

Abstract
This video presentation was created to show a method of harvesting the two most important highly vascular structures, not residing within the brain proper, that support forebrain function. They are the cerebral surface (superficial) vasculature along with associated meninges (MAV) and the choroid plexus which are necessary for cerebral blood flow and cerebrospinal fluid (CSF) homeostasis. The tissue harvested is suitable for biochemical and physiological analysis, and the MAV has been shown to be sensitive to damage produced by amphetamine and hyperthermia 1,2. As well, the major and minor cerebral vasculatures harvested in MAV are of potentially high interest when investigating concussive types of head trauma. The MAV dissected in this presentation consists of the pial and some of the arachnoid membrane (less dura) of the meninges and the major and minor cerebral surface vasculature. The choroid plexus dissected is the structure that resides in the lateral ventricles as described by Oldfield and McKinley3,4,5,6. The methods used for harvesting these two tissues also facilitate the harvesting of regional cortical tissue devoid of meninges and larger cerebral surface vasculature, and is compatible with harvesting other brain tissues such as striatum, hypothalamus, hippocampus, etc. The dissection of the two tissues takes from 5 to 10 min total. The gene expression levels for the dissected MAV and choroid plexus, as shown and described in this presentation can be found at GSE23093 (MAV) and GSE29733 (choroid plexus) at the NCBI GEO repository. This data has been, and is being, used to help further understand the functioning of the MAV and choroid plexus and how neurotoxic events such as severe hyperthermia and AMPH adversely affect their function.
Keywords: Neuroscience, Issue 69, Medicine, Anatomy, Physiology, Toxicology, brain, dissection, choroid plexus, meninges and associated vasculature
Protocol
Although not shown in the video, rats were first given an overdose of 300 mg/kg of pentobarbital, resulting in anesthesia in less than 3 min, and then killed by decapitation. Their brains were then rapidly but carefully removed from the skull and chilled in ice-cold normal saline for 5 min. It is important to let the brain cool for this amount of time prior to dissecting the MAV so that the MAV can be separated from the surface of the cortex (top of layer I). The brain was then placed in the bottom of a glass Petri dish resting on ice that was full (1 cm deep) of ice-cold normal saline or 0.1 M sodium phosphate-buffered saline pH=7.4. All the dissection is done with the brain for the most part immersed in saline.
1. Removal of the Pineal Gland
Place the brain dorsal side up (relative to the bottom of the Petri dish). The pineal gland is located at the most caudal ventral region between the two hemispheres just rostral to the cerebellum (pineal recess), and is just below or at the surface of the meninges over the colliculi. Remove the pineal gland first using two small bent tip forceps. It is pink in color like the cortex but is often surrounded in remnants of blood and vasculature residual from brain removal.
2. Removal of the MAV
Flip the brain over "upside down" and separate the vertebral artery, smaller arteries and meninges covering the pons from the meninges and vasculature of the forebrain. Removal of the forebrain MAV is then started. It is important to start ventrally in the dissection process. The large major arteries of the circle of Willis, middle cerebral arteries (MCA) and anterior arteries are the strongest and serve as "scaffolding" by which the smaller superficial arteries and arterioles of the pial membrane can then be removed from the cortex with the remainder of the meninges. See review by Scremin7 for more visual representations and details of the cerebral surface vasculature.
Starting with either hemisphere, use the small bent tip forceps to sever the posterior communicating artery just posterior to the internal carotid juncture. Thus, separating the more anterior and posterior arterial trees of the forebrain. Repeat the same process for the contralateral hemisphere.
Gripping the MCA and the anterior artery with the forceps, pull gently in an anterior-dorsal direction so that the MAV covering the ventral anterior cortex (piriform, olfactory tubercle) is lifted over and around the olfactory tract. This removes much of the MAV covering the anterior cortex (cingulated and orbital).
Turn the brain at a 45 to 90° angle to the Petri dish. The lateral, more anterior regions of the MAV surrounding the lateral cortical regions (granular insular, secondary somatosensory and auditory cortex) can be freed from the cortex by gripping the MCA and associated arteries and gently pulling dorsally along the cortex. If necessary, the ends of the forceps can be used to lift and free the major arteries from the cortex during dissection.
