

Abstract
Burmese is an old and popular cat breed, however, several health concerns, such as hypokalemia and a craniofacial defect, are prevalent, endangering the general health of the breed. Hypokalemia, a subnormal serum potassium ion concentration ([K+]), most often occurs as a secondary problem but can occur as a primary problem, such as hypokalaemic periodic paralysis in humans, and as feline hypokalaemic periodic polymyopathy primarily in Burmese. The most characteristic clinical sign of hypokalemia in Burmese is a skeletal muscle weakness that is frequently episodic in nature, either generalized, or sometimes localized to the cervical and thoracic limb girdle muscles. Burmese hypokalemia is suspected to be a single locus autosomal recessive trait. A genome wide case-control study using the illumina Infinium Feline 63K iSelect DNA array was performed using 35 cases and 25 controls from the Burmese breed that identified a locus on chromosome E1 associated with hypokalemia. Within approximately 1.2 Mb of the highest associated SNP, two candidate genes were identified, KCNH4 and WNK4. Direct sequencing of the genes revealed a nonsense mutation, producing a premature stop codon within WNK4 (c.2899C>T), leading to a truncated protein that lacks the C-terminal coiled-coil domain and the highly conserved Akt1/SGK phosphorylation site. All cases were homozygous for the mutation. Although the exact mechanism causing hypokalemia has not been determined, extrapolation from the homologous human and mouse genes suggests the mechanism may involve a potassium-losing nephropathy. A genetic test to screen for the genetic defect within the active breeding population has been developed, which should lead to eradication of the mutation and improved general health within the breed. Moreover, the identified mutation may help clarify the role of the protein in K+ regulation and the cat represents the first animal model for WNK4-associated hypokalemia.
Introduction
Potassium is the most abundant cation in mammals [1], [2]. The resting membrane potential of cells is affected by the relationship between intracellular and extracellular potassium concentrations and the resting potassium conductance [3]. Since the extracellular potassium greatly affects the tendency of cells to fire action potentials, potassium plays a crucial role in the function of nervous tissue and muscle (skeletal, cardiac and smooth) throughout the body [1], [2], implying perturbations can be debilitating or even life-threatening. To maintain ideal body homeostasis, potassium excretion and dietary intake must be balanced [4]–[6]. Abnormalities of potassium homeostasis can occur as a primary condition, or as a secondary disorder [7]–[10]. Inherited hypokalemia has been discovered in a variety of mammals by genetic studies of individuals and families affected by clinical disease. A classic syndrome of myopathic weakness, hyperkalemic periodic paralysis (HYPP), has been defined genetically in humans [11]–[14] and horses [15]–[18]. Genetic studies have shown that in humans, HYPP is attributable to a channelopathy associated with abnormal sodium conductance, usually inherited as an autosomal dominant trait. Primary hypokalemic disorders have been documented in humans, manifesting as episodic weakness associated with low serum potassium [19].
Feline hypokalaemic periodic paralysis or Burmese hypokalaemic periodic polymyopathy (BHP) has been recognized since the seminal cases described by Blaxter and colleagues [20]. The disease is characterized by muscle weakness associated with intermittent hypokalemia [21]. Genetic studies suggest an autosomal recessive condition in Burmese cats [20]. Blaxter [20] provided a full description of the condition in the Burmese cat breed of the United Kingdom, Jones and collaborators recorded similar findings within Burmese cats from New Zealand [22], while Mason and Lantinga documented the condition in cats in Australia and the Netherlands, respectively [23], [24]. BHP has also been diagnosed elsewhere in Europe [15], however the disease has not been diagnosed within the Burmese cat population within the United States.
Classically, signs of BHP are episodic but in some cats, the weakness is incessant. During an episode, muscle pain (myalgia) from palpation can be a prominent sign. Cats can present with severe generalized muscle weakness, although more commonly weakness of the cervical muscles as evidenced by ventroflexion of the head and neck, head bobbing and dorsal protrusion of the scapulae [20], [23], [25]–[27]. The gait becomes short and maximal recruitment of motor units gives rise to muscle tremor. Cats with more generalized weakness have a crouching gait, especially evident in the hind limbs. Clinical signs of hypokalemia are thought to develop once serum potassium concentrations decreases below 3.0 mmol/l, although the rate with which the potassium concentration decreases and the lowest concentration achieved during an episode probably also influence the onset and the severity of signs. Although the course of the disease is usually affected by veterinary intervention, including potassium supplementation, serum potassium usually spontaneously normalizes after an episode. The disease usually becomes evident when kittens are two to six months of age, although some cases have not been detected until 2 years of age. Clinical signs may be triggered by stress or exercise. In some patients, the condition improves spontaneously or with therapy over a period of time, with some cats eventually not requiring on-going medication. However, some cats with BHP have life-time requirements for a potassium supplementation.
