestingly, HPM treatment was able to decrease the observed changes in UC treated rats and return the colon to a more normal state, as the colonic mucosa surface of these rats was smooth, there was accidental edema, the vascular texture was clear, and congestion, edema and erosion were significantly reduced compared to the model group. As shown in Figure 1, the general morphological score of the colonic tissue in the model group was significantly higher than the normal group [6.0 (5.5-7.0) vs 0.0 (0.0-0.0), P = 0.000]. After treatment, the scores were lower in both the HPM group [3.5 (2.0-4.0) vs 6.0 (5.5-7.0), P = 0.01] and the control SA group [3.0 (1.5-3.5) vs 6.0 (5.5-7.0), P = 0.002]. This data suggests that HPM is able to dampen the tissue-damaging effects of UC.
Figure 1.
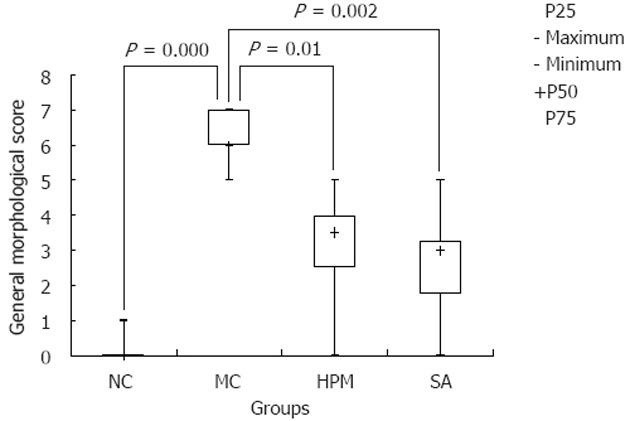
Herb-partition moxibustion inhibits tissue damage in colonic tissues of rats with ulcerative colitis. The general morphological score of the colonic tissue in the model control (MC) group was significantly higher than the normal control (NC) group (P = 0.000). After treatment, the scores were lower in both the herb-partition moxibustion (HPM) group (P = 0.01) and the control sulfasalazine (SA) group (P = 0.002).
HPM inhibits immunopathology in colonic tissues of UC rats
To evaluate whether HPM was also able to regulate the inflammatory cell infiltration that causes tissue damage in UC, we evaluated the extent of immunopathology by looking for inflammatory cell tissue infiltration by HE staining. As shown in Figure 2A-D, the colonic mucosa and mucosa villi were damaged or missing in the UC model group, and large mononuclear cell and macrophage infiltration appeared in the mucosa or submucosa with congestion, edema, and ulceration as compared to the normal group, where the colonic mucosa epithelium was complete and the colonic gland was regularly arranged with inconspicuous inflammatory cell infiltration. However, HPM treatment decreased the inflammatory cell infiltration and the ensuing immunopathology, as we observed only slight submucosal edema and inflammatory cell infiltration, and the colonic mucosa epithelium and the colonic gland were more regularly arranged than in the model group. Additionally, new epithelial cells were observed to be covering the ulcers that developed under UC conditions, indicating that HPM treatment could induce recovery of these ulcers. Positive control SA treatment showed similar recovery of the UC model as the HPM group. As shown in Figure 2E, the histopathological scores for the colonic tissue in the model group were significantly higher than the normal group [5.00 (4.50-5.50) vs 0.00 (0.00-0.50), P = 0.000]. After treatment, the scores were lower in both the HPM group [3.00 (2.00-3.50) vs 5.00 (4.50-5.50), P = 0.007] and SA group [3.00 (2.50-3.50) vs 5.00 (4.50-5.50), P = 0.015]. Taken together, these data indicate that HPM treatment can inhibit inflammatory cell infiltration under UC conditions and therefore limit the resulting immunopathology in the colon tissue.
Figure 2.
