

Abstract
Eyes absent (Eya) is a highly conserved transcription cofactor and protein phosphatase that plays an essential role in eye development and survival in Drosophila. Ectopic eye induction assays using cDNA transgenes have suggested that mitogen activated protein kinase (MAPK) activates Eya by phosphorylating it on two consensus target sites, S402 and S407, and that this activation potentiates the ability of Eya to drive eye formation. However, this mechanism has never been tested in normal eye development. In the current study, we generated a series of genomic rescue transgenes to investigate how loss- and gain-of-function mutations at these two MAPK target sites within Eya affect Drosophila survival and normal eye formation: eya+GR, the wild-type control; eyaSAGR, which lacks phosphorylation at the two target residues; and eyaSDEGR, which contains phosphomimetic amino acids at the same two residues. Contrary to the previous studies in ectopic eye development, all eya genomic transgenes tested rescue both eye formation and survival equally effectively. We conclude that, in contrast to ectopic eye formation, MAPK-mediated phosphorylation of Eya on S402 and S407 does not play a role in normal development. This is the first study in Drosophila to evaluate the difference in outcomes between genomic rescue and ectopic cDNA-based overexpression of the same gene. These findings indicate similar genomic rescue strategies may prove useful for re-evaluating other long-standing Drosophila developmental models.
Introduction
eyes absent (eya) encodes a highly conserved transcriptional coactivator and protein phosphatase whose homologs play vital roles in human development [1], [2], [3], [4]. Mutations in human EYA1 lead to the autosomal dominant disorder known as branchio-oto-renal (BOR) syndrome, characterized by craniofacial anomalies, hearing loss, and kidney defects [5]. In addition, EYA overexpression occurs in a number of human solid tissue tumors, and correlates with poor prognosis in breast cancer [6], [7], [8], [9].
Drosophila melanogaster has a single eya gene, which is essential for survival [1], [10]. Eya regulates development of the gonads, muscle, and the eye, where it has been most extensively studied [1], [11], [12], [13]. The compound eye of adult Drosophila arises from a larval epithelial structure known as the eye imaginal disc. The eye disc initially consists of undifferentiated, proliferating cells. Later, at the onset of the third instar larval stage, an indentation called the morphogenetic furrow forms at the posterior margin of the eye disc and sweeps toward the anterior margin, triggering the onset of differentiation [14]. Eya expression begins in the eye disc during the second instar stage, prior to furrow initiation. Once the furrow starts, Eya continues to be expressed in a domain anterior to the furrow as well as in differentiating cells in the posterior part of the disc [1]. Eye discs that lack eya begin to develop normally, but the furrow fails to initiate, differentiation does not occur, and the eye disc undergoes widespread apoptosis, resulting in complete loss of the adult eye [1]. eya also appears to be required for differentiation or survival of photoreceptor cells behind the furrow [15]. Hence, understanding the regulation of Eya function may provide essential insight into the mechanisms of eye development.
Eya is a member of the retinal determination (RD) network, a small group of highly conserved transcriptional regulators that are both necessary for eye development and sufficient to trigger ectopic eye formation when overexpressed in other imaginal discs. Other key members of the RD network include Eyeless, Sine oculis, and Dachshund [16], [17], [18], [19]. Eye development involves complex regulatory interactions among the RD members as well as signaling pathways [20], [21], [22], [23], [24]. While the regulation of RD genes has been studied extensively at the level of transcription [25], [26], [27], [28], [29], relatively little is known about the role of post-translational modification in regulating RD factors.
Previous studies have suggested that one mechanism of post-translational regulation of Eya activity in the eye is through phosphorylation by mitogen activated protein kinases (MAPK). Two Eya residues, S402 and S407, strongly match the MAPK target motif [30]. These residues have been shown to undergo phosphorylation in vitro by the MAPK family kinases Erk and Nemo (Nmo) [31], [32]. Using ectopic eye induction as an assay of Eya activity, these studies have suggested that phosphorylation by Erk and Nmo at these residues activates Eya [31], [32]. A transgene encoding a protein that cannot be phosphorylated at these residues (UAS-eyaSA) shows a lower frequency of ectopic eye induction compared with a wild-type UAS-eya transgene. In contrast, UAS-eyaSDE, which encodes a phosphomimetic protein, induces ectopic eyes more frequently than wild-type UAS-eya [31]. Genetic interaction studies, likewise using ectopic eye induction, have supported this model. Specifically, co-overexpression of eya with a hyperactive allele of rolled (rl, which encodes Erk) or with nmo leads synergistically to more and larger ectopic eyes. Conversely, loss of one copy of rl or nmo leads to weaker ectopic eye induction by eya [31], [33]. UAS-eyaSA also loses the ability to synergize with UAS-nmo in ectopic eye induction [32]. Together, these results have led to the currently accepted model that MAPK-mediated phosphorylation of Eya on S402 and S407 positively regulates Eya in development.
The above studies all utilized ectopic overexpression assays, in which the cDNA for the gene of interest is expressed in a defined spatiotemporal domain using the Gal4/UAS system [34]. This approach has yielded many insights into the regulatory relationships among genes, as well as helping discern the in vivo function of protein domains and motifs [16], [19], [26], [35], [36], [37], [38]. However, Gal4/UAS assays face certain limitations: the levels of transgene expression differ from those of the endogenous gene, and random integration of the transgene into the genome leads to position effects, which make direct comparison of distinct transgene lines problematic. Moreover, not all findings of an ectopic expression experiment may be applicable to normal development, where the gene of interest acts in a different cellular context and in the presence of different binding partners and signaling pathways that may affect its function. More recently, the development of genomic rescue transgenes has made it possible to analyze the function of protein domains and motifs in the context of normal development [39]. However, to date, genomic rescue has not yet been employed to verify the native function of genes previously defined in ectopic studies. For these reasons, we sought to analyze the function of the MAPK target residues of Eya in the context of normal rather than ectopic eye development, using a genomic rescue strategy.
In the current study, we generate a series of eya genomic rescue constructs that fully rescue eye development, as well as all other known eya mutant phenotypes. Surprisingly, we find that in contrast to the effect of Eya phosphorylation on ectopic eye induction [31], [32], neither loss of MAPK target sites S402 and S407 nor phosphomimetic mutations at these sites affects normal eye development or survival. Our study is the first example of a genomic rescue system yielding results different from those of a cDNA-based ectopic overexpression assay, and underscores the importance of studying a gene in its native context.