A genome-wide association study (GWAS) using the illumina Infinium feline 63K iSelect DNA array was performed on hypokalemic and healthy Burmese to localize the gene responsible for the hypokalemic periodic polymyopathy. A significant association was identified on cat chromosome E1, which led to a mutation discovery in WNK4. A genetic test has been developed and carrier cats can be easily identified prior to mating, avoiding the generation of affected cats. The WNK4 mutation in the Burmese cat is linked for the first time with hypokalemia in a mammalian species; a novel model for human disease.
Results
Clinical Presentation
Clinical presentation of cats diagnosed with hypokalemia included: episodic or incessant musculoskeletal weakness frequently characteristically by ventroflexion of the head and neck ( Figure 1 ). Myopathic weakness was additionally associated with a stiff/stilted gait, muscle tremor, head bobbing and dorsal protrusion of one or both scapulae. Some cats were thought to have myalgia on the basis of pain on muscle palpation or a shifting lameness, which resolved with oral potassium supplementation. In some cats, a weaving or a crouching gait, hypermetric forelimbs, a wide-based hind-limb stance were also observed.
Figure 1. Phenotypes of Burmese cats and Burmese affected with hypokalemia.

a. Unaffected Burmese of champagne coloration, b. hypokalemia affected Burmese of red coloration, displaying ventroflexion of the head and neck.
Most cats were diagnosed when young adults, between two and six months of age, although occasional cats were diagnosed at a later age. Although all cats considered as having hypokalemia had reduced [K+] (by definition) at some point in time, this was not always a consistent finding in affected cats, and sometimes it was necessary to collect sequential blood samples to identify the time when cats were unequivocally hypokalemic. Suggestive signs in Burmese cats and sometimes increased serum CK or aspartate aminotransferase (AST) activities were helpful in focusing the clinicians’ attention on the possibility of a myopathy. The demonstration of overt hypokalemia was further substantiated by demonstrating an improvement in muscle strength after oral potassium supplementation with potassium gluconate or chloride, with a concurrent return of serum muscle enzyme activities to the reference range.
Genome-wide Association Study
The affected Burmese cats originated from Australia, UK and Germany (samples from both countries were grouped in the Europe category), therefore unaffected, control cats were selected from the same countries with an established minimal relationship to the cases (
Table 1
). Seventy-two cats, including 38 cases (36 Burmese and 2 random bred cats) and 34 controls, were submitted for SNP array genotyping. Ten cats, including 1 case and 9 controls, had genotyping rates <90% and were excluded from analysis, but included in the cohort of samples that were genotyped for the identified mutation (
Table 1
). Testing for stratification by MDS revealed that the cats were not in one genetic cluster (
Figure 2a
). Even though the distribution of the cases and controls was not equally dense across the cluster, only two obvious affected outliers, the random bred cats, were identified and excluded from the analysis, thus the final number of cases included in the analyses was 35 Burmese. Additionally, 10 cases from the same geographical region, European Burmese, did not group together within the cluster (
Figure 2b
). Twelve samples had a 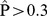 and the two random bred samples had a
and the two random bred samples had a 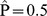 suggesting that the two samples are sibs. The detected genomic inflation factor (λ ) was 1.8 (
Figure 2c
). To reduce λ, several case-control studies were conducted after several samples pruning (Figure S1). The sample size range from 26–50 while the p-value varies from 7.22×10−5–1.66×10−9. The highest λ is detected when none of the Burmese samples is excluded from the analyses. The lowest λ = 1.09 is detected when the most genetically similar control is selected for each case (Table S2).
suggesting that the two samples are sibs. The detected genomic inflation factor (λ ) was 1.8 (
Figure 2c
). To reduce λ, several case-control studies were conducted after several samples pruning (Figure S1). The sample size range from 26–50 while the p-value varies from 7.22×10−5–1.66×10−9. The highest λ is detected when none of the Burmese samples is excluded from the analyses. The lowest λ = 1.09 is detected when the most genetically similar control is selected for each case (Table S2).