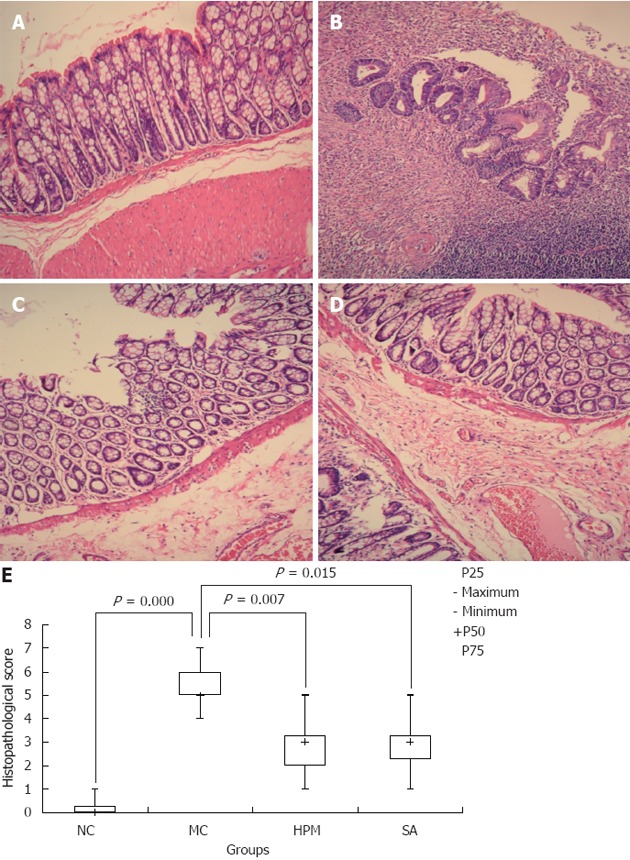
Herb-partition moxibustion inhibits immunopathology and decreases the histopathological scores in colonic tissues of rats with ulcerative colitis. A: Normal; B: Ulcerative colitis (UC); C: Herb-partition moxibustion (HPM); D: Sulfasalazine (SA); E: The histopathological scores for the colonic tissue in the UC model control (MC) group were significantly higher than the normal control (NC) group (P = 0.000). After treatment, the scores were lower in both the HPM group (P = 0.007) and SA group (P = 0.015). The colonic mucosa was damaged in the UC model group with cell infiltration, congestion, edema, and ulceration. After HPM treatment, there are only slight submucosal edema and inflammatory cell infiltration, and the colonic mucosa epithelium and the colonic gland were more regularly arranged than in the model group, new epithelial cells were observed to be covering the ulcers. Positive control SA treatment showed similar recovery of the UC model as the HPM group.
HPM treatment rebalances the colonic intestinal flora of UC rats
Because the deregulation of intestinal flora is now understood to be a major component of UC pathogenesis, we tested whether HPM could restore the balance of the bacteria species back to normal by using the readout of the levels of symbiotic and pathogenic bacteria. To do this, we tested the levels of species-specific bacterial DNA by detecting 16S RNA from rat feces among the different groups. As shown in Figure 3, DNA copies of the symbiotic groups Bifidobacterium and Lactobacillus were both significantly decreased in the UC group compared to the normal group (0.045 ± 0.036 × 105 vs 6.707 ± 1.595 × 105 for Bifidobacterium, 0.0015 ± 0.0014 × 107 vs 3.254 ± 1.395 × 107 for Lactobacillus, P = 0.000). After HPM treatment, the levels were restored, because DNA copies of Bifidobacterium and Lactobacillus were both significantly increased in the HPM group and SA group compared with the model group (1.395 ± 1.339, 1.461 ± 1.152 vs 0.045 ± 0.036 × 105, P = 0.001 in HPM group and P = 0.000 in SA group for Bifidobacterium, and 0.395 ± 0.325, 0.851 ± 0.651 vs 0.0015 ± 0.0014 × 107, P = 0.000 for Lactobacillus). As shown in Figure 4, the DNA copies of the pathogenic bacteria B. fragilis and E. coli were both significantly increased in the UC rats compared to the normal rats (1.285 ± 1.039 vs 0.097 ± 0.063 × 105 for B. fragilis, 1.691 ± 0.683 vs 1.691 ± 0.683 × 105 for E. coli, P = 0.000). After HPM treatment, the DNA copies of B. fragilis and E. coli were both significantly decreased in the HPM group and SA group compared with the UC model group (0.351 ± 0.181 × 105, 0.416 ± 0.329 × 105 vs 1.285 ± 1.039 × 105, P = 0.001 for B. fragilis and 0.244 ± 0.107 × 105, 0.628 ± 0.257 × 105 vs 1.691 ± 0.683 × 105 P = 0.000 in HPM group and P = 0.003 in SA group for E. coli). These data indicate that HPM treatment can rebalance the intestinal flora toward more normal levels after alteration under UC conditions.
Figure 3.