Results
The transgene eya+GR rescues eya mutant phenotypes
Prior evidence that MAPK-mediated phosphorylation activates Eya came from ectopic eye induction studies, which relied on Gal4/UAS-mediated overexpression of cDNA-based eya transgenes [31], [32]. To investigate how phosphorylation regulates Eya function during normal eye development, we generated genomic rescue transgenes, which offer two key advantages over the Gal4/UAS system. First, the genomic transgene contains regulatory sequences that drive expression of the gene of interest in a wild-type pattern and at levels matching those of the native gene. Second, the transgene is inserted in a specific site in the genome, allowing comparison of independent transgenic lines without confounding position effects [39]. We made eya+GR (Genomic Rescue), a 58.8 kb fragment encompassing the eya gene and flanking regions, in the P[acman] vector using recombineering [39] (Fig. 1). We used a large genomic rescue fragment to increase the chances of including regulatory regions necessary for all eya expression, as eya enhancers are currently not fully characterized. The transgene was introduced into P2, a specific and reproducible “attP” insertion site on the third chromosome [39].
Figure 1. Construction of eya+GR and point mutations.

A. Schematic of the eya+GR transgene (adapted from http://flybase.org). The red box indicates the extent of the 58.8 kb genomic rescue fragment, which includes the entire eya gene, as well as the upstream sequence up to the nearest gene (osm-6) and the entire gene that lies immediately downstream of eya, CG31637. CG31637 encodes a predicted sulfotransferase and has no known mutant phenotypes. The transgene was made in attB-P[acman]-ApR (see Materials and Methods). B. Schematic of the eya gene. Yellow diamonds indicate the MAPK target sites S402 and S407, both of which are in the PST (proline-serine-threonine rich) transcriptional coactivator domain of Eya, indicated by the maroon rectangle.
We tested the ability of eya+GR to rescue eye development in eya2 mutants. eya2 has a deletion of an eye-specific enhancer, leading to loss of eya expression only in the eye disc. Consequently, eya2 homozygous adults are viable and fertile, but completely lack eyes [1], [40]. One copy of eya+GR rescues eye formation in eya2 homozygotes. The eyes of eya2; eya+GR/+ flies are indistinguishable from wild type by external morphology and size, and sections show wild-type arrangement and number of rhabdomeres per ommatidium (Fig. 2).
Figure 2. eya+GR, eyaSAGR, and eyaSDEGR rescue eye development and survival in eya mutants.

External (A–E, J–M) and internal (F–I, N–Q) adult eye morphology is fully rescued with genomic transgenes in eya mutants. Homozygous eya2 adults carrying a single copy of eya+GR (B, G), eyaSAGR (C, H), or eyaSDEGR (D, I) have eye morphology indistinguishable from wild type Canton S (A, F), whereas eya2 homozygotes without a rescue construct show complete loss of the compound eye (E). eyacliIID/Df flies die due to lack of all endogenous eya function, and one copy of eya+GR (I, M), eyaSAGR (J, N), or eyaSDEGR (K, O) restores viability and normal eye morphology. Two copies of eyaSDEGR (M, Q) rescue viability and eye morphology in eya null adults equally well as one copy of eyaSDEGR, indicating no gain-of-function phenotype due to the phosphomimetic EyaSDE protein.
We also tested whether eya+GR can rescue eyacliIID, a null allele that results in embryonic lethality [10]. eyacliIID fails to complement Df(2L)BSC354 (hereafter referred to as Df), a molecularly defined deficiency [41] that uncovers eya. One copy of eya+GR fully rescues the lethality of eyacliIID/Df flies, and the adult eyes of rescued flies are indistinguishable from wild-type, both by external morphology and in sections (Fig. 2). We refer to eyacliIID/Df; eya+GR/+ flies hereafter as eya−; eya+GR flies. The late third instar larval eye discs of eya−; eya+GR are indistinguishable from wild type eye discs in size and morphology, and immunohistochemistry reveals similar Eya levels and expression patterns (Fig. 3). Likewise, differentiation proceeds normally, as shown by the R8 photoreceptor marker Senseless (Sens) (Fig. 3). eya−; eya+GR flies are present at Mendelian ratios (Table 1), indicating that the rescued flies do not have a survival disadvantage compared with their eyacliIID/CyO or Df/CyO siblings. Hence, a single copy of the eya+GR transgene is functionally equivalent to the single copy of endogenous wild-type eya on the CyO chromosome of eya−/CyO flies. We also tested the function of rescued eyes using electroretinograms (ERG) and found no difference between wild-type (Canton S) and eya−; eya+GR flies (Fig. 4). In addition, Eya regulates photoreceptor axon targeting to the brain [42]. We analyzed photoreceptor axon projections in eya−; eya+GR adults and third instar larvae, and found no difference in axon projections compared with Df/CyO (Fig. 5). Altogether, these observations indicate that one copy of our wild-type rescue transgene behaves similarly to an endogenous copy of eya.
Figure 3. eya+GR, eyaSAGR, and eyaSDEGR rescue larval eye disc development.

Late third instar eye imaginal discs from Df/+ (A), eyacliIID/Df; eya+GR/+ (B), eyacliIID/Df; eyaSAGR/+ (C), and eyacliIID/Df; eyaSDEGR/+ (D) larvae have been stained with anti-Eya (A–D) and anti-Sens to mark differentiating R8 photoreceptors (E–H); merged images shown in I–L. Eya expression pattern and levels are similar between heterozygous larvae and null eya larvae rescued with one copy of each eya transgene. Differentiation occurs normally in rescued eye discs.
Table 1. eya*GR fully rescues viability in eya− flies.
| Genotype | Cy obs. | Cy exp. | Non-Cy obs. | Non-Cy exp. | Total | ?2 |
| eya+GR | 278 | 273 | 131 | 136 | 409 | 0.28 |
| eyaSAGR | 216 | 223 | 119 | 112 | 335 | 0.66 |
| eyaSDEGR | 261 | 259 | 127 | 129 | 388 | 0.046 |
Progeny from w; eyacliIID/CyO; eya*GR×w; Df(2L)BSC354/CyO cross are present at Mendelian ratios, indicating that eya− flies rescued with a single copy of eya+GR, eyaSAGR, or eyaSDEGR do not have a survival disadvantage compared with eya−/CyO siblings. χ2 critical (1 d.f. P 0.05) = 3.84.
