

Abstract
Iridescence results from structures that generate color. Iridescence of bacterial colonies has recently been described and illustrated. The glitter-like iridescence class, created especially for a few strains of Cellulophaga lytica, exhibits an intense iridescence under direct illumination. Such color appearance effects were previously associated with other bacteria from the Bacteroidetes phylum, but without clear elucidation and illustration. To this end, we compared various bacterial strains to which the iridescent trait was attributed. All Cellulophaga species and additional Bacteroidetes strains from marine and terrestrial environments were investigated. A selection of bacteria, mostly marine in origin, were found to be iridescent. Although a common pattern of reflected wavelengths was recorded for the species investigated, optical spectroscopy and physical measurements revealed a range of different glitter-like iridescence intensity and color profiles. Importantly, gliding motility was found to be a common feature of all iridescent colonies. Dynamic analyses of “glitter” formation at the edges of C. lytica colonies showed that iridescence was correlated with layer superposition. Both gliding motility, and unknown cell-to-cell communication processes, may be required for the establishment, in time and space, of the necessary periodic structures responsible for the iridescent appearance of Bacteroidetes.
Introduction
Iridescence is a property of structural color arising from the interaction of light with micron- and sub-micron-sized periodic structures. The intense colors created exhibit angle-dependent color changes [1], [2], [3]. This form of color-appearance is found in natural or artificial organic/inorganic objects (e.g. opals, soap bubbles, compact discs) and in living organisms. Iridescence has been well studied in higher organisms, particularly Insecta [4], [5], [6], Aves [7], [8], [9], and fishes [2], [10]. Iridescence is also encountered in viruses [11], in marine organisms [12] and diatoms [13]. In animals, both communicative (e.g. species and sex recognition, predation avoidance) and non-communicative roles (e.g. photoprotection, thermoregulation, water repellency, friction reduction) were attributed to iridescence [14].
Iridescence expressed by bacterial colonies has been a vague concept in the literature. Only a few authors have studied the structural colors of prokaryotes [15], [16], [17], [18], [19], [20]. Ambiguous terms have been used for describing diverse visual effects of colonies or concentrated cell suspensions, but often with imprecise definitions. Images were rarely published and consequently the development of understanding of the visual colors was challenging. The comparison of a wide range of bacteria in [21] presented direct evidence of at least four categories of bacterial structural color. A novel and intense glitter-like iridescence class was created especially for a few strains of the marine bacterium Cellulophaga lytica. Glitter-like iridescence observed macroscopically under oblique direct illumination arose from bright millimeter- and sub-millimeter-sized centers of color distributed across the colony. The dominant glitter-like colors were observed to be green at the colonies' centers and were red and blue-violet at the colonies' peripheral edges. Importantly, and for the first time, the structural origin of the bacterial color appearance was verified by spectrophotometric data showing the reflected wavelengths under a broad range of light incidence angles. Several strains of C. lytica were compared and, interestingly, the intensity levels of iridescence were strongly different within the species.
The “iridescence” or “metallic tinge” of C. lytica colonies were mentioned only superficially in previous studies [22], [23]. Vague terms have been employed such as “greenish metallic iridescence” for the Cellulophaga genus [24] and “glistening”, “shine” or “shiny” for the Bacteriodetes [25]. These vague descriptions did not yield conclusions about the origin and nature of the observed colored appearances. Moreover, for some sample strains, relevant information relating to appearance was sometimes lost. For example, Flavobacterium [Cytophaga] johnsoniae and “Sporocytophaga cauliformis” were described as “greenish iridescent” [26] but without mention of the phenomenon in a more recent edition [24].
It was recently shown in C. lytica CECT 8139 that culture conditions can be the determining factor for the onset of iridescent appearance [27]. Thus, iridescent traits of many strains may have been overlooked due to non-optimal or inappropriate culture media conditions. In Cellulophaga spp. and other Flavobacteriales and Cytophagales, a close inspection of the optical properties may help to clarify the microbiology and ecology of iridescence.
In this study, we thoroughly examined the iridescence phenomena within all Cellulophaga species, and some closely related species from the Bacteroidetes phylum, on various culture media. Several C. lytica strains were included in order to evaluate the phenomenon within the species. Bacteroidetes strains described in previous studies as “green iridescent”, “iridescent”, “shiny” or “glistening” were selected to elucidate the color appearance effects. Illustrations of colonies are presented at the macroscopic and microscopic levels together with additional spectrophotometric data to describe the various intensities and iridescence profiles. Special attention was paid to the potential link between iridescence and gliding motility.
Materials and Methods
Bacterial strains
Strains are listed in Table 1 with recommended culture conditions and information on previous descriptions of their iridescence. “Reddish-greenish iridescence” and “metallic tinge” have been mentioned for Cellulophaga sp. [22], [26], so all strains belonging to this genus were included. Four C. lytica strains were incorporated in the study, including the genome-sequenced strain Lim 21T also registered as DSM 7489 and CIP 103822 [28]. The non-iridescent mutant of C. lytica CECT 8139 was obtained by spontaneous mutagenesis after cell-suspension dilution and following culture growth on MA.
Table 1. Bacteria from Bacteroidetes phylum with culture conditions and previous mentions of iridescence.