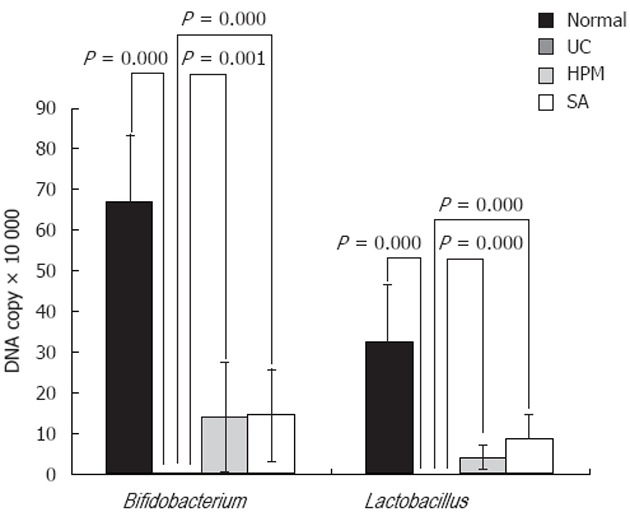
Herb-partition moxibustion treatment increases the colonic Bifidobacterium and Lactobacillus of rats with ulcerative colitis. The DNA copies of the symbiotic groups Bifidobacterium and Lactobacillus were both significantly decreased in the ulcerative colitis (UC) group compared to the normal control (NC) group (P = 0.000). After herb-partition moxibustion (HPM) treatment, the DNA copies of Bifidobacterium and Lactobacillus were both significantly increased in the HPM group (P = 0.001 and P = 0.000) and sulfasalazine (SA) group (P = 0.000 and P = 0.000) compared with the UC model group.
Figure 4.
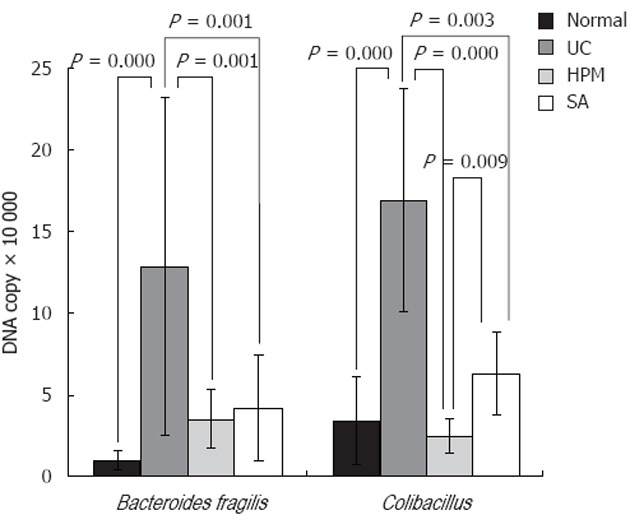
Herb-partition moxibustion treatment decreases the colonic pathogenic bacteria Bacteroides fragilis and Escherichia coli of rats with ulcerative colitis. The DNA copies of the pathogenic bacteria Bacteroides fragilis (B. fragilis), and Escherichia coli (E. coli) were both significantly increased in the ulcerative colitis (UC) rats compared to the normal control (NC) rats (P = 0.000). After herb-partition moxibustion (HPM) treatment, the DNA copies of B. fragilis and E. coli were both significantly decreased in the HPM group (P = 0.001 and P = 0.000) and sulfasalazine (SA) group (P = 0.001 and P = 0.003) compared with the UC model group.
HPM inhibits the secretion of the pro-inflammatory mediators TNF-α and IL-12 in the colon tissue of UC rats
To determine whether the HPM-induced reduction of inflammation was due to an effect of HPM on the secretion of pro-inflammatory cytokines, we measured TNF-α and IL-12 levels in colon tissue by immunohistochemistry. As shown in Figure 5A1-E1, TNF-α expression was significantly increased in the UC model group compared to the normal group (650 313.82 ± 65 996.76 vs 48 384.84 ± 9438.98, P = 0.000). After treatment, TNF-α expression was significantly decreased in both the HPM group and SA group compared with the UC model group (231 783.33 ± 50 222.65, 283 668.65 ± 44 978.06 vs 650 313.82 ± 65 996.76, P = 0.000). There was a difference in TNF-α expression between the HPM group and positive control SA group (231 783.33 ± 50 222.65 vs 283 668.65 ± 44 978.06, P = 0.034). As shown in Figure 5A2-E2, IL-12 expression was significantly increased in the model group compared to the normal group (901 708.26 ± 215 867.35 vs 76 799.88 ± 15 270.78, P = 0.000). HPM treatment inhibits IL-12 expression because it was significantly decreased in both the HPM group and positive control SA group compared with the model group (333 652.88 ± 121 428.18, 512 202.17 ± 95 369.17 vs 901 708.26 ± 215 867.35, P = 0.001 in the HPM group and P = 0.012 in the SA group). There was a difference in IL-12 expression between the HPM group and the SA group (333 652.88 ± 121 428.18 vs 512 202.17 ± 95 369.17, P = 0.033). This data indicates that HPM treatment exerts its anti-inflammatory effects through the inhibition of pro-inflammatory cytokine secretion, such as TNF-α and IL-12.