Now remove the more posterior lateral regions of the MAV. (Note, it is best to switch from one hemisphere to the other during this harvesting process to ensure that the MAV remains cold and hydrated. Also, if resistance in freeing the MAV from the cortex is encountered on one hemisphere switch to the contralateral hemisphere temporarily and then go back to the original hemisphere later.)
To remove the most dorsal regions of the MAV surrounding the primary somatosensory and motor cortex turn the brain dorsal side up (relative to the bottom of the Petri dish). To finally free the MAV from brain, use the ends of the forceps along the sagital sinus. This portion of the harvested MAV can be either kept temporarily in ice cold saline until testing and analysis or frozen for later processing. Often the MAV covering the most posterior/caudal regions of the cortex (entorhinal, visual and most caudal auditory cortices) remains attached to the cortex. Its removal is shown later in the video when the MAV between the retrosplenial cortex and overlying the colliculi are dissected.
3. Removal of the Choroid Plexus
Position the brain dorsal side up and hold in place with the larger forceps Push the smaller forceps down through the midline between the hemispheres and then use the ends to puncture through the cortex and corpus callosum (≈-3.3 mm from bregma) into the top of the midline of the hippocampus.
Use the forceps to pull the cortex with callosum away from the dorsal hippocampus and septum, exposing most of the lateral ventricle. The choroid plexus can be located and identified by the wavy red line demarcating the major artery that runs its length. Use the two ends of the forceps to pry the lateral walls of the third ventricle and enlarge it at the most caudal end (≈-4.3 mm from bregma8).
Using the small forceps, pull this end of the choroid plexus free. Then go to the very anterior region between the septum and caudate/putamen (most rostral extent, ≈+1.6 mm from Bregma) and with the forceps enlarge this section of the ventricle and pull the choroid plexus free. Perform the same procedure on the contralateral hemisphere to obtain the second remaining choroid plexus. Again this tissue can be either kept temporarily in ice cold saline until testing and analysis or frozen for later processing.
4. Removal of the Remaining MAV in the 3rd Ventricle Overlying the Thalamus and the Colliculi as well as that on the more Caudal Regions of the Cortex including Retrosplenial, Auditory and Visual Cortex
Remove the entire hippocampus from both hemispheres exposing the MAV over the thalamus and colliculi. The removal of this portion of the MAV is much less difficult than the removal of MAV over the cortex.
Pull this portion of the MAV free by grasping the larger vasculature overlying the anterior portion of the thalamus and gently pulling caudally. This vasculature is connected to the supra collicular network and supplies blood to the dorsal hippocampus and dorsal thalamus network. Pull it away caudally and gradually free the supracollicular network until reaching the last portions of the caudal arterial circle vasculature. At this point, more care must be taken to remove any remaining posterior ventral MAV on the surface of the granular retrosplenial, primary auditory and visual cortices. As with the other section of MAV covering the more anterior portions of the cortex, this tissue can be either kept temporarily in ice cold saline until testing and analysis or frozen for later processing. Any remaining posterior ventral MAV harvested with this second MAV section should be removed and added to the first dissected MAV section covering the cortex if they are to be analyzed separately.
5. Representative Results
When the dissection is executed correctly, the resulting MAV tissues should be in two intact entities weighing about 35 to 45 mg total. The initially dissected MAV surrounding most of the cortex should weigh about 25 mg and the MAV covering the thalamus, colliculi and occipital cortex should weigh about 15 mg. It should be slightly pinkish in color from the residual blood in the vasculature. The bilateral choroid plexus harvested should be in two intact tissue samples with each weighing 1 to 2 mg. Although excess water can be removed from the MAV prior to storage or processing using tissue paper such as that used in lens cleaning, to avoid tissue loss it is not a good idea to remove excess water from the dissected choroid plexus. Figure 1 was generated for comparing expression profiles in three regions (striatum, parietal cortex and MAV) under control conditions using Agilent-014879 Whole Rat Genome 4x44K 60mer oligonucleotide arrays (G4131F, Agilent Technologies, Palo Alto, CA; NCBI GEO Accession # GPL7294; GSE23093 and GSE29733 that are present at the NCBI GEO repository). The two plots in the figure pictorially show the expression in 1) the striatum compared to the parietal cortex and 2) the MAV compared to the parietal cortex. It is clear that the striatum and parietal cortex are much more closely related to each other than the MAV. The data comparing the choroid plexus with the MAV will be presented in a future publication. However, it is likely that the expression profiles in the MAV and choroid plexus are more closely related to each other than they are to the striatum and parietal cortex. Figure 2 shows the differential gene expression response to hyperthermia (EIH) versus AMPH in MAV compared to control and each other.
