

Abstract
After its release into the synaptic cleft, dopamine exerts its biological properties via its pre- and post-synaptic targets1. The dopamine signal is terminated by diffusion2-3, extracellular enzymes4, and membrane transporters5. The dopamine transporter, located in the peri-synaptic cleft of dopamine neurons clears the released amines through an inward dopamine flux (uptake). The dopamine transporter can also work in reverse direction to release amines from inside to outside in a process called outward transport or efflux of dopamine5. More than 20 years ago Sulzer et al. reported the dopamine transporter can operate in two modes of activity: forward (uptake) and reverse (efflux)5. The neurotransmitter released via efflux through the transporter can move a large amount of dopamine to the extracellular space, and has been shown to play a major regulatory role in extracellular dopamine homeostasis6. Here we describe how simultaneous patch clamp and amperometry recording can be used to measure released dopamine via the efflux mechanism with millisecond time resolution when the membrane potential is controlled. For this, whole-cell current and oxidative (amperometric) signals are measured simultaneously using an Axopatch 200B amplifier (Molecular Devices, with a low-pass Bessel filter set at 1,000 Hz for whole-cell current recording). For amperometry recording a carbon fiber electrode is connected to a second amplifier (Axopatch 200B) and is placed adjacent to the plasma membrane and held at +700 mV. The whole-cell and oxidative (amperometric) currents can be recorded and the current-voltage relationship can be generated using a voltage step protocol. Unlike the usual amperometric calibration, which requires conversion to concentration, the current is reported directly without considering the effective volume7. Thus, the resulting data represent a lower limit to dopamine efflux because some transmitter is lost to the bulk solution.
Keywords: Neuroscience, Issue 69, Cellular Biology, Physiology, Medicine, Simultaneous Patch Clamp and Voltametry, In Vitro Voltametry, Dopamine, Oxidation, Whole-cell Patch Clamp, Dopamine Transporter, Reverse transport, Efflux
Protocol
1. Equipment and Supplies
Mount a Faraday cage on top of the anti vibration table (TMI) to decrease the background noise.
The simultaneous patch clamp amperometry recording system requires an inverted microscope with excellent DIC optics and a long working distance lens. Connect the microscope lighthouse to a car battery. This DC light source for the system will further decrease the electrical noise.
Hydraulic micromanipulators (Siskiyou) further decrease noise. In our configuration, we use a right handed manipulator for the whole cell recording, and the left handed for amperometry.
2. Prepare Electrodes for Recording
Pull patch electrodes using quartz pipettes on a P-2000 puller (Sutter). Our pull lasts approximately 5 sec, with two heat cycles. This heat time has resulted in consistent resistance (3-4 MΩ) in our whole cell patch pipettes.
Fill the electrode with the pipette solution containing 2 mM dopamine and mount it on the right manipulator. Wrap the container holding the pipette solution containing DA with aluminum foil. Keep on ice. Dopamine is oxidizable. Keeping the solution on ice, protected from light will decrease the oxidation rate of dopamine.
Gently remove a ProCFE (Dagan) low noise carbon fiber amperometric electrode from the storage box, fill with mercury, mount onto the amperometric adaptor (as depicted in Figure 1), and then mount on the right maniupulator. Protect the tip of the carbon fiber from damage by holding the far end of the carbon fiber. Inspect the electrode with a lab microscope to ensure the tip is clean and intact.
Examine the integrity of the amperometric electrode by putting the electrode in a glass bottom Petri dish containing external solution. Record a baseline current in the absence of dopamine. Add 10 μl of a 1mM DA solution to the dish. A good amperometric electrode records an increase in the oxidative current. Repeat this step at the start and end of each experiment to make certain the amperometric electrode works properly.
3. Prepare the Primary Neuronal Culture of Dopamine Neurons or Cells Engineered to Express Dopamine Transporter in Glass Bottom Petri Dishes
Gently wash the cells or dopamine neurons three times with warm external solution.
Mount the glass bottom Petri dish onto the microscope stage.
4. Visualize Cell and Perform Experiment
Find the correct focal point to clearly visualize the cells. Place positive pressure on the patch electrode. Then, gently bring both electrodes down into the solution, and close to the cell.
Position the amperometric electrode next to the cell (on left side), and the patch electrode on the right.
Attain a gigaohm seal on the cell with the patch electrode. Rupture the seal with suction to achieve whole cell configuration.
Allow 5-8 min for dialysis of internal solution containing dopamine into the cell.
Use desired voltage step or ramp protocol. Simultaneously acquire data from both the patch pipette and the amperometric electrode to measure whole cell currents and reverse transport of dopamine through the dopamine transporter while the membrane potential is controlled via the patch pipette.