Figure 4. Animals rescued with eya*GR have normal eye function.

Representative ERG traces of Canton S (wild-type) (A), eyacliIID/Df; eya+GR/+ (B), eyacliIID/Df; eyaSAGR/+ (C), and eyacliIID/Df; eyaSDEGR/+ (D) adults. eya null adults rescued with a single copy of eyaSAGR or eyaSDEGR show ERG responses indistinguishable from flies rescued with eya+GR or wild-type flies.
Figure 5. Photoreceptor axon projections are normal in flies rescued with eya*GR.
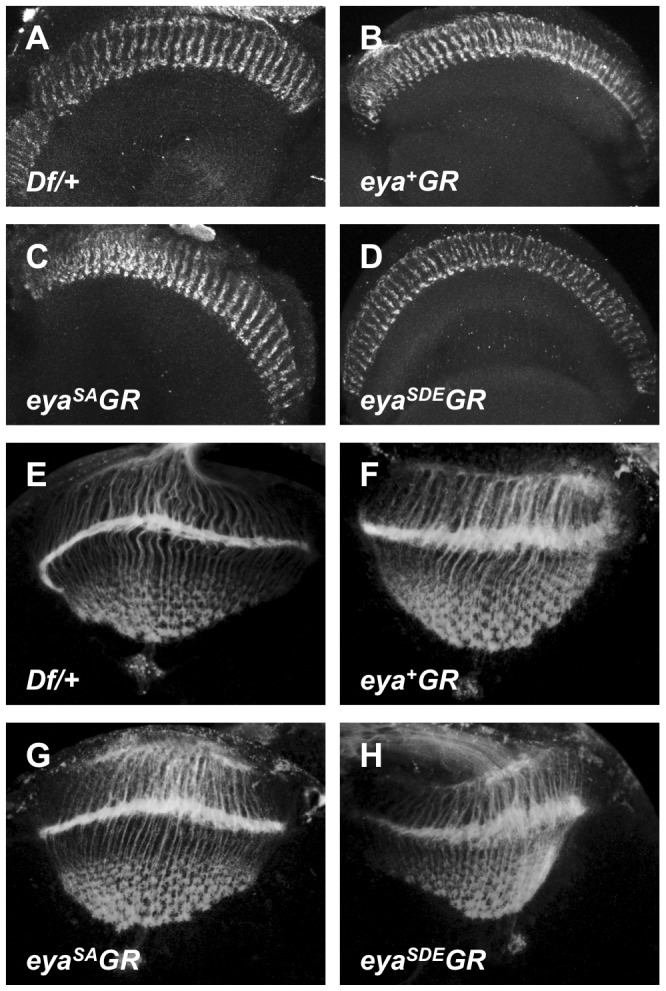
Photoreceptor axon projections to the optic lobe of Df/+ (A, E), eyacliIID/Df; eya+GR/+ (B, F), eyacliIID/Df; eyaSAGR/+ (C, G), and eyacliIID/Df; eyaSDEGR/+ (D, H) are indistinguishable from each other in both adults (A–D) and wandering third instar larvae (E–H). Axon projections are visualized with anti-Chaoptin (24B10).
Since Eya is required for somatic gonad development, and a partial loss-of-function eya allele causes male and female sterility [11], [12], [43], we tested whether eya+GR rescues fertility in eyacliIID homozygotes. eyacliIID; eya+GR females are fertile (data not shown), and eya+GR rescued males produce indistinguishable numbers of progeny from eya heterozygous control males when crossed to w virgin females (Fig. 6). In addition to being required for survival and for eye and gonad development, eya regulates muscle development [13]. The eyacliIID; eya+GR adults appear to move normally and are able to fly (data not shown), indicating that gross muscular defects in these flies are unlikely. In summary, we conclude that the eya+GR construct fully rescues all known aspects of eya function in Drosophila, and provides a critical tool for examining the role of specific Eya residues in normal eye development and survival.
Figure 6. eya*GR rescues fertility in eya null males.
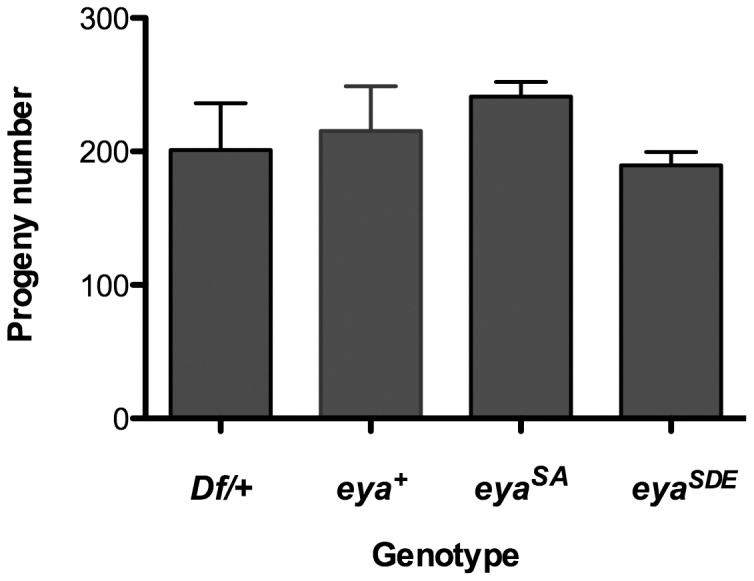
The numbers of progeny are not statistically different (P = 0.156) among w virgin females crossed to Df/CyO control, eya−; eya+GR, eya−; eyaSAGR, and eya−; eyaSDEGR males. Error bars indicate standard deviation. n = 3 crosses for each genotype.
The phosphorylation state of Eya residues S402 and S407 does not affect eye morphology or survival
Having found that eya+GR rescues eya loss-of-function phenotypes, we next generated two point-mutant genomic rescue constructs to investigate the role of Eya phosphorylation during normal eye development. Eya residues S402 and S407 are two serines that undergo Erk- and Nmo-mediated phosphorylation [31], [32]. In the first point-mutant genomic construct, eyaSAGR, we mutated S402 and S407 to alanines, which cannot be phosphorylated. Conversely, in the second point-mutant genomic construct, eyaSDEGR, we mutated S402 and S407 to aspartate and glutamate, respectively, mimicking constitutive phosphorylation. We integrated both constructs into the same attP site as eya+GR in order to avoid differences among the rescue phenotypes due to position effects (see Materials and Methods).