The gene expression of more than 11,000 genes with official NCBI gene symbols in MAV of control animals was compared to those recorded in the striatum and parietal cortex. Over 2000 of these genes had a 2.5-fold or more differing expression in the MAV compared to the two neuronal-rich tissues, and 550 genes had a 10-fold greater difference in expression. Slightly more than 40% (253) of these were genes with a decreased expression of 10-fold or more in MAV and were related to neuronal function. There were 343 genes with over a 10-fold or greater expression in MAV compared to parietal cortex and striatum. This list of genes was used as a starting point with which to determine genes with a very high enrichment in the MAV. Table 1 lists the genes with greater than 15-fold expression in MAV compared to parietal cortex and striatum and categorizes them with respect to function. Many of these genes were related to the vascular system and/or the immune system. These types of genes should be enriched about 5- to 10- fold to because of the increased vasculature itself and blood present within it. However, even for genes with known general vascular functions, an increase above 15-fold indicates enrichment in MAV. There were also numbers of genes with very large fold changes that were; 1) extracellular matrix proteins (not specifically related to vasculature), 2) solute transporters and 3) lipid and retinoic acid metabolism. As well, few growth and differentiation gene or transcription factors were found.
 Figure 1. Comparison of the Gene Expression in MAV with Parietal Cortex and Striatum. Volcano plot comparing the gene expression levels of the parietal cortex and striatum (top plot) and the parietal cortex and MAV (bottom plot). The red symbols denote the genes that are significantly different between regions at p <0.05 and have a 1.5-fold or greater differential expression between regions. The black symbols represent genes which do not significantly differ between regions (p >0.05) and are less than 1.5-fold differing in expression. The pink symbols indicate genes with less than a 1.5-fold difference in expression but statistically significantly different at p<0.05. The yellow symbols identify genes which have a 1.5-fold or greater expression but this change is not statistically significant at p<0.05.
Figure 1. Comparison of the Gene Expression in MAV with Parietal Cortex and Striatum. Volcano plot comparing the gene expression levels of the parietal cortex and striatum (top plot) and the parietal cortex and MAV (bottom plot). The red symbols denote the genes that are significantly different between regions at p <0.05 and have a 1.5-fold or greater differential expression between regions. The black symbols represent genes which do not significantly differ between regions (p >0.05) and are less than 1.5-fold differing in expression. The pink symbols indicate genes with less than a 1.5-fold difference in expression but statistically significantly different at p<0.05. The yellow symbols identify genes which have a 1.5-fold or greater expression but this change is not statistically significant at p<0.05.
Abbreviations
AMPH amphetamine EIH environmentally-induced hyperthermia MAV meninges and associated cerebral vasculature Norm normothermic controls
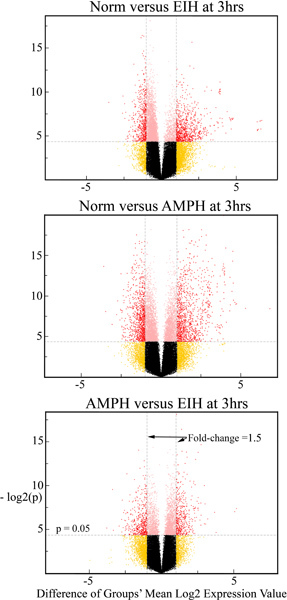 Figure 2. Effects of hyperthermia and amphetamine on gene expression in MAV. Volcano plots comparing gene expression levels in the MAV after saline, EIH or AMPH at the 3 hr time point. The red symbols denote the genes that are deemed to be significantly different while the black dots/circles represent genes which do not significantly differ between regions. Additional statistical analysis was used to verify which expression differences between control and treatments were in fact statistically significant.
