

Abstract
The brain renin-angiotensin system (RAS) has been highlighted as having a pathological role in stroke, dementia, and neurodegenerative disease. Particularly, in dementia, epidemiological studies indicate a preventive effect of RAS blockade on cognitive impairment in Alzheimer disease (AD). Moreover, basic experiments suggest a role of brain angiotensin II in neural injury, neuroinflammation, and cognitive function and that RAS blockade attenuates cognitive impairment in rodent dementia models of AD. Therefore, RAS regulation is expected to have therapeutic potential for AD. Here, we discuss the role of angiotensin II in cognitive impairment and AD. Angiotensin II binds to the type 2 receptor (AT2) and works mainly by binding with the type 1 receptor (AT1). AT2 receptor signaling plays a role in protection against multiple-organ damage. A direct AT2 receptor agonist is now available and is expected to reduce inflammation and oxidative stress and enhance cell differentiation. We and other groups reported that AT2 receptor activation enhances neuronal differentiation and neurite outgrowth in the brain. Here, we also review the effect of the AT2 receptor on cognitive function. RAS modulation may be a new therapeutic option for dementia including AD in the future.
1. Introduction
The renin-angiotensin system (RAS) in the brain is well known to be involved in systemic blood pressure control, including the regulation of cerebral blood flow [1]. Angiotensin II, a major player in RAS mainly via the angiotensin type 1 (AT1) receptor, plays an important role in the pathophysiology of tissue dysfunction [2, 3]; therefore, RAS blockade by AT1 receptor blockers (ARBs) and angiotensin converting enzyme inhibitors (ACEIs), which are widely used as antihypertensive drugs, is expected to prevent multiple-organ damage. Cognitive impairment and dementia are common serious health problems that impair quality of life in the elderly. Previous reports indicate the possibility that treatment with antihypertensive agents prevents the impairment of quality of life including cognitive performance [4, 5]. Possible beneficial effects of RAS blockade on cognitive function are also being highlighted in the clinical field [6, 7]. An epidemiological study by Li et al. recently showed that male subjects treated with ARBs exhibited a significant reduction in the incidence and progression of Alzheimer disease (AD) and dementia compared with those treated with ACEIs and other cardiovascular drugs [8]. Moreover, Davies et al. also reported that patients diagnosed with dementia had fewer prescriptions for ARBs and ACEIs. Interestingly, the inverse associations with AD were stronger for ARBs compared with ACEIs [9]. In contrast, Ohrui et al. demonstrated that long-term use of ACEIs may have a protective role against the development of AD, probably through their direct effects on RAS in the brain [10]. In a subanalysis of the Study on Cognition and Prognosis in the Elderly (SCOPE) trial, hypertensives treated with an ARB, candesartan, showed less decline of specific areas of cognitive function such as attention and episodic memory [11]. However, almost all large clinical intervention trials have shown no significant difference in the incidence of dementia between treatment with ARBs or ACEIs and the placebo group. The Ongoing telmisartan alone and in combination with ramipril global endpoint trial (ONTARGET) and the parallel telmisartan randomized assessment study in ACE intolerant subjects with cardiovascular disease (TRANSCEND) trial showed no clear effects on cognitive outcomes [12]. The reason why RAS blockade failed to prevent dementia may be the short-term observation for the long-term preclinical disease stage of dementia; however, the detailed explanation is not clear. Another reason is the selection of hypertensive patients, who have high cardiovascular disease morbidity, in these trials. A large number of these patients are likely to go on to develop dementia, most likely with strong vascular underpinning. In these trials, vascular dementia and AD are not well distinguished because most studies focused on dementia as subanalysis. As described in the review by Kehoe and Passmore, RAS has multifunctional involvement not only in vascular dementia but also in AD [13]. Therefore, in such specific groups with cardiovascular risk, the distinction of dementia subtype is very important in comparing the incidence of dementia.
The effect of angiotensin II on cognition has been examined in basic studies. Although the blood-brain barrier is impermeable for all RAS components, the local brain RAS has possible physiological and pharmacological functions in the neuronal system [14]. Gard reviewed the contradictory role of angiotensin II in memory and learning in animal studies [15]. Angiotensin II enhances memory and learning in rodents [16, 17], but other studies suggest that angiotensin II decreases cognition [18]. To assess the paradoxical effect of angiotensin II on cognitive function, we therefore performed cognitive tests in mice with continuous activation of angiotensin II, using transgenic mice carrying both the human renin and angiotensinogen genes (hRN/hANG-Tg) [19]. Interestingly, the avoidance rate in hRN/hANG-Tg mice did not increase from 14 weeks of age; however, that from 8 to 13 weeks of age tended to be higher than that in wild-type mice. These findings suggest that the acute or subacute effect of angiotensin II may enhance cognitive function, but chronic treatment with angiotensin II may exhaust neural function and result in cognitive impairment. Angiotensin II induces cerebrovascular remodeling, promotes vascular inflammation and oxidative stress, and results in impairment of regulation of cerebral blood flow (CBF) [20, 21]. Moreover, endothelial function in cerebral vessels was impaired in a genetic model of angiotensin-II-dependent hypertension [22, 23]. On the other hand, Lanz et al. showed that angiotensin II induced sustained central nervous system (CNS) inflammation via transforming growth factor- (TGF-)β in an experimental autoimmune encephalomyelitis (EAE) mouse model [24]. Furthermore, angiotensin II induced astrocyte senescence, which is involved in age-associated neurodegenerative disease via superoxide production [25]. In contrast, a centrally active ACE inhibitor, perindopril, was reported to prevent cognitive impairment in chronic central hypoperfusion rats [26] and Alzheimer disease model mice [27]. These reports indicate that continuous angiotensin II stimulation impairs cognitive function via stimulation of the AT1 receptor with “environmental degradation of neurons” such as a decrease in CBF and an increase in oxidative stress, CNS inflammation, and cellular senescence in the brain. Such multiple stimuli by angiotensin II induce cognitive impairment following neuronal degeneration.
