

Abstract
Understanding how the mucosal immune system in the human female reproductive tract might prevent or facilitate HIV infection has important implications for the design of effective interventions. We and others have established cohorts of highly-exposed, HIV-seronegative individuals, such as HIV-uninfected commercial sex workers, who have remained HIV-negative after more than 5 years of active prostitution. Observations obtained in studies of such individuals, who represent a model of natural immunity to HIV, indicate that HIV resistance may be associated with the host's capacity to preserve systemic integrity by constraining immune activity and controlling inflammatory conditions at the mucosal point of entry. This likely necessitates the orchestration of balanced, first-line and adaptive immune responses.
1. Introduction
At the end of 2010, 34 million people were living with HIV/AIDS world-wide. In that year, a total of 2.7 million people were infected by HIV, mostly through heterosexual intercourse, and 60% of new HIV infections affected women in sub-Saharan Africa [1]. Needless to say, the design of effective vaccines and microbicides to prevent HIV infection remains a global priority. High levels of anti-inflammatory and neutralizing proteins, such as antiproteases and HIV-specific immunoglobulins (Ig), are found in the genital mucosa of highly exposed HIV-seronegative (HESN) individuals, such as HIV-uninfected, “resistant” commercial sex workers (CSWs) [2, 3]. This suggests that efforts to develop effective microbicides and vaccines should aim at mimicking and/or soliciting innate and adaptive immune responses, such as those seen in the context of natural immunity to HIV. From such a viewpoint, vaccine approaches to specifically induced mucosal responses seem very promising. Indeed, genital IgA and IgG, elicited through combined intra-muscular and intranasal vaccination against HIV-gp41, delivered via virosome in nonhuman primates, prevented systemic HIV invasion by blocking transcytosis and by mediating antibody-dependent cellular cytotoxicity (ADCC) [4]. These animals lacked serum-neutralizing antibody activity, highlighting the role of effector antibodies at the mucosal point of entry, and their importance in preventing the dissemination of HIV infection [5]. In humans, the RV144 vaccine regimen (canarypox prime, HIV gp120 envelope (Env) glycoprotein boost) elicits protective responses, the nature of which remains to be defined in terms of generation and effector mechanisms [6]. Reduced rates of HIV acquisition without significant effects on initial viral loads or CD4 T-cell counts have led to the hypothesis of a transient, protective B-cell response. Moreover, binding of IgG antibodies to variable regions 1 and 2 (V1, V2) of Env has been shown to be inversely correlated with HIV infection rates [7]. Unfortunately, mucosal samples were not collected during the RV144 trial to assess mucosal Env-specific Ig levels, which we predict may constitute better correlates of protection. Success in conceiving effective vaccines most likely relies on their capacity to establish rapid, first-line immune responses at the mucosal point of entry as well as long-term protection, which operates both at the mucosal and systemic levels.
A better understanding of the mechanisms of transmission and HIV-specific immune responses at the initial site of infection is therefore pivotal to the design of preventive strategies. Most observations relating to these events have been obtained with simian immunodeficiency virus (SIV) infection in nonhuman primates (reviewed in [8, 9]). In humans, findings in HESN individuals, such as HIV-uninfected CSWs, who represent a model of natural immunity to HIV, may thus yield important clues to the development of preventive approaches. As such, the current perspective on cumulative data, reported by us and others, supports the notion that HIV “resistance” in these highly exposed CSWs may be associated with their capacity to control genital inflammatory conditions and recruitment of HIV target cells at the initial site of infection. This could be achieved by locally constraining immune activity to mucosal sites and preserving peripheral integrity, a process that likely involves genetic factors and orchestration of strong innate and adaptive immune responses.
2. Immunology of the Female Genital (FGT)
FGT immunology has been reviewed recently [10] and will only be summarized here briefly. The FGT is subdivided into 3 major areas presenting distinct phenotypic profiles: the nonsterile vagina and ectocervix colonized by commensal microflora, the sterile endometrium and fallopian tubes, and the endocervix in which sterility may be temporally related to menstrual cycle phase. Thus, FGT immunity is tightly regulated by a hormonal/inflammatory process throughout the menstrual cycle, having to deal with the pressure of procreation and microbial control. The innate immune compartment of the FGT involves the mucous lining of a tight epithelial cell (EC) barrier, stratified at the vaginal and ectocervical levels, as well as dendritic cells (DCs), Langerhans cells (LCs), macrophages, natural killer (NK) cells, and neutrophils, which confer protection through the production of antimicrobial agents, chemokines, and cytokines [10, 11] (Figure 1). Control of flora and invading pathogens is modulated via pattern recognition receptors (PRRs), such as toll-like receptors (TLRs) and NOD-like receptors, which recognise specific common microbial/pathogen-associated molecular patterns. As such, genital ECs form an uninterrupted barrier between the lumen and underlying cells and express PRRs, such as TLR-1 to -9, indicating the potential to respond to a wide range of microbes/pathogens [10–14]. DCs also express a large spectrum of PRRs, and the interaction between ECs and submucosal DCs likely modulates the maintenance of homeostatic balance between tolerance and inflammation in the FGT [10–15]. FGT-associated lymphoid organs are part of mucosal-associated lymphoid tissue, (MALT) which also includes gastrointestinal lymphoid tissue (GALT). Unlike GALT, the FGT does not contain M cells or organised lymphoid crypts or follicles in the submucosa. Rather, the upper FGT contains unique lymphoid aggregates constituted of CD8+ T cells that surround a central B cell core, which are encapsulated by macrophages [10] (Figure 1). Immunisation at the FGT level has been shown to elicit local CD8+ cytotoxic T lymphocyte (CTL), IgG, and IgA responses. Although immune induction mechanisms in the FGT remain poorly understood, it is likely that DCs migrate to FGT mucosal-associated lymphoid structures to induce first-line B-cell responses and to regulate adaptive lymphocyte responses [10, 16–18]. Interestingly, detailed characterisation of the Ig repertoire of cervical and systemic B cells from a HESN individual in Kenya disclosed that site-specific responses occur with unique regulation of tolerance and recruitment into local memory or blast B cell compartments. Also, the infusion of systemic post-germinal center B cells to the human cervix seems to be a common event [19]. These findings suggest that cervical B cell populations largely contribute to protection against HIV, by producing first-line and mature mucosal HIV-specific IgG and IgA, which are correlates of control “resistance” to HIV infection in the FGT of HESN women. Understanding how B cell populations are recruited and maintained in the FGT is crucial for the design of preventive approaches, to block infection by HIV at its main point of entry.
Figure 1.