Figure 2. Effects of hyperthermia and amphetamine on gene expression in MAV. Volcano plots comparing gene expression levels in the MAV after saline, EIH or AMPH at the 3 hr time point. The red symbols denote the genes that are deemed to be significantly different while the black dots/circles represent genes which do not significantly differ between regions. Additional statistical analysis was used to verify which expression differences between control and treatments were in fact statistically significant.
| MAV Expression Brain Expression | NCBI Gene Symbols | Tissue Specificity or Reported Function(s) |
| > 50-fold | Anxa2a, Col8a1, Des, Esm1, Glycam1, Kla, Lect1, Lepr, Lum, Myh11, Ogn, Tagln, Thbs2, Tnnt2 | Vasculature & Heart |
| > 50-fold | Ccl19, Cd74a, Defb1, Lrrc21, Mgl1, Mrc1, Msln, Pla2g5, Prg4, RT1-Bb, RT1-Da | Immune System |
| > 50-fold | Adh1, Aebp1, Aldh1a2, Gstm2 | Retinoic Acid & Lipid Processing |
| > 50-fold | Cdh1, Col3a1, Col1a2, Colec12, Cpxm2, Cpz, Dcn, Emp3, Gpc3, Nupr1, Omd, Pcolce Slamf9, Tmem27, Tspan8 | Extracellular Matrix & cell-cell junctions |
| > 50-fold | Aqp1, Asgr1, Kcnj13, Slc5a5, Slc6a13, Slc6a20, Slc22a6, Sned1, Ttr | Ion & Solute Homeostasis |
| > 50-fold | Alx3, Cdkn1c, Foxc2, Igfbp2, Ifitm1, Ifitm2, Nkx6-1, Osr1, Prrx2, Sfrp1, Tbx15. Upk1b, Wisp2, Wnt6 | Development & Transcription Regulation |
| > 50-fold | Gpha2, Mfap5, Mpzl2 Plac8, Scgb1c1, Sostdc1, Steap1 | Unknown & Miscellaneous |
| 30 to 50-fold | Anxa1a , Angpt2, C6a, Gjb2, Ptgis, Thbd, Timp1 | Vasculature & Heart |
| 30 to 50-fold | Casp12, Ccl2, Ccr1, Cd14, Cxcl10, Ifitm3, Klra5, Lgals1, Lgals3, Ms4a4a, Ms4a7, Plscr1, Spp1, Xcl1 | Immune System |
| 30 to 50-fold | Col6a3, Cpxm1, Efemp1, Fmod, Mgp, Nid2 | Extracellular Matrix & cell-cell junctions |
| 30 to 50-fold | Cp, Cubn, Gcgr, S100a6, Scn7a, Sct, Slc4a5, Slc16a11, Slco1a5 | Ion & Solute homeostasis |
| 30 to 50-fold | Cfd, Ch25h, Crabp2, Rarres2 | Retinoic Acid & Lipid Processing |
| 30 to 50-fold | Bmp6, Casp12, Dab2, Folr1, Igf2, Msx1, Tbx18, Twist1, Wnt5b | Development & Transcription Regulation |
| 30 to 50-fold | Cela3b, Cln6, C1qtnf7, Copz2, Dhrs7c, Gng11, Prss23, Srpx, Vim | Unknown & Miscellaneous |
| 15 to 30-fold | Adm, Angpt1, Angptl2, Bgn, Cklfa, Cnn1,Cox8h, Ctsk, F13a1, Gja5, Klf4, Lox, Lyz, Myl9, Procr, Pros1, RT1-Ba, Serpinb10, Serping1, Serpinf1, Tgm2, Tnmd, Trim63, Txnipa, Vamp5, Vtn | Vasculature & Heart |
| 15 to 30-fold | Adaa, Bst2, Ccl6, Cd40, Cd68, Ctsc, Dap, Faim3, Fkbp9, Fxyd5, Fmo1, Glipr1, Ier3, Ifi47, Igsf6, Msc, Nfatc4, RT1-Db1, Serpinb1a, Tir4, Tubb6 | Immune System |
| 15 to 30-fold | Adamtsl4, Col1a1, Crb3, Dpt, Egfl3, Fbn1, Fbln1, Fbln5, Fn1, Itgb4, Lama2, Lgals3bp, Loxl1, Mfap4, Mmp14, Mmp23, Ppic, Serpinh1, Timp3 | Extracellular Matrix & cell-cell junctions |
| 15 to 30-fold | Cybrd1, Selenbp1, Slc2a4, Slc9a2, Slc13a3, Slc16a4, Slc22a8, Slc22a18 | Ion & Solute homeostasis |
| 15 to 30-fold | Agpat2, Bdh2, Cyp26b1, Lpar3, Ltb4dh, Olr1, Pon3, Rbp1, Rbp4 | Retinoic Acid & Lipid Processing |
| 15 to 30-fold | Atf3, Dkk4, Eya2, Ifitm7, Ltbp1, Mustn1, Nr2f2, Ptrf, Sphk1, Tgfbi, Tcea3 Tcfap2b, Wnt2b, Wnt5a, Zic1 | Development & Transcription Regulation |