Based on previous data [31], we expected that eyaSAGR would result in partial or no rescue of eye formation in eya mutants, and that eyaSDEGR would lead either to full rescue or to a phenotype partially resembling Eya overexpression in the eye. Unexpectedly, both eyaSAGR and eyaSDEGR (hereafter referred to collectively as eya*GR) rescue eye formation in eya2 homozygotes to the same extent as eya+GR. The external morphology of eya2; eya*GR/+ eyes is indistinguishable from eya2; eya+GR/+ eyes (Fig. 2). Sections reveal the normal number and arrangement of photoreceptors in eya2; eya*GR/+ eyes (Fig. 2).
We also tested the ability of eya*GR to restore survival and eye formation to eya null mutants. eya−; eya*GR adults are viable and present at Mendelian ratios (Table 1). Moreover, the eyes of eya−; eya*GR adults appear wild-type in external morphology and in sections (Fig. 2). Because a single copy of eya*GR appears to be functionally equivalent to one copy of endogenous eya in restoring viability, we performed the remaining experiments on eya null flies with one copy of eya*GR. Since eyaSDEGR encodes a phosphomimetic function that may act as a hypermorph, we considered the possibility that eya null flies rescued with two copies of eyaSDEGR may display gain-of-function phenotypes not seen in flies with one copy of eyaSDEGR. However, eyacliIID homozygous adults with two copies of eyaSDEGR have the same external and internal eye morphology as null flies with a single copy of eya*GR (Fig. 2).
Based on the adult eye phenotypes of eya−; eya*GR flies, we expected to observe normal Eya expression in the developing eye discs of eya−; eya*GR larvae. Immunohistochemistry reveals similar Eya levels and expression patterns among Df/+, eya−; eya+GR, and eya−; eya*GR late third instar larval eye discs (Fig. 3). This is consistent with previous findings that the SA and SDE mutations do not affect Eya stability [31]. Loss of eya from the eye disc leads to a failure of differentiation and to widespread apoptosis, which causes a severe reduction in eye disc size in the third instar [1]. By contrast, the eye discs of eya*GR-rescued flies are indistinguishable from eya−; eya+GR and wild type eye discs in size and morphology, and differentiation proceeds normally, as shown by the R8 photoreceptor marker Senseless (Sens) (Fig. 3).
In addition to restoring survival and eye development, eya*GR transgenes also rescue fertility and muscle development. We observed that eyacliIID; eya*GR females are fertile (data not shown), and we quantified the number of progeny produced by eya*GR rescued males crossed to w virgin females (Fig. 6). eya*GR rescued males produce statistically the same numbers of progeny as males rescued with eya+GR and eya heterozygous males (Fig. 6). Similarly to eya−; eya+GR adults, the eya−; eya*GR adults appear to move normally, making it unlikely that S402 and S407 phosphorylation regulates Eya in muscle development (data not shown). We conclude that, in contrast to previous findings using ectopic eye induction as an assay [31], MAPK-mediated phosphorylation of S402 and S407 does not regulate Eya during normal Drosophila development.
Phosphorylation of Eya residues S402 and S407 is not required for eye function
Electroretinogram (ERG) assays indicate that normal response to light is rescued in eya−; eya*GR eyes (Fig. 4). In addition, we analyzed photoreceptor axon projections to the adult brain in eya−; eya*GR flies. Since tyrosine phosphorylation of Eya by the Abelson kinase regulates photoreceptor axon projections [42], we asked whether Eya phosphorylation by MAPK might also play a role in this process. However, we found no difference in axon projections between eya−; eya*GR and eya−; eya+GR adults (Fig. 5). Likewise, eya null third instar larvae rescued with a copy of eya*GR show an even pattern of axon projections at the lamina of the optic lobe that is the same as in eya heterozygotes, rather than the irregular gaps and thickenings in the lamina plexus previously reported in eya loss-of-function mutant larvae [42]. We conclude that phosphorylation of Eya at S402 and S407 is not required for eye development or function.
Discussion
Drosophila eyes absent (eya) is essential for survival [1], [10] and is required for eye development, as well as regulating development of the gonads and muscles [1], [11], [12], [13]. Previous studies using ectopic overexpression of eya cDNA transgenes during the past decade have suggested that phosphorylation by MAPK may activate Eya during Drosophila eye development [31], [32]. Similar to the Drosophila studies, a recent report in mice showed that loss of conserved MAPK target sites in murine Eya1 reduced its ability to induce ectopic hair cell formation in the cochlea [44]. These cumulative ectopic expression studies in multiple species have led to the current model that Eya function is activated by phosphorylation at specific, conserved MAPK target sites. However, in contrast to these ectopic studies, we show that both the genomic rescue transgene eyaSAGR, which leads to loss of phosphorylation on two MAPK target residues, and eyaSDEGR, which encodes a protein with phosphomimetic amino acids at the same two residues, rescue normal eye development and survival in eya mutants as effectively as the wild-type eya+GR transgene. While we cannot rule out subtle phenotypes, overall our data indicate that phosphorylation of Eya on S402 and S407 does not regulate Eya function during normal development.
Differences between ectopic eye induction vs. normal eye development
The ability of eyaSAGR and eyaSDEGR to rescue eye development may be explained by the difference in context between the eye disc and other discs being reprogrammed to form ectopic eyes. Overexpression of a retinal determination network (RD) transgene such as eya can trigger ectopic eye formation only in retinal “hot spots”, small subsets of cells in the antenna, leg, wing, and haltere imaginal discs [45]. Some of these “hot spots” overlap with sites capable of transdetermination (a process in which one type of imaginal disc assumes the fate of another, in response to injury or genetic manipulation) [46], [47]. This suggests that ectopic eye induction is confined to populations of cells that have a high level of developmental plasticity, perhaps due to their chromatin state or the activity of signaling pathways [45]. Transforming these cell populations into ectopic retinal tissue may require different factors from those needed for normal eye development.
For example, sine oculis (so) encodes a transcription factor in the RD network that is both necessary and sufficient for eye development [18], [48]. A UAS transgene encoding a constitutively repressive form of So can still induce ectopic eyes in the antenna, but it cannot rescue the loss of the normal eye in so1 mutants. Conversely, a UAS transgene encoding So fused to a strong transcriptional activator domain fails to trigger ectopic eye formation, but it restores normal eye development in so1 mutants. These results indicate different requirements for transcriptional repression vs. activation by So in ectopic and normal eye formation [49]. In another example of a difference between ectopic and normal eye development, the kinase Nmo synergizes with Eya to induce ectopic expression of dachshund (dac) and lozenge (lz), yet neither dac nor lz expression is affected in nmo loss-of-function clones in the eye [32], [33].
