

Abstract
The surgical resection of primary colorectal cancer is associated with an enhanced risk of liver metastases. Moreover, bacterial translocation or anastomic leakage during resection has been shown to correlate with a poor long-term surgical outcome, suggesting that bacterial products may contribute to the formation of metastases. Driven by these premises, we investigated the role of the bacterial product lipopolysaccharide (LPS) in the generation of liver metastases. Intraperitoneal injection of LPS led to enhanced tumor-cell adhesion to the rat liver as early as 1.5 h post-administration. Furthermore, a rapid loss of the expression of the tight junction protein zonula occludens-1 (ZO-1) was observed, suggesting that LPS disrupts the integrity of the microvasculature. LPS addition to endothelial-macrophage co-cultures damaged endothelial monolayers and caused the formation of intercellular gaps, which was accompanied by increased tumor-cell adhesion. These results suggest that macrophages are involved in the endothelial damage resulting from exposure to LPS. Interestingly, the expression levels of of ZO-1 were not affected by LPS treatment in rats in which liver macrophages had been depleted as well as in rats that had been treated with a reactive oxygen species (ROS) scavenger. In both settings, decreased tumor-cell adhesion was observed. Taken together, our findings indicate that LPS induces ROS release by macrophages, resulting in the damage of the vascular lining of the liver and hence allowing increased tumor-cell adherence. Thus, peri-operative treatments that prevent the activation of macrophages and—as a consequence—limit endothelial damage and tumor-cell adhesion may significantly improve the long-term outcome of cancer patients undergoing surgical tumor resection.
Keywords: endothelial damage, lipopolysaccharide, liver metastases, macrophages, surgery
Introduction
Colorectal cancer (CRC) is the most prevalent malignancy of the digestive tract in men and women in developed countries. CRC is diagnosed approximately one million times per year worldwide, and over half a million patients die from this disease annually.1-3 The surgical removal of primary CRC is the preferred therapeutic option and the only treatment that can provide long-term disease-free survival.4 However, 20–50% of the patients that do not have evidence of metastatic disease at the time of resection develop liver metastases within 5 y.5 This indicates that these patients either have undetectable micro-metastases or circulating tumor cells that can grow out at distant sites, even though surgery successfully removes the primary neoplastic lesion.
It has previously been demonstrated that surgery can paradoxically contribute to tumor recurrence and liver metastasis development.6-8 First, free circulating tumor cells have been detected in the peripheral blood of CRC patients prior to surgery.9 Moreover, the number of circulating tumor cells has been shown to increase during, or shortly after, resection in, both peripheral and portal blood, suggesting that the surgical handling of the tumor may result in tumor-cell shedding and dissemination.9,10 Second, previous studies have proved that surgery leads to the enhanced adhesion of tumor cells, which can proliferate and generate metastases, suggesting that surgical trauma augments metastasis outgrowth.7,8,11,12 Interestingly, it has also been shown that patients exhibiting bacterial translocation after surgical tumor resection have significantly reduced disease-free survival.13 In line with this notion, the treatment of mice with bacterial endotoxins reportedly promoted the development of lung and liver metastases.14,15 These data suggest that bacterial spillage during surgery may contribute to the growth of metastases.
Bacterial products such as lipopolysaccharide (LPS, an important component of the outer membrane of Gram-negative bacteria) have been detected in the plasma of surgerized patients.16,17 The concentration of LPS in the peripheral blood was found to increase 1 h after surgery and to normalize 24 h later. Elevations in the circulating levels of LPS were accompanied by intestinal permeability, suggesting that the epithelial barrier was partially compromised upon surgery.18 Finally, the translocation of viable bacteria from the gut lumen to local mesenteric lymph nodes has been documented in CRC patients undergoing colectomy.13,19,20
It has recently been demonstrated that LPS is detected by human CRC cells via Toll-like receptor 4 (TLR4), leading to enhanced cell adhesion.14 In addition, LPS can activate potent immune responses through TLRs that are expressed by a wide variety of cells from the immune system, including macrophages.21 Previously, we have shown that macrophages can release reactive oxygen species (ROS) upon surgery.11 ROS have been reported to disrupt the endothelial barrier, leading to vascular permeability in tissues, as well as enhanced tumor-cell adhesion in the liver.22-24 Because macrophages express TLR4, we hypothesized that they may play a role in the pro-metastatic activity of LPS. In the present study, we investigated the effects of LPS-stimulated macrophages on endothelial integrity in macrophage-endothelial co-cultures. In addition, we determined the biological activity of LPS in the rat liver and its role in tumor-cell adhesion in vivo.
Results
LPS stimulates tumor-cell adhesion in the rat liver by disrupting organ integrity
To investigate the effects of LPS on tumor-cell adhesion in the liver, rats were intraperitoneally injected with PBS or LPS, followed by the inoculation of tumor cells into the portal circulation. The number of tumor cells persisting in the liver of rats that had been pre-treated with LPS was significantly increased 1.5 h after injection of LPS, as compared with the amount of tumor cells found in the liver of PBS-treated rats (Fig. 1A and B). Immunofluorescent staining of rat liver samples demonstrated that the administration of LPS results in decreased expression of the tight junction protein zonula occludens-1 (ZO-1; Figure 1C), pointing to a situation of endothelial stress and loss of liver microvascular integrity.

Figure 1. LPS stimulates tumor-cell adhesion in the liver by disrupting endothelial integrity. (A) Tumor cells in the livers of rats that had been treated with PBS or LPS and were sacrificed 1.5 h after tumor-cell inoculation. Red: DiI-labeled CC531s, blue: cell nuclei, arrowheads point to CC531s cells. (B) Quantification of tumor cells in the livers of rats after PBS or LPS treatment. n = 4 per group; *p < 0.05. (C) ZO-1 staining in the livers of rats that had been treated with PBS or LPS and were sacrificed after 1.5 h. Green: ZO-1; blue: nuclei.