| Species | Strains | Origin | Country | Reference | Culture | Mention of iridescence | ||
| Medium | T°C | Description | Reference | |||||
| Bacteria with glitter-like iridescence | ||||||||
| Cellulophaga lytica | CECT 8139 | Surface of an anemone Actinia equine | Oléron island, France | [21] | MA | 20°C | Glitter-like | [21] |
| DSM 2040 | Seawater aquarium | La Jolla, USA | [30] | CYT | 20°C | Glitter-like | [21] | |
| DSM 74891(s) | Beach mud from limon | Costa Rica | Reddish-greenish iridescence | [26] # | ||||
| CIP 1038221(s) | Greenish metallic iridescence | [24] # | ||||||
| CIP 1038221(s) | Metallic tinge | [22] | ||||||
| Bacteria with suspicion of glitter-like iridescence | ||||||||
| Promyxobacterium flavum | DSM 3577 | Rhizosphere of tomato plant | Russia | [43] | NA | 25°C | Greenish iridescent | [26] # |
| Flavobacterium [Cytophaga] succinicans | DSM 4002 | Eroded fin of a fingerling chinook salmon | USA | [26] # | CY | 25°C | Greenish iridescent | [26] # |
| Sporocytophaga cauliformis | DSM 3657 | Water | Lake Bodensee, Germany | [26] # | CY | 20°C | Greenish iridescent | [26] # |
| Flavobacterium [Cytophaga] johnsoniae | DSM 2064(s) | ND | ND | [44] | CY | 30°C | Almost greenish iridescent | [26] # |
| Flavobacterium [Cytophaga] flevensis | DSM 1076 | Freshwater lake | The Netherlands | [32] | C/10 | 20°C | Iridescent | [26] # |
| Bacteria with suspicion of iridescence | ||||||||
| Cellulophaga baltica | CIP 106307 | Surface of brown alga Fucus serratus | Baltic Sea, Denmark | [22] | MA | 25°C | Metallic tinge | [22] |
| Cellulophaga fucicola | CIP 106308 | Surface of brown alga Fucus serratus | Kattegat sea, Denmark | [22] | MA | 25°C | Metallic tinge | [22] |
| Cellulophaga pacifica | LMG 21938 | Seawater | Gulf Peter, Japan | [45] | MA | 28°C | Shiny | [45] |
| Psychroflexus [Flavobacterium] gondwanensis | DSM 5423 | Organic lake | Antarctica | [46] | MA | 28°C | Shiny | [26] # |
| Flavobacterium aquatile | DSM 1132(s) | Deep well | Kent, England | [33] | Flavo | 30°C | Glistening | [26] # |
| Maribacter dokdonensis | DSM 17201 | Seawater | South Korea | [47] | MA | 28°C | Glistening | [48] |
| Bacteria with no description of iridescence | ||||||||
| Cellulophaga algicola | DSM 14237(s) | Diatom Melosira and macrophyta | Eastern Antarctic coast | [49] | MA | 10°C | - | - |
| Cellulophaga tyrosinoxydans | DSM 21164 | Seawater | Korea | [50] | MA | 28°C | - | - |
| Cellulophaga geojensis | CCUG 60801 | Marine sand | Geoje island, Korea | [51] | MA | 25°C | - | - |
| Leeuwenhoekiella [Cytophaga] marinoflava | DSM 3653 | Seawater | Scotland | [26] # | CYT | 20°C | - | - |
| Flavobacterium degerlachei | DSM 15718 | Microbial mat | Antarctica | [52] | R2A | 20°C | - | - |
The strain DSM 7489 was also registered as Lim-21 and CIP 103822 [28].
Genome sequenced strains.
These references correspond to editions of the “Bergey's Manual of Systematic Bacteriology” or “The Prokaryotes”.
ND: not detailed.
Culture conditions
Most of the culture media employed in this study comprised recommended media by strains catalogue DSMZ, CIP and BCCM/LMG (Table 1). For each strain, the culture media tested and the growth temperatures are listed in Table 2. Bacteria from the Bacteroidetes phylum have marine, freshwater or soil origins, thus both marine media (i) and terrestrial media (ii) were prepared.
Table 2. Iridescence phenomenon of strains belonging to the Bacteroidetes phylum.
| Strains | Media | T°C | ||||||||||
| MA | SYP | CYT | LN | sNA | ||||||||
| Growth 1 | Iridescence 2 | Growth | Iridescence | Growth | Iridescence | Growth | Iridescence | Growth | Iridescence | |||
| Marine strains | ||||||||||||
| Maribacter dokdonensis ** | DSM 17201 | ++ | + G | ++ | + G | ++ | +++ G | ++ | + G | + | − | 28°C |
| Cellulophaga lytica * | CECT 8139 | ++ | +++ G/R | ++ | ++ G/R | ++ | ++ G/R | ++ | ++ G | + | − | 25°C |
| DSM 2040 | ++ | +++ G/R | ++ | ++ G/R | ++ | +++ G/R | ++ | +++ G | + | − | 25°C | |
| DSM 7489(s) | ++ | + G/R | ++ | + G/R | ++ | + G/R | ++ | + G | + | − | 25°C | |
| DSM 2039 | ++ | − | ++ | − | ++ | ++ Y | ++ | ++ G | + | − | 25°C | |
| CIP 103822 | ++ | − | ++ | − | ++ | − | ++ | − | + | − | 25°C | |
| Cellulophaga baltica * | CIP 106307 | ++ | + R | ++ | + R | ++ | +++ R | ++ | + R | + | − | 25°C |
| Cellulophaga fucicola * | CIP 106308 | ++ | + G/R | ++ | + G/R | ++ | ++ G/R | ++ | + G | + | − | 25°C |
| Cellulophaga pacifica ** | LMG 21938 | ++ | + G/R | ++ | + G/R | ++ | ++ G/R | ++ | − | + | − | 25°C |
| Cellulophaga geojensis | CCUG 60801 | ++ | + G/R | ++ | + G/R | ++ | ++ G/R | ++ | ++ G | + | − | 25°C |
| Cellulophaga tyrosinoxydans | DSM 21164 | ++ | − | ++ | − | ++ | ++ G | ++ | + G | + | − | 25°C |
| Cellulophaga algicola | DSM 14237(s) | + | − | ++ | − | ++ | − | + | − | − | 25°C | |
| Psychroflexus gondwanesis ** | DSM 5423 | ++ | − | ++ | − | + | − | + | − | + | − | 25°C |
| Zobellia uliginosa | DSM 2061 | ++ | − | ++ | − | + | − | + | − | + | − | 25°C |
| Leeuwenhoekiella marinoflava | DSM 3653 | ++ | − | ++ | − | + | − | + | − | + | − | 20°C |
| Flavobacterium degerlachei | DSM 15718 | ++ | − | ++ | − | + | − | + | − | + | − | 20°C |
| Terrestrial strains | CY | Flavo | C/10 | TSA | NA | |||||||
| Flavobacterium johnsoniae * | DSM 2064(s) | ++ | ++ G/R | ++ | + G/R | ++ | ++ G/R | ++ | − | ++ | + G/R | 30°C |
| Flavobacterium aquatile ** | DSM 1132(s) | ++ | − | ++ | − | ++ | − | ++ | − | ++ | − | 30°C |
| Flavobacterium succinicans * | DSM 4002 | ++ | + R | ++ | − | ++ | − | ++ | − | ++ | − | 25°C |
| Promyxobacterium flavum * | DSM 3577 | ++ | − | ++ | − | ++ | − | ++ | − | ++ | − | 25°C |
| Flavobacterium flevensis * | DSM 1076 | ++ | − | ++ | − | ++ | − | ++ | − | ++ | − | 20°C |
| Sporocytophaga cauliformis * | DSM 3657 | ++ | + R | ++ | − | ++ | + R | ++ | − | ++ | + R | 20°C |
Strains previously described with “metallic tinge” or “greenish” appearances.