DISCUSSION
In normal physiological conditions, the overgrowth of pathogenic microorganisms is kept in check by the balance of gut flora species as well as the interaction between microbes and the host’s immune system. These mechanisms limit the pathogenic microorganisms from sticking to the intestinal mucosa and act as a biological barrier to the host[7,27,28]. Altered bacterial composition and function of the gut flora in UC results in increased immune stimulation, epithelial dysfunction, and enhanced mucosal permeability[7]. Moreover, this barrier dysfunction leads to increased recognition of pathogens and their metabolites by the immune system and cause an abnormal intestinal immune response[7] that constantly stimulates intestinal epithelium cells, among other effects. This immune response leads to the production of a large amount of cytokines, such as TNF-α, IL-12 and IL-6, that further aggravate the local immune response of the intestinal mucosa and cause continual damage of the intestinal mucosa[29,30]. In recent years, studies have widely focused on the relationship between intestinal bacteria and UC. Changes in gut flora have been considered as key in the initiation and maintenance stages of the inflammatory processes in UC[7,10,31-35]. There is evidence to support that UC patients undergo alterations in intestinal flora: the amount of Enterobacter, Enterococcus,
and small-bowel Clostridium increased significantly, while Bifidobacteria and Lactobacilli decreased significantly in acute UC. In patients undergoing remission from UC, bacteroid and Bifidobacteria were increased markedly and small-bowel Clostridium was decreased significantly[36,37]. In some studies, bacteroids were significantly increased[38] and Bifidobacteria and Lactobacilli in the gut were both decreased in acute and remission UC[39]. Other studies have indicated that Bifidobacteria, Lactobacilli, and Fusobacterium in the gut of UC patients decreased significantly, while Bacteroides, E. coli, and Desulfovibrio increased remarkably[3-6]. The results of the present study showed that Bifidobacteria and Lactobacilli decreased significantly, while E. coli and B. fragilis significantly increased by testing the bacterial DNA copy numbers in the stool of UC rats as compared to normal rats. These results indicate that gut flora alterations also occur in the rat model of UC used in these studies and provide further evidence that alterations in gut flora play an important role in the pathogenesis of UC.
In recent years, modulation of gut flora has been suggested as an approach to treat UC. The balance between beneficial and detrimental bacterial species determines homeostasis vs inflammation. This balance can be manipulated by antibiotics, probiotics, and prebiotics to treat and prevent relapses of UC[40]. Additionally, alternative treatments or complementary therapy, such as the use of probiotics, have been considered good candidates for treatment of UC because there are less problems with resistance, it means less risk for the development of bacterial resistance to antibiotic treatment, and there are less potential side effects and fewer ecological concerns than using antibiotic drugs[41]. Our previous studies in humans have indicated that HPM treatment is safe and has a therapeutic effect on UC patients[12-14]. To determine the mechanism of how HPM might control UC, the aim of this study was to analyze whether HPM can modulate the gut flora in a rat model of UC. The results shown here demonstrate that HPM treatment does indeed alter the intestinal flora bacterial composition, as Bifidobacteria and Lactobacilli increased significantly, while E. coli and B. fragilis decreased remarkably in the stool of HPM-treated UC rats as compared to the stool of UC rats without treatment. Damage to the colonic mucosa was also remarkably improved after HPM treatment of UC rats, which indicated that the alteration of the gut flora back towards normal rat flora correlated with increased recovery of gut pathology in the UC rats treated with HPM. Therefore, HPM could modulate the alteration of gut flora in the UC rat model, recover intestinal micro-ecological balance, and inhibit the development and/or maintenance of gut pathology. The resultant modulation of gut flora by HPM is similar to that of probiotic supplements[29], and therefore deserves further study as an alternative treatment for UC symptoms.
IL-12 and TNF-α are pre-inflammation factors and play an important role in the pathology of intestinal bowel disease (IBD). IL-12 from dendritic cells induces the differentiation of CD4+ T cells to Th1 cells. IL-12 also stimulates natural killer cells and T cells to secrete γ-interferon and TNF-α cytokines to mediate the inflammation. TNF-α cooperates with γ-interferon and can change the function of the intestinal barrier; it can also enhance the permeability of the mucous membrane or induce apoptosis of bowel epithelial cells. This tissue damage is the key process in the colonic inflammatory response in UC[3,39] and is closely related to the pathogenesis of UC[4,5]. Studies have revealed that IL-12 protein and mRNA expression is increased and correlates with the active index score of UC[5,6]. IL-12 and TNF-α are significantly increased in the peripheral blood of patients with IBD[40-43]. These previous studies indicate the relevance of IL-12 and TNF-α cytokines in the initiation and maintenance of UC pathogenesis.