Taken together, our results and previous studies [31], [32] indicate that the function of Eya phosphorylation differs between ectopic and normal eye development. One explanation may be that a higher level of Eya activity is needed to reprogram antenna into eye, compared with the level required for normal eye formation. Alternatively, phosphorylation may improve Eya's ability to regulate a target (or targets) in the antenna that is not normally expressed in the eye disc or relevant to normal eye development.
Transgene position effects in previous ectopic eye studies
In addition to the differences between normal and ectopic eye development, the transgenic systems used in previous studies vs. the current study may have contributed to the difference in results as well. The previous study [31] tested the effect of phosphorylation on Eya function using UAS-eya transgenes that integrated randomly in the genome. While the average efficiency of eye induction was higher for UAS-eya+ (49%) than for UAS-eyaSA (19%), and lower for UAS-eya+ than for UAS-eyaSDE (81%), these averages do not reveal the large differences among lines expressing the same transgene. Among eight independent UAS-eya+ transgenic lines, the frequency of ectopic eye induction ranged from 3% to 78% [31], suggesting considerable position effects. In contrast, we used site-specific integration of eya genomic rescue transgenes, which is expected to minimize the differences among transgenic lines due to position effects.
Alternative mechanisms for Eya regulation
While our results indicate that S402 and S407 of Eya are not required for survival or eye formation, we do not rule out the possibility that Erk and/or Nmo regulates Eya in the eye by phosphorylation on other serine or threonine residues. S402 and S407 are the only Eya residues that match the “strong” consensus MAPK target site, but 14 additional previously unstudied Eya residues match a less stringent MAPK consensus [30]. Ten of these “weak” MAPK consensus sites reside along with S402 and S407 in the Eya proline-serine-threonine rich (PST) domain, which is robustly phosphorylated by Erk and Nmo in vitro. While phosphorylation of S402 and S407 appears to account for approximately 80% of Nmo-mediated and more than 90% of Erk-mediated phosphorylation of the Eya PST domain in vitro, when S402 and S407 are both mutated to alanines, the point-mutant PST domain can still be weakly phosphorylated in vitro by MAPK [31], [32]. Whereas Nmo appears to phosphorylate only the N-terminal part of Eya, which includes the PST domain, rather than the C-terminal domain [32], the ability of Erk to phosphorylate Eya C-terminal domain has not been assayed. It remains to be tested which Eya residues besides S402 and S407 can be phosphorylated by Erk and/or Nmo, and whether such phosphorylation regulates Eya in vivo. The observed interaction between Eya and MAPK might also be due to MAPK-mediated phosphorylation of the Eya binding partner and transcription factor So, which undergoes serine/threonine phosphorylation in cell culture [50]. Future studies, perhaps using mass spectrometry, will be needed to elucidate the extent of Eya and/or So phosphorylation by MAPK.
The investigation of MAPK-mediated phosphorylation of Eya was prompted by the finding that eya interacts genetically with the Egfr pathway in the eye [51]. Activation of the Egfr pathway in the eye leads to activation of the MAPK Erk, which can then phosphorylate Eya (reviewed by [52]). However, if MAPK does not regulate Eya by phosphorylation, the genetic interaction between eya and the Egfr pathway may be due to transcriptional activation of eya by Egfr signaling. A recent study has shown that Egfr signaling regulates eya expression in both the eye disc and the embryo, and that loss of pointed, which encodes a transcription factor that acts downstream of Egfr, causes strong reduction in Eya expression in eye disc clones [53].
In summary, Eya and its homologs play an essential role in regulating multiple aspects of development from Drosophila to humans. A long-standing model of Eya regulation, based on ectopic eye induction assays, has posited that MAPK-mediated phosphorylation at residues S402 and S407 potentiates Eya activity [31], [32]. Using genomic rescue transgenes, we demonstrate that unlike ectopic eye development, normal eye formation and survival are unaffected by either loss of phosphorylation or phosphomimetic mutations at the two previously studied MAPK target residues of Eya. This is the first study to use a genomic rescue approach in Drosophila to reassess the biological relevance of ectopic overexpression studies of the same gene. These findings indicate similar genomic rescue strategies may prove useful for re-evaluating other Drosophila developmental models.
Materials and Methods
Construction of eya+GR
A 58.8 kb fragment encompassing the eya gene and flanking regions was cloned into the attB-P[acman]-ApR vector using recombineering as described previously [39]. The resulting construct was end-sequenced and integrated into the P{CaryP}attP2 site (abbreviated P2), which is located at 3L:11,063,638, using φC31 integrase [54]. Site-specific integration was confirmed by PCR with attB/attP primers [39].
Recombineering-induced point mutagenesis of eya+GR
The codons encoding S402 and S407 (TCC and TCG, respectively) in eya+GR were mutated to GCC and GCG to make eyaSAGR and to GAC and GAG to make eyaSDEGR. We used two-step recombineering with the catSacB cassette, which provides positive selection (cat, chloramphenicol resistance) and negative selection (SacB, sucrose sensitivity). The protocol was performed as described previously [55]. Since S402 and S407 are only five amino acid residues apart, both codons were targeted in one recombineering event. Putative recombinants were tested by sequencing, and true positives were tested for rearrangements by restriction digest fingerprinting. Both point mutant transgenes were injected into P2, the same site used for eya+GR, and site-specific integration was verified by PCR with attB/attP primers [39]. The presence of point mutations was verified by restriction digest of genomic PCR products. Both S407A and S407E mutations create a PvuI site restriction enzyme site (CGATCG) that is not present in wild-type eya (CGATCT). We performed PCR on genomic DNA from wild-type, eya+GR, eyaSAGR, and eyaSDEGR adults using MAPK1-for and MAPK2-rev primers that flank the S402 and S407 codons and give a 1,058 bp product. The PCR product was digested with PvuI and run on a gel. The enzyme cut the eyaSAGR and eyaSDEGR PCR product only, resulting in 500 and 558 bp bands. Primer sequences are available on request.