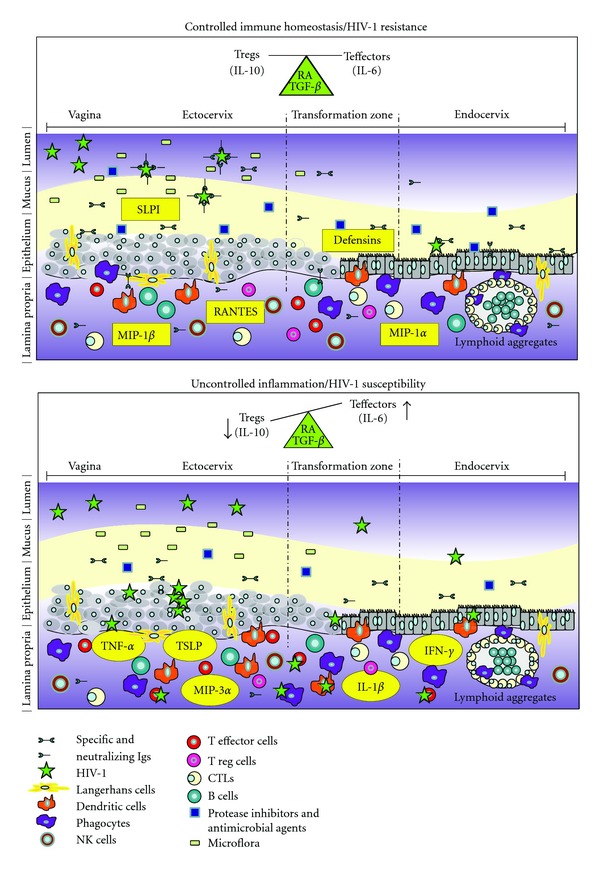
Qualitative and quantitative differences in mucosal innate and adaptive immune components are associated with the outcome of HIV infection in the female genital tract (FGT).
3. HIV Transmission in the FGT
Until now, the cascade of events leading to HIV infection after heterosexual transmission remains unclear. Several reports in humans and rhesus macaques suggest that LCs and DCs on mucosal surfaces are the earliest cell types to be exposed, and possibly infected by HIV or SIV, and migrate to the lamina propria and draining lymphoid tissues to facilitate transmission of the virus to permissive cells (reviewed in [8, 9, 20–23]). The most likely scenario has been proposed recently by the group of Haase [9, 24]. After genital administration to macaques, SIV establishes foci of infection in the vaginal sub-mucosa within a matter of days through a scheme involving macrophage inflammatory protein-3alpha (MIP-3α) (CCL20) production by ECs and early recruitment of interferon-alpha (IFN-α) producing plasmacytoid DC (pDC) as well as DC and CD4+ T-cell effectors, rapidly accessing draining lymph nodes and establishing systemic invasion by days 10–14. Recent studies with genital explants indicate that LCs can elaborate protrusions across the stratified epithelium into the lumen of the FGT to capture HIV mainly through the surface expression of langerin (CD207) [8, 20–23]. LCs may then enable HIV transmission to sub-mucosal DCs that express PRRs, such as DC-SIGN (CD209), and to CD4+ CCR5+ effector target T lymphocytes and/or migrate to draining lymphoid tissues. DCs are also thought to have the capacity to establish protrusions across the epithelium, enabling direct HIV transmission to permissive populations in the lamina propria or after their migration to draining lymphoid compartments [8, 20–23].
Although HIV does not productively infect ECs, it can be transcytosed, reaching sub-mucosal DC populations and effector target T lymphocytes in the lamina propria [25–30]. Furthermore, HIV likely facilitates its incursion through the genital epithelium by inducing a proinflammatory milieu that affects tight junction proteins and enhances microbial translocation [31]. HIV has also been shown to be internalized by FGT ECs via gp340, a scavenger receptor, subsequently promoting the production of proinflammatory thymic stromal lymphopoietin (TSLP) via TLR-7 signalling, which then activates DCs and promotes HIV transmission to CD4+ T cells [32, 33]. The galactosyl ceramide receptor induces HIV endocytosis in ECs and DCs, allowing transcytosis and transfer to susceptible CD4+ T cells [34].
HIV acquisition may depend on the level of inflammation and availability/permissiveness of target populations, such as activated CD4+ T cells expressing CCR5 and α4β7 [35, 36]. In human peripheral blood, CD4+ T cells expressing CCR6+ are most permissive to HIV infection [37, 38]. In the gut, which represents a major viral reservoir, mucosal Th17 effectors are the main targets of HIV/SIV [39–42]. These mucosal Th17 cells are mostly α4β7+ CD103+, express RORγt+ and have been reported to require factors, such as transforming growth factor-beta (TGF-β), interleukin (IL)-1, IL-6, IL-21, and IL-23, for their differentiation [43]. They express CCR6, a major ligand of MIP-3α, which is mainly secreted by mucosal ECs and is known to also attract immature LCs and DCs [44, 45]. It has been determined that homeostatic balance between mucosal T effector versus T regulatory (Treg) populations in the gut is modulated by EC and DC cross-talk, and is highly influenced by factors, such as retinoic acid (RA) and TGF-β [46, 47] (Figure 1). TGF-β is also known to influence FGT integrity [48, 49], and RA is involved in the regulation of ovarian function and FGT immune status by its modulatory effect on sexual hormones [50]. Also, oestrogen can upregulate RA and TGF-β production and signalling in the human endometrium [51, 52].
Thus, ECs and DCs appear to play a critical role in HIV infection by sensing through PRRs and orchestrating the dynamics of cellular populations, inflammatory conditions, and adaptive immune responses. The fact that TLR expression and responsiveness are increased in viraemic HIV infections suggests that TLR modulation is likely to influence HIV infection [53]. From this viewpoint, modulation of inflammatory responses through TLR agonists is a promising therapeutic approach in diseases with an imbalance in T cell responses, such as allergy and asthma, and could be seen as impacting inflammatory conditions and immune status in the FGT, the mucosal point of entry for the virus.
4. Nonpathogenic SIV Infections Provide Novel Insights into the Pathogenesis of Human HIV Infection
Similar to pathogenic HIV and SIV infections in susceptible hosts, SIV infections in the natural host Sooty mangabeys result in high viral replication and massive depletion of gut mucosal effector CD4+ T cells [54]. However, a major distinction from pathogenic infection is the rapidly developing anti-inflammatory milieu that prevents chronic activation, apoptosis, and proliferation of T cells in SIV-infected Sooty mangabeys. This contributes to the maintenance of mucosal barrier integrity, preventing microbial translocation from the gut, which is the hallmark of pathogenic infections [55]. The control of disease progression appears to be linked to better management of aberrant immune activation caused by SIV infection. Indeed, the early onset of anti-inflammatory IL-10 production and Treg activity seems to be favoured in SIV nonpathogenic infections [54]. Furthermore, it was recently demonstrated that Sooty mangabeys generated less Th17 effector target cells than highly-susceptible macaques [40]. Importantly, the capacity to manage inflammatory conditions in Sooty mangabeys is associated with a low type I IFN gene profile. The latter appears to be linked to genetic polymorphisms in the type I IFN regulatory factor-7 (IRF-7) gene involved in the regulation of IFN production downstream of TLR-7 and -9 signalling, which are intracellular ligands for lentiviral ssRNA viruses, such as SIV and HIV, and CpG DNA, respectively [56]. Moreover, early blocking of MIP-3α and pro-inflammatory cytokines in the FGT of SIV-susceptible macaques prevented cellular recruitment, establishment of an inflammatory milieu, and infection despite repeated intravaginal exposure to high SIV doses [24]. Therefore, low inflammatory conditions are beneficial to the host in the context of HIV/SIV, and we believe that preventive approaches, such as microbicides, should be designed to induce and maintain a low inflammatory milieu.