Strains previously described with “glistening” or “shiny” appearances.
Apparent growth: −, none; +, moderate; ++, good; +++, intense.
Iridescence and observed colors: −, none; +, low; ++, good; +++, bright. R, red; Y, Yellow; G, Green; G/R, middle green with red edges.
Genome sequenced strains.
(i) Marine media comprised ready-to-use Marine agar (MA) medium from Dutscher (Laboratorios Conda, S.A. Pronadisa®) and salted Nutrient agar (sNA) (Dutscher) with a supplement of NaCl 25 g L−1. Prepared media were made with artificial seawater (ASW) Instant Ocean© (30 g L−1). Cytophaga agar (CYT) contained 1 g tryptone, 0.5 g yeast extract, 0.5 g CaCl2.2 H2O, 0.5 g MgSO4.7 H2O, and 15 g agar in 1 L of ASW [22], [29], [30]. This medium was adapted with casein replaced by tryptone because C. lytica does not degrade casein. Low nutrient medium (LN) contained 15 g of agar in 1 L of ASW [31]. Recommended for Zobellia uliginosa, Seawater yeast peptone agar (SY) contained 5 g peptone, 3 g yeast extract, 12 g agar, 250 ml distilled water and 750 ml ASW.
(ii) Terrestrial media comprised ready-to-use Trypticase soy agar (TSA) and Nutrient agar (NA) (Dutsher). CY-agar (CY) was prepared by adding 3 g casitone, 1.36 g CaCl2.2H2O, 1 g yeast extract and 15 g of agar in 1 L of distilled water [26]. Recommended for Flavobacterium flevensis, C/10 contained 3 g casitone, 1.36 g CaCl2.2H2O and 15 g of agar in 1 L of distilled water [32]. Flavobacterium aquatile medium (Flav) was prepared by adding 2 g Na-caseinate, 0.5 g yeast extract, 1 g proteose peptone, 0.5 g K2HPO4 and 15 g of agar in 1 L of distilled water [33], [34].
Imaging colonies' iridescence appearance
The iridescence appearance of colonies was observed with the aid of a streaking procedure [21]. One colony from a 24 h-old plate was subcultured on a new plate drawing a 5 cm-linear streak in triplicate. Cultures were photographed after 24 h of growth in a dark room using an experimental arrangement of oblique epi-illumination at a fixed illumination angle of 60°. The light source was a lamp (Kaiser RB 218 N HF copy lighting unit) of 18 W, 5400 K, the operating voltage correspond to AC 220–240 V, 50 Hz with an operating frequency of 40 kHz. The camera was a Nikon D1500 18–55 VR on Av program with f 22 (12.1 Mega pixels). The lens was a macro, large-size used in superfin mode.
Gliding motility
Gliding motility was observed using timelapse microscopy. Pictures of colony edges were taken using an Olympus SZ61 stereomicroscope every 1 min for 20 min. The plates were observed under transverse illumination using a SCOTT KL 1500-halogen light source (150 W). For closest observations, colony edges were also observed in phase-contrast using an automated and inverted epi-fluorescence microscope TE2000-E-PFS (Nikon, France). The microscope was equipped with “The Perfect Focus System” that automatically maintains focus on a point of interest. Images were recorded with a CoolSNAP HQ 2 (Roper Scientific, Roper Scientific SARL, France) and a 40×/0.75 DLL “Plan-Apochromat” or a 100×/1.4 DLL objective. Due to the short working distances of 100× or 40× objectives, a 1 cm sided cube of colony growing on agar was cut and inverted on a coverslip.
Physical measurement of glitter-like iridescence (microspectrophotometry)
The diverse intensity and iridescence profiles of glitter-like iridescence were measured by microspectrophotometry [35]. Selected bacterial strains were included so that their various color-appearance effects could be evaluated. Samples were epi-illuminated and reflected light was collected from the same side of the sample. The angle of illumination and of detection could be separately set and controlled to a resolution of 0.5°. The illumination plane and the detection plane were offset from each other by 3° to enable unobstructed detection over the full collection angle range. Illumination was directed onto the sample through an Ocean Optics UV-Vis-NIR optical fiber that was connected to an Ocean Optics HPX-2000 light source, spanning approximately 300 nm to 850 nm. The reflected light was collected using a similar optical fiber that was connected to an Ocean Optics USB 4000-UV-VIS spectrometer. For a series of chosen fixed-incidence illumination angles, the collection fiber was stepped in 5° angle steps in an arc over the sample, and reflection spectra were recorded at each position. In this way, the dependence of reflected color with angle, and hence the extent of each sample's iridescence, could be measured and assessed.
Results
Iridescence within the Bacteroidetes phylum
In relation to previous descriptions of optical effects (Table 1), the glitter-like iridescence of several Bacteroidetes strains was investigated on several culture media (Table 2). The best media for iridescence observation were CYT and CY respectively for marine and terrestrial strains. The sNA and TSA-rich media did not appear to give rise to discernable iridescence. The brightness of the iridescence and the observed colors previously reported as having an “iridescent profile” were different within the species (Table 2, Figure 1). However, the glitter-like iridescence that was defined as being green with red edges was common to most iridescent Bacteroidetes. Red or yellow iridescence was also observed in a few species. A green iridescence profile was typical of most transparent colonies growing on LN medium. C. baltica exhibited a red iridescence after 24 h of growth but could acquire the green and red iridescence after 48 h/72 h (data not shown). Except for C. algicola and C. lytica CIP 103822, all Cellulophaga spp. were iridescent (Figure 1 A, B).
Figure 1. Iridescence colors of Bacteroidetes.

Iridescence was observed after 24 h of growth (or 48 h for Z. uliginosa and P. gondwanensis) on CYT medium (A, B, C), for marine strains, or CY medium (D) for terrestrial strains.