Because our previous findings suggest that macrophages promote tumor-cell adhesion by disrupting sinus endothelial barriers,11 we studied the effects of LPS-stimulated macrophages on endothelial cells. Because macrophages and sinus endothelial cells are in close proximity to each other in the liver, we developed a model allowing us to study endothelial cells and macrophages in intimate contact (Fig. 2A). Confluent endothelial cell monolayers were cultured on the upper side of transwells membranes, and macrophages on the lower side. Neither the incubation of human umbilical vein endothelial cells (HUVECs) with 100 ng/mL LPS nor the co-culture of HUVECs with macrophages in the absence LPS affected endothelial integrity (Fig. 2B). However, the addition of 10 ng/mL LPS to HUVEC-macrophage co-cultures disrupted the integrity of endothelial monolayers and promoted the formation of intercellular gaps between endothelial cells, which were further increased when 100 or 500 ng/mL LPS was added. Addition of the LPS inhibitor polymixin B (PMB) significantly prevented endothelial cell damage (Fig. 2C). Next, we investigated tumor-cell adhesion after incubation of HUVEC-macrophage co-cultures with LPS. Intercellular gaps in the endothelial monolayers that were formed upon LPS administration, contained high numbers of adherent tumor cells (Fig. 2D, right panel). In contrast, no tumor cells attached to endothelial monolayers were detected in the absence of LPS (Fig. 2D, left panel).

Figure 2. LPS leads to endothelial damage and causes enhanced tumor-cell adhesion. (A) Schematic overview of the co-culture of HUVEC and macrophages (MΦ) in vitro. (B) HUVEC-macrophages co-cultures were incubated with different concentrations of LPS. Red: HUVECs fluorescently stained with Ulex-Rhodamin, blue: cell nuclei. (C) Quantification of damaged endothelial area (%). *p < 0.05; **p < 0.01; ***p < 0.001. (D) Tumor-cell adherence after exposure of HUVEC-MΦ co-cultures to control or LPS. Red: HUVECs, blue: cell nuclei, light blue: SW620 tumor cells. Dotted lines indicate damaged areas without endothelial cells.
Since these data reflected the activation of macrophages by LPS, we used scanning and transmission electron microscopy (SEM and TEM, respectively) to visualize the effects of LPS stimulation on macrophages. After 30 min incubation with PBS, macrophages showed some extent of adherence to the culture substrate (Fig. 3A, left panels). Conversely, macrophages that received LPS were tightly attached to the substrate and widely spread, pointing to a robust activation (Fig. 3A, right panels). Moreover, LPS-stimulated macrophages formed a continuous monolayer after 2 h, while their PBS-treated counterparts did not do so and persisted as non-confluent populations. Furthermore, PBS-treated macrophages contained no intracellular vacuoles and showed almost no filopodia (Fig. 3B, left panel), reflecting a reduced cellular activity. In contrast, LPS-stimulated macrophages contained several intracellular vacuoles and exhibited multiple filopodia, indicating a high cellular activity (Fig. 3B, right panel). Thus, incubation of macrophages with LPS caused their rapid activation. This is consistent with our in vivo data showing increased tumor-cell adhesion as early as 1.5 h after tumor-cell injection in rats that were pre-injected with LPS (Fig. 1A and B).
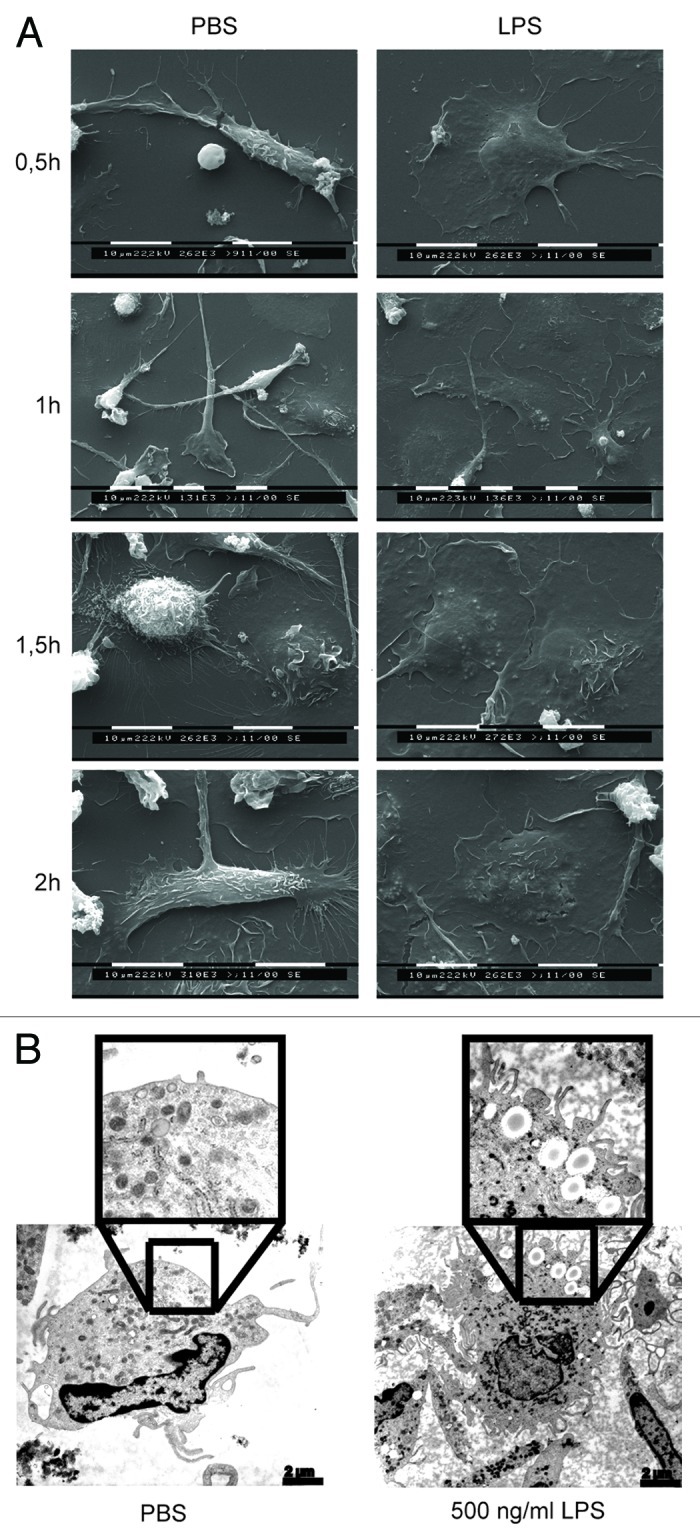
Figure 3. LPS stimulates rapid macrophage activation. (A) Human macrophages were incubated with either PBS or LPS and cells were analyzed by SEM at different time points. (B) Macrophages were treated with PBS or LPS for 2 h and then analyzed by TEM. Representative pictures are shown.