Representative Results
Combined patch clamp with amperometry can measure voltage-dependent DAT-mediated DA efflux. Figure 2A shows a representative experimental configuration and recording of DAT-mediated DA efflux when the intracellular milieu and the membrane potential are clamped by a whole-cell patch pipette. Using this technique, cells expressing YFP-DAT proteins are voltage-clamped with a whole cell patch pipette while an amperometric electrode is placed onto the plasma membrane (Figure 2A). The whole cell electrode is filled with a pipette solution containing 2 mM DA. The amperometric electrode touching the plasma membrane is held at +700 mV, a potential greater than the redox potential of DA. DAT-mediated currents (whole-cell-Figure 2C and amperometric-Figure 2D) are recorded by stepping the whole-cell pipette to a membrane voltage between -60 and +100 mV from a holding potential of -40 mV (Figure 2B). To determine the specific DAT-mediated DA efflux, amperometric currents can be recorded in the absence and presence of a DAT antagonist. The DAT-mediated amperometric current is then determined by subtracting the traces recorded in the presence of DAT antagonist from amperometric traces recorded without antagonist. An upward deflection (positive deflection, Figure 2D) in the amperometric currents corresponds to an outward flux of DA. Moving the carbon fiber away from the patch causes the oxidative response to become smaller and slower. As expected for DA oxidation, the oxidative response decreases when the carbon fiber voltage is reduced to +300 mV and will disappear completely on further reduction. Figure 2E-2F are representative of acceptable and unacceptable recordings. Figure 2E is an example of a tight GΩ whole-cell seal and the corresponding amperometric signal. Figure 2F represents a leaky whole-cell patch. The corresponding amperometric recording measures leaked DA.
 Figure 1. Preparation and assembly of amperometric electrode. Gently remove a ProCFE low noise carbon fiber amperometric electrode from the storage box, fill with mercury. Prepare the adaptor by threading the silver wire into the patch clamp holder, then thread the adaptor onto the patch clamp holder and gently mount the amperometric electrode. The tip of the carbon fiber should not touch anything.
Figure 1. Preparation and assembly of amperometric electrode. Gently remove a ProCFE low noise carbon fiber amperometric electrode from the storage box, fill with mercury. Prepare the adaptor by threading the silver wire into the patch clamp holder, then thread the adaptor onto the patch clamp holder and gently mount the amperometric electrode. The tip of the carbon fiber should not touch anything.
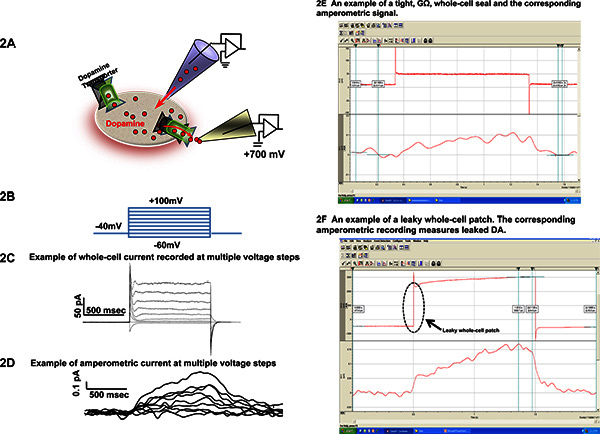 Figure 2. Representative whole-cell and amperometric recording at multiple voltages when the patch pipette is in whole-cell configuration. (A) Cartoon illustrating the experimental configuration. Cells expressing dopamine transporter were voltage clamped with a whole-cell patch pipette while an amperometric electrode was placed near the cell membrane. The oxidation of DA results in a positive amperometric current. (B) Illustration of voltage clamp waveform protocol. (C) DAT-mediated currents recorded by stepping the membrane voltage between -60 and +100 mV from a holding potential of -40 mV with the whole-cell pipette. The pipette solution contained 2 mM dopamine, as described previously7-9. (D) The amperometric current acquired simultaneously with the whole-cell current represented in panel C (above). At the start of the voltage step, for voltages higher than +20 mV, the amperometric electrode records an oxidation current (positive) that increased during the entire duration of the voltage step. (E) and (F) are representative of acceptable and unacceptable whole-cell patch clamp experiments, respectively. Figure 2E is an example of a tight GΩ whole-cell seal and the corresponding amperometric signal. Figure 2F represents a leaky whole-cell patch. The seal resistance reached the MΩ range instead of GΩ prior or after breaking into the cell to achieve whole-cell mode. The corresponding amperomtric recording measures leaked DA. Click here to view larger figure.
