

Abstract
We have developed a fully automated system for operant behavior testing and neuronal activity recording by which multiple cognitive brain functions can be investigated in a single task sequence. The unique feature of this system is a custom-made, acoustically transparent chamber that eliminates many of the issues associated with auditory cue control in most commercially available chambers. The ease with which operant devices can be added or replaced makes this system quite versatile, allowing for the implementation of a variety of auditory, visual, and olfactory behavioral tasks. Automation of the system allows fine temporal (10 ms) control and precise time-stamping of each event in a predesigned behavioral sequence. When combined with a multi-channel electrophysiology recording system, multiple cognitive brain functions, such as motivation, attention, decision-making, patience, and rewards, can be examined sequentially or independently.
Keywords: Neuroscience, Issue 63, auditory behavioral task, acoustic chamber, cognition test, multi-channel recording, electrophysiology, attention, motivation, decision, patience, rat, two-alternative choice pitch discrimination task, behavior
Protocol
System Overview
The system comprises three main components: (1) a double-walled sound proof room (Industrial Acoustical Company, Bronx, New York); (2) a multiple channel electrophysiological recording system (Neuralynx, Bozeman, MT); and (3) a fully automated, customized behavioral testing system from the Med Associates Inc. (St. Albans, VT).
As shown in Figure 1A, the operant chamber is located inside the sound-proof room. A commutator (Model SL-36, Dragonfly Research and Development, Inc., Ridgeley, West Virginia) for connecting cables from headstage to the electrophysiological recording system (Figure1A-a), and a video camera for monitoring and recording animal behaviors are mounted above the operant chamber (Figure1A-b).
Custom-Designed Operant Chamber
The custom-designed, acoustically transparent operant chamber (Figure1A-d) consists of three acoustically transparent walls and one modular-operation panel (Figure 1B). Three speakers (Cage Tweeter, ENV_224BM, Med Associates) mounted on the top of the middle and two side panels are used for emitting auditory cues. Auditory cues are generated by a calibrated, programmable audio generator (ANL-926). A stimulus light (ENV_221M) and two triple-stimulus LED displays (ENV_222M) are located on the middle and side panels, respectively. These stimulus lights can be used for auditory-visual multi-sensory behavioral tests. A nose poke device with three color LED lights (ENV_114M) is mounted at the bottom of the middle panel. An infrared detector installed within the nose poke unit is used to signal nose poking and holding period. The LED lights within the nose poke unit can be used for training nose-hold inside the hole. A movable response lever (ENV_112CM) is mounted on each side of the operant panel. The mobility of these levers allows flexible control of the presence of the levers, which can be effectively used for both initial task training and the study of several important cognitive functions of the brain (see below). Four pairs of infrared beam sources and detectors (EVN_253SD) controlled by a four channel IR controller (ENV_253) are placed at the bottom of the chamber to both indicate the positions of the animal and control other devices based on an animal's position (Figure 1A-e). Two pellet dispensers each with a built-in infrared sentry (ENV_203M-45IR) are used for dispensing rewards into the pellet receptacles (Figure 1A-c). The infrared sentry is used for monitoring failure of pellet dispenser and providing warning signals.
System Configuration
The overview of system connections and hardware components is illustrated in Figure 2. Functionally, there are two parallel, interactive sub-systems: one for behavior training and the other for electrophysiological recording. The two sub-systems are synchronized through the MED-PC IV Software platform (SOF-735). The computer sends commands to behavioral devices and TTL pulses to neural recording system (signal flows indicated by the red arrows in Figure 2), and receives signals generated by animal's responses and neural activities (signal flows indicated by the green and blue arrows, respectively, in Figure 2). These parallel, interactive systems enable recordings of synchronized behavioral/neuronal data and allow manipulation of animal's behavior based on neural activity, or vice versa.
Behavior training sub-system A SmartCtrl Connection Panel (SG-716B) serves as a two-way communication panel: i.e. sending the control signals (red arrow in Figure 2) from computer to the behavioral devices (listed in red box), and transmit animal's response signals (green arrow in Figure 2) back to the computer. The outputs of the four channel IR controller (ENV_253) are also routed into the connection panel. An interface card (SmartCtrl interface card, DIG-716B) and a decode card (DIG-700F) communicate the signals from the connection panel to a PCI card (DIG-704PCI) installed in the computer. The auditory cues are generated by the Stimulus Generator (ANL-926), which is also controlled by the MED-PC IV Software through the decode card (DIG-700F). As illustrated in Figure 2, all the interface cards are hosted in a Tabletop Interface Cabinet (SG-6080D). This cabinet also supplies powers for all the behavior devices.