Previous studies showed that HPM could rectify abnormal mucosal immune responses[43], regulate the expression of IL-1 and IL-1β[44,45], and improve intestinal mucus damage by down-regulating the expression of TGF-β1 and IGF-1 in the inflamed tissue of UC[46,47]. The results of the present study indicate that TNF-α and IL-12 are expressed at low levels in the colonic mucosa and the submucosa of normal rats. However, in UC rats, TNF-α and IL-12 were expressed at high levels that correlated with the observed higher morphological and histopathological scores. After HPM treatment, however, the expression of TNF-α and IL-12 decreased and the general morphological and histopathological scores were lower than those of the UC group, indicating that HPM treatment had an anti-inflammatory effect and improved the UC-mediated pathology. These results further support the previous reports that suggest that HPM can improve the inflammatory response induced by UC, and that the mechanism may be regulation of the expression of inflammatory cytokines IL-6, IL-1β, TNF-α, and IL-12 in the colon mucosa of UC rats.
Additionally, the intestinal flora and their products have been found to trigger cytokine expression, such as inducing TNF-α in macrophage and epithelial cell systems of inflammatory bowel disease[48]. In addition to restoring beneficial intestinal flora, probiotics may enhance host protective immunity such as down-regulation of pro-inflammatory cytokines, IL-12 and TNF-α in colitis[49,50]. Rifaximin administration decreased the protein and mRNA levels of IL-12 and TNF-α, and caused a significant reduction of colon bacterial translocation towards mesenteric lymph nodes, in colon of 2, 4, 6-trinitrobenzene sulfonic acid-induced colitis in mice[51]. The results of this study showed that HPM can modulate intestinal flora towards a more normal flora and inhibit the expression of TNF-α and IL-12 in UC rats. These indicate that there may be a relationship between the release of IL-12 /TNF-α and the modulation of bacterial flora.
In conclusion, we suggest that HPM can modulate intestinal flora towards a more normal flora and inhibit the expression of the pro-inflammatory mediators TNF-α and IL-12 in UC rats. Therefore, whether HPM can improve the intestinal inflammation response of UC by modulating intestinal flora deserves further study as a treatment for patients suffering with UC.
COMMENTS
Background
Intestinal flora plays an important role in human health. Previous studies indicated that herb-partition moxibustion (HPM) has a beneficial effect on ulcerative colitis (UC), not just to relieve the symptoms, but also to improve the stool property, but whether the effect of HPM is related to intestinal flora remains unknown.
Research frontiers
More and more data show that the key role of bacteria in the pathogenesis of inflammatory bowel disease, which has become a hot spot of study.
Innovations and breakthroughs
This study is the first to report on the effects of HPM on intestinal flora of UC. HPM treatment can regulate intestinal flora of the UC model rat.
Applications
The experimental data can be used in the further study of moxibustion therapy in the treatment of inflammatory bowel disease.
Peer review
This is a good original study in which authors observed the relative proportions of both the symbiotic and most prevalent bacteria, and the conditioned pathogenic flora to explore the effects of HPM on modulating the intestinal flora of rats in a model of UC. They also investigated whether HPM regulates the expression of pro-inflammatory cytokines in colon tissues of UC rats. They concluded that HPM treatment regulates intestinal flora and inhibit the expression of interleukin-12 and tumor necrosis factor, indicating that HPM can improve colonic immune response.
Footnotes
Supported by National Natural Science Foundation of China, No. 81001549; National Basic Research Program of China (973 program), No. 2009CB522900; Shanghai Health System of Outstanding Young Talent Cultivation Program, No. XYQ2011068; Shanghai Rising-Star Program, No. 10QA1406100