To investigate the role of macrophages in tumor-cell adherence in the rat liver upon in vivo LPS administration, we used control rats and rats in which liver macrophages (also known as Kupffer cells, KCs) had been depleted by injection of clodronate-containing liposomes.25 The absence of liver macrophages or newly recruited monocytes upon treatment clonodrate-bearing liposomes was confirmed by staining for ED2 (a marker for tissue-resident macrophages, data not shown) and ED1 (a marker of newly recruited monocytes, data not shown). All animals were treated with LPS and tumor cells were injected into the portal circulation. The number of tumor cells in the livers of KC-depleted rats that had been pre-treated with LPS was drastically decreased as compared with the number of tumor cells found in the livers of control rats pre-treated with LPS 1.5 h after tumor-cell inoculation (Fig. 4A and B). Moreover, higher expression levels of ZO-1 were observed in the livers of KC-depleted rats receiving with LPS, as compared with control rats similarly treated (Fig. 4C). Thus, macrophages appear to regulate the adherence of tumor cells in the livers of LPS-treated animals by disrupting organ integrity.

Figure 4. LPS-induced tumor-cell adhesion is Kupffer cell (KC)-dependent. (A) Tumor cells in the livers of control or KC-depleted rats that were treated with LPS. Red: DiI-labeled CC531s, blue: cell nuclei, arrow heads point to CC531s cells. (B) Quantification of tumor cells in the livers of control or KC-depleted rats that were treated with LPS. n = 4 per group; ***p < 0.001 (C) ZO-1 staining in the livers of control or KC-depleted rats that were treated with LPS. Green: ZO-1, blue: cell nuclei.
LPS stimulation of macrophages leads to damaged endothelial layer through ROS production
It has previously been shown that ROS induce endothelial damage.11 Therefore, we investigated whether the damage to endothelial monolayers in LPS-treated co-cultures of HUVECs and macrophages was mediated by ROS. The incubation of co-cultures with 100 ng/mL LPS resulted in an endothelial damage that was reduced in the presence of the ROS-scavenging enzymes superoxide dismutase (SOD) or catalase. Moreover, endothelial damage was virtually abolished when SOD and catalase were employed in combination (Fig. 5A and B).

Figure 5. LPS-induced damage is mediated by ROS. (A) Co-culture of HUVECs and macrophages (MΦ) were incubated with 100 ng/mL LPS in the presence or absence of catalase and/or SOD. Red: HUVECs, blue: cell nuclei. Dotted lines indicate areas without endothelial cells. (B) Quantification of damaged endothelial area (%). *p < 0.05; **p < 0.01.
Next, we investigated the role of ROS in LPS-induced tumor-cell adhesion in vivo by treating rats with the anti-oxidant edaravone, which is clinically used to treat ischemic stroke.26,27 Animals were treated with PBS, edaravone, LPS or LPS plus edaravone. The liver of rats that were treated with LPS contained a significantly higher number of tumor cells than the livers of PBS- or edaravone-treated rats (Fig. 6A and B). Importantly, significantly lower numbers of tumor cells were observed in livers of animals which were treated LPS plus edaravone, as compared with rats receiving LPS only. Furthermore, the expression levels of ZO-1 did not decreased in the livers of rats that had been treated with LPS plus edaravone (Fig. 6C). Altogether, these results indicate that LPS-induced tumor-cell adhesion in the liver is a ROS-dependent process.

Figure 6. LPS-induced tumor-cell adhesion in the liver is prevented by treatment with edaravone (EDA). (A) Tumor cells in the livers of rats that were treated with PBS, EDA, LPS or LPS plus EDA. Rats were sacrificed 1.5 h after tumor-cell inoculation. Red: DiI-labeled CC531s cells, blue: cell nuclei, arrow heads point to CC531s cells (B) Quantification of the tumor cells in the livers of rats that were treated with PBS, EDA, LPS or LPS plus EDA. n = 4 per group; **p < 0.01; ***p < 0.001. (C) ZO-1 staining in the livers of rats that were either treated with PBS, EDA, LPS or LPS plus EDA. Green: ZO-1, blue: cell nuclei.