2. Effects of Angiotensin II on Amyloid β Metabolism and Cholinergic System
There are two major proposed pathomechanisms of AD; the amyloid cascade hypothesis and the cholinergic hypothesis. Amyloid β (Aβ) is a 39–42 amino acid peptide, produced by cleavage of amyloid precursor protein (APP) [28]. Aβ (1–42) causes the neurodegenerative abnormalities that lead to clinical AD [29]. Although the effect of angiotensin converting enzyme on Aβ metabolism is one of the hot topics in the relation between RAS and AD [30], it seems that angiotensin II does not directly affect Aβ secretion or secretase activity via activation of the AT1 receptor [31]. On the other hand, blockade of RAS may affect Aβ metabolism. For example, an ARB, valsartan, was able to attenuate oligomerization of amyloid β peptides into high molecular weight oligomeric peptides [32]. Moreover, treatment with valsartan also disrupted the development of amyloid β-mediated cognitive impairment in Tg2576 mice, a model of Alzheimer disease; however, it is reported that this beneficial effect is not observed with treatment with other ARBs. We previously reported that Aβ (1–40) concentration in the brain of ddY mice that underwent intracerebroventricular injection of Aβ (1–40) was significantly decreased by treatment with an ARB, telmisartan [33]. Moreover, Danielyan et al. reported that intranasal administration of losartan exerts direct neuroprotective effects via its Aβ-reducing and anti-inflammatory effects in the central nervous system [34]. These results indicate that treatment with ARBs may have a beneficial effect on Aβ-induced brain injury through unknown mechanisms on Aβ metabolism by angiotensin II inhibition. On the other hand, brain-penetrating ACEIs such as perindopril prevent cognitive impairment in mice with intracerebroventricular Aβ (1–40) injection via attenuation of oxidative stress and hippocampal astrocyte activation [35]. ACE activity is increased in the hippocampus of these AD mice and suppressed by perindopril treatment. Although there is concern that ACEIs may enhance brain Aβ (1–42) deposition from basic research [36] because ACE converts Aβ (1–42), which plays a causative role in the development of Alzheimer disease, to Aβ (1–40) [37], recent pilot clinical trials showed that ramipril inhibits cerebrospinal fluid (CSF) ACE activity, but did not influence CSF Aβ (1–42) and cognition [38]. The effects of other RAS components involving angiotensin-II-generating enzymes on cognition have also been discussed. Aβ clearance is induced by many kinds of degrading enzyme such as neprilysin (NEP), insulin-degrading enzyme, and endothelin-converting enzyme. Angiotensin II is also generated by degradation of angiotensinogen and angiotensin I by tonins, cathepsins, and chymases as well as ACE. Gene polymorphism in cathepsin G, one of the angiotensin generating enzymes, showed no significant association with AD [39]. In our knowledge, no report has examined the relation between tonin, chymase, and dementia; however, inhibition of angiotensin generating enzymes may also inhibit Aβ degradation. Therefore, it is difficult to assess the effect on Aβ metabolism of drugs that inhibit angiotensin II based on degrading angiotensinogen. Further investigation is necessary to understand the relation among angiotensin II, ACE, other degrading enzymes, and Aβ metabolism.
In the cholinergic hypothesis, AD is also characterized by a loss of neurons, especially those expressing nicotinic acetylcholine receptors (nAChR) [40, 41]. To improve the cognitive deficit in AD, one promising drug target currently under investigation is the neuronal nicotinic alpha7 acetylcholine receptor (α7nAChR) [42, 43]. Although there are few reports about the correlation between α7nAChR and angiotensin II, Marrero's group has demonstrated that angiotensin II blocks nicotine-mediated neuroprotection against Aβ (1–42) via activation of the tyrosine phosphatase, SHP-1 [44]. They also showed that angiotensin II inhibits α7nAChR-induced activation of the JAK2-PI-3 K cascade in PC12 cells through AT2 receptor-induced SHP-1 activation [45]. However, AT2 receptor-induced SHP-1 activation also induces cerebellar development and neural differentiation [46, 47]. Moreover, Aβ triggered AT2 receptor oligomerization in the hippocampus [48] and impaired coupling of the muscarinic acetylcholine receptor (mAChR) to heterotrimeric GTP-binding proteins (Gα q/11) [49]. Therefore, the AT2 receptor may interact with the cholinergic system; however, the actual effect of angiotensin II mediated by AChRs is still an enigma (Figure 1).