5. Factors Associated with Susceptibility/Resistance to HIV Infection
The number of sexual partners and failure to use condoms are the best documented behavioural risk factors for sexual HIV transmission. Among the most compelling biological risk factors are the presence of vaginosis and sexually-transmitted infections, high viral load and low CD4+ T lymphocyte counts in infectious contact, and possibly viral virulence and tropism (reviewed in [57]). In Figure 1, top panel: controlled immune homeostasis results in resistance to HIV infection at the mucosal point of entry. Homeostatic balance between mucosal T effector versus T regulatory (Treg) populations is modulated by epithelial cell (EC) and dendritic cell (DC) cross-talk and is influenced by factors, such as retinoic acid (RA) and transforming growth factor-beta (TGF-β). Host factors associated with resistance to HIV infection involve the modulation of mucosal innate factors, such as defensins, secretory leukocyte proteaseinhibitor (SLPI), and other antiproteases as well as variations in frequencies and activities of DC, B, T, and natural killer (NK) cell populations [2, 58] (Table 1). HIV-specific mucosal IgA blocks viral transcytosis through the epithelium, and IgG is involved in antibody-dependent cellular cytotoxicity(ADCC). High levels of β-chemokines, such as macrophage inflammatory protein-1alpha (MIP-1α), MIP-1β and regulated upon activation, normal T-cell expressed and secreted (RANTES), which are natural CCR5 (major HIV coreceptor) ligands, can block cell viral entry to the FGT mucosa. Bottom panel: uncontrolled inflammation results in susceptibility to HIV infection at the mucosal point of entry. Unbalanced ratios between mucosal T effector versus Treg populations favouring high levels of T effectors are the hallmark of an inflammatory environment. Established vaginal inflammation can facilitate HIV infection through epithelium damage and recruitment of CD4+ T effectors, prime targets of HIV. The virus can also bind to ECs, be captured by Langerhans cells or DCs, and subsequently transcytosed and transferred to productively infect CD4+ target cells. Host factors associated with susceptibility to HIV infection are inflammatory markers, such as tumour necrosis factor-alpha (TNF-α), interferon-gamma (IFN-γ), IFN-α, interleukin-1beta (IL-1β), as well as thymic stromal lymphopoietin (TSLP) and MIP-3α secreted primarilyby ECs after downstream HIV signalling, favouring the recruitment of plasmacytoid DC (pDC) and CD4+ target cells. The promotion of an inflammatory milieu will contribute to infection and dissemination of HIV across the genital tract.
Table 1.
Genetic and genital mucosa host factors associated with resistance to HIV-1 infection in several HESN cohorts.
| HIV-resistant host factors | |
|---|---|
| Genetic | Genital mucosa |
| KIR3D L1/S1 [59] | Protease inhibitors (SLPI, lactoferrin, serpins, cystatins, trappin-2/elafin) [2, 84–87] |
| KIR2D L2/L3 [60] | |
| HLA class 1 alleles [61–65] | Defensins (α, β) [2] |
| IRF-1 [66] | CC-β chemokines [9, 79, 80] |
| TLR 9 [67] | APOBEC3G, TRIM5α, tetherin, SAMHD1, LEDGF/p75 [81–83] |
| CCR5Δ32 [68–76] | Elevated DC and NK cell frequencies/activities [58] |
| CCR2b [77, 78] | CD4+- and CD8+-specific immune responses and reduced T-cell |
| MIP-1α [79] | activation [2, 88–97] |
| MIP-1β [80] |
Cross-clade neutralizing specific IgA (transcytosis inhibition and ADCC activities) [2, 88–94, 98, 99] |
ADCC: antibody-dependent cellular cytotoxicity; APOBEC: apolipoprotein B mRNA-editing catalytic polypeptide-like; CTL: cytotoxic T lymphocyte; DC: dendritic cell; HESN: highly-exposed HIV-seronegative; HLA: human leucocyte antigen; IRF: interferon-regulating factor; KIR: killing inhibitory receptor; LEDGF: Lens epithelium-derived growth factor; MIP: macrophage inflammatory protein; NK: natural killer; SAMHD: sterile alpha motif and HD domain; SLPI: secretory leukocyte protease inhibitor; TLR: Toll-like receptor; TRIM: tripartite motif.
NK cells represent a critical component of the host innate immune response against viral infections. The killing inhibitory receptor (KIR)3DL1/S1 locus has been linked with both slow progression to AIDS and resistance to HIV infection in a high-risk cohort of i.v. drug users from Montreal [59], and the KIR2DL2/DL3 locus has also been associated with resistance to HIV infection among African CSWs [60]. Functional modulation of NK cell responses (IFN-γ), NK activation (CD69), and degranulation (CD107a) markers has been correlated with resistance to HIV infection in several independent cohorts of HESN individuals [58]. Human leukocyte antigen (HLA) alleles, which are KIR ligands, are also associated with susceptibility/resistance to HIV infection and disease progression [61–65]. Other factors include IRF-1 [66], TLR-9 [67], and chemokine receptor/ligand polymorphisms, such as CCR5 [68–76], CCR2b [77, 78], CCL3 (MIP-1α) [79], and CCL4 (MIP-1β) [80]. Viral restriction factors, such as apolipoprotein B mRNA-editing catalytic polypeptide-like (APOBEC) 3G, tripartite motif (TRIM) 5α, tetherin, and sterile alpha motif and HD domain 1 (SAMHD1), exert anti-HIV activity. Lens epithelium-derived growth factor (LEDGF/p75) may also contribute to HIV resistance [81, 82]. Indeed, relatively low levels of LEDGF/p75 occurred in blood CD4+ T lymphocytes of HESN subjects enrolled in a Senegalese cohort of HIV-serodiscordant couples [83].
In a prospective cohort study of female CSWs in Nairobi, Kenya, over a 13-year period, a small group of women were found to be persistently IgG-seronegative and resistant to infection [102]. HIV resistance in this cohort has been associated with factors, such as trappin-2/elafin [84–86], serpins and cystatins in genital samples [87], certain HLA class I and II alleles [62], IRF-1 polymorphisms [66], and HIV-specific immune responses. Indeed, HIV-specific CD4+ T cell and CD8+ CTL responses as well as cross-clade neutralizing IgA have been encountered in both the blood and genital tract of resistant women [2, 88–94]. HIV-resistant CSWs from the Kenyan cohort had increased cervical CD4+ T cell counts compared to HIV-infected CSWs [92]. Moreover, CD4+ T cells in HIV-resistant women had a low activation profile but a much greater ability to proliferate in response to HIV p24 peptides than HIV-infected CSWs [93]. Also, in resistant women, higher levels of HIV-specific CTLs were noted in the cervix than in blood [89]. Recent studies have demonstrated that the quality of T cell responses in the context of HIV may be a major determinant of disease progression [95–97]. In a cohort of HESN women from the Ivory Coast, HIV-specific mucosal IgA was shown to block viral transcytosis through tight epithelial barriers [98, 99]. Thus, HIV-specific immune responses in CSWs prevail in the FGT and may be important in preventing heterosexual HIV infection. Interestingly, there is a clear indication of clustering of both resistance and HIV-specific CTL responses among HIV-resistant CSWs, suggesting that genetic factors could be involved in “protective” immune responses [103]. However, the durability and protective efficacy of CTL responses in these subjects are not absolute. Late seroconversion occurred in some HIV-resistant CSWs despite HIV-specific CTL responses [104]. Seroconversion happened in the absence of detectable CTL escape mutations and was related to the waning of HIV-specific CD8+ CTL responses due to reduced sexual activity and thus renewed antigenic exposure. These findings suggest that production and maintenance of HIV-specific effector responses and low-level immune activation may depend on genetically determined genital HIV-specific immune responses induced upon initial contact with HIV and low ongoing viral exposure.