Electrophysiological recording sub-system Animals' response signals received by the computer are instantaneously sent to the neural recording system via the SuperPort TTL card (DIG-726) and the Cheetah Digital Interface Box (Neuralynx, Bozeman, MT) (Figure 2). These behavior events are time-stamped and recorded simultaneously with neural activities. Neural spikes detected online from the Neuralynx recording system can be used as input signals to the behavioral control sub-system for manipulating or interfering animal's behavior. Conversely, the animal's response signals can be used as triggers for manipulating or interfering neuronal activities when combined with electric or optogenetic stimulation techniques. These approaches will be valuable for elucidating causalities between neuronal activities and behaviors.
Programming and data processing The behavioral control programs are written with Trans IV software (Thomas A. Tatham and MED Associates) and compiled with Pascal compiler. Automation of each training step is realized by loading the Trans IV program into the MED-PC IV Software. Training parameters can also be adjusted online by trainers while the MED-PC IV Software is running. The Trans IV codes need to be specific for both system setup and behavioral task. Standard training programs are, however, freely available from the MED Associates and can be modified to meet individual laboratory's specific needs. Training programs used in our setups are also freely available upon request.
Behavioral data are saved automatically by the MED-PC IV Software. The saved data can be translated into Microsoft Excel files by using the MED-PC To Excel program (MPC2XL, Thomas A. Tatham and MED Associates). The translated Excel files can then be imported and analyzed in a MATLAB environment (MathWorks, Natick, MA). The neural data along with the behavioral event timestamps recorded with the Cheetah software (Cheetah 5, Neuralynx, Bozeman, MT) can also be imported into MATLAB for analysis.
Training
To illustrate the operation of this system, we describe here a Two-Alternative Choice Pitch Discrimination Task, designed to examine the frequency discrimination threshold of a rat. A schematic illustration of the task is shown in Figure 3.
1. Pre-training
Begin with naïve adult male, Sprague-Dawley, rats, age ~60 days.
Before training, limit the food intake until the animal's weight is ~90% of baseline ad libidum weight.
2. Cage Acclimation
To prepare the chamber for acclimation, retract the levers and block the nose poke hole with a rubber stopper (made from the plunger of a 60 cc syringe) to prevent animal from activating the nose-poke device.
Place ~20 fully nutritional pellets (45 mg, Product # F0021, BioServ, Frenchtown, NJ) in each pellet receptacle (food cup).
Place a naïve animal in the chamber for acclimation. The rat will soon start to explore the food cups and eat the pellets.
When both food cups are empty, use the software control to dispense a single pellet into each tray. In doing so, the rat will learn to associate food with the cup.
Force the rat to move to both sides of the chamber by dispensing pellets randomly into each food cup. One 30 minute session is usually enough for establishing the food-cup association. In one 30 minute session, the rat usually obtains 200 - 300 pellets, which are enough to maintain their body weight in a constant level of ~90% of baseline.
3. Lever-push Training
In a new session, extend both levers into the chamber and leave the food cup empty.
Next, place an acclimated rat into the chamber. When the animal enters the vicinity of a lever, dispense a pellet manually through the Med-PC IV software. Also provide rewards when the rat shows interest in the lever, such as sniffing, touching, or climbing. An accidental lever push should also trigger a reward automatically by the program.
To encourage lever pushing and to force exploration of both levers, allow the animal to push each lever consecutively a limited number of times. When the limit is reached, retract the lever. When both levers have retracted, extend them to repeat the procedure.
Gradually decrease the limit until the lever retracts every time it is pushed. One to two 30 minute sessions are usually enough for establishing the lever push - food reward association.
4. Nose-poke Training
In a new session, retract the levers, and remove the rubber stopper from the nose poke. Place several pellets inside the nose poke to encourage rat's interest in exploring the nose-poke device.
Reintroduce the animal into the chamber. Extend one of the two levers randomly when the rat sniffs the nose-poke hole for food pellets.
Seeing the extended lever, the rat will approach and push the lever to obtain a food pellet. After the lever is pressed and the reward is dispensed, retract the lever to encourage the rat to explore the nose poke device. It usually takes about 20 to 30 minutes to learn the task sequence: Nose-poke →Lever extension→Lever push→Rewards.