Figure 1.
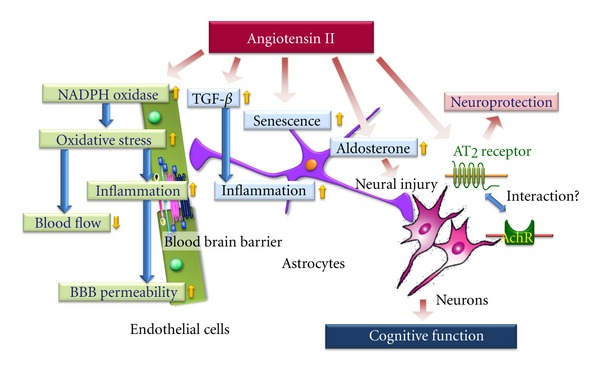
Possible effect of angiotensin II on neurovascular unit. AT2: angiotensin II type 2 receptor, AchR: acetylcholine receptor, BBB: blood brain barrier, and TGF-β: transforming growth factor β.
3. Effects of Angiotensin II on Neurovascular Unit
Nonneuronal cells such as vascular cells and glia (astrocytes, microglia and oligodendroglia) comprise the “neurovascular unit” and could play important roles in disease pathogenesis [50]. Especially, CBF functions in concert as a part of the neurovascular unit to maintain homeostasis of the cerebral microenvironment [51]. Iadecola and colleagues demonstrated that angiotensin II increases the production of reactive oxygen species (ROS) in cerebral microvessels via gp91phox (nox-2), a subunit of NADPH oxidase [20, 51]. Moreover, recently they also demonstrated that slow infusion of the pressor angiotensin II induces attenuation of the increase in CBF induced by neural activity (whisker stimulation) and by endothelium-dependent vasodilators, without elevation of mean arterial pressure (MAP) [52]. Such an effect of angiotensin II reduces blood supply and contributes to increased susceptibility to dementia. Interestingly, this angiotensin-induced cerebrovascular dysregulation was attenuated in female compared with male mice [53]. This sexual dimorphism of the cerebral blood-vessel response to angiotensin II may be implicated in the sex difference in cognitive impairment reported in epidemiological studies [54]. On the other hand, Takeda et al. demonstrated that the ARB olmesartan ameliorates amyloid β-induced impairment of functional hyperemia evoked by whisker stimulation via a decrease in oxidative stress in brain microvessels [55]. Recently, Zhang et al. reported that angiotensin II increases cerebral microvasculature inflammation via induction of oxidative stress and leads to immune-endothelial interaction, resulting in enhancement of BBB permeability [56]. Therefore, angiotensin-II-induced oxidative stress may have a key role in dysfunction of the neurovascular unit (Figure 1).
On the other hand, several reports indicate the effect of angiotensin II on astrocytes to be neuroinflammation, neuronal damage and astrocyte senescence. For example, Lanz et al. clearly demonstrated that angiotensin II acts as a paracrine mediator, sustaining inflammation in the CNS via TGF-β upregulation in astrocytes [24]. We also reported that aldosterone secretion induced by angiotensin II in astrocytes enhances neuronal damage due to angiotensin II [57]. Moreover, Liu et al. showed that angiotensin II induces astrocyte senescence via superoxide production [25]. These findings of astrocyte dysfunction induced by angiotensin II also explain the crucial role of angiotensin II in dysfunction of the neurovascular unit (Figure 1).
4. Effect of AT2 Receptors on Cognition and Dementia
The major actions of angiotensin II are mediated by the AT1 receptor, whereas the role of a second receptor subtype known as the angiotensin II type 2 (AT2) receptor is suggested to be protecting of the brain [58]. In the brain, AT2 receptors are expressed not only in the vascular wall but also in areas related to learning and control of motor activity [59, 60]. Mice with deletion of the AT2 receptor were reported to exhibit worse cognitive function compared with wild-type mice [60]. Reinecke et al. demonstrated the possibility that stimulation of the AT2 receptor may promote cell differentiation and regeneration in neuronal tissue [61] and that AT2 receptor stimulation supported neuronal survival and neurite outgrowth in response to ischemia-induced neuronal injury [62]. We also demonstrated that AT2 receptor signaling enhanced neural differentiation and the repair of damaged DNA through induction of a neural differentiating factor, methyl methanesulfonate-sensitive 2 (MMS2), which is one of the ubiquitin conjugating enzyme variants [47]. Moreover, Gallo-Payet et al. reported that angiotensin II induces neural differentiation and neurite outgrowth via mitogen-activated protein kinase [63] or nitric oxide [64] through AT2 receptor activation, and is involved in cerebellar development [65]. Therefore, direct AT2 receptor stimulation is expected to have a beneficial effect on cognitive function. We examined the possibility that direct stimulation of the AT2 receptor by a newly generated direct AT2 receptor agonist, Compound 21 (C21), would enhance cognitive function [66]. Daily intraperitoneal injection of C21 for 2 weeks significantly enhanced spatial learning evaluated by the Morris water maze test in C57BL6 mice, but this effect was not observed in AT2 receptor-deficient mice. C21 treatment increased cerebral blood flow assessed by laser speckle flowmetry and hippocampal field-excitatory postsynaptic potential. Moreover, treatment with C21 prevented cognitive decline in an Alzheimer disease mouse model with intracerebroventricular injection of amyloid β (1–40). AT2 receptor activation is reported to stimulate the release of NO/cGMP and may mediate vascular relaxation and blood flow indirectly through modulation of bradykinin release [67]. In our model, C21-induced cognitive enhancement was attenuated by coadministration of icatibant, a bradykinin B2 receptor antagonist. Therefore, direct activation of the AT2 receptor improves spatial learning via an increase in microcirculation, partly through modulation of bradykinin. The preventive effect of AT2 receptor signaling on dementia is summarized in Figure 2. Clinical use of C21 is expected to be a new therapeutic option in patients with dementia.