Discussion
The surgical resection of primary tumors is the preferred therapeutic option for CRC patients and can provide long-term disease-free survival. However, surgical trauma has been paradoxically correlated with development of liver metastases.8 In addition, it has been demonstrated that exposure to LPS promotes the formation of lung and liver metastases.14,15 The resection of part of the colon may potentially be accompanied by bacterial contamination, as suggested by the detection of increased LPS concentrations in the portal and peripheral blood of patients subjected to colectomy. Moreover, patients exhibiting bacterial translocation from the gut lumen to mesenteric lymph nodes have a comparatively short disease-free survival,13 indicating that the exposure to bacterial products like LPS may lead to poor clinical outcome. In the current study, we demonstrated that LPS promotes tumor-cell adhesion in the livers of rats as early as 1.5 h after the inoculation of tumor cells via the portal vein. It has recently been shown that LPS can directly increase CRC cell adhesion via a TLR4-dependent mechanism.14 This promotes the functional activation of β1 integrin, which is an important adhesion molecule specific for the extracellular matrix (ECM). However, whereas β1 integrin appeared to be essential for the adhesion of CC531s CRC cells to damaged peritoneal surfaces,7 it was not required for adherence to the liver vasculature.8 This suggests that our findings cannot be explained by a direct effect of LPS on CRC cells.
Upon binding to TLR4 on the surface of cells from the immune system, LPS also operate as a potent activator of inflammatory and immune responses.21,28 The liver contains resident macrophages, which are in intimate contact with sinusoidal endothelial cells and capable of releasing ROS upon stimulation with LPS.29,30 In line with previous results,29,30 we also observed that LPS promote the activation of macrophages. Previously, inflammatory mediators such as LPS have been suggested to induce the M1 macrophage phenotype, corresponding to so-called “classically-activated” macrophages that are able to produce and release ROS.28 It is therefore likely that rats treated with LPS will develop macrophages with an M1-like phenotype. In our hands, the activation of macrophages by LPS resulted in damaged endothelial monolayers in vitro, potentially resulting in the exposure of the sub-endothelial ECM. Moreover, the addition of tumor cells to LPS-treated HUVEC-macrophage co-cultures demonstrated that tumor cells preferentially attach to intercellular gaps, rather to endothelial cells. Furthermore, we detected a decreased expression of ZO-1 in the livers of rats that had been treated with LPS. This indicates that LPS disrupts liver microvasculature. It has previously been shown that the exposure of the sub-endothelial ECM in the liver vasculature leads to increased tumor-cell adhesion.8 Therefore, we propose that LPS causes endothelial damage in vivo as well, allowing free circulating tumor cells to adhere to exposed ECM. This hypothesis is supported by the results of a previous study, demonstrating that an increased binding of polymorphonuclear cells (PMNs) to the liver microvasculature upon LPS treatment was due to an altered interaction with the ECM.31 As we also observed significant accumulation of PMNs in the livers of rats receiving LPS (data not shown), it is likely that both PMNs and tumor cells use similar mechanisms to adhere to the liver ECM exposed upon damage of the microvasculature. Importantly, the downregulation of endothelial tight junction molecules (reflecting a situation of endothelial cell stress) in liver samples of patients after surgery has been reported, indicating that the exposure of the sub-endothelial ECM after surgery can occur in patients as well.11 Furthermore, enhanced numbers of circulating tumor cells during surgery have been detected in CRC patients.9,10,32 Therefore, LPS-induced endothelial damage may promote the adherence of tumor cells in the liver and worsen disease outcome.
The damage to endothelial monolayers mediated by LPS was prevented by the addition of ROS-scavenging enzymes, indicating an essential role for ROS in this process. Both PMNs and macrophages can be activated by LPS and are potent ROS producers.21,33 Therefore, these cells may contribute to endothelial damage in vivo. The depletion of liver macrophages as well as the administration of the antioxidant edaravone drastically decreased tumor-cell adherence in the liver and preserved ZO-1 expression, demonstrating an essential role for ROS production by macrophages. Of note, an impairment of ROS-scavenging systems has been observed both in cancer patients and tumor-bearing mice. Catalase activity, which neutralizes H2O2, was decreased in patients bearing rectal, gastric, pancreatic and colic tumors as well as in leukocytes and livers of tumor-bearing mice.34,35 Thus, the production of ROS following surgery-associated bacterial translocation in cancer patients with an imbalanced ROS-neutralizing system can result in damaged endothelial lining of the liver. We confirmed that LPS is an important factor for the induction of ROS production by macrophages. However, other bacterial components may be involved in enhancing tumor-cell adhesion, since the TLR1/2 ligand Pam3CSK4 as well as the TLR5 ligand flagellin36 induced significant ROS production by macrophages (Fig. S1). The TLR2 ligand peptidoglycan from Staphylococcus aureus did not induce ROS production. Thus, surgery may lead to contamination with different TLR ligands that may be involved in enhanced tumor-cell adhesion.
Because we previously demonstrated that enhanced tumor-cell adherence promotes the development of liver metastases,8 we also investigated tumor development after LPS injection. In spite of increased tumor-cell adhesion, rats that received LPS developed less liver metastases as compared with PBS treated rats (data not shown). Previous studies demonstrated that LPS injection leads to tumor outgrowth from HT29 or 4T1 cells.14,15 However, HT29 cells were grown in immunocompromised mice and 4T1 cells are poorly immunogenic, whereas CC531s cells have been reported to induce immune responses.8,37-40 Since TLR ligands can induce antitumor immunity,41-43 we stained tumor samples for infiltrating CD8+ and CD4+ T cells as well as NK cells (data not shown). Tumors from the livers of rats that were treated with LPS contained significantly higher numbers of all these cell populations, suggesting that LPS stimulates immune responses against immunogenic CC531s cells, resulting in the inhibition of liver metastases. Alternatively, it is possible that the result of the study employing 4T1 mammary carcinoma cells were affected by endotoxin tolerance, resulting in the unresponsiveness of the immune system, as a five times higher concentration of LPS was used.44-46
In conclusion, LPS exposure can have opposing effects on tumor-cell adherence and tumor development. First, by binding to TLR4, LPS induces the production of ROS by macrophages, hence initiating endothelial damage and promoting the exposure of the sub-endothelial ECM, to which circulating tumor cells can adhere. Second, LPS may stimulate the clearance of immunogenic tumor cells by promoting the initiation of adaptive immune responses. However, the lack of specific tumor-associated antigens is a major problem in most clinically manifest tumors.47 Therefore, it is likely that many patients may not benefit from LPS-induced antitumor immune responses because tumor antigens are poorly recognized by immune cells. Additionally, exposure to high LPS concentrations — as it occurs during anastomotic leakage — may induce endotoxin tolerance, explaining (at least in part) the poor survival of these patients. Increased understanding of the short-term and long-term activation of immune cells after surgery may help to identify patients at risk for the development of post-surgical liver metastases. Designing suitable peri-operative therapies to reduce this risk may ultimately improve disease outcome in cancer patients undergoing surgical resection.