5. Cue Training
In a new session, play the auditory cues following a nose-poke event with a short delay (100 to 250 ms). Extend both left and right levers shortly (100 ms) after each auditory cue presentation.
Reward the rat only when it pushes the lever that is indicated by the auditory cue. The animal will gradually learn to associate a specific auditory cue with one lever. The animal is then free to begin a new trial with the sequence: Nose-poke → Cue→Lever push→Reward/No Reward (Figure 3). Because of high false rate in the initial learning sessions, supplementary food should be given to maintain the body weight after each training session.
In the next several 30 minute sessions, let the rat practice the newly learned task until a consistent performance level is reached (see Figure 3A for a typical learning curve). Once the task is mastered, a rat can obtain about 200 - 300 pellets in each 30 minute session, which are enough to maintain its body weight.
6. Representative Results
Following the above protocol, we trained rats to recognize two different patterns of pure tone pulse trains consisting of six tone pips with either the same frequency (F,F,F,F,F,F) or different frequencies (F, F-ΔF, F, F-ΔF, F, F-ΔF) 1-5. Each tone pip is 200 ms in duration and the tone pip interval is 400 ms. In the present study, F was set to be 10 kHz and ΔF ranged from 1 to 50% of the F (Figure 3, top). Typically, ΔF was set at relatively large value during training sessions: 5 kHz, 4 kHz, 3 kHz, 2 kHz, and 1 kHz, to facilitate the training. Each pure tone pulse train with different ΔF value was presented randomly in a given session.
The Two-Alternative Choice Pitch Discrimination Task is illustrated in Figure 3. Rats were trained to poke their nose into the nose hole (Figure 1B and Figure 3, bottom) to start a trial. Nose poke triggers broadcasting of the auditory cues. Upon recognition of the cues, rats need to run to the correct side of the chamber, approach the lever, wait for the lever to extend into the chamber, and then push the lever within a certain time window (lever active time, 1 to 2 seconds) to obtain a reward (Figure 3). The hit rate was calculated for each ΔF value as the number of hit trials divided by the number of total trials for each individual ΔF value. A criterion of 75% hit rate was used to indicate that the rat had learned the task. A typical learning curve of one rat is presented in Figure 4A. Each colored line depicts the learning progress for each pulse train with different ΔF (ΔF0 represents the constant pulse train). On average, it took about seven sessions of training (starting from the first session of auditory cue presentation, Step 5 Cue Training) to reach the 75% hit rate criterion.
This system also allows quantitative characterization of animal's behavior in performing the task designed by experimenters. Three measurements that are widely used in animal behavioral studies are shown in Figure 4 B - D. The reaction time, reflecting mainly an animal's attentiveness to the task, was measured as elapse of time between onset of auditory cue and lever push. The inter-trial interval, reflecting how committed an animal was to the task in which the animal initiated each trial and was not punished by time-out in false trial, was plotted in Figure 4C. The temporal variation of performance within a session, representing the dynamic patterns of an animal's overall performance and reflecting improvement/adaptation that could occur within a single session, was plotted in Figure 4D. Each session was divided into early, middle, and late stage (10 minutes per stage). Accumulated numbers of rewards in each stage were used in this measurement.
The neural basis of several cognitive behaviors (Figure 5A and see Discussion) can also be addressed with this system by recording the neural activities of animals performing a task. Examples of neural activity recorded simultaneously in the nucleus basalis (NB) and ventral tegmental area (VTA) of a rat brain are shown in Figure 5B and C. The firings of neurons are time-stamped to relate to each event of the trial (such as nose-poke, lever push, auditory cue presentation and recognition, and actual receiving of reward) and analyzed in relation to these behavioral task events. Results of combined behavioral and neural activity recording with this system will be fruitful in elucidating the neural basis of a variety of cognitive behaviors.