The recent finding that the frequency of immunosuppressive Treg lymphocytes was increased in the blood of HIV-resistant women [105] is consistent with the notion that the host's capacity to control and/or maintain low levels of immune activation may contribute to protection against infection. According to this view, in a cohort of highly HIV-exposed Beninese CSWs, we found that HIV-uninfected CSWs had significantly lower genital levels of tumour necrosis factor-alpha (TNF-α) and IFN-γ than HIV-infected CSWs [100] (Table 2). These observations suggest that the capacity to maintain a low-key inflammatory profile at the initial site of exposure is associated with protection against HIV infection in HESN individuals. In contrast, serum IL-2, IL-10, and TNF-α levels were significantly higher in HIV-uninfected CSWs than in HIV-infected CSWs. Importantly, when assessing the serum effector (IL-6) to regulatory (IL-10) ratio, we determined that it was lower in HIV-uninfected CSWs (0.9) than in HIV-infected CSWs (1.5). Moreover, the relatively normal cytokine levels found in the serum of HIV-uninfected CSWs (similar to nonexposed women from the general population) may be reflective of their capacity to maintain integrity of the systemic immune compartment by stopping HIV dissemination beyond the genital tract. In contrast, the low levels of cytokines observed in the serum of HIV-infected CSWs could reflect active recruitment of cytokine-producing cells to the genital mucosa in response to HIV infection. In this respect, HIV-infected CSWs had significantly higher blood and genital levels of monocyte chemotactic protein-3 (MCP-3) and monokine induced by gamma interferon (MIG) compared to both HIV-uninfected CSW and non-CSW groups [101]. In the HIV-infected group, MCP-3 and MIG levels were significantly higher in the genital mucosa than in blood, indicating a chemotactic gradient favouring the recruitment of immune cells contributing to the mucosal inflammatory response observed in these women. However, HIV-uninfected CSWs had significantly higher MIP-1α levels in the genital mucosa than both HIV-infected CSWs and HIV-uninfected non-CSW women. Moreover, serum MIP-1α (CCL3) and MIP-1β (CCL4) levels were higher in HIV-uninfected CSWs than the other groups. Interestingly, MIP-1α and MIP-1β are natural ligands of the HIV coreceptor CCR5, and high copy numbers of CCL3 and CCL4 genes have been previously associated with lower risk of HIV infection [79, 80], possibly by competing/blocking viral entry mediated by the co-receptor CCR5 [9]. Finally, recent evidence indicates that local microflora may also play a pivotal role in shaping host immune responses [2, 106], and thus may be a potential ally in the modulation of a mucosal immune compartment favourable to the maintenance of low inflammatory conditions. Thus, the immune events involved in natural immunity “resistance” to HIV may share some similarities with those associated with the control of the mucosal commensal microflora, which are thought to involve mucosal Igs and balanced Treg/T effector responses in absence of inflammation, and which cellular niches are maintained by repeated antigenic exposure, such as likely encountered by HESN individuals.
Table 2.
Cytokines and chemokines significantly associated with resistance to HIV-1 infection in the Beninese HIV-1-uninfected and infected CSW cohort.
| Cytokines/chemokines | HIV-resistant CSWs | HIV-infected CSWs |
|---|---|---|
| Genital mucosa | ||
| TNF-α | ↓ | ↑ |
| IFN-γ | ↓ | ↑ |
| MIP-1α | ↑↑ | ↓ |
| MCP-3 | ↓ | ↑↑ |
| MIG | ↓ | ↑↑ |
| Blood | ||
| IL-2 | ↑ | ↓ |
| IL-10 | ↑ | ↓ |
| TNF-α | ↑ | ↓ |
| MIP-1α | ↑ | ↓ |
| MIP-1β | ↑ | ↓ |
| MCP-3 | ↓ | ↑ |
| MIG | ↓ | ↑ |
6. Conclusion
Overall, resistance in the context of HIV infection may be associated with the host's capacity to induce a strong innate and HIV-specific immune response and, at the same time, control/maintain low inflammatory conditions and fewer HIV target cells at the initial exposure site (Figure 1). Understanding how the mucosal immune system in the human FGT might prevent or facilitate HIV infection has important implications for the design of effective interventions and may help develop strategies to modulate mucosal inflammatory conditions, to establish quick, long-lasting, first-line mucosal defence against HIV.
Funding
This work was supported by Grants from the Canadian Institutes of Health Research (no. MOP-119406) and the Réseau SIDA et Maladies Infectieuses, Fonds de la recherche en santé du Québec (FRSQ).
Acknowledgment
The authors are thankful to Mr. Christian Charbonneau for his graphic arts expertise and to Mr. Ovid Da Silva for medical editing.
References
- 1. http://www.unaids.org/globalreport/Global_report.htm.