Three terrestrial strains were iridescent. F. johnsoniae had moderate iridescence intensity which was higher than the other strains, S. cauliformis and F. succinicans (Figure 1 D).
Spectrophotometric profiles of glitter-like iridescences
Optical reflection associated with different glitter-like iridescent profiles observed macroscopically was quantified using microspectrophotometry (Figure 2). The data shows optical reflection bands that unequivocally represent iridescence, and therefore structurally-generated color, of bacterial colonies on account of the change in their reflected wavelengths with angle. A common form of spectral map (presenting sample intensity vs wavelength vs detection angle) was typical to the iridescence profiles of all structurally colored samples. Main reflected wavelengths (700 nm, 530 nm, 380 nm, and 310 nm) are represented by the areas with higher intensity in coloring, visible for example on C. lytica DSM 2040 (Figure 2 A). A higher intensity of green-red (G/R) iridescence results in narrower band of reflected wavelengths (Figure 2 A).
Figure 2. Spectral properties of the different glitter-like iridescence profiles.

The color maps show the angle-dependent spectral reflectance of Cellulophaga spp. colonies grown 24 h on different media. Samples were illuminated at a fixed light angle of −70°. Scattered wavelengths from 300 nm to 850 nm were recorded at different detection angles from −65° to 70° with 5° angle step resolution. Results are shown using a color intensity scale. As shown in the small inserts (upper left in each color map), iridescence profiles correspond to: (A) green and red iridescence profile with high intensity from C. lytica DSM 2040, moderate intensity from C. lytica CECT 8139, and low intensity from C. lytica DSM 7489; (B) other color profiles with green profile from C. lytica DSM 2040 on LN, yellow profile from C. lytica DSM 2039 and red iridescence profile from C. baltica CIP 106307; (C) non-iridescent controls with C. lytica CIP 103822, C. lytica CECT 8139 on sNA and the CYT agar control. Wavelength values: UV, <400 nm; violet, 400 to 435 nm; blue, 435 to 490 nm; cyan, 490 to 520 nm; green, 520 to 560 nm; yellow, 560 to 590 nm; orange, 590 to 620 nm; red, 620 to 700 nm; and infrared, >700 nm.
The main scattering peaks were conserved for the different color profiles but with a displacement within the spectra (Figure 2 B). For the green profile of LN-grown colonies, the main peak was at lower wavelengths, i.e at 500 nm instead of 530 nm for the G/R profile (Figure 2 B). Higher wavelengths at 600 nm and 635 nm were obtained for yellow and red iridescent profiles, respectively. A fifth peak at 380 nm was discerned for the red iridescent C. baltica (Figure 2 B). Under non-iridescent conditions with C. lytica CIP 103822 on CYT medium and C. lytica CECT 8139 on sNA, no reflection peaks were observed. The background of reflected colors was due to pigmentation and the agar medium (Figure 2 C).
Correlation between gliding motility and iridescence
Gliding bacteria move individually or in swarms, resulting in the formation of typical colonies that have thin spreading edges [36]. Gliding motility might be required for the establishment of periodic structures within the iridescent colonies. In order to test this hypothesis, we attempted to isolate a mutant of C. lytica CECT 8139 impaired in gliding motility. A spontaneous gliding-negative mutant was obtained. Interestingly, this mutant is almost non-iridescent, harboring only a residual iridescence when the colony edges were observed microscopically (Figure 3 A). A strong link between gliding motility and wild-type iridescence may thus exist. To reinforce this hypothesis, a rigorous microscopic inspection of colony edges was performed in the Bacteroidetes strains (Figure 3 B, C). Since iridescence and colony edges were not in the same focal plane, the optical effects were hard to observe in this test. All iridescent colonies exhibited spreading edges, a hallmark of gliding bacteria. Interestingly, these typical edges were absent in colonies that appeared non-iridescent at the macroscopic level (e.g. C. algicola and C. lytica CIP 103822). However, typical edges of gliding bacteria were found in some non-iridescent strains, in particular for terrestrial bacteria. Moreover, no correlation was found between the intensity of iridescence and the gliding rates for the tested strains (Table 3).
Figure 3. Edges patterns of Bacteroidetes colonies.

Marine bacteria (A, B) and terrestrial bacteria (C) were grown 24 h on CYT and CY media, respectively. In (A), pictures of the entire colonies of C. lytica CECT 8139 and its mutant (impaired in gliding motility) are shown for comparison at the macroscopic level. The mutant was not iridescent on SYP, CYT, LN and sNA culture media (data not shown). All iridescent colonies exhibited typical spreading and jagged edges. Iridescent strains were: C. lytica CECT 8139 and DSM 2040 (and to a lesser extent DSM 7489), C. baltica CIP 106307, C. fucicola CIP 106308, C. pacifica LMG 21938, C. geojensis CCUG 60801, C. tyrosinoxydans DSM 21164, M. dokdonensis DSM 1720, F. johnsoniae DSM 2064, F. succinicans DSM 4002 and S. cauliformis DSM 3657.
Table 3. Gliding motility and iridescence on CYT or CY media.
| Strains | Iridescence1 | Gliding motility | ||
| Marine strains (CYT medium) | Typical edges2 | Rate3 | ||
| Cellulophaga lytica | CECT 8139 | ++ | ++ | Fast |
| CECT 8139 mutant | ∼ | − | ||
| DSM 2040 | +++ | ++ | Fast | |
| DSM 7489(s) | + | + | Moderate | |
| DSM 2039 | ++ | + | Slow | |
| CIP 103822 | − | − | ||
| Cellulophaga baltica | CIP 106307 | ++ | ++ | Slow |
| Cellulophaga fucicola | CIP 106308 | ++ | ++ | Moderate |
| Cellulophaga pacifica | LMG 21938 | ++ | ++ | Moderate |
| Cellulophaga geojensis | CCUG 60801 | ++ | ++ | Slow |
| Cellulophaga tyrosinoxydans | DSM 21164 | ++ | + | Slow |
| Cellulophaga algicola | DSM 14237(s) | − | − | |
| Psychroflexus gondwanensis | DSM 5423 | − | − | |
| Zobellia uliginosa | DSM 2061 | − | + | Moderate |
| Leeuwenhoekiella marinoflava | DSM 3653 | − | − | |
| Flavobacterium degerlachei | DSM 15718 | − | − | |
| Maribacter dokdonensis | DSM 17201 | ++ | ++ | Slow |
| Terrestrial strains (CY medium) | ||||
| Flavobacterium johnsoniae | DSM 2064(s) | ++ | ++ | Moderate |
| Flavobacterium aquatile | DSM 1132(s) | − | − | |
| Flavobacterium succinicans | DSM 4002 | + | ++ | Slow |
| Promyxobacterium flavum | DSM 3577 | − | ++ | Moderate |
| Flavobacterium flevensis | DSM 1076 | − | ++ | Slow |
| Sporocytophaga cauliformis | DSM 3657 | + | ++ | Fast |
Iridescence intensity: −, none; ∼, microscopically observed; +, low; ++, good; +++, intense.