Table 1. Domestic cats genotyped for the mutation associated with hypokalemia.
| Breed | Origin | Phenotype | No. | Genotype | ||
| C/C | C/T | T/T | ||||
| USA | Normal | 16 | 16 | 0 | 0 | |
| Asia | Affected* | 2 | 0 | 0 | 2 | |
| Burmese | Europe (UK | Affected* + | 22 | 0 | 0 | 22 |
| + Germany) | Normal+ | 16 | 10 | 6 | 0 | |
| Australia | Affected* | 8 | 0 | 0 | 8 | |
| Normal | 0 | 0 | 0 | 0 | ||
| Random Bred | UK | Affected* | 2 | 0 | 0 | 2 |
| Pedigree | Europe | Affected* | 9 | 0 | 0 | 9 |
| Normal** | 26 | 10 | 16 | 0 | ||
| Bombay | USA | Normal | 13 | 13 | 0 | 0 |
| Random Bred | USA | Normal+ | 10 | 10 | 0 | 0 |
| Tonkinese | USA | Normal | 10 | 10 | 0 | 0 |
| Total | 134 |
Figure 2. Multi-dimensional scaling and QQ-plot of Burmese cats analyzed for hypokalemia.

a. Multi-dimensional scaling showing the distribution in 2 dimensions of the cases and controls of Burmese cats. The black asterisk indicates the location of the two random bred cat outliers excluded from the association analyses. b. Multi-dimensional scaling showing the differences between the geographical origins of the cats included in the analyses. The black asterisk indicates the 10 European Burmese cases that did not cluster with the majority of the Burmese samples included in the study. c. Q–Q plot showing an inflation in the test statistics with a λ = 1.8.
After evaluating the genotype qualities of the 62,897 SNPs on the array, 34,800 markers passed quality control, 27,152 SNPs had a MAF <5% and 1206 SNPs a GENO value >0.1. By analyzing 35 cases and 25 controls, a significant association was identified with several SNPs on chromosome E1 ( Figure 3 ), with the most significant association for a SNP at position 73,054,644 (the Whole Genome Shotgun project has been deposited at DDBJ/EML/GenBank under the accession AANG00000000. The sequence assembly used in this study is the second version, AANG02000000). A second association was detected with chromosome C2 with a SNP at position 5,583,090 (Table S2). After correcting P raw values for multiple hypotheses testing by permutation, six SNPs on chromosome E1 reached genome wide significance (P genome = 0.00002 for SNP E1 at position 73,054,644) and one less significant SNP on chromosome C2 position 5,583,090 (P genome = 0.046) (Figure S1). The strongest association on chromosome E1 was detected within a region between chromosome positions ∼72 to 73 Mb. A haplotype block was identified from position 73,054,644 to position 74,507,499 with a frequency of 75.7% across all cases, suggesting the most likely region associated with the disease (Figure S3). Within the same block, controls showed a 16.6% haplotype frequency. The two affected random bred samples excluded from the case-control study, showed a different haplotype compared to the affected Burmese cats. Inspection of genes within this haplotype block revealed two strong candidates involved in potassium level regulation, KCNH4 at ∼570 Kb and WNK4 at ∼1.2 Mb from the SNP with the highest association (Figure S4). The location of the two genes compared to the SNP position is presented in Figure S4. Since WNK4 is not annotated in the Felis catus genome, the distance between the two genes (KCNH4 and WNK4) in the human genome was used to infer the distance between the same genes in the cat assembly.
Figure 3. Manhattan plot summarizing the case – control GWAS for Burmese cats with hypokalemia.

The associated SNPs from 72 to 73 Mb on cat chromosome E1 (chromosome 14) obtained by analyzing 35 cases and 25 controls, correspond to a region containing the two candidate genes, KCNH4 at ∼570 Kb and WNK4 at ∼1.2 Mb from the SNP with the highest association (E1.73054644, P raw = 9.274×10−12 uncorrected).
KCNH4 and WNK4 Analyses
A partial KCNH4 (GenBank accession no. JQ522970) coding sequence (CDS), which was missing exons 1 and 13, was analyzed in four unrelated Burmese cats (two cases and two controls) and one random bred cat. Four variants were identified across the CDS and none were concordant with the phenotype (Table S3). Polymorphisms identified in the intronic sequence are reported in Table S3.
The entire CDS of WNK4 (GenBank accession no. JQ522971), including partial 5′ UTR and the 3′ UTR, was analyzed in the same four Burmese (2 cases and 2 controls) and a control random bred cat. In humans, WNK4 has one isoform and the length of the coding region within the transcript is 3,732 bp. In the cat, WNK4 has 19 exons, the boundaries were confirmed by genomic sequencing of the five cats used for the analyses of KCNH4, producing a 3,711 bp CDS that translates into 1236 amino acids. The differences between the two transcripts are located in exon 1 (3 bp), exon 13 (3 bp) and exon 14 (12 bp+3 bp). The average CDS identity between human and cat is 89.3% and at the protein level WNK4 is 90% identical to human. Three of the four identified exonic polymorphisms are missense mutations ( Table 2 ), the affected cats showed an AGTC haplotype and the controls an AGCT haplotype at the mutated positions. The first (c.340G>A) and second (c.2851G>A) result in alanine to threonine substitutions, the third mutation (c.2899C>T) causes a premature stop codon (CAG>TAG). Moreover, the mutation is validated by the RNA transcript sequence obtained from the affected Burmese cat blood. This mutation responsible for a premature WNK4 protein truncation lacking the C-terminal coiled coil domain and the Akt1/SGK phosphorylation site was further investigated. Several variants identified in the intronic regions ( Table 2 ) were highly variable and were not concordant with the disease phenotype in the cats analyzed.