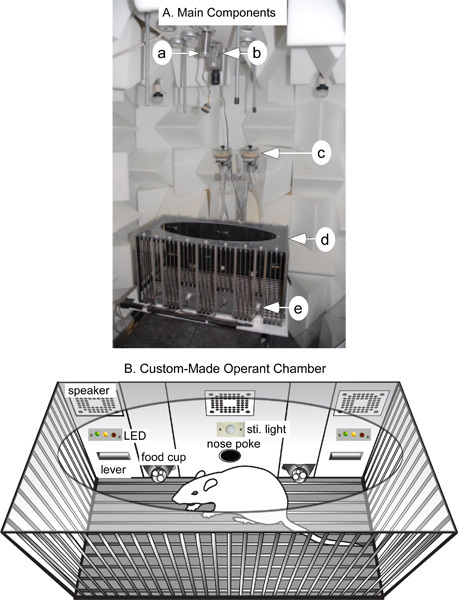 Figure 1. The main components of the system (A) and a schematic drawing of the custom-made auditory operant chamber (B). A. The operant chamber is located in a double-walled sound-proof room. Anechoic foam wedges are mounted throughout the walls of the room to eliminate sound scattering and deflection. a: Commutator for routing wires to neuronal activity recording system; b: Video camera for monitoring and recording animal behavior; c: Pellet dispensers; d: Operant chamber. e: Infrared receivers. B. The operant chamber comprises three acoustically transparent walls and one modular-operation panel. See Text for detailed descriptions.
Figure 1. The main components of the system (A) and a schematic drawing of the custom-made auditory operant chamber (B). A. The operant chamber is located in a double-walled sound-proof room. Anechoic foam wedges are mounted throughout the walls of the room to eliminate sound scattering and deflection. a: Commutator for routing wires to neuronal activity recording system; b: Video camera for monitoring and recording animal behavior; c: Pellet dispensers; d: Operant chamber. e: Infrared receivers. B. The operant chamber comprises three acoustically transparent walls and one modular-operation panel. See Text for detailed descriptions.
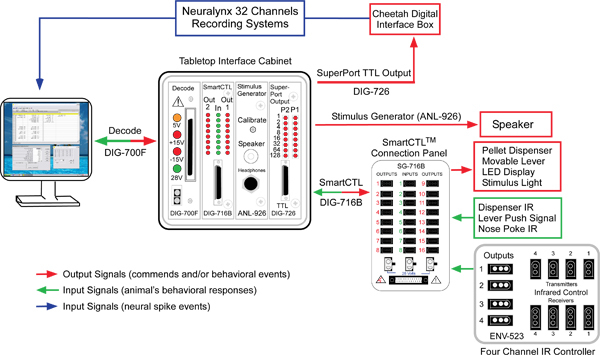 Figure 2. Schematic overview of the system. The system consists of two sub-systems: behavioral training and neural activity recording system. The two subsystems interactively communicate with each other via TTL pulses (see System Configuration for details). The red arrows represent commands and/or behavioral events sent out from the computer; the green arrows denote animal response signals feeding back to the computer; and the blue arrow represents signal inputs of neural spike events detected online with the Neuralynx recording system.
Figure 2. Schematic overview of the system. The system consists of two sub-systems: behavioral training and neural activity recording system. The two subsystems interactively communicate with each other via TTL pulses (see System Configuration for details). The red arrows represent commands and/or behavioral events sent out from the computer; the green arrows denote animal response signals feeding back to the computer; and the blue arrow represents signal inputs of neural spike events detected online with the Neuralynx recording system.
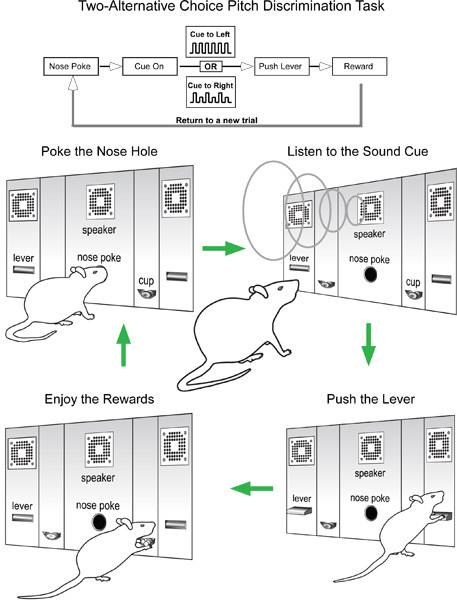 Figure 3. Two-Alternative Choice Frequency Discrimination Task. Top, Block diagram showing the basic task sequence. Bottom, Schematic depiction of main behavioral actions. Green arrows indicate the sequential flow of the task.
Figure 3. Two-Alternative Choice Frequency Discrimination Task. Top, Block diagram showing the basic task sequence. Bottom, Schematic depiction of main behavioral actions. Green arrows indicate the sequential flow of the task.