Typical gliding edges: −, none; +, moderate; ++, wide.
Gliding motility rates were qualified visually: Fast, spreading movements on the agar with visible distance covered after only 20 min; Moderate, spreading movements at a lower extend; Slow, few movements within the colony with short distance covered.
Genome sequenced strains.
Colony edges of C. lytica strains were also observed at higher magnification (Figure 4). C. lytica CECT 8139 moved as single cells and within coordinated groups. The mutant of C. lytica CECT 8139 displaying only a reduced iridescence at the colony edges still formed tiers but lacked gliding cells. Moreover, when C. lytica CIP 103822 and C. lytica CECT 8139 were plated in non iridescent conditions (CYT and sNA respectively), gliding cells were not observed. Conversely, iridescent C. lytica CECT 8139 exhibited fast gliding motility (see Movies S1 and S2 as supplemental data). Taken together these results suggest a correlation between iridescence and gliding motility.
Figure 4. Phase contrast observation of colony edges of different C. lytica strains at high magnifications.
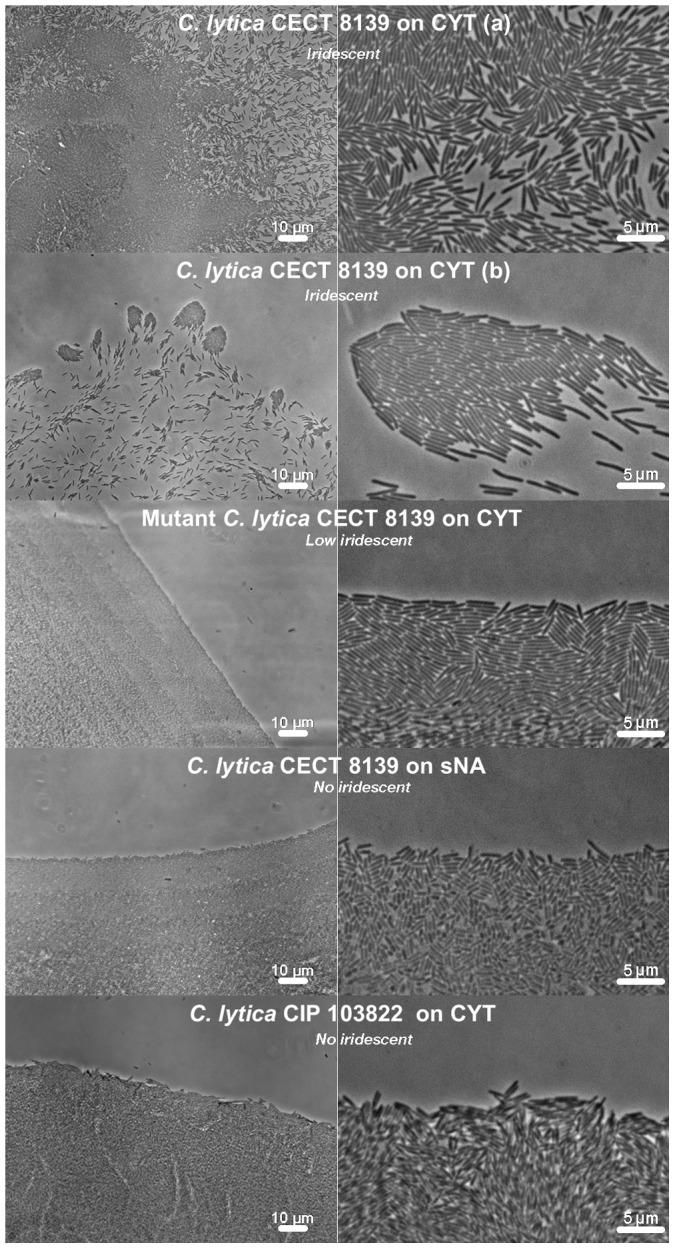
Iridescent C. lytica CECT 8139 growing on CYT was observed at different colony positions: an edge (a) and an extreme edge (b). Low or non-iridescent conditions were CECT 8139 gliding defective mutant on CYT, C. lytica CECT 8139 grown on sNA and CIP 103822 strains grown on CYT. Magnification was ×400 (left column) and ×1000 (right column).
Dynamic formation of an iridescent glitter
We designed an experimental device to monitor the appearance of iridescence over time in a gliding colony. Intense iridescent colonies of C. lytica DSM 2040 were used to observe the kinetic formation of glitters (Figure 5). Colonies appeared to move rapidly and the formation of a glitter centre could be followed (black arrows). After only 20 minutes the first non-iridescent layer was visualized and 10 minutes later, likely due to layer superposition, a glitter effect could be observed. The iridescence was conserved even with rapid micro-colony movement, suggesting the existence of a complex ordered structure continuously being formed, for example, if the cells follow the exact same path in the glitter area. These effects were more easily observed on the video sequence (see Movie S3).
Figure 5. Dynamic formation of iridescence.

A glitter centre (black arrow) was observed on the edges of a C. lytica DSM 2040 colony after 24 h incubation on CYT medium. Colony was photographed every 5 min for 60 min. Scales bars (A) 0.5 mm and (B) 0.1 mm.