Material and Methods
Colon carcinoma cell culture
The rat colon carcinoma cell line CC531s is a moderately differentiated and immunogenic cell line.48 CC531s cells were cultured under standard culture conditions in RPMI 1640 medium (Invitrogen) supplemented with 10% heat-inactivated fetal calf serum (FCS), 2 mM glutamine, 50 U/mL penicillin and 50 U/mL streptomycin (Gibco; hereafter referred as CC531s complete growth medium). The human colon carcinoma cell line SW620 (ATCC, Manassas, VA) was cultured in DMEM (Gibco) containing 10% FCS (Gibco), 2 mM glutamine, 50 U/mL penicillin and 50 U/mL streptomycin. Cell suspensions were prepared by enzymatic detachment using a trypsin-EDTA solution (Gibco), and contained single tumor cells as well as small clusters (2–8 cells). Viability was assessed by tryphan blue exclusion and always exceeded 95%. For in vivo experiments, CC531s cells (5 × 106 cells/mL) were fluorescently labeled by incubating cells in CC531s complete growth medium containing 50 μg/mL 1,1-dioctadecyl-3,3,3,3-tetramethylindocarbo cyanine perchlorate (DiI, Sigma-Aldrich) for 30 min at 37°C and subsequently washed with HBSS (Gibco) according to the manufacturer's instructions. SW620 cells (4.5 × 106 cells/mL) were fluorescently labeled by incubation at 37°C for 20 min in DMEM/ supplemented with 10% FCS and containing calcein-AM (0.5 μM, Invitrogen), after which cells were washed with Hanks’ balanced salt solution containing 0.5% bovine serum albumin and diluted (2 × 106 cells/mL).
Endothelial cell cultures
Human umbilical veins were treated with collagenase (Worthington) for 20 min at 37°C according to a standard procedure.49 HUVECs were harvested and cultured to confluence till passage 5 in M199 medium supplemented with 10% heat-inactivated human serum, 10% heat-inactivated new born calf serum, 5000 U/mL heparin, 10 μg/mL basic fibroblast growth factor (Peprotech Inc.), 2 mM glutamine, 50 U/mL penicillin and 50 U/mL streptomycin (Gibco) in standard culture conditions (hereafter referred to as endothelial growth medium).
Endothelial-macrophage co-cultures
Monocytes were isolated with CD14 MicroBeads (MACS Miltenyi Biotec) from freshly obtained donor blood, according to the manufacturer’s instructions. All healthy donors gave informed consent according to the guidelines of the medical ethical committee of the VUmc. Cells were resuspended in DMEM medium containing 10% heat-inactivated human serum, 2 mM glutamine, 50 U/mL penicillin and 50 U/mL streptomycin (Gibco; referred as macrophage growth medium). Two × 105 monocytes were added to transwells (Corning Inc.) with 0.4 μm pore size polyester membrane inserts, which had been turned upside down. Transwells were incubated for three hours to allow optimal adhesion of monocytes to membranes (on bottom side). Transwells were then turned and placed in 24 well plates for 7 d to allow differentiation into macrophages in macrophage growth medium. Hereafter, confluent HUVEC monolayers (5 × 105 cells/well) were cultured on gelatine coating on the upper site of filters. Cells were incubated with different concentrations of LPS for 30 min in M199 with or without the LPS scavenger polymyxin B sulfate (PMB, 50 μg/mL, Applichem). Alternatively, 5000 U/mL SOD (from bovine erythrocytes, Sigma Aldrich) and catalase (from bovine liver, Sigma Aldrich) were added.
Tumor-cell adhesion after exposure to LPS of HUVEC-macrophage co-cultures was investigated by adding 2 × 105 calcein-labeled SW620 cells, which were allowed to adhere for 30 min at 37°C. After washing, cells were fixated with 1% glutaraldehyde for 10 min at room temperature (RT). HUVECs monolayers were stained with Rhodamine-labeled Ulex Europaeus Agglutinin I (Vectors Labs) for 15 min at RT. Cell nuclei were stained with Hoechst (Invitrogen). Membranes were washed, removed from the inserts, mounted and 5 random pictures were made with a Leica DM6000 microscope (Leica Microsystems). Areas of damaged HUVECs monolayers were measured with AnalySIS software (Soft Imaging System GmbH).
Scanning and transmission electron microscopy (SEM and TEM)
Freshly isolated human monocytes were allowed to differentiate into macrophages for 7 d in macrophage growth medium in teflon Erlenmeyer flasks (Nalgene). Macrophages were seeded into 6-well culture plates (Greiner Bio One) and incubated for different periods with PBS or 500 ng/mL LPS. SEM was performed as described previously.11 Alternatively, 1 h after incubation with PBS or LPS, macrophages were fixed in 2.5% glutaraldehyde, post-fixed with 1% osmium tetroxide, dehydrated in ethanol infiltrated with propylene oxide and embedded in Agar 100 Resin. Ultrathin sections were stained with uranylacetate and leadcitrate and examined with TEM (Philips CM 100 Bio Twin).