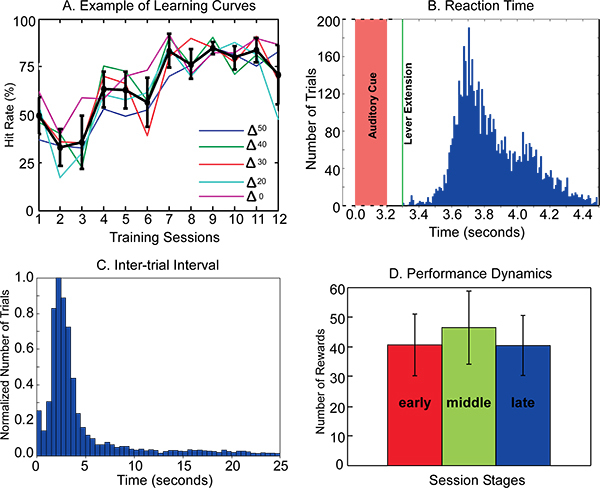 Figure 4. Representative results of Two-alternative Choice Frequency Discrimination Task. A. Learning curves: Each colored line represents the learning progress of a rat on discriminating each frequency variation (ΔFs). The dark line represents the averaged learning curve for frequency variations. B. Distribution of reaction time measured as time laps from the onset of cue to lever push. C. Distribution of inter-trial interval. D. Temporal dynamics of performance within a session measured with accumulated rewards obtained in early, middle, and late stage of a session. All the data in B - D was obtained in the last stage when the rat's performance was over the 75% hit rate.
Figure 4. Representative results of Two-alternative Choice Frequency Discrimination Task. A. Learning curves: Each colored line represents the learning progress of a rat on discriminating each frequency variation (ΔFs). The dark line represents the averaged learning curve for frequency variations. B. Distribution of reaction time measured as time laps from the onset of cue to lever push. C. Distribution of inter-trial interval. D. Temporal dynamics of performance within a session measured with accumulated rewards obtained in early, middle, and late stage of a session. All the data in B - D was obtained in the last stage when the rat's performance was over the 75% hit rate.
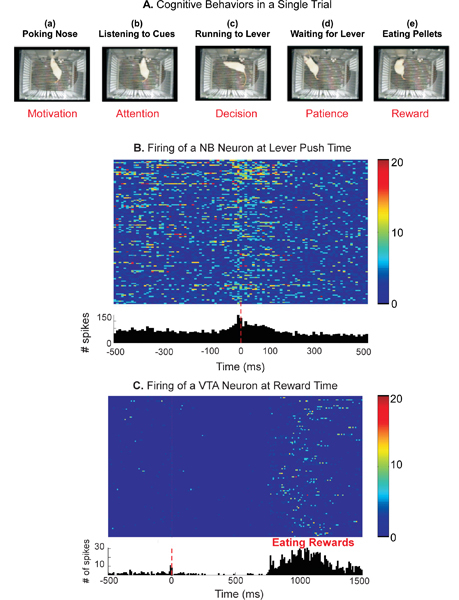 Figure 5. Example of cognitive behaviors and brain functions that can be investigated using the system. A. Cognitive behaviors. The top captions describe each action in a sequence of one trial. The bottom captions indicate the cognitive behaviors that can be studied. Note that both levers were extracted in all pictures except for in d where the lever is in the processing of extending into the chamber. B. Firing patterns of a neuron recorded in the NB of a rat performing the two-choice auditory task. Top, Raster image plot of firing across each trial. Each colored rectangle represents the firing of the cell and the firing rate is coded by the color. Bottom, Peri-event histogram of the firing rate. Note the building-up of the firing before the action (pushing lever at time zero microseconds, indicated by the red-dotted vertical line) and gradual dispersion of the firing after the action. C. Firing patterns of a neuron recorded in the VTA of a rat performing the two-choice auditory task. Top, Raster image plot of the firing of a single VTA neuron across each trial. Each colored rectangle represents the firing of the VTA neuron and its firing rate is coded by color. Bottom, Peri-event histogram of the firing rate shown in the raster image. Note the sparse firing immediately before the action of pushing lever (at time zero microseconds, indicated by the vertical red-dotted line) and vigorous firing during the time period when the rat is obtaining the reward. The activity of this neuron is virtually silent between these two actions. Tungsten stereotrodes implanted in the brain were used to record neural activities simultaneously from the NB and VTA while the rat was performing the task. Spike-sorting was performed off-line using SpikeSort 3D Software (Neuralynx, Bozeman, MT).