- 2.Schellenberg JJ, Plummer FA. The microbiological context of HIV resistance: vaginal microbiota and mucosal inflammation at the viral point of entry. International Journal of Inflammation. 2012;2012:10 pages. doi: 10.1155/2012/131243.131243 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Moir S, Fauci AS. B cells in HIV infection and disease. Nature Reviews Immunology. 2009;9(4):235–245. doi: 10.1038/nri2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bomsel M, Tudor D, Drillet AS, et al. Immunization with HIV-1 gp41 subunit virosomes induces mucosal antibodies protecting nonhuman primates against vaginal SHIV challenges. Immunity. 2011;34(2):269–280. doi: 10.1016/j.immuni.2011.01.015. [DOI] [PubMed] [Google Scholar]
- 5.McElrath MJ. Standing guard at the mucosa. Immunity. 2011;34(2):146–148. doi: 10.1016/j.immuni.2011.02.014. [DOI] [PubMed] [Google Scholar]
- 6.Tomaras GD, Haynes BF. Strategies for eliciting HIV-1 inhibitory antibodies. Current Opinion in HIV and AIDS. 2010;5(5):421–427. doi: 10.1097/COH.0b013e32833d2d45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Haynes BF, Gilbert PB, McElrath MJ, et al. Immune-correlates analysis of an HIV vaccine efficacy trial. The New England Journal of Medicine. 2012:1275–1286. doi: 10.1056/NEJMoa1113425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hladik F, McElrath MJ. Setting the stage: host invasion by HIV. Nature Reviews Immunology. 2008;8(6):447–457. doi: 10.1038/nri2302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Haase AT. Early events in sexual transmission of hiv and siv and opportunities for interventions. Annual Review of Medicine. 2011;62:127–139. doi: 10.1146/annurev-med-080709-124959. [DOI] [PubMed] [Google Scholar]
- 10.Hickey DK, Patel MV, Fahey JV, Wira CR. Innate and adaptive immunity at mucosal surfaces of the female reproductive tract: stratification and integration of immune protection against the transmission of sexually transmitted infections. Journal of Reproductive Immunology. 2011;88(2):185–194. doi: 10.1016/j.jri.2011.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Quayle AJ. The innate and early immune response to pathogen challenge in the female genital tract and the pivotal role of epithelial cells. Journal of Reproductive Immunology. 2002;57(1-2):61–79. doi: 10.1016/s0165-0378(02)00019-0. [DOI] [PubMed] [Google Scholar]
- 12.Horne AW, Stock SJ, King AE. Innate immunity and disorders of the female reproductive tract. Reproduction. 2008;135(6):739–749. doi: 10.1530/REP-07-0564. [DOI] [PubMed] [Google Scholar]
- 13.Triforiova RT, Doncel GF, Fichorova RN. Polyanionic microbicides modify toll-like receptor-mediated cervicovaginal immune responses7. Antimicrobial Agents and Chemotherapy. 2009;53(4):1490–1500. doi: 10.1128/AAC.01152-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gribar SC, Richardson WM, Sodhi CP, Hackam DJ. No longer an innocent bystander: epithelial toll-like receptor signaling in the development of mucosal inflammation. Molecular Medicine. 2008;14(9-10):645–659. doi: 10.2119/2008-00035.Gribar. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Den Dunnen J, Gringhuis SI, Geijtenbeek TBH. Innate signaling by the C-type lectin DC-SIGN dictates immune responses. Cancer Immunology, Immunotherapy. 2009;58(7):1149–1157. doi: 10.1007/s00262-008-0615-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pala P, Gomez-Roman VR, Gilmour J, Kaleebu P. An African perspective on mucosal immunity and HIV-1. Mucosal Immunology. 2009;2(4):300–314. doi: 10.1038/mi.2009.23. [DOI] [PubMed] [Google Scholar]
- 17.Shacklett BL. Immune responses to HIV and SIV in mucosal tissues: ‘location, location, location’. Current Opinion in HIV and AIDS. 2010;5(2):128–134. doi: 10.1097/COH.0b013e328335c178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cerutti A, Puga I, Cols M. Innate control of B cell responses. Trends in Immunology. 2011;32(5):202–211. doi: 10.1016/j.it.2011.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gaudet RG, Breden F, Plummer F, Berry JD. Molecular characterization of the cervical and systemic B-cell repertoire: unique, yet overlapping, immune compartments of an HIV-1 resistant individual. mAbs. 2011;3(2):184–194. doi: 10.4161/mabs.3.2.14858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Derby N, Martinelli E, Robbiani M. Myeloid dendritic cells in HIV-1 infection. Current Opinion in HIV and AIDS. 2011;6:379–384. doi: 10.1097/COH.0b013e3283499d63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.van der Vlist M, van der Aar AMG, Gringhuis SI, Geijtenbeek TBH. Innate signaling in HIV-1 infection of dendritic cells. Current Opinion in HIV and AIDS. 2011;6:348–352. doi: 10.1097/COH.0b013e328349a2d1. [DOI] [PubMed] [Google Scholar]
- 22.Smed-Sörensen A, Loré K. Dendritic cells at the interface of innate and adaptive immunity to HIV-1. Current Opinion in HIV and AIDS. 2011;6:405–410. doi: 10.1097/COH.0b013e328349b06b. [DOI] [PubMed] [Google Scholar]
- 23.Blanchet F, Moris A, Mitchell JP, Piguet V. A look at HIV journey: from dendritic cells to infection spread in CD4+ T cells. Current Opinion in HIV and AIDS. 2011 doi: 10.1097/COH.0b013e328349b0a0. [DOI] [PubMed] [Google Scholar]
- 24.Li Q, Estes JD, Schlievert PM, et al. Glycerol monolaurate prevents mucosal SIV transmission. Nature. 2009;458(7241):1034–1038. doi: 10.1038/nature07831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bouschbacher M, Bomsel M, Verronèse E, et al. Early events in HIV transmission through a human reconstructed vaginal mucosa. AIDS. 2008;22(11):1257–1266. doi: 10.1097/QAD.0b013e3282f736f4. [DOI] [PubMed] [Google Scholar]
- 26.Fox CH, Kotler D, Tierney A, Wilson CS, Fauci AS. Detection of HIV-1 RNA in the lamina propria of patients with AIDS and gastrointestinal disease. Journal of Infectious Diseases. 1989;159(3):467–471. doi: 10.1093/infdis/159.3.467. [DOI] [PubMed] [Google Scholar]
- 27.Asin SN, Fanger MW, Wildt-Perinic D, Ware PL, Wira CR, Howell AL. Transmission of HIV-1 by primary human uterine epithelial cells and stromal fibroblasts. Journal of Infectious Diseases. 2004;190(2):236–245. doi: 10.1086/421910. [DOI] [PubMed] [Google Scholar]
- 28.Hocini H, Becquart P, Bouhlal H, et al. Active and selective transcytosis of cell-free human immunodeficiency virus through a tight polarized monolayer of human endometrial cells. Journal of Virology. 2001;75(11):5370–5374. doi: 10.1128/JVI.75.11.5370-5374.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dezzutti CS, Guenthner PC, Cummins JE, et al. Cervical and prostate primary epithelial cells are not productively infected but sequester human immunodeficiency virus type 1. Journal of Infectious Diseases. 2001;183(8):1204–1213. doi: 10.1086/319676. [DOI] [PubMed] [Google Scholar]