Animal models
Male inbred Wag/Rij rats (180–220 g) were housed under standard laboratory conditions and had access to food and water ad libitum. The Committee for Animal Research of the VUmc approved the experiments according to institutional and national guidelines.
Portal veins of rats were catheterized as described previously.8 Catheters were flushed every 3 d with glycerol (Merck) containing heparin (50 IU/mL) to prevent obstruction of catheters by blood clots. All animals were allowed to recover 10 d. To investigate tumor-cell adhesion following an i.p. injection with PBS or 10 µg/kg LPS, 2 × 106 DiI-labeled CC531s cells were injected through the catheter and animals were sacrificed after 1.5 h (n = 4 per group). Liver samples were snap-frozen. In additional experiments, KCs were depleted by intravenous injection of clodronate (Roche Diagnostics) encapsulated in liposomes25 or not (PBS administration), 2 d prior to tumor-cell injection (n = 4 per group). Alternatively, rats received 125 mg/kg of the ROS scavenger edaravone (3-Methyl-1-Phenyl-2-Pyrazolin-5-one, Calbiochem, Darmstadt, Germany) or the vehicle ethanol (± 6%) i.p. Thirty minutes prior to and immediately after LPS or PBS injection (n = 4 per group). Animals were sacrificed 1.5 h after inoculation of fluorescently labeled CC531s cells. Liver samples were snap frozen for immunofluorescent analyses.
Fluorescence microscopy
Cryostat liver tissue sections of 5 μm were fixed for 10 min in acetone and air-dried. After blocking with 10% normal goat serum for 15 min, slides were incubated for 1 h with primary antibodies against ZO-1 (Zymed Laboratories) at RT. Rabbit serum was used as isotype control. After washing, visualization was achieved by incubation with Alexa 488-labeled secondary antibodies (Molecular Probes Inc.). Nuclei were stained with Hoechst (Molecular Probes Inc.). Sections were washed, mounted and examined with a Leica DM6000 fluorescence microscope (Leica Microsystems). Tumor-cell numbers at different time points after surgery were determined (20 stitched fields per liver sample, 5 liver samples per animal).
Statistical analyses
For comparisons between two groups Student’s t tests were used. Comparisons between multiple groups (> 2) were performed with ANOVA. Statistical significance was accepted at p < 0.05. Results are presented as means ± SEM.
Supplementary Material
Acknowledgments
This project was financially supported by Dutch Cancer Foundation (K.W.F.) project VU2011–4931 and the Netherlands Organization for Scientific Research (N.W.O.) Mosaic Bursary 017.008.057. We thank Jan van Marle (Department of Cell Biology and Histology, Academic Medical Center, University of Amsterdam, the Netherlands) for facilitating electron microscopy experiments.
Glossary
Abbreviations:
- CRC
colorectal cancer
- DiI
1,1-dioctadecyl-3,3,3,3-tetramethylindocarbo cyanine perchlorate
- ECM
extracellular matrix
- i.p.
intraperitoneal
- KC
Kupffer cells
- LPS
lipopolysaccharide
- PMB
polymyxin B
- PMN
polymorphonuclear cell
- ROS
reactive oxygen species
- RT
room temperature
- SEM
scanning electron microscopy
- SOD
superoxide dismutase
- TEM
transmission electron microscopy
- TLR
Toll-like receptor
- ZO-1
zonula occludens-1
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Footnotes
Previously published online: www.landesbioscience.com/journals/oncoimmunology/article/22303
References
- 1.Center MM, Jemal A, Ward E. International trends in colorectal cancer incidence rates. Cancer Epidemiol Biomarkers Prev. 2009;18:1688–94. doi: 10.1158/1055-9965.EPI-09-0090. [DOI] [PubMed] [Google Scholar]
- 2.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 3.Weitz J, Koch M, Debus J, Höhler T, Galle PR, Büchler MW. Colorectal cancer. Lancet. 2005;365:153–65. doi: 10.1016/S0140-6736(05)17706-X. [DOI] [PubMed] [Google Scholar]
- 4.Zdenkowski N, Chen S, van der Westhuizen A, Ackland S. Curative strategies for liver metastases from colorectal cancer: a review. Oncologist. 2012;17:201–11. doi: 10.1634/theoncologist.2011-0300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bird NC, Mangnall D, Majeed AW. Biology of colorectal liver metastases: A review. J Surg Oncol. 2006;94:68–80. doi: 10.1002/jso.20558. [DOI] [PubMed] [Google Scholar]
- 6.Coffey JC, Wang JH, Smith MJ, Bouchier-Hayes D, Cotter TG, Redmond HP. Excisional surgery for cancer cure: therapy at a cost. Lancet Oncol. 2003;4:760–8. doi: 10.1016/S1470-2045(03)01282-8. [DOI] [PubMed] [Google Scholar]
- 7.Oosterling SJ, van der Bij GJ, Bögels M, ten Raa S, Post JA, Meijer GA, et al. Anti-beta1 integrin antibody reduces surgery-induced adhesion of colon carcinoma cells to traumatized peritoneal surfaces. Ann Surg. 2008;247:85–94. doi: 10.1097/SLA.0b013e3181588583. [DOI] [PubMed] [Google Scholar]
- 8.van der Bij GJ, Oosterling SJ, Bögels M, Bhoelan F, Fluitsma DM, Beelen RH, et al. Blocking alpha2 integrins on rat CC531s colon carcinoma cells prevents operation-induced augmentation of liver metastases outgrowth. Hepatology. 2008;47:532–43. doi: 10.1002/hep.22013. [DOI] [PubMed] [Google Scholar]