Histology and imaging of adult eyes
Adult eye sections were performed as described previously [56]. Images of eye sections and whole adult eyes were taken with a Zeiss Axioplan 2 microscope and AxioVision software. Images of whole-mount adult eyes were processed with CZ Focus software. All images were further processed with Adobe Photoshop software.
Immunohistochemistry of adult brains and 3rd instar eye discs
Brains were dissected and stained as previously described [57] out of adults and late wandering third instar larvae. For larval eye disc dissections, w/Y; eyacliIID/BSC354; eya*GR/+ males were crossed with w; BSC354/CyO, GFP females and progeny larvae were scored for absence of GFP expression (w; eyacliIID/BSC354; eya*GR/+). Eye imaginal discs were dissected out of wandering third instar larvae in 1× PBS and fixed in 4% formaldehyde in PBS 20 minutes on ice. Discs were washed with PBS, PAXD (1× PBS with 1% BSA, 0.3% Triton X-100, and 0.3% sodium deoxycholate), and PAXDG (5% normal goat serum in PAXD) on ice, 10 minutes per wash. Discs were then incubated with primary antibody in PAXDG at 4°C overnight. Subsequent steps were at room temperature. The following day the discs were washed 3× with PAXDG, 10 minutes per wash, and incubated in secondary antibody in PAXDG 2 hours. The discs were washed with PAXDG, PAXD, and PBS, 10 minutes per wash, and post-fixed in 4% formaldehyde in PBS 15 minutes. The discs were then washed twice with PBS (first wash quick, second wash 10 minutes) and incubated in Vectashield (Vector Laboratories, Inc.). Primary antibodies used were 1∶100 mouse anti-Chaoptin (24B10, Developmental Studies Hybridoma Bank), 1∶200 mouse anti-Eya (10H6, Developmental Studies Hybridoma Bank) and 1∶100 guinea pig anti-Sens (gift from H. Bellen). Secondary antibodies used were Cy3 goat anti-mouse (1∶200 for brains, 1∶500 for discs; Jackson ImmunoResearch) and 1∶500 Alexa Fluor 488 goat anti-guinea pig (Molecular Probes). Images were taken with a Zeiss LSM 510 confocal microscope and processed with Image J and Adobe Photoshop software.
Electroretinogram recordings
Electroretinograms were performed as described previously [58]. Six three-day-old adults were assayed for each genotype.
Survival assay
w/Y; eyacliIID/CyO; eya*GR males were crossed to w; Df(2L)BSC354/CyO, Kr-GFP virgin females. Expected (Mendelian) progeny ratios were 2/3 Cy (eyacliIID/CyO and BSC354/CyO) and 1/3 non-Cy (eyacliIID/BSC354). Based on observed numbers of adult progeny, for eya+GR, χ2 calculated = 0.28. For eyaSAGR, χ2 calculated = 0.66. For eyaSDEGR, χ2 calculated = 0.046. χ2 critical (1 d.f. P 0.05) = 3.84. For all genotypes, χ2 calculated<χ2 critical. Hence, for all genotypes, observed progeny ratios are not significantly different from expected ratios.
Male fertility assay
Five males of each genotype (w/Y; Df(2L)BSC354/CyO control, w/Y; eyacliIID/Df(2L)BSC354; eya+GR/+, w/Y; eyacliIID/Df(2L)BSC354; eyaSAGR/+, and w/Y; eyacliIID/Df(2L)BSC354; eyaSDEGR/+) were crossed to ten w1118 virgin females in triplicate. The flies were allowed to lay eggs for three days before being removed. We counted progeny that eclosed between days 9 and 16 after setting the cross, and the results were analyzed with one-way ANOVA.
Acknowledgments
We thank Umesh Karandikar, Matthew Rose, Georg Halder, Hamed Jafar-Nejad and Susan Rose for comments on the manuscript, and Gabriela David and Manish Jaiswal for assistance with Drosophila brain dissections.
Funding Statement
This study was supported by the Retina Research Foundation (http://retinaresearchfnd.org/), the National Eye Institute (www.nei.nih.gov) grant R01 EY011232 and the NEI/NIH Core Grant for Vision Research EY-002520. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Bonini NM, Leiserson WM, Benzer S (1993) The eyes absent gene: genetic control of cell survival and differentiation in the developing Drosophila eye. Cell 72: 379–395. [DOI] [PubMed] [Google Scholar]
- 2. Silver SJ, Davies EL, Doyon L, Rebay I (2003) Functional dissection of eyes absent reveals new modes of regulation within the retinal determination gene network. Mol Cell Biol 23: 5989–5999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Rayapureddi JP, Kattamuri C, Steinmetz BD, Frankfort BJ, Ostrin EJ, et al. (2003) Eyes absent represents a class of protein tyrosine phosphatases. Nature 426: 295–298 doi:10.1038/nature02093. [DOI] [PubMed] [Google Scholar]
- 4. Tootle TL, Silver SJ, Davies EL, Newman V, Latek RR, et al. (2003) The transcription factor Eyes absent is a protein tyrosine phosphatase. Nature 426: 299–302 doi:10.1038/nature02097. [DOI] [PubMed] [Google Scholar]
- 5. Abdelhak S, Kalatzis V, Heilig R, Compain S, Samson D, et al. (1997) A human homologue of the Drosophila eyes absent gene underlies branchio-oto-renal (BOR) syndrome and identifies a novel gene family. Nat Genet 15: 157–164 doi:10.1038/ng0297-157. [DOI] [PubMed] [Google Scholar]
- 6. Zhang L, Yang N, Huang J, Buckanovich RJ, Liang S, et al. (2005) Transcriptional coactivator Drosophila eyes absent homologue 2 is up-regulated in epithelial ovarian cancer and promotes tumor growth. Cancer Res 65: 925–932. [PubMed] [Google Scholar]
- 7. Pandey RN, Rani R, Yeo E-J, Spencer M, Hu S, et al. (2010) The Eyes Absent phosphatase-transactivator proteins promote proliferation, transformation, migration, and invasion of tumor cells. Oncogene 29: 3715–3722 doi:10.1038/onc.2010.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Miller SJ, Lan ZD, Hardiman A, Wu J, Kordich JJ, et al. (2010) Inhibition of Eyes Absent Homolog 4 expression induces malignant peripheral nerve sheath tumor necrosis. Oncogene 29: 368–379 doi:10.1038/onc.2009.360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Robin TP, Smith A, McKinsey E, Reaves L, Jedlicka P, et al. (2012) EWS/FLI1 regulates EYA3 in Ewing's sarcoma via modulation of microRNA-708, resulting in increased cell survival and chemoresistance. Molecular cancer research: MCR. Available: http://www.ncbi.nlm.nih.gov/pubmed/22723308. Accessed 13 July 2012. [DOI] [PMC free article] [PubMed]