Peer reviewer: Mitsunori Yamakawa, Professor, Department of Pathological Diagnostics, Yamagata University, Faculty of Medicine, 2-2-2 Iida-Nishi, Yamagata 990-9585, Japan
S- Editor Gou SX L- Editor O’Neill M E- Editor Xiong L
References
- 1.Watanabe S, Yamakawa M, Hiroaki T, Kawata S, Kimura O. Correlation of dendritic cell infiltration with active crypt inflammation in ulcerative colitis. Clin Immunol. 2007;122:288–297. doi: 10.1016/j.clim.2006.10.013. [DOI] [PubMed] [Google Scholar]
- 2.Zhang SZ, Zhang DC. Immunological mechanism of ulcerative colitis. Zhongguo Jianyan Yixue Zazhi. 2006;27:419–421. [Google Scholar]
- 3.Macfarlane S, Furrie E, Kennedy A, Cummings JH, Macfarlane GT. Mucosal bacteria in ulcerative colitis. Br J Nutr. 2005;93 Suppl 1:S67–S72. doi: 10.1079/bjn20041347. [DOI] [PubMed] [Google Scholar]
- 4.Tamboli CP, Neut C, Desreumaux P, Colombel JF. Dysbiosis in inflammatory bowel disease. Gut. 2004;53:1–4. doi: 10.1136/gut.53.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Andoh A, Imaeda H, Aomatsu T, Inatomi O, Bamba S, Sasaki M, Saito Y, Tsujikawa T, Fujiyama Y. Comparison of the fecal microbiota profiles between ulcerative colitis and Crohn’s disease using terminal restriction fragment length polymorphism analysis. J Gastroenterol. 2011;46:479–486. doi: 10.1007/s00535-010-0368-4. [DOI] [PubMed] [Google Scholar]
- 6.Rowan F, Docherty NG, Murphy M, Murphy B, Calvin Coffey J, O’Connell PR. Desulfovibrio bacterial species are increased in ulcerative colitis. Dis Colon Rectum. 2010;53:1530–1536. doi: 10.1007/DCR.0b013e3181f1e620. [DOI] [PubMed] [Google Scholar]
- 7.Sartor RB. Microbial influences in inflammatory bowel diseases. Gastroenterology. 2008;134:577–594. doi: 10.1053/j.gastro.2007.11.059. [DOI] [PubMed] [Google Scholar]
- 8.De Jager PL, Franchimont D, Waliszewska A, Bitton A, Cohen A, Langelier D, Belaiche J, Vermeire S, Farwell L, Goris A, et al. The role of the Toll receptor pathway in susceptibility to inflammatory bowel diseases. Genes Immun. 2007;8:387–397. doi: 10.1038/sj.gene.6364398. [DOI] [PubMed] [Google Scholar]
- 9.Subramanian S, Campbell BJ, Rhodes JM. Bacteria in the pathogenesis of inflammatory bowel disease. Curr Opin Infect Dis. 2006;19:475–484. doi: 10.1097/01.qco.0000244054.69253.f3. [DOI] [PubMed] [Google Scholar]
- 10.Underhill D, Braun J. Current understanding of fungal microflora in inflammatory bowel disease pathogenesis. Inflamm Bowel Dis. 2008;14:1147–1153. doi: 10.1002/ibd.20402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ma BX, Chong GQ. Study on the immunoregulation effect of moxibustion. Jiansu Zhongyiyao. 2008;40:86–87. [Google Scholar]
- 12.Joos S, Wildau N, Kohnen R, Szecsenyi J, Schuppan D, Willich SN, Hahn EG, Brinkhaus B. Acupuncture and moxibustion in the treatment of ulcerative colitis: a randomized controlled study. Scand J Gastroenterol. 2006;41:1056–1063. doi: 10.1080/00365520600580688. [DOI] [PubMed] [Google Scholar]
- 13.Mu JP, Wu HG, Zhang ZQ, Liu HR, Zhu Y, Shi Z, Wang XM. Meta-analysis on acupuncture and moxibustion for treatment of ulcerative colitis. Zhongguo Zhenjiu. 2007;27:687–690. [PubMed] [Google Scholar]
- 14.Zhou EH, Liu HR, Wu HG, Shi Z, Zhang W, Zhu Y, Shi DR, Zhou S. Down-regulation of protein and mRNA expression of IL-8 and ICAM-1 in colon tissue of ulcerative colitis patients by partition-herb moxibustion. Dig Dis Sci. 2009;54:2198–2206. doi: 10.1007/s10620-008-0620-4. [DOI] [PubMed] [Google Scholar]
- 15.Wu HG, Liu HR, Tan LY, Gong YJ, Shi Y, Zhao TP, Yi Y, Yang Y. Electroacupuncture and moxibustion promote neutrophil apoptosis and improve ulcerative colitis in rats. Dig Dis Sci. 2007;52:379–384. doi: 10.1007/s10620-006-9561-y. [DOI] [PubMed] [Google Scholar]
- 16.Shi Y, Zhou EH, Wu HG, Zhou CL, Wang QY, Qi L. Moxibustion treatment restoring the intestinal epithelium barrier in rats with Crohn’s disease by down-regulating tumor necrosis factor alpha, tumor necrosis factor receptor 1, and tumor necrosis factor receptor 2. Chin J Integr Med. 2011;17:212–217. doi: 10.1007/s11655-011-0669-3. [DOI] [PubMed] [Google Scholar]