- 9.Wind J, Tuynman JB, Tibbe AG, Swennenhuis JF, Richel DJ, van Berge Henegouwen MI, et al. Circulating tumour cells during laparoscopic and open surgery for primary colonic cancer in portal and peripheral blood. Eur J Surg Oncol. 2009;35:942–50. doi: 10.1016/j.ejso.2008.12.003. [DOI] [PubMed] [Google Scholar]
- 10.Sastre J, Maestro ML, Puente J, Veganzones S, Alfonso R, Rafael S, et al. Circulating tumor cells in colorectal cancer: correlation with clinical and pathological variables. Ann Oncol. 2008;19:935–8. doi: 10.1093/annonc/mdm583. [DOI] [PubMed] [Google Scholar]
- 11.Gül N, Bögels M, Grewal S, van der Meer AJ, Rojas LB, Fluitsma DM, et al. Surgery-induced reactive oxygen species enhance colon carcinoma cell binding by disrupting the liver endothelial cell lining. Gut. 2011;60:1076–86. doi: 10.1136/gut.2010.224717. [DOI] [PubMed] [Google Scholar]
- 12.van der Bij GJ, Oosterling SJ, Beelen RH, Meijer S, Coffey JC, van Egmond M. The perioperative period is an underutilized window of therapeutic opportunity in patients with colorectal cancer. Ann Surg. 2009;249:727–34. doi: 10.1097/SLA.0b013e3181a3ddbd. [DOI] [PubMed] [Google Scholar]
- 13.Chin KF, Kallam R, O’Boyle C, MacFie J. Bacterial translocation may influence the long-term survival in colorectal cancer patients. Dis Colon Rectum. 2007;50:323–30. doi: 10.1007/s10350-006-0827-4. [DOI] [PubMed] [Google Scholar]
- 14.Hsu RY, Chan CH, Spicer JD, Rousseau MC, Giannias B, Rousseau S, et al. LPS-induced TLR4 signaling in human colorectal cancer cells increases beta1 integrin-mediated cell adhesion and liver metastasis. Cancer Res. 2011;71:1989–98. doi: 10.1158/0008-5472.CAN-10-2833. [DOI] [PubMed] [Google Scholar]
- 15.Pidgeon GP, Harmey JH, Kay E, Da Costa M, Redmond HP, Bouchier-Hayes DJ. The role of endotoxin/lipopolysaccharide in surgically induced tumour growth in a murine model of metastatic disease. Br J Cancer. 1999;81:1311–7. doi: 10.1038/sj.bjc.6694369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Buttenschoen K, Buttenschoen DC, Berger D, Vasilescu C, Schafheutle S, Goeltenboth B, et al. Endotoxemia and acute-phase proteins in major abdominal surgery. Am J Surg. 2001;181:36–43. doi: 10.1016/S0002-9610(00)00534-1. [DOI] [PubMed] [Google Scholar]
- 17.Buttenschoen K, Schneider ME, Utz K, Kornmann M, Beger HG, Carli Buttenschoen D. Effect of major abdominal surgery on endotoxin release and expression of Toll-like receptors 2/4. Langenbecks Arch Surg. 2009;394:293–302. doi: 10.1007/s00423-008-0357-8. [DOI] [PubMed] [Google Scholar]
- 18.Schietroma M, Carlei F, Cappelli S, Amicucci G. Intestinal permeability and systemic endotoxemia after laparotomic or laparoscopic cholecystectomy. Ann Surg. 2006;243:359–63. doi: 10.1097/01.sla.0000201455.89037.f6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Koratzanis G, Giamarellos-Bourboulis EJ, Papalambros E, Giamarellou H. Bacterial translocation following intrabdominal surgery. Any influence of antimicrobial prophylaxis? Int J Antimicrob Agents. 2002;20:457–60. doi: 10.1016/S0924-8579(02)00237-6. [DOI] [PubMed] [Google Scholar]
- 20.Reddy BS, MacFie J, Gatt M, Macfarlane-Smith L, Bitzopoulou K, Snelling AM. Commensal bacteria do translocate across the intestinal barrier in surgical patients. Clin Nutr. 2007;26:208–15. doi: 10.1016/j.clnu.2006.10.006. [DOI] [PubMed] [Google Scholar]
- 21.Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010;11:373–84. doi: 10.1038/ni.1863. [DOI] [PubMed] [Google Scholar]
- 22.Boueiz A, Hassoun PM. Regulation of endothelial barrier function by reactive oxygen and nitrogen species. . Microvasc Res. 2009;77:26–34. doi: 10.1016/j.mvr.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 23.Mittal A, Phillips AR, Loveday B, Windsor JA. The potential role for xanthine oxidase inhibition in major intra-abdominal surgery. World J Surg. 2008;32:288–95. doi: 10.1007/s00268-007-9336-4. [DOI] [PubMed] [Google Scholar]
- 24.Bentes de Souza AM, Rogers MS, Wang CC, Yuen PM, Ng PS. Comparison of peritoneal oxidative stress during laparoscopy and laparotomy. J Am Assoc Gynecol Laparosc. 2003;10:65–74. doi: 10.1016/S1074-3804(05)60237-X. [DOI] [PubMed] [Google Scholar]
- 25.Van Rooijen N, Sanders A. Liposome mediated depletion of macrophages: mechanism of action, preparation of liposomes and applications. J Immunol Methods. 1994;174:83–93. doi: 10.1016/0022-1759(94)90012-4. [DOI] [PubMed] [Google Scholar]
- 26.Feng S, Yang Q, Liu M, Li W, Yuan W, Zhang S, et al. Edaravone for acute ischaemic stroke. Cochrane Database Syst Rev. 2011:CD007230. doi: 10.1002/14651858.CD007230.pub2. [DOI] [PubMed] [Google Scholar]