- 30.Wu Z, Chen Z, Phillips DM. Human genital epithelial cells capture cell-free human immunodeficiency virus type 1 and transmit the virus to CD4+ cells: implications for mechanisms of sexual transmission. Journal of Infectious Diseases. 2003;188(10):1473–1482. doi: 10.1086/379248. [DOI] [PubMed] [Google Scholar]
- 31.Nazli A, Chan O, Dobson-Belaire WN, et al. Exposure to HIV-1 directly impairs mucosal epithelial barrier integrity allowing microbial translocation. PLoS Pathogens. 2010;6(4) doi: 10.1371/journal.ppat.1000852.e1000852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fontenot D, He H, Hanabuchi S, et al. TSLP production by epithelial cells exposed to immunodeficiency virus triggers DC-mediated mucosal infection of CD4+ T cells. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(39):16776–16781. doi: 10.1073/pnas.0907347106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Fontenot D. The role of mucosal epithelial cells in HIV infection [Dissertations and Theses] UT Graduate School of Biomedical Sciences at Houston; 2010. http://digitalcommons.library.tmc.edu/utgsbs_dissertations/28. [Google Scholar]
- 34.Yu H, Tudor D, Alfsen A, Labrosse B, Clavel F, Bomsel M. Peptide P5 (residues 628–683), comprising the entire membrane proximal region of HIV-1 gp41 and its calcium-binding site, is a potent inhibitor of HIV-1 infection. Retrovirology. 2008;5, article 93 doi: 10.1186/1742-4690-5-93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.McKinnon LR, Kaul R. Quality and quantity: mucosal CD4+ T cells and HIV susceptibility. Current Opinion in HIV and AIDS. 2012;7(2):195–202. doi: 10.1097/COH.0b013e3283504941. [DOI] [PubMed] [Google Scholar]
- 36.McKinnon LR, Nyanga B, Chege D, et al. Characterization of a human cervical CD4 + T cell subset coexpressing multiple markers of HIV susceptibility. Journal of Immunology. 2011;187(11):6032–6042. doi: 10.4049/jimmunol.1101836. [DOI] [PubMed] [Google Scholar]
- 37.Gosselin A, Monteiro P, Chomont N, et al. Peripheral blood CCR4+CCR6+ and CXCR3+CCR6+ CD4+ T cells are highly permissive to HIV-1 infection. Journal of Immunology. 2010;184(3):1604–1616. doi: 10.4049/jimmunol.0903058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Monteiro P, Gosselin A, Wacleche VS, et al. Memory CCR6+CD4+ T cells are preferential targets for productive HIV type 1 infection regardless of their expression of integrin β7. Journal of Immunology. 2011;186(8):4618–4630. doi: 10.4049/jimmunol.1004151. [DOI] [PubMed] [Google Scholar]
- 39.Kader M, Wang X, Piatak M, et al. α4+ β7hiCD4+ memory T cells harbor most Th-17 cells and are preferentially infected during acute SIV infection. Mucosal Immunology. 2009;2(5):439–449. doi: 10.1038/mi.2009.90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Brenchley JM, Paiardini M, Knox KS, et al. Differential Th17 CD4 T-cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood. 2008;112(7):2826–2835. doi: 10.1182/blood-2008-05-159301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Favre D, Lederer S, Kanwar B, et al. Critical loss of the balance between Th17 and T regulatory cell populations in pathogenic SIV infection. PLoS Pathogens. 2009;5(2) doi: 10.1371/journal.ppat.1000295.e1000295 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Arthos J, Cicala C, Martinelli E, et al. HIV-1 envelope protein binds to and signals through integrin α4β7, the gut mucosal homing receptor for peripheral T cells. Nature Immunology. 2008;9(3):301–309. doi: 10.1038/ni1566. [DOI] [PubMed] [Google Scholar]
- 43.Manel N, Unutmaz D, Littman DR. The differentiation of human TH-17 cells requires transforming growth factor-β and induction of the nuclear receptor RORγt. Nature Immunology. 2008;9(6):641–649. doi: 10.1038/ni.1610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Schutyser E, Struyf S, Van Damme J. The CC chemokine CCL20 and its receptor CCR6. Cytokine and Growth Factor Reviews. 2003;14(5):409–426. doi: 10.1016/s1359-6101(03)00049-2. [DOI] [PubMed] [Google Scholar]
- 45.Cremel M, Berlier W, Hamzeh H, et al. Characterization of CCL20 secretion by human epithelial vaginal cells: involvement in Langerhans cell precursor attraction. Journal of Leukocyte Biology. 2005;78(1):158–166. doi: 10.1189/jlb.0305147. [DOI] [PubMed] [Google Scholar]
- 46.Iliev ID, Mileti E, Matteoli G, Chieppa M, Rescigno M. Intestinal epithelial cells promote colitis-protective regulatory T-cell differentiation through dendritic cell conditioning. Mucosal Immunology. 2009;2(4):340–350. doi: 10.1038/mi.2009.13. [DOI] [PubMed] [Google Scholar]
- 47.Rescigno M, Di Sabatino A. Dendritic cells in intestinal homeostasis and disease. The Journal of Clinical Investigation. 2009;119(9):2441–2450. doi: 10.1172/JCI39134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Li Q, Agno JE, Edson MA, Nagaraja AK, Nagashima T, Matzuk MM. Transforming growth factor β receptor type 1 is essential for female reproductive tract integrity and function. PLoS Genetics. 2011;7(10) doi: 10.1371/journal.pgen.1002320.e1002320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sharkey DJ, Macpherson AM, Tremellen KP, Mottershead DG, Gilchrist RB, Robertson SA. TGF-β mediates proinflammatory seminal fluid signaling in human cervical epithelial cells. Journal of Immunology. 2012;189(2):1024–1035. doi: 10.4049/jimmunol.1200005. [DOI] [PubMed] [Google Scholar]
- 50.Irnidayanti Y. Effect of retinoic acid on fetus reproductive organ mice (Mus musculus) SwissWebster. International Journal of Medical and Biological Sciences. 2012;6:223–225. [Google Scholar]
- 51.Deng L, Shipley GL, Loose-Mitchell DS, et al. Coordinate regulation of the production and signaling of retinoic acid by estrogen in the human endometrium. Journal of Clinical Endocrinology and Metabolism. 2003;88(5):2157–2163. doi: 10.1210/jc.2002-021844. [DOI] [PubMed] [Google Scholar]
- 52.Takahashi T, Eitzman B, Bossert NL, et al. Transforming growth factors β1, β2, and β3 messenger RNA and protein expression in mouse uterus and vagina during estrogen-induced growth: a comparison to other estrogen-regulated genes. Cell Growth and Differentiation. 1994;5(9):919–935. [PubMed] [Google Scholar]
- 53.Lester RT, Yao XD, Ball TB, et al. Toll-like receptor expression and responsiveness are increased in viraemic HIV-1 infection. AIDS. 2008;22(6):685–694. doi: 10.1097/QAD.0b013e3282f4de35. [DOI] [PubMed] [Google Scholar]
- 54.Pandrea I, Sodora DL, Silvestri G, Apetrei C. Into the wild: simian immunodeficiency virus (SIV) infection in natural hosts. Trends in Immunology. 2008;29(9):419–428. doi: 10.1016/j.it.2008.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Brenchley JM, Price DA, Schacker TW, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nature Medicine. 2006;12(12):1365–1371. doi: 10.1038/nm1511. [DOI] [PubMed] [Google Scholar]
- 56.Mandl JN, Barry AP, Vanderford TH, et al. Divergent TLR7 and TLR9 signaling and type I interferon production distinguish pathogenic and nonpathogenic AIDS virus infections. Nature Medicine. 2008;14(10):1077–1087. doi: 10.1038/nm.1871. [DOI] [PubMed] [Google Scholar]