Discussion
Iridescence in prokaryotes is a concept that has only recently been elucidated [21]. The intense glitter-like iridescence observed under direct illumination was typical of some C. lytica colonies. Understanding the bases of such iridescence and its cellular function requires its identification in bacteria and its correlation with the environment and specificities of the iridescent strains. Therefore, we set out to document iridescence in the Bacteroidetes phylum. We found that the previous descriptive terms such as “shiny”, “glistening”, “metallic tinge” or “greenish iridescent” were clearly correlated with iridescence effects for marine bacteria, but this correlation was not clear for terrestrial strains. Marine bacteria and in particular Cellulophaga spp. were found to exhibit the most intense iridescence effects. However, the iridescent trait was scattered within the Bacteroidetes and no phylogenetic link was found between the iridescent species.
As recently shown by [27], culture conditions were confirmed to influence strongly the observation of the iridescence. Nutrient-rich media were found to inhibit both gliding motility and iridescence. Specific physiological states might thus be required for the production of metabolites involved in the onset of iridescence. The repression of iridescence under certain conditions may explain, in part, the lack of previously reported descriptions and illustrations.
As several strains did not produce the previously reported effects it was essential to illustrate colonies' iridescence properties precisely. This was achieved using a specific epi-illumination experimental arrangement. Descriptive terms must be chosen carefully and the observation conditions must be precisely defined, in particular because a diverse set of iridescence profiles have been highlighted in this study. The green and red iridescence defined as “glitter-like” iridescence was observed in several marine Bacteroidetes and F. johnsoniae but with different intensity levels. Evidence of distinct color variations was also collected. For these reasons, the common glitter-like iridescence class could be divided in different sub-classes referring to the color.
In spite of these differences in intensities and/or reflected colors, strong similarities were found between the intensity-color maps of their iridescence. Similar periodic structures, with minor variations of dimensions and periodicity, are therefore inferred to be responsible for the iridescent appearances observed.
It is still unknown whether bacterial iridescence occurs in natural habitats. However, iridescence was conserved under conditions that mimic the marine habitat [27]. Moreover, Bacteroidetes are widely distributed in nature, and Cellulophaga spp. are found in rocky shore ecosystems where stressful conditions of salinity, temperature variation, desiccation and light exposure select adapted organisms. The iridescence of bacterial colonies or biofilms might therefore have eco-biological roles by strongly reflecting specific wavelengths. UV and IR wavelengths were particularly well reflected by colonies exhibiting glitter-like iridescence. These typical reflections may have photoprotective or thermoregulatory roles. Non-communicative functions of iridescence have been reported in higher organisms [2], [6], [14]. For example, the reed frog Hyperolius viridiflacus adapts its skin color appearance and reflectance with the arid environmental conditions of African savannas [37]. The measured spectral maxima in reflectance are in the IR wavelengths from 800 nm to 1100 nm with additional reflectance of UV. Periodic structures are also responsible for reducing mechanical friction in some burrowing animals [38]. A similar mechanical role might also be envisaged for bacterial iridescence.
A strong correlation was found between gliding motility and iridescence in this study. Gliding motility appeared as a necessary -but not sufficient- condition for the formation of iridescent colonies. In gliding motility, the cell movements proceed without pili or flagella, but correspond to “a smooth translocation of cells over a surface by an active process that requires the expenditure of energy” [36]. The cellular mechanisms are not perfectly known in the Bacteroidetes [39]. At the colony level, one of the characteristics is spreading edges. In this study, all iridescent colonies possessed typical edges of gliding bacteria, and the motility was particularly rapid for some strains. Moreover, dynamic analyses of glitter-centre formation indicated that the iridescent structures themselves could move. A fast gliding motility was thus compatible with the maintenance of a certain periodicity of disposition within the cell population. Such process likely requires fine-tuning and a high degree of cell-to-cell communication. This is a major question that remains unanswered. Among the strains tested, only three have their genome completely sequenced, namely F. johnsoniae DSM2064 (UW101), C. algicola DSM 14237 and C. lytica DSM 7489 [28], [40]. F. johnsoniae might constitute a good candidate to characterize the genetic bases of iridescence. Accordingly, F. johnsoniae has already been used as a model organism for studies of Cytophaga-Flavobacterium gliding motility, and a wide variety of genetic tools have been developed for its manipulation [41], [42]. Moreover, genome comparisons between iridescent and non-iridescent Bacteroidetes strains might help us to identify key genes controlling both gliding motility and dynamic formation of iridescent glitter centers. This approach is currently being used in our laboratory for several strains of C. lytica.
Supporting Information
Gliding motility of C. lytica CECT 8139 observed at colony edges by phase contrast microscopy. The bacterium was grown on Cytophaga agar medium and movements were recorded during 1 minute (one photo every three seconds). Gliding reverse movements were visible. Motility was reduced in the central parts of the colony.
(MOV)
Gliding motility of C. lytica CECT 8139 observed at colony extreme edges by phase contrast microscopy. The bacterium was grown on Cytophaga agar medium and movements were recorded during 1 minute (one photo every three seconds). A collective motion and the organization of cells into larger moving clusters were observed.
(MOV)
Dynamic formation of an iridescent glitter on the edges of a C. lytica DSM 2040 colony after 24 h incubation on Cytophaga agar medium. The phenomenon was observed with a total duration of 50 minutes (one photo every minute). On the edges, spreading movements were well visible. Several layers seemed needed to observe the red iridescence from the glitter.
(MOV)
Acknowledgments
ER acknowledges Jean-Jacques Levrel from the C.H.U. Pontchaillou (Rennes, France) for photography advices.
Funding Statement
PV acknowledges the support of AFOSR grant FA9550-10-1-0020. BK was a PhD student with a grant from the Ministère de la recherche et de l'enseignement supérieur. ER acknowledges the support of CNRS grant AIR75515 (“Bactéridescence” project). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Fox DL (1976) Animal biochromes and structural colors: physical, chemical, distributional & physiological features of colored bodies in the animal world, 2nd ed. University of California Press: Berkeley. 433 p.