Figure 2. Representative whole-cell and amperometric recording at multiple voltages when the patch pipette is in whole-cell configuration. (A) Cartoon illustrating the experimental configuration. Cells expressing dopamine transporter were voltage clamped with a whole-cell patch pipette while an amperometric electrode was placed near the cell membrane. The oxidation of DA results in a positive amperometric current. (B) Illustration of voltage clamp waveform protocol. (C) DAT-mediated currents recorded by stepping the membrane voltage between -60 and +100 mV from a holding potential of -40 mV with the whole-cell pipette. The pipette solution contained 2 mM dopamine, as described previously7-9. (D) The amperometric current acquired simultaneously with the whole-cell current represented in panel C (above). At the start of the voltage step, for voltages higher than +20 mV, the amperometric electrode records an oxidation current (positive) that increased during the entire duration of the voltage step. (E) and (F) are representative of acceptable and unacceptable whole-cell patch clamp experiments, respectively. Figure 2E is an example of a tight GΩ whole-cell seal and the corresponding amperometric signal. Figure 2F represents a leaky whole-cell patch. The seal resistance reached the MΩ range instead of GΩ prior or after breaking into the cell to achieve whole-cell mode. The corresponding amperomtric recording measures leaked DA. Click here to view larger figure.
Discussion
Simultaneous voltage-clamp and amperometry has the following benefits. All cell types are accessible and can be used for recording. The identification of the cells or neuron where the recordings are done is simple and straightforward. In particular, if the cell is fluorescently labeled by adding a fluorescent tag to the protein of interest the experimenter can easily select the target cell or neuron. The experimental configuration allows uniform and controlled delivery of pharmacological agents either via the patch pipette, or by addition of these agents to the bath solution10. The simultaneous patch clamp amperometry technique allows investigation of the role of different channels and transporters on the release of oxidizable transmitter with millisecond time resolution while the membranepotential is controlled. The same information cannot be obtained through biochemical methods8. One of the unique aspects of this approach is that it allows manipulation of the intracellular environment by dialysis via the patch pipette7,11. However, this technique has the disadvantage that the amperometric electrode can detect only oxidizable transmitters. In addition, the sensitivity of the amperometric electrode decreases in the presence of substantial basal, spontaneous release of an oxidizable compound; thus, measurement of the release after, for example, preloading of an oxidizable compound that is spontaneously released can produce a decrease in the oxidative current. The main disadvantage of this technique is the requirement of technical expertise and expensive equipment.
Disclosures
No conflicts of interest declared.
Acknowledgments
We thank Dr. Sanika Chirwa for critical review of this manuscript. This work was supported by National Institutes of Health (DA026947, DA021471, and NS071122).
References
- Michael AC, Ikeda M, Justice JB., Jr Mechanisms contributing to the recovery of striatal releasable dopamine following MFB stimulation. Brain Res. 1987;421:325–335. doi: 10.1016/0006-8993(87)91302-3. [DOI] [PubMed] [Google Scholar]
- Gonon F. Prolonged and extrasynaptic excitatory action of dopamine mediated by D1 receptors in the rat striatum in vivo. J. Neurosci. 1997;17:5972–5978. doi: 10.1523/JNEUROSCI.17-15-05972.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sulzer D, Pothos EN. Regulation of quantal size by presynaptic mechanisms. Rev. Neurosci. 2000;11:159–212. doi: 10.1515/revneuro.2000.11.2-3.159. [DOI] [PubMed] [Google Scholar]
- Napolitano A, Cesura AM, Da Prada M. The role of monoamine oxidase and catechol O-methyltransferase in dopaminergic neurotransmission. J. Neural. Transm. Suppl. 1995;45:35–45. [PubMed] [Google Scholar]
- Sulzer D, Maidment NT, Rayport S. Amphetamine and other weak bases act to promote reverse transport of dopamine in ventral midbrain neurons. J. Neurochem. 1993;60:527–535. doi: 10.1111/j.1471-4159.1993.tb03181.x. [DOI] [PubMed] [Google Scholar]
- Salahpour A, et al. Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc. Natl. Acad. Sci. U.S.A. 2008;105:4405–4410. doi: 10.1073/pnas.0707646105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khoshbouei H, Wang H, Lechleiter JD, Javitch JA, Galli A. Amphetamine-induced dopamine efflux. A voltage-sensitive and intracellular Na+-dependent mechanism. J. Biol. Chem. 2003;278:12070–12077. doi: 10.1074/jbc.M212815200. [DOI] [PubMed] [Google Scholar]
- Goodwin JS, et al. Amphetamine and methamphetamine differentially affect dopamine transporters in vitro and in. 2009. pp. 284–2978. [DOI] [PMC free article] [PubMed]
- Kahlig KM, et al. Amphetamine induces dopamine efflux through a dopamine transporter channel. Proc. Natl. Acad. Sci. U.S.A. 2005;102:3495–3500. doi: 10.1073/pnas.0407737102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swant J, Chirwa S, Stanwood G, Khoshbouei H. Methamphetamine reduces LTP and increases baseline synaptic transmission in the CA1 region of mouse hippocampus. PLoS One. 2010;5:e11382. doi: 10.1371/journal.pone.0011382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gnegy ME, et al. Intracellular Ca2+ regulates amphetamine-induced dopamine efflux and currents mediated by the human dopamine transporter. Mol. Pharmacol. 2004;66:137–143. doi: 10.1124/mol.66.1.137. [DOI] [PubMed] [Google Scholar]