- 17.The Ministry of Science and Technology of the People¡¯s Republic of China. Guidance Suggestions for the Care and Use of Laboratory Animals. 2006. [Google Scholar]
- 18.Peppelenbosch MP, van Deventer SJ. T cell apoptosis and inflammatory bowel disease. Gut. 2004;53:1556–1558. doi: 10.1136/gut.2004.040824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wu HG, Gong X, Yao LQ, Zhang W, Shi Y, Liu HR, Gong YJ, Zhou LB, Zhu Y. Mechanisms of acupuncture and moxibustion in regulation of epithelial cell apoptosis in rat ulcerative colitis. World J Gastroenterol. 2004;10:682–688. doi: 10.3748/wjg.v10.i5.682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Xu SY, Bian RL, Chen X. Pharmacology experiment methodology. Beijing: People’s Medical Publishing House; 2002. p. 892. [Google Scholar]
- 21.Sugai GC, Freire Ade O, Tabosa A, Yamamura Y, Tufik S, Mello LE. Serotonin involvement in the electroacupuncture- and moxibustion-induced gastric emptying in rats. Physiol Behav. 2004;82:855–861. doi: 10.1016/j.physbeh.2004.07.003. [DOI] [PubMed] [Google Scholar]
- 22.Li ZR. Experimental acupuncture science. Beijing: China press of Traditional Chinese Medicine; 2007. p. 56. [Google Scholar]
- 23.Wu HG, Zhou LB, Shi DR, Liu SM, Liu HR, Zhang BM, Chen HP, Zhang LS. Morphological study on colonic pathology in ulcerative colitis treated by moxibustion. World J Gastroenterol. 2000;6:861–865. doi: 10.3748/wjg.v6.i6.861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fan H, Qiu MY, Mei JJ, Shen GX, Liu SL, Chen R. Effects of four regulating-intestine prescriptions on pathology and ultrastructure of colon tissue in rats with ulcerative colitis. World J Gastroenterol. 2005;11:4800–4806. doi: 10.3748/wjg.v11.i31.4800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhou YY. Review of the clinical application of fibercoloscope. Zhonghua Xiaohua Zazhi. 1984;4:70–72. [Google Scholar]
- 26.Chen YZ. Practical Internal Medicine. Beijing: People’s Medical Publishing House; 2001. p. 17854. [Google Scholar]
- 27.Hold GL, Mukhopadhya I, Monie TP. Innate immune sensors and gastrointestinal bacterial infections. Clin Dev Immunol. 2011;2011:579650. doi: 10.1155/2011/579650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Schirbel A, Fiocchi C. Inflammatory bowel disease: Established and evolving considerations on its etiopathogenesis and therapy. J Dig Dis. 2010;11:266–276. doi: 10.1111/j.1751-2980.2010.00449.x. [DOI] [PubMed] [Google Scholar]
- 29.Sasai M, Tatematsu M, Oshiumi H, Funami K, Matsumoto M, Hatakeyama S, Seya T. Direct binding of TRAF2 and TRAF6 to TICAM-1/TRIF adaptor participates in activation of the Toll-like receptor 3/4 pathway. Mol Immunol. 2010;47:1283–1291. doi: 10.1016/j.molimm.2009.12.002. [DOI] [PubMed] [Google Scholar]
- 30.Byrd-Leifer CA, Block EF, Takeda K, Akira S, Ding A. The role of MyD88 and TLR4 in the LPS-mimetic activity of Taxol. Eur J Immunol. 2001;31:2448–2457. doi: 10.1002/1521-4141(200108)31:8<2448::aid-immu2448>3.0.co;2-n. [DOI] [PubMed] [Google Scholar]
- 31.Bai AP, Ouyong Q. Probiotics treatment of inflammatory bowel disease. Zhonghua Xiaohua Zazhi. 2005;10:240–242. [Google Scholar]
- 32.Hans W, Schölmerich J, Gross V, Falk W. The role of the resident intestinal flora in acute and chronic dextran sulfate sodium-induced colitis in mice. Eur J Gastroenterol Hepatol. 2000;12:267–273. doi: 10.1097/00042737-200012030-00002. [DOI] [PubMed] [Google Scholar]
- 33.Balfour Sartor R. Bacteria in Crohn’s disease: mechanisms of inflammation and therapeutic implications. J Clin Gastroenterol. 2007;41 Suppl 1:S37–S43. doi: 10.1097/MCG.0b013e31802db364. [DOI] [PubMed] [Google Scholar]
- 34.Flanagan P, Campbell BJ, Rhodes JM. Bacteria in the pathogenesis of inflammatory bowel disease. Biochem Soc Trans. 2011;39:1067–1072. doi: 10.1042/BST0391067. [DOI] [PubMed] [Google Scholar]
- 35.van Lierop PP, Samsom JN, Escher JC, Nieuwenhuis EE. Role of the innate immune system in the pathogenesis of inflammatory bowel disease. J Pediatr Gastroenterol Nutr. 2009;48:142–151. doi: 10.1097/MPG.0b013e3181821964. [DOI] [PubMed] [Google Scholar]