- 2. Land M (1972) The physics and biology of animal reflectors. Prog Biophys Mol Biol 24: 75–106. [DOI] [PubMed] [Google Scholar]
- 3. Vukusic P, Sambles JR, Lawrence CR, Wootton RJ (2001) Structural color: now you see it-now you don't. Nature 410: 36. [DOI] [PubMed] [Google Scholar]
- 4. Vukusic P, Sambles JR (2003) Photonic structures in biology. Nature 424: 6950, 852–855. [DOI] [PubMed] [Google Scholar]
- 5. Ghiradella H (1991) Light and color on the wing: structural colors in butterflies and moths. Appl Optics 30: 3492–3500. [DOI] [PubMed] [Google Scholar]
- 6. Seago AE, Brady P, Vigneron J-P, Schultz TD (2009) Gold bugs and beyond: a review of iridescence and structural color mechanisms in beetles (Coleoptera). J R Soc Interface 6: S165–S184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Greenewalt CH, Brandt W, Friel DD (1960) The iridescent colors of hummingbird feathers. Proc Am Philos Soc 104: 249–253. [Google Scholar]
- 8. Prum RO, Torres R (2003) Structural coloration of avian skin: convergent evolution of coherently scattering dermal collagen arrays. J Exp Biol 206: 2409–2429. [DOI] [PubMed] [Google Scholar]
- 9. Doucet SM, Shawkey MD, Hill GE, Montgomerie R (2006) Iridescent plumage in satin bowerbirds: structure, mechanisms and nanostructural predictors of individual variation in color. J Exp Biol 209: 380–390. [DOI] [PubMed] [Google Scholar]
- 10. Lythgoe JN, Shand J (1989) The structural basis for iridescent color changes in dermal and corneal iridophores in fish. J Exp Biol 141: 313–325. [Google Scholar]
- 11. Williams RC, Smith KM (1958) The polyhedral form of the Tipula iridescent virus. Biochim Biophys Acta 28: 464–469. [DOI] [PubMed] [Google Scholar]
- 12. Welch V, Vigneron J-P, Lousse V, Parker A (2006) Optical properties of the iridescent organ of the comb-jellyfish Beroë cucumis (Ctenophora). Phys Rev E 73: 0419161–0419167. [DOI] [PubMed] [Google Scholar]
- 13. Noyes J, Sumper M, Vukusic P (2008) Light manipulation in a marine diatom. J Mater Res 23: 3229–3232. [Google Scholar]
- 14. Doucet SM, Meadows MG (2009) Iridescence: a functional perspective. J R Soc Interface 6: S115–S132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Engboek HC (1950) The phenomenon of iridescence in bacterial cultures, with particular reference to Pfeiffer's bacillus. Acta Pathol Microbiol 27: 388–393. [Google Scholar]
- 16. Guillot M (1941) Relation entre certaines propriétés optiques des bactéries et leur forme géométrique vraie. Diffraction de la lumière par les cultures et symétrie de la bactérie. C R Hebd Seances Acad Sci 1100–1103. [Google Scholar]
- 17. Nogrady G, Guérault A (1964) Conditions favorisant l'iridescence bactérienne et l'observation du phénomène. Rev Can Biol 23: 367–373. [PubMed] [Google Scholar]
- 18. Pijper A (1918) Diffraction phenomena in cultures of microöganisms. Med J S Afr 14: 211–218. [Google Scholar]
- 19. Pijper A (1923) Diffraction in biological structures. I. The structure of colonies of the coli-typhoid group. S Afr Med Rec 17: 243–248. [Google Scholar]
- 20. Ponder E (1934) Diffraction patterns produced by bacteria. J Exp Biol 11: 54–57. [Google Scholar]
- 21. Kientz B, Vukusic P, Luke S, Rosenfeld E (2012) Iridescence of a marine bacterium and classification of prokaryotic structural colors. Appl Environ Microbiol 78: 2092–2099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Johansen JE, Nielsen P, Sjøholm C (1999) Description of Cellulophaga baltica gen. nov., sp. nov. and Cellulophaga fucicola gen. nov., sp. nov. and reclassification of [Cytophaga] lytica to Cellulophaga lytica gen. nov., comb. nov. Int J Syst Evol Micr 49: 1231–1240. [DOI] [PubMed] [Google Scholar]
- 23. Skerratt JH, Bowman JP, Hallegraeff G, James S, Nichols PD (2002) Algicidal bacteria associated with blooms of a toxic dinoflagellate in a temperate Australian estuary. Mar Ecol Prog Ser 244: 1–15. [Google Scholar]
- 24.Bowman JP (2011) Genus IX. Cellulophaga Johansen, Nielsen and Sjøholm 1999, 1238VP In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, Ludwig W, Whitman WB, editors. Bergey's manual of systematic bacteriology, The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes, 2nd ed., vol 4. Springer Press: New York Dordrecht Heidelberg London, pp. 176–180.
- 25.Bernardet J-F, Nakagawa Y (2006) An introduction to the family Flavobacteriaceae. In: Dworkin M, Falkow S, Rosenberg E, Schleifer K-H, Stackebrandt E, editors. The Prokaryotes, 3rd ed., vol. 7. Springer, New York Press: NY, pp. 455–480.
- 26.Reichenbach H (1989) Genus I. Cytophaga Winogradsky 1929, 577AL emend. In: Staley JT, Bryant MP, Pfennig N, Holt JG, editors. Bergey's manual of systematic Bacteriology, vol. 3. Williams & Wilkins Press: Baltimore, pp. 2015–2050.
- 27. Kientz B, Marié P, Rosenfeld E (2012) Effect of abiotic factors on the unique glitter-like iridescence of Cellulophaga lytica . FEMS Microbiol Lett 333 2: 101–8. [DOI] [PubMed] [Google Scholar]
- 28. Pati A, Abt B, Teshima H, Nolan M, Lapidus A, et al. (2011) Complete genome sequence of Cellulophaga lytica type strain (LIM-21T). Stand Genomic Sci 4: 221–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Lewin RA (1969) A Classification of Flexibacteria. Microbiology 58: 189–206. [DOI] [PubMed] [Google Scholar]
- 30. Lewin RA, Lounsbery DM (1969) Isolation, cultivation and characterization of Flexibacteria. J Gen Microbiol 58: 145–170. [DOI] [PubMed] [Google Scholar]
- 31. Jensen PR, Kauffman CA, Fenical W (1996) High recovery of culturable bacteria from the surfaces of marine algae. Mar Biol 126: 1–7. [Google Scholar]
- 32. van der Meulen HJ, Harder W, Veldkamp H (1974) Isolation and characterization of Cytophaga flevensis sp. n., a new agarolytic flexibacterium. Antonie van Leeuwenhoek J Microbiol Serol 40: 329–346. [DOI] [PubMed] [Google Scholar]
- 33. Weeks OB (1955) Flavobacterium aquatile (Frankland and Frankland) Bergey et al., type species of the genus Flavobacterium . J Bacteriol 69: 649–658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Reichenbach H, Dworkin M (1981) The Order Cytophagales (with addenda on the genera Herpetosiphon, Saprospira, and Flexithrix). In: Starr MP, Stolp H, Trüper HG, Balows A, Schlegel HG, editors. The Prokaryotes: a handbook on habitats, isolation, and identification of bacteria, 1st ed., vol. 1. Springer-Verlag Press: Berlin, pp. 356–379.