Figure 2.
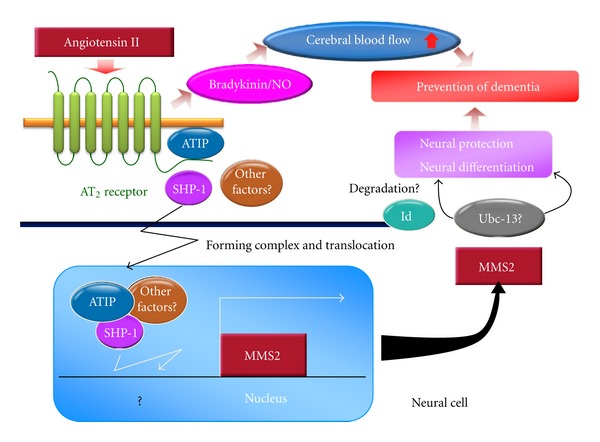
Effect of angiotensin II type 2 receptor signaling on cognitive function. AT2: angiotensin II type 2 receptor, ATIP: AT2 receptor-interacting protein, Id1: inhibitor of DNA binding protein 1, MMS2: methyl methanesulfonate-sensitive 2, NO: nitric oxide, SHP-1: Src homology 2 domain-containing protein-tyrosine phosphatase 1, and Ubc-13: ubiquitin conjugating enzyme 13.
5. Conclusion
Continuous stimulation with angiotensin II may damage neurons via multiple cascades through AT1 receptor stimulation. On the other hand, stimulation of the AT2 receptor is expected to prevent neural damage and cognitive impairment (Figure 3). However, it is difficult to perform clinical intervention studies to confirm the results of animal studies because of the long-term progression of cognitive impairment. Moreover, in clinical practice, it is not possible to exclude the antihypertensive effect of RAS blockade on cognition in patients with hypertension. However, RAS modulation may be a new therapeutic option for dementia including AD in the future. Therefore, the hypothesis that RAS regulation affects future cognitive function should be confirmed with carefully designed clinical studies.
Figure 3.
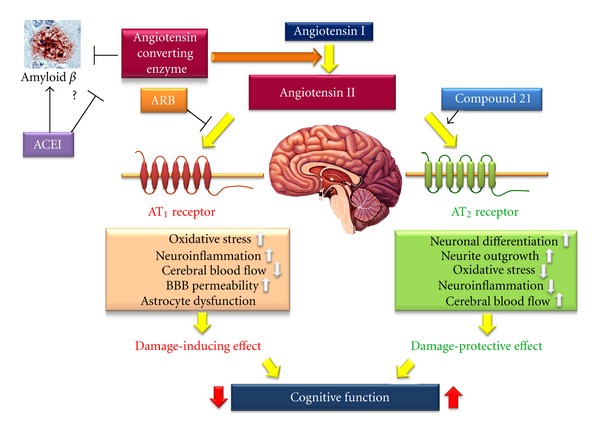
Effect of angiotensin II on cognitive function. ACE: angiotensin converting enzyme inhibitor, AT1: angiotensin II type 1 receptor, AT2: angiotensin II type 2 receptor, and ARB: angiotensin II type 1 receptor blocker.
Conflict of Interests
The authors declare no conflict of interests.
Acknowledgment
This work was supported by grants from the Ministry of Education, Science, Sports, and Culture of Japan to M. Mogi, J. Iwanami, and M. Horiuchi.
Abbreviations
- AD:
Alzheimer disease
- ARB:
Angiotensin II type 1 (AT1) receptor blocker
- AT1 receptor:
Angiotensin II type 1 receptor
- AT2 receptor:
Angiotensin II type 2 receptor
- ACE:
Angiotensin converting enzyme
- CBF:
Cerebral surface blood flow
- hRN/hANG-Tg:
Human renin and angiotensinogen genes
- RAS:
Renin-angiotensin system
- si:
Small interfering
- MMS2:
Methyl methanesulfonate-sensitive 2.