Figure 5. Example of cognitive behaviors and brain functions that can be investigated using the system. A. Cognitive behaviors. The top captions describe each action in a sequence of one trial. The bottom captions indicate the cognitive behaviors that can be studied. Note that both levers were extracted in all pictures except for in d where the lever is in the processing of extending into the chamber. B. Firing patterns of a neuron recorded in the NB of a rat performing the two-choice auditory task. Top, Raster image plot of firing across each trial. Each colored rectangle represents the firing of the cell and the firing rate is coded by the color. Bottom, Peri-event histogram of the firing rate. Note the building-up of the firing before the action (pushing lever at time zero microseconds, indicated by the red-dotted vertical line) and gradual dispersion of the firing after the action. C. Firing patterns of a neuron recorded in the VTA of a rat performing the two-choice auditory task. Top, Raster image plot of the firing of a single VTA neuron across each trial. Each colored rectangle represents the firing of the VTA neuron and its firing rate is coded by color. Bottom, Peri-event histogram of the firing rate shown in the raster image. Note the sparse firing immediately before the action of pushing lever (at time zero microseconds, indicated by the vertical red-dotted line) and vigorous firing during the time period when the rat is obtaining the reward. The activity of this neuron is virtually silent between these two actions. Tungsten stereotrodes implanted in the brain were used to record neural activities simultaneously from the NB and VTA while the rat was performing the task. Spike-sorting was performed off-line using SpikeSort 3D Software (Neuralynx, Bozeman, MT).
Discussion
A critical aspect in design of any auditory behavioral task is elimination of unwanted sounds resulting from scattering and deflection in the test environment. Poor sound control can have a significant effect on the behavior being tested and will produce misleading or even uninterpretable results. The behavioral chamber used in the system described here is specifically designed to be acoustically transparent in order to avoid sound deflection from the chamber walls. Indeed, when measured from the center of the chamber, sound deflection was effectively undetectable (data not shown).
Although we developed this system mainly for studying the auditory system, it can be easily adapted by other researchers to study other sensory systems. Modifications can be made easily on both software and hardware for different tasks without changing the overall configuration of the system. The modular operation panel makes the system particularly versatile by allowing for addition and/or substitution of different devices for new behavioral tasks. For instance, olfactory behavioral tasks can be implanted by delivering olfactory stimulus into the nose poke device. The five choice illuminated nose poke wall with olfactory stimulus from the MED Associates (ENV-115A-OF) can be easily installed on the operation panel for complex olfactory tasks. In addition, all the operant devices can be easily replaced with those that are designed for mice without changing the configuration of the system.
Precise temporal control of each operant device, as well as high resolution recording of single events in a given trial, enable accurate manipulation of the devices for customizing the design of behavioral tasks to address different cognitive brain functions (see below). When combined with neuronal activity recording, a rich variety of issues in the field of neurosciences can be studied with this system. For instance, in the auditory task described above, the following questions related to brain cognitive functions can be investigated in a single trial:
(1). Motivation: Since each trial is initiated by animal's "self-motivated" nose-poke action (Figure 5A-a and Figure 3), motivation can thus be assessed quantitatively by measuring the total number of trials performed by an animal in a given session, or the number of consecutive trials performed6,7.
(2). Attention: The key for obtaining a reward in a trial is to correctly recognize the auditory cues. In about 25% of rats that could not be trained to perform the task, lack of attending to the auditory cues was the major factor. In contrast, in the rats who learned the task, a momentary pause of behaviors was apparent during auditory cue presentation (see Figure 5A-b and Figure 3). By using this system, it is thus possible to (i) screen rats for "attention deficits" and (ii) study neural mechanisms of attention when combined with neuronal recordings while the animal is attending to the auditory cues8-10.
(3). Decision: Upon recognition of the auditory cues, the animal has to decide which direction to turn to approach the correct lever within a limited time window (Figure 5A-c). It is thus also an effective paradigm for studying decision-making11,12.
(4). Patience: The timing of lever extension can be controlled such that the animal has to wait for the lever after arriving at the spot where the lever will extend (Figure 5A-d). By varying the length of waiting, the extent of an animal's patience can be tested and quantified13.
(5). Rewards: The final goal of the task is to obtain the reward (Figure 5A-e and Figure 3). Behavioral tasks using this system can thus be easily designed to study many aspects of reward-decision issues and the function of the value systems of the brain14-17.