- 10. Nüsslein-Volhard C, Wieschaus E, Kluding H (1984) Mutations affecting the pattern of the larval cuticle inDrosophila melanogaster. Wilhelm Roux's Archives of Developmental Biology 193: 267–282 doi:10.1007/BF00848156. [DOI] [PubMed] [Google Scholar]
- 11. Boyle M, Bonini N, DiNardo S (1997) Expression and function of clift in the development of somatic gonadal precursors within the Drosophila mesoderm. Development 124: 971–982. [DOI] [PubMed] [Google Scholar]
- 12. Bonini NM, Leiserson WM, Benzer S (1998) Multiple roles of the eyes absent gene in Drosophila. Dev Biol 196: 42–57 doi:10.1006/dbio.1997.8845. [DOI] [PubMed] [Google Scholar]
- 13. Liu Y-H, Jakobsen JS, Valentin G, Amarantos I, Gilmour DT, et al. (2009) A systematic analysis of Tinman function reveals Eya and JAK-STAT signaling as essential regulators of muscle development. Dev Cell 16: 280–291 doi:10.1016/j.devcel.2009.01.006. [DOI] [PubMed] [Google Scholar]
- 14. Ready DF, Hanson TE, Benzer S (1976) Development of the Drosophila retina, a neurocrystalline lattice. Dev Biol 53: 217–240. [DOI] [PubMed] [Google Scholar]
- 15. Pignoni F, Hu B, Zavitz KH, Xiao J, Garrity PA, et al. (1997) The eye-specification proteins So and Eya form a complex and regulate multiple steps in Drosophila eye development. Cell 91: 881–891. [DOI] [PubMed] [Google Scholar]
- 16. Bonini NM, Bui QT, Gray-Board GL, Warrick JM (1997) The Drosophila eyes absent gene directs ectopic eye formation in a pathway conserved between flies and vertebrates. Development 124: 4819–4826. [DOI] [PubMed] [Google Scholar]
- 17. Halder G, Callaerts P, Gehring WJ (1995) Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science 267: 1788–1792. [DOI] [PubMed] [Google Scholar]
- 18. Weasner B, Salzer C, Kumar JP (2007) Sine oculis, a member of the SIX family of transcription factors, directs eye formation. Dev Biol 303: 756–771 doi:10.1016/j.ydbio.2006.10.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Shen W, Mardon G (1997) Ectopic eye development in Drosophila induced by directed dachshund expression. Development 124: 45–52. [DOI] [PubMed] [Google Scholar]
- 20. Hazelett DJ, Bourouis M, Walldorf U, Treisman JE (1998) decapentaplegic and wingless are regulated by eyes absent and eyegone and interact to direct the pattern of retinal differentiation in the eye disc. Development 125: 3741–3751. [DOI] [PubMed] [Google Scholar]
- 21. Chen R, Halder G, Zhang Z, Mardon G (1999) Signaling by the TGF-beta homolog decapentaplegic functions reiteratively within the network of genes controlling retinal cell fate determination in Drosophila. Development 126: 935–943. [DOI] [PubMed] [Google Scholar]
- 22. Curtiss J, Mlodzik M (2000) Morphogenetic furrow initiation and progression during eye development in Drosophila: the roles of decapentaplegic, hedgehog and eyes absent. Development 127: 1325–1336. [DOI] [PubMed] [Google Scholar]
- 23. Bessa J, Gebelein B, Pichaud F, Casares F, Mann RS (2002) Combinatorial control of Drosophila eye development by eyeless, homothorax, and teashirt. Genes Dev 16: 2415–2427 doi:10.1101/gad.1009002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Firth LC, Baker NE (2009) Retinal determination genes as targets and possible effectors of extracellular signals. Dev Biol 327: 366–375 doi:10.1016/j.ydbio.2008.12.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Czerny T, Halder G, Kloter U, Souabni A, Gehring WJ, et al. (1999) twin of eyeless, a second Pax-6 gene of Drosophila, acts upstream of eyeless in the control of eye development. Mol Cell 3: 297–307. [DOI] [PubMed] [Google Scholar]
- 26. Halder G, Callaerts P, Flister S, Walldorf U, Kloter U, et al. (1998) Eyeless initiates the expression of both sine oculis and eyes absent during Drosophila compound eye development. Development 125: 2181–2191. [DOI] [PubMed] [Google Scholar]
- 27. Niimi T, Seimiya M, Kloter U, Flister S, Gehring WJ (1999) Direct regulatory interaction of the eyeless protein with an eye-specific enhancer in the sine oculis gene during eye induction in Drosophila. Development 126: 2253–2260. [DOI] [PubMed] [Google Scholar]
- 28. Pauli T, Seimiya M, Blanco J, Gehring WJ (2005) Identification of functional sine oculis motifs in the autoregulatory element of its own gene, in the eyeless enhancer and in the signalling gene hedgehog. Development 132: 2771–2782 doi:10.1242/dev.01841. [DOI] [PubMed] [Google Scholar]
- 29. Pappu KS, Ostrin EJ, Middlebrooks BW, Sili BT, Chen R, et al. (2005) Dual regulation and redundant function of two eye-specific enhancers of the Drosophila retinal determination gene dachshund. Development 132: 2895–2905 doi:10.1242/dev.01869. [DOI] [PubMed] [Google Scholar]
- 30. Clark-Lewis I, Sanghera JS, Pelech SL (1991) Definition of a consensus sequence for peptide substrate recognition by p44mpk, the meiosis-activated myelin basic protein kinase. J Biol Chem 266: 15180–15184. [PubMed] [Google Scholar]
- 31. Hsiao FC, Williams A, Davies EL, Rebay I (2001) Eyes absent mediates cross-talk between retinal determination genes and the receptor tyrosine kinase signaling pathway. Dev Cell 1: 51–61. [DOI] [PubMed] [Google Scholar]
- 32. Morillo SA, Braid LR, Verheyen EM, Rebay I (2012) Nemo phosphorylates Eyes absent and enhances output from the Eya-Sine oculis transcriptional complex during Drosophila retinal determination. Dev Biol 365: 267–276 doi:10.1016/j.ydbio.2012.02.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Braid LR, Verheyen EM (2008) Drosophila nemo promotes eye specification directed by the retinal determination gene network. Genetics 180: 283–299 doi:10.1534/genetics.108.092155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Brand AH, Perrimon N (1993) Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118: 401–415. [DOI] [PubMed] [Google Scholar]