References
- 1.de Gasparo M, Catt KJ, Inagami T, Wright JW, Unger T. International union of pharmacology. XXIII. The angiotensin II receptors. Pharmacological Reviews. 2000;52(3):415–472. [PubMed] [Google Scholar]
- 2.Schmieder RE, Hilgers KF, Schlaich MP, Schmidt BM. Renin-angiotensin system and cardiovascular risk. The Lancet. 2007;369(9568):1208–1219. doi: 10.1016/S0140-6736(07)60242-6. [DOI] [PubMed] [Google Scholar]
- 3.Iwanami J, Mogi M, Iwai M, Horiuchi M. Inhibition of the renin-angiotensin system and target organ protection. Hypertension Research. 2009;32(4):229–237. doi: 10.1038/hr.2009.5. [DOI] [PubMed] [Google Scholar]
- 4.Fogari R, Zoppi A. Effect of antihypertensive agents on quality of life in the elderly. Drugs and Aging. 2004;21(6):377–393. doi: 10.2165/00002512-200421060-00003. [DOI] [PubMed] [Google Scholar]
- 5.Fletcher A. Quality of life in the management of hypertension. Clinical and Experimental Hypertension. 1999;21(5-6):961–972. doi: 10.3109/10641969909061024. [DOI] [PubMed] [Google Scholar]
- 6.Wright JW, Harding JW. The brain RAS and Alzheimer’s disease. Experimental Neurology. 2010;223(2):326–333. doi: 10.1016/j.expneurol.2009.09.012. [DOI] [PubMed] [Google Scholar]
- 7.Mogi M, Horiuchi M. Effects of angiotensin II receptor blockers on dementia. Hypertension Research. 2009;32(9):738–740. doi: 10.1038/hr.2009.110. [DOI] [PubMed] [Google Scholar]
- 8.Li NC, Lee A, Whitmer RA, et al. Use of angiotensin receptor blockers and risk of dementia in a predominantly male population: prospective cohort analysis. British Medical Journal. 2010;340:p. b5465. doi: 10.1136/bmj.b5465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Davies NM, Kehoe PG, Ben-Shlomo Y, Martin RM. Associations of anti-hypertensive treatments with Alzheimer’s disease, vascular dementia, and other dementias. Journal of Alzheimer’s Disease. 2011;26(4):699–708. doi: 10.3233/JAD-2011-110347. [DOI] [PubMed] [Google Scholar]
- 10.Ohrui T, Matsui T, Yamaya M, et al. Angiotensin-converting enzyme inhibitors and incidence of Alzheimer’s disease in Japan [7] Journal of the American Geriatrics Society. 2004;52(4):649–650. doi: 10.1111/j.1532-5415.2004.52178_7.x. [DOI] [PubMed] [Google Scholar]
- 11.Saxby BK, Harrington F, Wesnes KA, McKeith IG, Ford GA. Candesartan and cognitive decline in older patients with hypertension: a substudy of the SCOPE trial. Neurology. 2008;70(19):1858–1866. doi: 10.1212/01.wnl.0000311447.85948.78. [DOI] [PubMed] [Google Scholar]
- 12.Anderson C, Teo K, Gao P, et al. Renin-angiotensin system blockade and cognitive function in patients at high risk of cardiovascular disease: analysis of data from the ONTARGET and TRANSCEND studies. The Lancet Neurology. 2011;10(1):43–53. doi: 10.1016/S1474-4422(10)70250-7. [DOI] [PubMed] [Google Scholar]
- 13.Kehoe PG, Passmore PA. The renin-angiotensin system and antihypertensive drugs in Alzheimer’s disease: current standing of the angiotensin hypothesis? Journal of Alzheimer’s Disease. 2012;30(supplement 2):S251–S268. doi: 10.3233/JAD-2012-111376. [DOI] [PubMed] [Google Scholar]
- 14.Wright JW, Harding JW. Brain renin-angiotensin-A new look at an old system. Progress in Neurobiology. 2011;95(1):49–67. doi: 10.1016/j.pneurobio.2011.07.001. [DOI] [PubMed] [Google Scholar]
- 15.Gard PR. The role of angiotensin II in cognition and behaviour. European Journal of Pharmacology. 2002;438(1-2):1–14. doi: 10.1016/s0014-2999(02)01283-9. [DOI] [PubMed] [Google Scholar]
- 16.Georgiev V, Yonkov D. Participation of angiotensin II in learning and memory. I. Interaction of angiotensin II with saralasin. Methods and Findings in Experimental and Clinical Pharmacology. 1985;7(8):415–418. [PubMed] [Google Scholar]
- 17.Kułakowska A, Karwowska W, Wiśniewski K, Braszko JJ. Losartan influences behavioural effects of angiotensin II in rats. Pharmacological Research. 1996;34(3-4):109–115. doi: 10.1006/phrs.1996.0073. [DOI] [PubMed] [Google Scholar]
- 18.Raghavendra V, Chopra K, Kulkarni SK. Involvement of cholinergic system in losartan-induced facilitation of spatial and short-term working memory. Neuropeptides. 1998;32(5):417–421. doi: 10.1016/s0143-4179(98)90065-8. [DOI] [PubMed] [Google Scholar]
- 19.Inaba S, Iwai M, Furuno M, et al. Continuous activation of renin-angiotensin system impairs cognitive function in renin/angiotensinogen transgenic mice. Hypertension. 2009;53(2):356–362. doi: 10.1161/HYPERTENSIONAHA.108.123612. [DOI] [PubMed] [Google Scholar]