- 36.Cui HH, Chen CL, Sun Y, et al. Changing of the flora and humoral immunofunction in patients with inflammatory bowel diseases. Zhongguo Weishengtaixue Zazhi. 2003;15:151–155. [Google Scholar]
- 37.Cui HH. Changes of the flora and human immune response in patients with inflammatory bowel diseases. Weichangbingxue he Ganzangbingxue Zazhi. 2003;12:276–278. [Google Scholar]
- 38.Lucke K, Miehlke S, Jacobs E, Schuppler M. Prevalence of Bacteroides and Prevotella spp. in ulcerative colitis. J Med Microbiol. 2006;55:617–624. doi: 10.1099/jmm.0.46198-0. [DOI] [PubMed] [Google Scholar]
- 39.Dotan I, Rachmilewitz D. Probiotics in inflammatory bowel disease: possible mechanisms of action. Curr Opin Gastroenterol. 2005;21:426–430. [PubMed] [Google Scholar]
- 40.Sartor RB. Microbial-host interactions in inflammatory bowel diseases and experimental colitis. Nestle Nutr Workshop Ser Pediatr Program. 2009;64:121–32; discussion 132-137, 251-257. doi: 10.1159/000235787. [DOI] [PubMed] [Google Scholar]
- 41.Tursi A, Brandimarte G, Papa A, Giglio A, Elisei W, Giorgetti GM, Forti G, Morini S, Hassan C, Pistoia MA, et al. Treatment of relapsing mild-to-moderate ulcerative colitis with the probiotic VSL#3 as adjunctive to a standard pharmaceutical treatment: a double-blind, randomized, placebo-controlled study. Am J Gastroenterol. 2010;105:2218–2227. doi: 10.1038/ajg.2010.218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Brauchle M, Angermeyer K, Hübner G, Werner S. Large induction of keratinocyte growth factor expression by serum growth factors and pro-inflammatory cytokines in cultured fibroblasts. Oncogene. 1994;9:3199–3204. [PubMed] [Google Scholar]
- 43.D’Haens G, Daperno M. Advances in biologic therapy for ulcerative colitis and Crohn’s disease. Curr Gastroenterol Rep. 2006;8:506–512. doi: 10.1007/s11894-006-0041-5. [DOI] [PubMed] [Google Scholar]
- 44.Verri WA, Souto FO, Vieira SM, Almeida SC, Fukada SY, Xu D, Alves-Filho JC, Cunha TM, Guerrero AT, Mattos-Guimaraes RB, et al. IL-33 induces neutrophil migration in rheumatoid arthritis and is a target of anti-TNF therapy. Ann Rheum Dis. 2010;69:1697–1703. doi: 10.1136/ard.2009.122655. [DOI] [PubMed] [Google Scholar]
- 45.Pang YH, Zheng CQ, Yang XZ, Zhang WJ. Increased expression and activation of IL-12-induced Stat4 signaling in the mucosa of ulcerative colitis patients. Cell Immunol. 2007;248:115–120. doi: 10.1016/j.cellimm.2007.10.003. [DOI] [PubMed] [Google Scholar]
- 46.Nielsen OH, Kirman I, Rüdiger N, Hendel J, Vainer B. Upregulation of interleukin-12 and -17 in active inflammatory bowel disease. Scand J Gastroenterol. 2003;38:180–185. doi: 10.1080/00365520310000672. [DOI] [PubMed] [Google Scholar]
- 47.Tessner TG, Cohn SM, Schloemann S, Stenson WF. Prostaglandins prevent decreased epithelial cell proliferation associated with dextran sodium sulfate injury in mice. Gastroenterology. 1998;115:874–882. doi: 10.1016/s0016-5085(98)70259-8. [DOI] [PubMed] [Google Scholar]
- 48.Klapproth JM, Sasaki M. Bacterial induction of proinflammatory cytokines in inflammatory bowel disease. Inflamm Bowel Dis. 2010;16:2173–2179. doi: 10.1002/ibd.21332. [DOI] [PubMed] [Google Scholar]
- 49.Chen CC, Kong MS, Lai MW, Chao HC, Chang KW, Chen SY, Huang YC, Chiu CH, Li WC, Lin PY, et al. Probiotics have clinical, microbiologic, and immunologic efficacy in acute infectious diarrhea. Pediatr Infect Dis J. 2010;29:135–138. doi: 10.1097/inf.0b013e3181b530bf. [DOI] [PubMed] [Google Scholar]
- 50.Nanda Kumar NS, Balamurugan R, Jayakanthan K, Pulimood A, Pugazhendhi S, Ramakrishna BS. Probiotic administration alters the gut flora and attenuates colitis in mice administered dextran sodium sulfate. J Gastroenterol Hepatol. 2008;23:1834–1839. doi: 10.1111/j.1440-1746.2008.05723.x. [DOI] [PubMed] [Google Scholar]
- 51.Fiorucci S, Distrutti E, Mencarelli A, Barbanti M, Palazzini E, Morelli A. Inhibition of intestinal bacterial translocation with rifaximin modulates lamina propria monocytic cells reactivity and protects against inflammation in a rodent model of colitis. Digestion. 2002;66:246–256. doi: 10.1159/000068362. [DOI] [PubMed] [Google Scholar]