Disclosures
Production and Free Access of this video-article is sponsored by Med Associates, Inc.
Acknowledgments
This work was supported by the Neurosciences Research Foundation and grants from The Blasker-Rose-Miah Fund of The San Diego Foundation and The G. Harold and Leila Y. Mathers Charitable Foundation.
References
- Zheng W. Auditory map reorganization and pitch discrimination in adult rats chronically exposed to low-level ambient noise. Front. Syst.Neurosci. 2012;6:65. doi: 10.3389/fnsys.2012.00065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talwar SK, Gerstein GL. Auditory frequency discrimination in the white rat. Hear Res. 1998;126:135–150. doi: 10.1016/s0378-5955(98)00162-2. [DOI] [PubMed] [Google Scholar]
- Talwar SK, Gerstein GL. A signal detection analysis of auditory-frequency discrimination in the rat. J. Acoust. Soc. Am. 1999;105:1784–1800. doi: 10.1121/1.426716. [DOI] [PubMed] [Google Scholar]
- Sloan AM, Dodd OT, Rennaker RL. Frequency discrimination in rats measured with tone-step stimuli and discrete pure tones. Hear Res. 2009;251:60–69. doi: 10.1016/j.heares.2009.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Syka J, Rybalko N, Brozek G, Jilek M. Auditory frequency and intensity discrimination in pigmented rats. Hear Res. 1996;100:107–113. doi: 10.1016/0378-5955(96)00101-3. [DOI] [PubMed] [Google Scholar]
- Eldar E, Morris G, Niv Y. The effects of motivation on response rate: A hidden semi-Markov model analysis of behavioral dynamics. J. Neurosci. Methods. 2011. [DOI] [PubMed]
- Bieszczad KM, Weinberger NM. Learning strategy trumps motivational level in determining learning-induced auditory cortical plasticity. Neurobiology of Learning and Memory. 2010;93:229–239. doi: 10.1016/j.nlm.2009.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sagvolden TImpulsiveness, overactivity poorer sustained attention improve by chronic treatment with low doses of l-amphetamine in an animal model of Attention-Deficit/Hyperactivity Disorder (ADHD) Behav. Brain Funct. 2011;7:6. doi: 10.1186/1744-9081-7-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sagvolden T, Johansen EB. Rat Models of ADHD. Curr. Top Behav. Neurosci. 2011. [DOI] [PubMed]
- Brown DC, Nichols JA, 2nd, Thomas F, Dinh L, Atzori M. Nicotinic modulation of auditory attentional shift in the rat. Behavioural brain research. 2010;210:273–279. doi: 10.1016/j.bbr.2010.02.046. [DOI] [PubMed] [Google Scholar]
- Johnson KO. Sensory discrimination: neural processes preceding discrimination decision. J. Neurophysiol. 1980;43:1793–1815. doi: 10.1152/jn.1980.43.6.1793. [DOI] [PubMed] [Google Scholar]
- Johnson KO. Sensory discrimination: decision process. J. Neurophysiol. 1980;43:1771–1792. doi: 10.1152/jn.1980.43.6.1771. [DOI] [PubMed] [Google Scholar]
- Fraser LM, et al. Measuring anxiety- and locomotion-related behaviours in mice: a new way of using old tests. Psychopharmacology (Berl) 2010;211:99–112. doi: 10.1007/s00213-010-1873-0. [DOI] [PubMed] [Google Scholar]
- Hui GK, et al. Conditioned tone control of brain reward behavior produces highly specific representational gain in the primary auditory cortex. Neurobiology of Learning and Memory. 2009;92:27–34. doi: 10.1016/j.nlm.2009.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feduccia AA, Duvauchelle CL. Auditory stimuli enhance MDMA-conditioned reward and MDMA-induced nucleus accumbens dopamine, serotonin and locomotor responses. Brain Res. Bull. 2008;77:189–196. doi: 10.1016/j.brainresbull.2008.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kudoh M, Shibuki K. Sound sequence discrimination learning motivated by reward requires dopaminergic D2 receptor activation in the rat auditory cortex. Learning & Memory. 2006;13:698–69. doi: 10.1101/lm.390506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiflett MW, et al. Cue-elicited reward-seeking requires extracellular signal-regulated kinase activation in the nucleus accumbens. J. Neurosci. 2008;28:1434–1443. doi: 10.1523/JNEUROSCI.2383-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]