- 57.Vermund SH, Tabereaux PB, Kaslow R. Textbook of AIDS Medicine. 1999. [Google Scholar]
- 58.Tomescu C, Abdulhaqq S, Montaner LJ. Evidence for the innate immune response as a correlate of protection in human immunodeficiency virus (HIV)-1 highly exposed seronegative subjects (HESN) Clinical and Experimental Immunology. 2011;164(2):158–169. doi: 10.1111/j.1365-2249.2011.04379.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Boulet S, Kleyman M, Kim JY, et al. A combined genotype of KIR3DL1 high expressing alleles and HLA-B∗57 is associated with a reduced risk of HIV infection. AIDS. 2008;22(12):1487–1491. doi: 10.1097/QAD.0b013e3282ffde7e. [DOI] [PubMed] [Google Scholar]
- 60.Jennes W, Verheyden S, Demanet C, et al. Cutting edge: resistance to HIV-1 infection among African female sex workers is associated with inhibitory KIR in the absence of their HLA ligands. Journal of Immunology. 2006;177(10):6588–6592. doi: 10.4049/jimmunol.177.10.6588. [DOI] [PubMed] [Google Scholar]
- 61.Matte C, Lajoie J, Lacaille J, Zijenah LS, Ward BJ, Roger M. Functionally active HLA-G polymorphisms are associated with the risk of heterosexual HIV-1 infection in African women. AIDS. 2004;18(3):427–431. doi: 10.1097/00002030-200402200-00008. [DOI] [PubMed] [Google Scholar]
- 62.MacDonald KS, Fowke KR, Kimani J, et al. Influence of HLA supertypes on susceptibility and resistance to human immunodeficiency virus type 1 infection. Journal of Infectious Diseases. 2000;181(5):1581–1589. doi: 10.1086/315472. [DOI] [PubMed] [Google Scholar]
- 63.Lockett SF, Robertson JR, Brettle RP, Yap PL, Middleton D, Brown AJL. Mismatched human leukocyte antigen alleles protect against heterosexual HIV transmission. Journal of Acquired Immune Deficiency Syndromes. 2001;27(3):277–280. doi: 10.1097/00126334-200107010-00010. [DOI] [PubMed] [Google Scholar]
- 64.Liu C, Carrington M, Kaslow RA, et al. Association of polymorphisms in human leukocyte antigen class I and transporter associated with antigen processing genes with resistance to human immunodeficiency virus type 1 infection. Journal of Infectious Diseases. 2003;187(9):1404–1410. doi: 10.1086/374394. [DOI] [PubMed] [Google Scholar]
- 65.Dorak MT, Tang J, Penman-Aguilar A, et al. Transmission of HIV-1 and HLA-B allele-sharing within serodiscordant heterosexual Zambian couples. The Lancet. 2004;363(9427):2137–2139. doi: 10.1016/S0140-6736(04)16505-7. [DOI] [PubMed] [Google Scholar]
- 66.Ball TB, Ji H, Kimani J, et al. Polymorphisms in IRF-1 associated with resistance to HIV-1 infection in highly exposed uninfected Kenyan sex workers. AIDS. 2007;21(9):1091–1101. doi: 10.1097/QAD.0b013e3280ef6ae1. [DOI] [PubMed] [Google Scholar]
- 67.Bochud PY, Hersberger M, Taffé P, et al. Polymorphisms in Toll-like receptor 9 influence the clinical course of HIV-1 infection. AIDS. 2007;21(4):441–446. doi: 10.1097/QAD.0b013e328012b8ac. [DOI] [PubMed] [Google Scholar]
- 68.Dean M, Carrington M, Winkler C, et al. Genetic restriction of HIV infection and progression to AIDS by a deletion allele of the CKR5 structural gene. Hemophilia Growth and Development Study, Multicenter AIDS Cohort Study, Multicenter Hemophilia Cohort Study, San Francisco City Cohort, ALIVE Study. Science. 273:1856–1862. doi: 10.1126/science.273.5283.1856. [DOI] [PubMed] [Google Scholar]
- 69.Huang Y, Paxton WA, Wolinsky SM, et al. The role of a mutant CCR5 allele in HIV-1 transmission and disease progression. Nature Medicine. 1996;2(11):1240–1243. doi: 10.1038/nm1196-1240. [DOI] [PubMed] [Google Scholar]
- 70.Eugen-Olsen J, Iversen AKN, Garred P, et al. Heterozygosity for a deletion in the CKR-5 gene leads to prolonged AIDS-free survival and slower CD4 T-cell decline in a cohort of HIV-seropositive individuals. AIDS. 1997;11(3):305–310. doi: 10.1097/00002030-199703110-00007. [DOI] [PubMed] [Google Scholar]
- 71.De Roda Husman AM, Koot M, Cornelissen M, et al. Association between CCR5 genotype and the clinical course of HIV-1 infection. Annals of Internal Medicine. 1997;127(10):882–890. doi: 10.7326/0003-4819-127-10-199711150-00004. [DOI] [PubMed] [Google Scholar]
- 72.Meyer L, Magierowska M, Hubert JB, et al. Early protective effect of CCR-5 Δ32 heterozygosity on HIV-1 disease progression: relationship with viral load. AIDS. 1997;11(11):F73–F78. doi: 10.1097/00002030-199711000-00001. [DOI] [PubMed] [Google Scholar]
- 73.Zimmerman PA, Buckler-White A, Alkhatib G, et al. Inherited resistance to HIV-1 conferred by an inactivating mutation in CC chemokine receptor 5: studies in populations with contrasting clinical phenotypes, defined racial background, and quantified risk. Molecular Medicine. 1997;3(1):23–36. [PMC free article] [PubMed] [Google Scholar]
- 74.Michael NL, Chang G, Louie LG, et al. The role of viral phenotype and CCR-5 gene defects in HIV-1 transmission and disease progression. Nature Medicine. 1997;3(3):338–340. doi: 10.1038/nm0397-338. [DOI] [PubMed] [Google Scholar]
- 75.Misrahi M, Teglas JP, N’Go N, et al. CCR5 chemokine receptor variant in HIV-1 mother-to-child transmission and disease progression in children. JAMA. 1998;279(4):277–280. doi: 10.1001/jama.279.4.277. [DOI] [PubMed] [Google Scholar]
- 76.Bakshi SS, Zhang L, Ho D, Than S, Pahwa SG. Distribution of CCR5δ32 in human immunodeficiency virus-infected children and its relationship to disease course. Clinical and Diagnostic Laboratory Immunology. 1998;5(1):38–40. doi: 10.1128/cdli.5.1.38-40.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Smith MW, Dean M, Carrington M, et al. Contrasting genetic influence of CCR2 and CCR5 variants on HIV infection and disease progression. Hemophilia Growth and Development Study (HGDS), Multicenter AIDS Cohort Study (MACS), Multicenter Hemophilia Cohort Study (MHCS), San Francisco City Cohort (SFCC), ALIVE Study. Science. 1997;277:959–965. doi: 10.1126/science.277.5328.959. [DOI] [PubMed] [Google Scholar]
- 78.Kostrikis LG, Huang Y, Moore JP, et al. A chemokine receptor CCR2 allele delays HIV-1 disease progression and is associated with a CCR5 promoter mutation. Nature Medicine. 1998;4(3):350–353. doi: 10.1038/nm0398-350. [DOI] [PubMed] [Google Scholar]
- 79.Gonzalez E, Kulkarni H, Bolivar H, et al. The influence of CCL3L1 gene-containing segmental duplications on HIV-1/AIDS susceptibility. Science. 2005;307(5714):1434–1440. doi: 10.1126/science.1101160. [DOI] [PubMed] [Google Scholar]
- 80.Colobran R, Adreani P, Ashhab Y, et al. Multiple products derived from two CCL4 Loci: high incidence of a new polymorphism in HIV+ patients. Journal of Immunology. 2005;174(9):5655–5664. doi: 10.4049/jimmunol.174.9.5655. [DOI] [PubMed] [Google Scholar]