- 20.Kazama K, Anrather J, Zhou P, et al. Angiotensin II impairs neurovascular coupling in neocortex through NADPH oxidase-derived radicals. Circulation Research. 2004;95(10):1019–1026. doi: 10.1161/01.RES.0000148637.85595.c5. [DOI] [PubMed] [Google Scholar]
- 21.Wei Y, Whaley-Connell AT, Chen K, et al. NADPH oxidase contributes to vascular inflammation, insulin resistance, and remodeling in the transgenic (mRen2) rat. Hypertension. 2007;50(2):384–391. doi: 10.1161/HYPERTENSIONAHA.107.089284. [DOI] [PubMed] [Google Scholar]
- 22.Didion SP, Sigmund CD, Faraci FM. Impaired endothelial function in transgenic mice expressing both human renin and human angiotensinogen. Stroke. 2000;31(3):760–765. doi: 10.1161/01.str.31.3.760. [DOI] [PubMed] [Google Scholar]
- 23.Faraci FM, Lamping KG, Modrick ML, Ryan MJ, Sigmund CD, Didion SP. Cerebral vascular effects of angiotensin II: new insights from genetic models. Journal of Cerebral Blood Flow and Metabolism. 2006;26(4):449–455. doi: 10.1038/sj.jcbfm.9600204. [DOI] [PubMed] [Google Scholar]
- 24.Lanz TV, Ding Z, Ho PP, et al. Angiotensin II sustains brain inflammation in mice via TGF-β . Journal of Clinical Investigation. 2010;120(8):2782–2794. doi: 10.1172/JCI41709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Liu G, Hosomi N, Hitomi H, et al. Angiotensin II induces human astrocyte senescence through reactive oxygen species production. Hypertension Research. 2011;34(4):479–483. doi: 10.1038/hr.2010.269. [DOI] [PubMed] [Google Scholar]
- 26.Yamada K, Horita T, Takayama M, et al. Effect of a centrally active angiotensin converting enzyme inhibitor, perindopril, on cognitive performance in chronic cerebral hypo-perfusion rats. Brain Research. 2011;1421(1):10–120. doi: 10.1016/j.brainres.2011.09.016. [DOI] [PubMed] [Google Scholar]
- 27.Yamada K, Uchida S, Takahashi S, et al. Effect of a centrally active angiotensin-converting enzyme inhibitor, perindopril, on cognitive performance in a mouse model of Alzheimer’s disease. Brain Research C. 2010;1352:176–186. doi: 10.1016/j.brainres.2010.07.006. [DOI] [PubMed] [Google Scholar]
- 28.Van Nostrand WE, Davis-Salinas J, Saporito-Irwin SM. Amyloid β-protein induces the cerebrovascular cellular pathology of Alzheimer’s disease and related disorders. Annals of the New York Academy of Sciences. 1996;777:297–302. doi: 10.1111/j.1749-6632.1996.tb34436.x. [DOI] [PubMed] [Google Scholar]
- 29.Hardy J, Selkoe DJ. The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics. Science. 2002;297(5580):353–356. doi: 10.1126/science.1072994. [DOI] [PubMed] [Google Scholar]
- 30.Zou K, Michikawa M. Angiotensin-converting enzyme as a potential target for treatment of Alzheimer’s disease: inhibition or activation? Reviews in the Neurosciences. 2008;19(4-5):203–212. doi: 10.1515/revneuro.2008.19.4-5.203. [DOI] [PubMed] [Google Scholar]
- 31.Wang BR, Shi JQ, Zhang YD, Zhu DL, Shi JP. Angiotensin II does not directly affect Aβ secretion or β-/γ-secretase activity via activation of angiotensin II type 1 receptor. Neuroscience Letters. 2011;500(2):103–107. doi: 10.1016/j.neulet.2011.06.014. [DOI] [PubMed] [Google Scholar]
- 32.Wang J, Ho L, Chen L, et al. Valsartan lowers brain β-amyloid protein levels and improves spatial learning in a mouse model of Alzheimer disease. Journal of Clinical Investigation. 2007;117(11):3393–3402. doi: 10.1172/JCI31547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Tsukuda K, Mogi M, Iwanami J, et al. Cognitive deficit in amyloid-β-injected mice was improved by pretreatment with a low dose of telmisartan partly because of peroxisome proliferator-activated receptor-γ activation. Hypertension. 2009;54(4):782–787. doi: 10.1161/HYPERTENSIONAHA.109.136879. [DOI] [PubMed] [Google Scholar]
- 34.Danielyan L, Klein R, Hanson LR, et al. Protective effects of intranasal losartan in the APP/PS1 transgenic mouse model of Alzheimer disease. Rejuvenation Research. 2010;13(2-3):195–201. doi: 10.1089/rej.2009.0944. [DOI] [PubMed] [Google Scholar]
- 35.Dong YF, Kataoka K, Tokutomi Y, et al. Perindopril, a centrally active angiotensin-converting enzyme inhibitor, prevents cognitive impairment in mouse models of Alzheimer’s disease. FASEB Journal. 2011;25(9):2911–2920. doi: 10.1096/fj.11-182873. [DOI] [PubMed] [Google Scholar]