- 35. Vukusic P, Stavenga DG (2009) Physical methods for investigating structural colours in biological systems. J R Soc Interface 6: S133–S148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. McBride MJ (2001) Bacterial gliding motility: multiple mechanisms for cell movement over surfaces. Annu Rev Microbiol 55: 49–75. [DOI] [PubMed] [Google Scholar]
- 37. Kobelt F, Linsenmair KE (1992) Adaptations of the reed frog Hyperolius viridiflavus (Amphibia: Anura: Hyperoliidae) to its arid environment. VI. The iridophores in the skin as radiation reflectors. J Comp Physiol B 162: 314–26. [DOI] [PubMed] [Google Scholar]
- 38. Meadows MG, Butler MW, Morehouse NI, Taylor LA, Toomey MB, et al. (2009) Iridescence: views from many angles. J R Soc Interface Suppl. 2: S107–S113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. McBride MJ (2004) Cytophaga-Flavobacterium gliding motility. J Mol Microbiol Biotechnol 7: 63–71. [DOI] [PubMed] [Google Scholar]
- 40. Abt B, Lu M, Misra M, Han C, Nolan M, et al. (2011) Complete genome sequence of Cellulophaga algicola type strain (IC166). Stand Genomic Sci 4: 72–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Braun TF, Khubbar MK, Saffarini DA, McBride MJ (2005) Flavobacterium johnsoniae gliding motility genes identified by mariner mutagenesis. J Bacteriol 18: 6943–6952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Rhodes RG, Pucker HG, McBride MJ (2011) Development and use of a gene deletion strategy for Flavobacterium johnsoniae to identify the redundant gliding motility genes remF, remG, remH, and remI. J Bacteriol 193: 2418–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Vozniakovskaya YM, Rybakova ZP (1969) Some new data on ecology and properties of bacteria belonging to Promyxobacterium genus. Mikrobiologiya 38: 135–143. [PubMed] [Google Scholar]
- 44. Stanier RY (1947) Studies on Nonfruiting Myxobacteria: I. Cytophaga johnsonae, n.sp., a Chitin-decomposing Myxobacterium. J Bacteriol 53: 297–315. [PMC free article] [PubMed] [Google Scholar]
- 45. Nedashkovskaya OI (2004) Cellulophaga pacifica sp. nov. Int J Syst Evol Microbiol 54: 609–613. [DOI] [PubMed] [Google Scholar]
- 46. Dobson SJ, James SR, Franzmann PD, McMeekin TA (1991) A numerical taxonomic study of some pigmented bacteria isolated from Organic Lake, an Antarctic hypersaline lake. Arch Microbiol 156: 56–61. [Google Scholar]
- 47. Yoon JH, Kang SJ, Lee SY, Lee CH, Oh TK (2005) Maribacter dokdonensis sp. nov., isolated from sea water off a Korean island, Dokdo. Int J Syst Evol Microbiol 55: 2051–2055. [DOI] [PubMed] [Google Scholar]
- 48.Bernardet JF, Bowman JP (2011) Genus I. Flavobacterium Bergey, Harrison, Breed, Hammer and Huntoon 1923, 97AL emend. Bernardet, Sergers, Vancanneyt, Berthe, Kersters and Vandamme 1996, 139, editors. Bergey's manual of systematic bacteriology, The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes, 2nd ed., vol. 4. Springer Press: New York Dordrecht Heidelberg London. pp. 112–176.
- 49. Bowman JP (2000) Description of Cellulophaga algicola sp. nov., isolated from the surfaces of Antarctic algae, and reclassification of Cytophaga uliginosa (ZoBell and Upham 1944) Reichenbach 1989 as Cellulophaga uliginosa comb. nov. Int J Syst Evol Microbiol 50: 1861–1868. [DOI] [PubMed] [Google Scholar]
- 50. Kahng HY, Chung BS, Lee DH, Jung JS, Park JH, Jeon CO (2009) Cellulophaga tyrosinoxydans sp. nov., a tyrosinase-producing bacterium isolated from seawater. Int J Syst Evol Microbiol 59: 654–657. [DOI] [PubMed] [Google Scholar]
- 51. Park S, Oh KH, Lee SY, Oh TK, Yoon JH (2012) Cellulophaga geojensis sp. nov., a member of the family Flavobacteriaceae isolated from marine sand. Int J Syst Evol Microbiol 62: 1354–1358. [DOI] [PubMed] [Google Scholar]
- 52. Van Trappen S, Vandecandelaere I, Mergaert J, Swings J (2004) Flavobacterium degerlachei sp. nov., Flavobacterium frigoris sp. nov. and Flavobacterium micromati sp. nov., novel psychrophilic bacteria isolated from microbial mats in Antarctic lakes. Int J Syst Evol Microbiol 54: 85–92. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Gliding motility of C. lytica CECT 8139 observed at colony edges by phase contrast microscopy. The bacterium was grown on Cytophaga agar medium and movements were recorded during 1 minute (one photo every three seconds). Gliding reverse movements were visible. Motility was reduced in the central parts of the colony.
(MOV)
Gliding motility of C. lytica CECT 8139 observed at colony extreme edges by phase contrast microscopy. The bacterium was grown on Cytophaga agar medium and movements were recorded during 1 minute (one photo every three seconds). A collective motion and the organization of cells into larger moving clusters were observed.
(MOV)
Dynamic formation of an iridescent glitter on the edges of a C. lytica DSM 2040 colony after 24 h incubation on Cytophaga agar medium. The phenomenon was observed with a total duration of 50 minutes (one photo every minute). On the edges, spreading movements were well visible. Several layers seemed needed to observe the red iridescence from the glitter.
(MOV)