| 15 to 30-fold | Adamts12, Adamtsl3, C1r, Crispld2, Crygn, Cyp1b1, Dse, Enpep, Enpp2, Epn3, Flna, Fmo3, Gnmt, Gprc5c, Hspb1, Klc3, Krt18, Krt19, Mdk, Mesp1, Ms4a6a, Ms4a6b, Ms4a11, Net1, Pdlim2, Phactr2, Plp2, Ppp1r3b, Pqlc3, Rin3, Spag11, Sult1a1, Tmem106a, Wfikkn2 | Unknown & Miscellaneous |
*Genes included in the table must be both more than 15-fold above expression in the striatum and parietal cortex and 5-fold above the background levels. The lower of the two ratios (MAV/striatum or MAV/parietal cortex) for each gene was used for grouping.
aGenes are found in endothelial cells but also likely play a major role in mediating immune responses.
Table 1. Genes with a 15-fold* or more increased expression in MAV compared to striatum and parietal cortex.
Discussion
Technical aspects of MAV and choroid plexus dissection
It is critical during the dissection of the MAV to have the brain "submerged" most of the time in the normal saline or sodium phosphate-buffered saline. Allowing the MAV and cortex to dehydrate during dissection will result in the MAV adhering tightly to cortical layer I. This will increase the amount of cortical tissue harvested with the MAV and/or result in the inability to remove these adhering MAV sections from the cortex. Also, patience is necessary during dissection. Again, if one encounters some resistance in removing the MAV from the cortex in one hemisphere during dissection, switch to the other hemisphere and go back later to the side where resistance was encountered. This method does take some practice to perfect and requires about 5 to 15 "practice" dissections before a consistently uniform harvest of MAV tissue is achieved. The method of starting the dissection and removal of the MAV at the arterial circle at the base of the brain facilitates the removal of the meninges by using the larger vasculature as support "scaffolding."
Much of the blood originally in the MAV vasculature after sacrifice should be ejected during the dissection, and less than 5% of the RNA harvested from MAV should be of blood origin. It is possible to remove almost all the blood by perfusing the animal with 50 to 70 ml of saline, using histological techniques, prior to decapitation and brain removal. However, it is then much more difficult to visualize the MAV tissues during dissection. The three most likely sources of "contamination" of the MAV tissues are the pineal gland, cortical layer I tissue and residual olfactory bulb tissue but it is also possible to get pituitary gland tissue in the preparation. Pineal contamination in the MAV is detected by the MAV containing a round 1 to 2 mm diameter sphere that is colored dark red. The primary function of the pineal gland is normally considered to be completely different from the MAV. Its primary function is to produce melatonin, which regulates aspects of the circadian rhythm9, and in lipoxygenation10 of lipid precursors. Although the gene Lox, which regulates lipoxygenase activity, was thought to be present in adults primarily in the pineal gland10, our results indicate that it is also consistently present at significant levels in MAV. Residual cortical or olfactory bulb contamination results in the pinkish web-like MAV tissue having some 1 to 2 mm denser tissues that are much paler in color enclosed in it. This can be spotted by "floating" the MAV on the surface of the buffer, and then removing the heavier cortical or bulb tissues using the two dissecting forceps.