- 36.Hemming ML, Selkoe DJ. Amyloid β-protein is degraded by cellular angiotensin-converting enzyme (ACE) and elevated by an ACE inhibitor. Journal of Biological Chemistry. 2005;280(45):37644–37650. doi: 10.1074/jbc.M508460200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zou K, Maeda T, Watanabe A, et al. Aβ42-to-Aβ40- and angiotensin-converting activities in different domains of angiotensin-converting enzyme. Journal of Biological Chemistry. 2009;284(46):31914–31920. doi: 10.1074/jbc.M109.011437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wharton W, Stein JH, Korcarz C, et al. The effects of ramipril in individuals at risk for Alzheimer’s disease: results of a pilot clinical trial. Journal of Alzheimer’s Disease. 2012;32(1):147–156. doi: 10.3233/JAD-2012-120763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bhojak TJ, DeKosky ST, Ganguli M, Kamboh MI. Genetic polymorphism in the cathepsin G gene and the risk of Alzheimer’s disease. Neuroscience Letters. 2001;309(2):138–140. doi: 10.1016/s0304-3940(01)02033-x. [DOI] [PubMed] [Google Scholar]
- 40.Kadir A, Almkvist O, Wall A, Långström B, Nordberg A. PET imaging of cortical 11C-nicotine binding correlates with the cognitive function of attention in Alzheimer’s disease. Psychopharmacology. 2006;188(4):509–520. doi: 10.1007/s00213-006-0447-7. [DOI] [PubMed] [Google Scholar]
- 41.Buckingham SD, Jones AK, Brown LA, Sattelle DB. Nicotinic acetylcholine receptor signalling: roles in alzheimer’s disease and amyloid neuroprotection. Pharmacological Reviews. 2009;61(1):39–61. doi: 10.1124/pr.108.000562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wang HY, Lee DHS, D’Andrea MR, Peterson PA, Shank RP, Reitz AB. β-Amyloid1-42 binds to α7 nicotinic acetylcholine receptor with high affinity. Implications for Alzheimer’s disease pathology. Journal of Biological Chemistry. 2000;275(8):5626–5632. doi: 10.1074/jbc.275.8.5626. [DOI] [PubMed] [Google Scholar]
- 43.Parri HR, Hernandez CM, Dineley KT. Research update: alpha7 nicotinic acetylcholine receptor mechanisms in Alzheimer’s disease. Biochemical Pharmacology. 2011;82(8):931–942. doi: 10.1016/j.bcp.2011.06.039. [DOI] [PubMed] [Google Scholar]
- 44.Shaw S, Bencherif M, Marrero MB. Angiotensin II blocks nicotine-mediated neuroprotection against β-amyloid (1–42) via activation of the tyrosine phosphatase SHP-1. Journal of Neuroscience. 2003;23(35):11224–11228. doi: 10.1523/JNEUROSCI.23-35-11224.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Marrero MB, Bencherif M. Convergence of alpha 7 nicotinic acetylcholine receptor-activated pathways for anti-apoptosis and anti-inflammation: central role for JAK2 activation of STAT3 and NF-κB. Brain Research C. 2009;1256:1–7. doi: 10.1016/j.brainres.2008.11.053. [DOI] [PubMed] [Google Scholar]
- 46.Seguin LR, Villarreal RS, Ciuffo GM. AT2 receptors recruit c-Src, SHP-1 and FAK upon activation by ang II in PND15 rat hindbrain. Neurochemistry International. 2012;60(2):199–207. doi: 10.1016/j.neuint.2011.11.008. [DOI] [PubMed] [Google Scholar]
- 47.Li JM, Mogi M, Tsukuda K, et al. Angiotensin II-induced neural differentiation via angiotensin II type 2 (AT2) receptor-MMS2 cascade involving interaction between AT 2 receptor-interacting protein and Src homology 2 domain-containing protein-tyrosine phosphatase 1. Molecular Endocrinology. 2007;21(2):499–511. doi: 10.1210/me.2006-0005. [DOI] [PubMed] [Google Scholar]
- 48.AbdAlla S, Lother H, El Missiry A, et al. Angiotensin II AT2 receptor oligomers mediate G-protein dysfunction in an animal model of Alzheimer disease. Journal of Biological Chemistry. 2009;284(10):6554–6565. doi: 10.1074/jbc.M807746200. [DOI] [PubMed] [Google Scholar]
- 49.Tienari PJ, De Strooper B, Ikonen E, et al. The β-amyloid domain is essential for axonal sorting of amyloid precursor protein. EMBO Journal. 1996;15(19):5218–5229. [PMC free article] [PubMed] [Google Scholar]
- 50.Iadecola C. The overlap between neurodegenerative and vascular factors in the pathogenesis of dementia. Acta Neuropathologica. 2010;120(3):287–296. doi: 10.1007/s00401-010-0718-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Girouard H, Park L, Anrather J, Zhou P, Iadecola C. Angiotensin II attenuates endothelium-dependent responses in the cerebral microcirculation through nox-2-derived radicals. Arteriosclerosis, Thrombosis, and Vascular Biology. 2006;26(4):826–832. doi: 10.1161/01.ATV.0000205849.22807.6e. [DOI] [PubMed] [Google Scholar]