- 27.Lapchak PA. A critical assessment of edaravone acute ischemic stroke efficacy trials: is edaravone an effective neuroprotective therapy? Expert Opin Pharmacother. 2010;11:1753–63. doi: 10.1517/14656566.2010.493558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sica A, Mantovani A. Macrophage plasticity and polarization: in vivo veritas. J Clin Invest. 2012;122:787–95. doi: 10.1172/JCI59643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Su GL, Klein RD, Aminlari A, Zhang HY, Steinstraesser L, Alarcon WH, et al. Kupffer cell activation by lipopolysaccharide in rats: role for lipopolysaccharide binding protein and toll-like receptor 4. Hepatology. 2000;31:932–6. doi: 10.1053/he.2000.5634. [DOI] [PubMed] [Google Scholar]
- 30.Spolarics Z. Endotoxemia, pentose cycle, and the oxidant/antioxidant balance in the hepatic sinusoid. J Leukoc Biol. 1998;63:534–41. doi: 10.1002/jlb.63.5.534. [DOI] [PubMed] [Google Scholar]
- 31.McDonald B, McAvoy EF, Lam F, Gill V, de la Motte C, Savani RC, et al. Interaction of CD44 and hyaluronan is the dominant mechanism for neutrophil sequestration in inflamed liver sinusoids. J Exp Med. 2008;205:915–27. doi: 10.1084/jem.20071765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Koch M, Kienle P, Hinz U, Antolovic D, Schmidt J, Herfarth C, et al. Detection of hematogenous tumor cell dissemination predicts tumor relapse in patients undergoing surgical resection of colorectal liver metastases. Ann Surg. 2005;241:199–205. doi: 10.1097/01.sla.0000151795.15068.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Freitas M, Lima JL, Fernandes E. Optical probes for detection and quantification of neutrophils’ oxidative burst. A review. Anal Chim Acta. 2009;649:8–23. doi: 10.1016/j.aca.2009.06.063. [DOI] [PubMed] [Google Scholar]
- 34.Kaplan JH, Groves JN. Liver and blood cell catalase activity of tumor-bearing mice. Cancer Res. 1972;32:1190–4. [PubMed] [Google Scholar]
- 35.Nishikawa M. Reactive oxygen species in tumor metastasis. Cancer Lett. 2008;266:53–9. doi: 10.1016/j.canlet.2008.02.031. [DOI] [PubMed] [Google Scholar]
- 36.Lee CC, Avalos AM, Ploegh HL. Accessory molecules for Toll-like receptors and their function. Nat Rev Immunol. 2012;12:168–79. doi: 10.1038/nri3151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chen L, Huang TG, Meseck M, Mandeli J, Fallon J, Woo SL. Rejection of metastatic 4T1 breast cancer by attenuation of Treg cells in combination with immune stimulation. Mol Ther. 2007;15:2194–202. doi: 10.1038/sj.mt.6300310. [DOI] [PubMed] [Google Scholar]
- 38.Tamai H, Watanabe S, Zheng R, Deguchi K, Cohen PA, Koski GK, et al. Effective treatment of spontaneous metastases derived from a poorly immunogenic murine mammary carcinoma by combined dendritic-tumor hybrid vaccination and adoptive transfer of sensitized T cells. Clin Immunol. 2008;127:66–77. doi: 10.1016/j.clim.2007.12.001. [DOI] [PubMed] [Google Scholar]
- 39.Van de Vrie W, Marquet RL, Eggermont AM. Cyclosporin A enhances locoregional metastasis of the CC531 rat colon tumour. J Cancer Res Clin Oncol. 1997;123:21–4. doi: 10.1007/BF01212610. [DOI] [PubMed] [Google Scholar]
- 40.Van Putte BP, Hendriks JM, Romijn S, Pauwels B, Vermorken JB, Van Schil PE. Combination chemotherapy with gemcitabine with isolated lung perfusion for the treatment of pulmonary metastases. J Thorac Cardiovasc Surg. 2005;130:125–30. doi: 10.1016/j.jtcvs.2004.07.045. [DOI] [PubMed] [Google Scholar]
- 41.Gao T, Bi H, Ma S, Lu J. The antitumor and immunostimulating activities of water soluble polysaccharides from Radix Aconiti, Radix Aconiti Lateralis and Radix Aconiti Kusnezoffii. Nat Prod Commun. 2010;5:447–55. [PubMed] [Google Scholar]
- 42.Grauer OM, Molling JW, Bennink E, Toonen LW, Sutmuller RP, Nierkens S, et al. TLR ligands in the local treatment of established intracerebral murine gliomas. J Immunol. 2008;181:6720–9. doi: 10.4049/jimmunol.181.10.6720. [DOI] [PubMed] [Google Scholar]
- 43.Wolska A, Lech-Marańda E, Robak T. Toll-like receptors and their role in carcinogenesis and anti-tumor treatment. Cell Mol Biol Lett. 2009;14:248–72. doi: 10.2478/s11658-008-0048-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Biswas SK, Lopez-Collazo E. Endotoxin tolerance: new mechanisms, molecules and clinical significance. Trends Immunol. 2009;30:475–87. doi: 10.1016/j.it.2009.07.009. [DOI] [PubMed] [Google Scholar]
- 45.Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010;11:889–96. doi: 10.1038/ni.1937. [DOI] [PubMed] [Google Scholar]
- 46.Cavaillon JM, Adib-Conquy M. Bench-to-bedside review: endotoxin tolerance as a model of leukocyte reprogramming in sepsis. Crit Care. 2006;10:233. doi: 10.1186/cc5055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Vergati M, Intrivici C, Huen NY, Schlom J, Tsang KY. Strategies for cancer vaccine development. J Biomed Biotechnol. 2010;2010 doi: 10.1155/2010/596432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Marquet RL, Westbroek DL, Jeekel J. Interferon treatment of a transplantable rat colon adenocarcinoma: importance of tumor site. Int J Cancer. 1984;33:689–92. doi: 10.1002/ijc.2910330521. [DOI] [PubMed] [Google Scholar]
- 49.Jaffe EA, Nachman RL, Becker CG, Minick CR. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J Clin Invest. 1973;52:2745–56. doi: 10.1172/JCI107470. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.