If contamination of the MAV from cortical layer I tissue, hypothalamic tissue or the pineal gland is present, then several genes will have a significantly greater expression than what is normally present in a "clean" dissection. Contamination of the MAV with pineal gland will result in higher expression of arylalkylamine N-acetyltransferase (Aanat) which is necessary for melatonin synthesis. Unfortunately, if pineal contamination occurs during MAV dissection, the levels of Lox in MAV will not be able to be accurately determined. Contamination of the MAV with cortical tissue will result in higher expression of neuron-specific genes such as hippocalcin (Hpca), parvalbumin (Pvalb) and/or neuronal pentraxin receptor (Nptxr). Contamination of MAV with hypothalamic tissue will result in the higher expression of growth hormone 1 (Gh1), proopiomelanocortin (Pomc). In the event that some contamination does exist in the harvested tissues, most of these genes can be identified and not used in gene expression analysis. This is particularly important for genes present in the circulating blood that may still be present in MAV or choroid plexus.
Interpretting gene expression data from harvested MAV and choroid plexus
The cerebral surface (superficial) vasculature along with associated meninges (MAV) and the choroid plexus are necessary for cerebral blood flow and cerebrospinal fluid (CSF) homeostasis4,11 ,12 ,13 ,14 ,15 ,16 ,17 . The tissue harvest is suitable for biochemical and physiological analysis, and the MAV has been shown to be sensitive to damage produced by amphetamine and hyperthermia1,2 through gene expression analysis. These results are commensurate with what is also known about severe hyperthermia and exposure to amphetamines to the brain vasculature in general 18,19,20 . Human clinical data supports the belief that the subdural vasculature is sensitive to damage by amphetamine and methamphetamine 21,22 . As well, the major and minor cerebral vasculatures including those in the pial layer of the meninges that are harvested in MAV are of potentially high interest when investigating concussive types of head trauma 23,24 25 . The current gene expression profile yielded from the MAV indicates that the pial and possibly the arachnoid meningeal membranes may play a greater role than expected in regulating CSF composition. In the future, through the use of dissecting microscope techniques, it may be possible to further separate the tissues comprising the MAV so that the pial and arachnoid membranes can be separated from the larger arterial vasculature present and possibly from each other. This will enable a better interpretation of the function of each of these 3 components of the harvested MAV.
Disclosures
We have nothing to disclose. The work presented here may not necessarily represent the views of the Food and Drug Administration.
Acknowledgments
This work was funded by NCTR/FDA.
References
- Thomas M, George NI, Patterson TA, Bowyer JF. Amphetamine and environmentally induced hyperthermia differentially alter the expression of genes regulating vascular tone and angiogenesis in the meninges and associated vasculature. Synapse. 2009;63:881–894. doi: 10.1002/syn.20661. [DOI] [PubMed] [Google Scholar]
- Thomas M, et al. Endoplasmic reticulum stress responses differ in meninges and associated vasculature, striatum, and parietal cortex after a neurotoxic amphetamine exposure. Synapse. 2010;64:579–593. doi: 10.1002/syn.20763. [DOI] [PubMed] [Google Scholar]
- Ek CJ, Dziegielewska KM, Habgood MD, Saunders NR. Barriers in the developing brain and Neurotoxicology. Neurotoxicology. 2012. [DOI] [PubMed]
- Oldfield BJM, Michael J. In: The Rat Nervous System. Paxinos G, editor. Vol. 2. San Diego: Academic Press; 1995. p. 391. [Google Scholar]
- Johanson CE, Stopa EG, McMillan PN. The blood-cerebrospinal fluid barrier: structure and functional significance. Methods Mol. Biol. 2011;686:101–131. doi: 10.1007/978-1-60761-938-3_4. [DOI] [PubMed] [Google Scholar]