- 52.Capone C, Faraco G, Park L, Cao X, Davisson RL, Iadecola C. The cerebrovascular dysfunction induced by slow pressor doses of angiotensin II precedes the development of hypertension. American Journal of Physiology-Heart and Circulatory Physiology. 2011;300(1):H397–H407. doi: 10.1152/ajpheart.00679.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Girouard H, Lessard A, Capone C, Milner TA, Iadecola C. The neurovascular dysfunction induced by angiotensin II in the mouse neocortex is sexually dimorphic. American Journal of Physiology-Heart and Circulatory Physiology. 2008;294(1):H156–H163. doi: 10.1152/ajpheart.01137.2007. [DOI] [PubMed] [Google Scholar]
- 54.Laughlin GA, McEvoy LK, von Muhlen D, et al. Sex differences in the association of Framingham cardiac risk score with cognitive decline in community-dwelling elders without clinical heart disease. Psychosomatic Medicine. 2011;73(8):683–689. doi: 10.1097/PSY.0b013e31822f9089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Takeda S, Sato N, Takeuchi D, et al. Angiotensin receptor blocker prevented β-amyloid-induced cognitive impairment associated with recovery of neurovascular coupling. Hypertension. 2009;54(6):1345–1352. doi: 10.1161/HYPERTENSIONAHA.109.138586. [DOI] [PubMed] [Google Scholar]
- 56.Zhang M, Mao Y, Ramirez SH, Tuma RF, Chabrashvili T. Angiotensin II induced cerebral microvascular inflammation and increased blood-brain barrier permeability via oxidative stress. Neuroscience. 2010;171(3):852–858. doi: 10.1016/j.neuroscience.2010.09.029. [DOI] [PubMed] [Google Scholar]
- 57.Min LJ, Mogi M, Iwanami J, et al. Angiotensin II and aldosterone-induced neuronal damage in neurons through an astrocyte-dependent mechanism. Hypertension Research. 2011;34(6):773–778. doi: 10.1038/hr.2011.38. [DOI] [PubMed] [Google Scholar]
- 58.Horiuchi M, Mogi M, Iwai M. The angiotensin II type 2 receptor in the brain. Journal of the Renin-Angiotensin-Aldosterone System. 2010;11(1):1–6. doi: 10.1177/1470320309347793. [DOI] [PubMed] [Google Scholar]
- 59.Iwai M, Liu HW, Chen R, et al. Possible inhibition of focal cerebral ischemia by angiotensin II type 2 receptor stimulation. Circulation. 2004;110(7):843–848. doi: 10.1161/01.CIR.0000138848.58269.80. [DOI] [PubMed] [Google Scholar]
- 60.Mogi M, Li JM, Iwanami J, et al. Angiotensin II type-2 receptor stimulation prevents neural damage by transcriptional activation of methyl methanesulfonate sensitive 2. Hypertension. 2006;48(1):141–148. doi: 10.1161/01.HYP.0000229648.67883.f9. [DOI] [PubMed] [Google Scholar]
- 61.Reinecke K, Lucius R, Reinecke A, Rickert U, Herdegen T, Unger T. Angiotensin II accelerates functional recovery in the rat sciatic nerve in vivo: role of the AT2 receptor and the transcription factor NF-kappaB. FASEB Journal. 2003;17(14):2094–2096. doi: 10.1096/fj.02-1193fje. [DOI] [PubMed] [Google Scholar]
- 62.Li J, Culman J, Hörtnagl H, et al. Angiotensin AT2 receptor protects against cerebral ischemia-induced neuronal injury. FASEB Journal. 2005;19(6):617–619. doi: 10.1096/fj.04-2960fje. [DOI] [PubMed] [Google Scholar]
- 63.Gendron L, Laflamme L, Rivard N, Asselin C, Payet MD, Gallo-Payet N. Signals from the AT2 (angiotensin type 2) receptor of angiotensin II inhibit p21(ras) and activate MAPK (mitogen-activated protein kinase) to induce morphological neuronal differentiation in NG108-15 cells. Molecular Endocrinology. 1999;13(9):1615–1626. doi: 10.1210/mend.13.9.0344. [DOI] [PubMed] [Google Scholar]
- 64.Cote F, Laflamme L, Payet MD, Gallo-Payet N. Nitric oxide, a new second messenger involved in the action of angiotensin II on neuronal differentiation of NG108-15 cells. Endocrine Research. 1998;24(3-4):403–407. doi: 10.3109/07435809809032622. [DOI] [PubMed] [Google Scholar]
- 65.Côté F, Do TH, Laflamme L, Gallo JM, Gallo-Payet N. Activation of the AT2 receptor of angiotensin II induces neurite outgrowth and cell migration in microexplant cultures of the cerebellum. Journal of Biological Chemistry. 1999;274(44):31686–31692. doi: 10.1074/jbc.274.44.31686. [DOI] [PubMed] [Google Scholar]
- 66.Jing F, Mogi M, Sakata A, et al. Direct stimulation of angiotensin II type 2 receptor enhances spatial memory. Journal of Cerebral Blood Flow and Metabolism. 2011;32(2):248–255. doi: 10.1038/jcbfm.2011.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Jöhren O, Dendorfer A, Dominiak P. Cardiovascular and renal function of angiotensin II type-2 receptors. Cardiovascular Research. 2004;62(3):460–467. doi: 10.1016/j.cardiores.2004.01.011. [DOI] [PubMed] [Google Scholar]