- 81.Malim MH, Bieniasz PD. HIV restriction factors and mechanisms of evasion. Cold Spring Harbour Perspectives in Medicine. 2012;2(5) doi: 10.1101/cshperspect.a006940.a006940 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Laguette N, Benkirane M. How Samhd1 changes our view of viral restriction. Trends in Immunology. 2012;33:26–33. doi: 10.1016/j.it.2011.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Mous K, Jennes W, Camara M, et al. Expression analysis of LEDGF/p75, APOBEC3G, TRIM5alpha, and tetherin in a Senegalese cohort of HIV-1-exposed seronegative individuals. PLoS One. 2012;7 doi: 10.1371/journal.pone.0033934.e33934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ghosh M, Shen Z, Fahey JV, Cu-Uvin S, Mayer K, Wira CR. Trappin-2/Elafin: a novel innate anti-human immunodeficiency virus-1 molecule of the human female reproductive tract. Immunology. 2010;129(2):207–219. doi: 10.1111/j.1365-2567.2009.03165.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ghosh M, Fahey JV, Shen Z, et al. Anti-HIV activity in cervical-vaginal secretions from HIV-positive and -negative women correlate with innate antimicrobial levels and IgG antibodies. PloS One. 2010;5(6) doi: 10.1371/journal.pone.0011366.e11366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Drannik AG, Nag K, Yao X-D, et al. Anti-HIV-1 activity of elafin is more potent than its precursor's, trappin-2, in genital epithelial cells. Journal of Virology. 2012;86(8):4599–4610. doi: 10.1128/JVI.06561-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Burgener A, Rahman S, Ahmad R, et al. Comprehensive proteomic study identifies serpin and cystatin antiproteases as novel correlates of HIV-1 resistance in the cervicovaginal mucosa of female Sex workers. Journal of Proteome Research. 2011;10(11):5139–5149. doi: 10.1021/pr200596r. [DOI] [PubMed] [Google Scholar]
- 88.Rowland-Jones SL, Dong T, Fowke KR, et al. Cytotoxic T cell responses to multiple conserved HIV epitopes in HIV- resistant prostitutes in Nairobi. The Journal of Clinical Investigation. 1998;102(9):1758–1765. doi: 10.1172/JCI4314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Kaul R, Plummer FA, Kimani J, et al. HIV-1-specific mucosal CD8+ lymphocyte responses in the cervix of HIV- 1-resistant prostitutes in Nairobi. Journal of Immunology. 2000;164(3):1602–1611. doi: 10.4049/jimmunol.164.3.1602. [DOI] [PubMed] [Google Scholar]
- 90.Kaul R, Trabattoni D, Bwayo JJ, et al. HIV-1-specific mucosal IgA in a cohort of HIV-1-resistant Kenyan sex workers. AIDS. 1999;13(1):23–29. doi: 10.1097/00002030-199901140-00004. [DOI] [PubMed] [Google Scholar]
- 91.Fowke KR, Kaul R, Rosenthal KL, et al. HIV-1-specific cellular immune responses among HIV-1-resistant sex workers. Immunology and Cell Biology. 2000;78(6):586–595. doi: 10.1046/j.1440-1711.2000.00944.x. [DOI] [PubMed] [Google Scholar]
- 92.Iqbal SM, Ball TB, Kimani J, et al. Elevated T cell counts and RANTES expression in the genital mucosa of HIV-1-resistant Kenyan commercial sex workers. Journal of Infectious Diseases. 2005;192(5):728–738. doi: 10.1086/432482. [DOI] [PubMed] [Google Scholar]
- 93.Alimonti JB, Koesters SA, Kimani J, et al. CD4+ T cell responses in HIV-exposed seronegative women are qualitatively distinct from those in HIV-infected women. Journal of Infectious Diseases. 2005;191(1):20–24. doi: 10.1086/425998. [DOI] [PubMed] [Google Scholar]
- 94.Devito C, Hinkula J, Kaul R, et al. Cross-clade HIV-1-specific neutralizing IgA in mucosal and systemic compartments of HIV-1-exposed, persistently seronegative subjects. Journal of Acquired Immune Deficiency Syndromes. 2002;30(4):413–420. doi: 10.1097/00042560-200208010-00007. [DOI] [PubMed] [Google Scholar]
- 95.Almeida JR, Price DA, Papagno L, et al. Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover. Journal of Experimental Medicine. 2007;204(10):2473–2485. doi: 10.1084/jem.20070784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Betts MR, Nason MC, West SM, et al. HIV nonprogressors preferentially maintain highly functional HIV-specific CD8+ T cells. Blood. 2006;107(12):4781–4789. doi: 10.1182/blood-2005-12-4818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Pereyra F, Addo MM, Kaufmann DE, et al. Genetic and immunologic heterogeneity among persons who control HIV infection in the absence of therapy. Journal of Infectious Diseases. 2008;197(4):563–571. doi: 10.1086/526786. [DOI] [PubMed] [Google Scholar]
- 98.Bélec L, Ghys PD, Hocini H, et al. Cervicovaginal secretory antibodies to human immunodeficiency virus type 1 (HIV-1) that block viral transcytosis through tight epithelial barriers in highly exposed HIV-1-seronegative African women. Journal of Infectious Diseases. 2001;184(11):1412–1422. doi: 10.1086/324375. [DOI] [PubMed] [Google Scholar]
- 99.Hocini H, Bomsel M. Infectious human immunodeficiency virus can rapidly penetrate a tight human epithelial barrier by transcytosis in a process impaired by mucosal immunoglobulins. Journal of Infectious Diseases. 1999;179(supplement 3):S448–S453. doi: 10.1086/314802. [DOI] [PubMed] [Google Scholar]
- 100.Lajoie J, Poudrier J, Massinga-Loembe M, et al. Differences in immunoregulatory cytokine expression patterns in the systemic and genital tract compartments of HIV-1-infected commercial sex workers in Benin. Mucosal Immunology. 2008;1(4):309–316. doi: 10.1038/mi.2008.18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Lajoie J, Poudrier J, Massinga Loembe M, et al. Chemokine expression patterns in the systemic and genital tract compartments are associated with HIV-1 infection in women from benin. Journal of Clinical Immunology. 2010;30(1):90–98. doi: 10.1007/s10875-009-9343-3. [DOI] [PubMed] [Google Scholar]
- 102.Fowke KR, Nagelkerke NJD, Kimani J, et al. Resistance to HIV-1 infection among persistently seronegative prostitutes in Nairobi, Kenya. The Lancet. 1996;348(9038):1347–1351. doi: 10.1016/S0140-6736(95)12269-2. [DOI] [PubMed] [Google Scholar]
- 103.Plummer FA, Ball TB, Kimani J, Fowke KR. Resistance to HIV-1 infection among highly exposed sex workers in Nairobi: what mediates protection and why does it develop? Immunology Letters. 1999;66(1–3):27–34. doi: 10.1016/s0165-2478(98)00182-5. [DOI] [PubMed] [Google Scholar]
- 104.Kaul R, Rowland-Jones SL, Kimani J, et al. Late seroconversion in HIV-resistant Nairobi prostitutes despite pre-existing HIV-specific CD8+ responses. The Journal of Clinical Investigation. 2001;107(3):341–349. doi: 10.1172/JCI10714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Card CM, McLaren PJ, Wachihi C, Kimani J, Plummer FA, Fowke KR. Decreased immune activation in resistance to HIV-1 infection is associated with an elevated frequency of CD4+CD25+FOXP3+ Regulatory T Cells. Journal of Infectious Diseases. 2009;199(9):1318–1322. doi: 10.1086/597801. [DOI] [PubMed] [Google Scholar]
- 106.Ahmed N, Hayashi T, Hasegawa A, et al. Suppression of human immunodeficiency virus type 1 replication in macrophages by commensal bacteria preferentially stimulating Toll-like receptor 4. Journal of General Virology. 2010;91(11):2804–2813. doi: 10.1099/vir.0.022442-0. [DOI] [PubMed] [Google Scholar]