- Johanson CE. Multiplicity of cerebrospinal fluid functions: ? challenges in health and disease. Cerebrospinal Fluid Res. 2008;5 doi: 10.1186/1743-8454-5-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scremin OU. In: The Rat Nervous System. Paxinos G, editor. Vol. 2. San Diego: Academic Press; 1995. p. 3. [Google Scholar]
- Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. San Diego: Academic Press; 1995. [Google Scholar]
- Johnston JD. Photoperiodic regulation of prolactin secretion: changes in intra-pituitary signalling and lactotroph heterogeneity. J. Endocrinol. 2004;180:351–356. doi: 10.1677/joe.0.1800351. [DOI] [PubMed] [Google Scholar]
- Catala A. The function of very long chain polyunsaturated fatty acids in the pineal gland. Biochim. Biophys. Acta. 2010;1801:95–99. doi: 10.1016/j.bbalip.2009.10.010. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Neurogenic control of cerebral circulation. Cephalalgia. 1985;5(2):25–33. doi: 10.1177/03331024850050S205. [DOI] [PubMed] [Google Scholar]
- Hamel E. Perivascular nerves and the regulation of cerebrovascular tone. J. Appl. Physiol. 2006;100:1059–1064. doi: 10.1152/japplphysiol.00954.2005. [DOI] [PubMed] [Google Scholar]
- Johanson CE, et al. Cognitive function and nigrostriatal markers in abstinent methamphetamine abusers. Psychopharmacology (Berl) 2006;185:327–338. doi: 10.1007/s00213-006-0330-6. [DOI] [PubMed] [Google Scholar]
- Johnston M, Papaiconomou C. Cerebrospinal Fluid Transport: a Lymphatic Perspective. News Physiol. Sci. 2002;17:227–230. doi: 10.1152/nips.01400.2002. [DOI] [PubMed] [Google Scholar]
- Johnston M, Zakharov A, Koh L, Armstrong D. Subarachnoid injection of Microfil reveals connections between cerebrospinal fluid and nasal lymphatics in the non-human primate. Neuropathol. Appl. Neurobiol. 2005;31:632–640. doi: 10.1111/j.1365-2990.2005.00679.x. [DOI] [PubMed] [Google Scholar]
- Johnston M, et al. Evidence of connections between cerebrospinal fluid and nasal lymphatic vessels in humans, non-human primates and other mammalian species. Cerebrospinal Fluid Res. 2004;1:2. doi: 10.1186/1743-8454-1-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kulik T, et al. Regulation of cerebral vasculature in normal and ischemic brain. Neuropharmacol. 2008;55:281–288. doi: 10.1016/j.neuropharm.2008.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowyer JF, Ali SF. High doses of methamphetamine that cause disruption of the blood-brain barrier in limbic regions produce extensive neuronal degeneration in mouse hippocampus. Synapse. 2006;60:521–532. doi: 10.1002/syn.20324. [DOI] [PubMed] [Google Scholar]
- Sharma HS, Westman J, Nyberg F. Pathophysiology of brain edema and cell changes following hyperthermic brain injury. Prog. Brain Res. 1998;115:351–412. doi: 10.1016/s0079-6123(08)62043-9. [DOI] [PubMed] [Google Scholar]
- Sharma HS, Sjoquist PO, Ali SF. Drugs of abuse-induced hyperthermia, blood-brain barrier dysfunction and neurotoxicity: neuroprotective effects of a new antioxidant compound H-290/51. 2007;13:1903–1923. doi: 10.2174/138161207780858375. [DOI] [PubMed] [Google Scholar]
- Ho EL, Josephson SA, Lee HS, Smith WS. Cerebrovascular complications of methamphetamine abuse. Neurocrit. Care. 2009;10:295–305. doi: 10.1007/s12028-008-9177-5. [DOI] [PubMed] [Google Scholar]
- Ohta K, et al. Delayed ischemic stroke associated with methamphetamine use. J. Emerg. Med. 2005;28:165–167. doi: 10.1016/j.jemermed.2004.06.015. [DOI] [PubMed] [Google Scholar]
- Brenner LA. Neuropsychological and neuroimaging findings in traumatic brain injury and post-traumatic stress disorder. Dialogues Clin. Neurosci. 2011;13:311–323. doi: 10.31887/DCNS.2011.13.3/lbrenner. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Case ME. Inflicted traumatic brain injury in infants and young children. Brain Pathol. 2008;18:571–582. doi: 10.1111/j.1750-3639.2008.00204.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maas AI, et al. Prognostic value of computerized tomography scan characteristics in traumatic brain injury: results from the IMPACT study. J. Neurotrauma. 2007;24:303–314. doi: 10.1089/neu.2006.0033. [DOI] [PubMed] [Google Scholar]