- 35. Chen R, Amoui M, Zhang Z, Mardon G (1997) Dachshund and eyes absent proteins form a complex and function synergistically to induce ectopic eye development in Drosophila. Cell 91: 893–903. [DOI] [PubMed] [Google Scholar]
- 36. Kango-Singh M, Singh A, Henry Sun Y (2003) Eyeless collaborates with Hedgehog and Decapentaplegic signaling in Drosophila eye induction. Dev Biol 256: 49–60. [DOI] [PubMed] [Google Scholar]
- 37. Mutsuddi M, Chaffee B, Cassidy J, Silver SJ, Tootle TL, et al. (2005) Using Drosophila to decipher how mutations associated with human branchio-oto-renal syndrome and optical defects compromise the protein tyrosine phosphatase and transcriptional functions of eyes absent. Genetics 170: 687–695 doi:10.1534/genetics.104.039156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Weasner BM, Weasner B, Deyoung SM, Michaels SD, Kumar JP (2009) Transcriptional activities of the Pax6 gene eyeless regulate tissue specificity of ectopic eye formation in Drosophila. Dev Biol 334: 492–502 doi:10.1016/j.ydbio.2009.04.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Venken KJT, He Y, Hoskins RA, Bellen HJ (2006) P[acman]: a BAC transgenic platform for targeted insertion of large DNA fragments in D. melanogaster. Science 314: 1747–1751 doi:10.1126/science.1134426. [DOI] [PubMed] [Google Scholar]
- 40. Zimmerman JE, Bui QT, Liu H, Bonini NM (2000) Molecular genetic analysis of Drosophila eyes absent mutants reveals an eye enhancer element. Genetics 154: 237–246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Parks AL, Cook KR, Belvin M, Dompe NA, Fawcett R, et al. (2004) Systematic generation of high-resolution deletion coverage of the Drosophila melanogaster genome. Nat Genet 36: 288–292 doi:10.1038/ng1312. [DOI] [PubMed] [Google Scholar]
- 42. Xiong W, Dabbouseh NM, Rebay I (2009) Interactions with the abelson tyrosine kinase reveal compartmentalization of eyes absent function between nucleus and cytoplasm. Dev Cell 16: 271–279 doi:10.1016/j.devcel.2008.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Fabrizio JJ, Boyle M, DiNardo S (2003) A somatic role for eyes absent (eya) and sine oculis (so) in Drosophila spermatocyte development. Dev Biol 258: 117–128. [DOI] [PubMed] [Google Scholar]
- 44. Ahmed M, Wong EYM, Sun J, Xu J, Wang F, et al. (2012) Eya1-Six1 interaction is sufficient to induce hair cell fate in the cochlea by activating Atoh1 expression in cooperation with Sox2. Dev Cell 22: 377–390 doi:10.1016/j.devcel.2011.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Salzer CL, Kumar JP (2010) Identification of retinal transformation hot spots in developing Drosophila epithelia. PLoS ONE 5: e8510 doi:10.1371/journal.pone.0008510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Hadorn E (1965) Problems of determination and transdetermination. Brookhaven Symp Biol 18: 148–161. [Google Scholar]
- 47. Maves L, Schubiger G (1995) Wingless induces transdetermination in developing Drosophila imaginal discs. Development 121: 1263–1272. [DOI] [PubMed] [Google Scholar]
- 48. Cheyette BN, Green PJ, Martin K, Garren H, Hartenstein V, et al. (1994) The Drosophila sine oculis locus encodes a homeodomain-containing protein required for the development of the entire visual system. Neuron 12: 977–996. [DOI] [PubMed] [Google Scholar]
- 49. Anderson AM, Weasner BM, Weasner BP, Kumar JP (2012) Dual transcriptional activities of SIX proteins define their roles in normal and ectopic eye development. Development 139: 991–1000 doi:10.1242/dev.077255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Rebay I, Silver SJ, Tootle TL (2005) New vision from Eyes absent: transcription factors as enzymes. Trends Genet 21: 163–171 doi:10.1016/j.tig.2005.01.005. [DOI] [PubMed] [Google Scholar]
- 51. Rebay I, Chen F, Hsiao F, Kolodziej PA, Kuang BH, et al. (2000) A genetic screen for novel components of the Ras/Mitogen-activated protein kinase signaling pathway that interact with the yan gene of Drosophila identifies split ends, a new RNA recognition motif-containing protein. Genetics 154: 695–712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Doroquez DB, Rebay I (2006) Signal integration during development: mechanisms of EGFR and Notch pathway function and cross-talk. Crit Rev Biochem Mol Biol 41: 339–385 doi:10.1080/10409230600914344. [DOI] [PubMed] [Google Scholar]
- 53. Salzer CL, Elias Y, Kumar JP (2010) The retinal determination gene eyes absent is regulated by the EGF receptor pathway throughout development in Drosophila. Genetics 184: 185–197 doi:10.1534/genetics.109.110122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Groth AC, Fish M, Nusse R, Calos MP (2004) Construction of transgenic Drosophila by using the site-specific integrase from phage phiC31. Genetics 166: 1775–1782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Thomason L, Court DL, Bubunenko M, Costantino N, Wilson H, et al. (2007) Recombineering: genetic engineering in bacteria using homologous recombination. Curr Protoc Mol Biol Chapter 1: Unit 1.16 doi:10.1002/0471142727.mb0116s78. [DOI] [PubMed] [Google Scholar]
- 56. Tomlinson A, Ready DF (1987) Cell fate in the Drosophila ommatidium. Dev Biol 123: 264–275. [DOI] [PubMed] [Google Scholar]
- 57. Wu JS, Luo L (2006) A protocol for dissecting Drosophila melanogaster brains for live imaging or immunostaining. Nat Protoc 1: 2110–2115 doi:10.1038/nprot.2006.336. [DOI] [PubMed] [Google Scholar]
- 58. Heisenberg M (1971) Separation of receptor and lamina potentials in the electroretinogram of normal and mutant Drosophila. J Exp Biol 55: 85–100. [DOI] [PubMed] [Google Scholar]