Table 2. SNP analyses of WNK4 in cats with and without hypokalemia.
| Breed | Phenotype | c.340 G>A | c.285 G>A | c.2899 T>C | c.3516 T>C | −20E6 C>G | +37E6 T>C | +32E9 A>G | −80E11 G>− | −64E11 G>− | +92E19 G>A | +154E19 A>G |
| Burmese | Case | A | G | T | C | C | T | A | C | G | A | G |
| Burmese | Case | A | G | T | C | C | T | A | C | G | A | G |
| Burmese | Control | A | G | C | T | T | T | R | Y | G | R | R |
| Burmese | Control | A | G | C | T | T | T | A | C | G | G | A |
| Random Bred | Control | G | R | C | T | T | T | A | C | G | G | A |
| Trace | − | G | G | C | T | T | C | G | T | − | G | A |
| AA | change | A -> T | A -> T | Q -> X | − | − | − | − | − | − | − | − |
WNK4 Genotyping Analyses
A multi-generational pedigree consisting of 35 cats, segregating for hypokalemia ( Figure 4 ), was genotyped for eight STRs [28]. All the relationships between cats were confirmed as reported (data not show). Nine affected cats, with either carrier or affected parents, were confirmed by genotyping as homozygous for the c.2899C>T mutation. Of the 26 unaffected cats, ten were homozygous wildtype and 16 were carriers.
Figure 4. Burmese cat pedigree segregating for hypokalemia.

Circles represent females and squares represent males. Black symbols indicate cats with classical clinical signs for hypokalemia, white symbols indicate healthy cats. The +/+, +/−, and −/− signs indicate cats that tested homozygous wildtype, heterozygous, and homozygous affected for hypokalemia, respectively. Cats with a question mark in the symbol indicate that the phenotypic data was not available; symbols with a diagonal line indicate that no DNA sample was available. The asterisk indicates the four affected cats included in the GWA study, the number sign (#) indicate two of the controls used for the GWA study.
All the unaffected cats from other USA breeds (Bombay and Tonkinese), as well as random bred cats, were homozygous wildtype (n = 33). Fifty Burmese cats from Asia, Australia and Europe (Germany and UK), including 34 GWAS affected and 16 unaffected cats, with available potassium concentrations were genotyped and all the affected cats were homozygous for the c.2899C>T mutation; the unaffected individuals were either carriers or homozygous wildtype ( Table 1 ).
A cohort of 268 cats from different breeds including Asian, Australian Mist, Burmese, Burmilla, Tiffanie and 2 unknown cats was used to estimate the disease allele frequency. The frequency of the affected allele within the population is estimated as 14.9% (Table S4).
Discussion
Variations in the flux and balance of several ions, such as K+, Ca2+, Na+, and Cl− play key roles in specific tissue functions. In skeletal muscle, perturbations of these electrolytes are responsible for periodic paralyses, paramyotonia, and myotonia [1], [29]. Periodic paralyses constitute a group of hereditary muscle disorders characterized by acute and reversible attacks of muscle weakness associated with decrease blood pressure [30] in humans and is reported in other species such as, cow [31], horse [32], [33], rat and dog [34], [35]. Hypokalemic periodic paralysis is the most frequent form of periodic paralysis in humans, with an estimated prevalence of 1/100,000 [36]. Death due to paralysis of respiratory muscles [37] or from cardiac arrhythmia, secondary to a severe decrease in blood potassium concentration has been reported [38].
This case-control genome-wide association study was performed on a cohort comprising of 35 cases and 25 controls from the Burmese breed. Although the disease is usually episodic, definitive diagnosis can be made by the clinical presentations. Polymyopathy was reported in the domestic cat in 1983 [25] and hypokalemic periodic paralysis in the Burmese breed was first reported the UK and later identified in Australia and New Zealand [20], [22], [25], [26], [39]. However, Burmese cats in the USA have not been reported with hypokalemia. Due to differences in breed standards and health concerns, the Burmese in the USA are generally isolated from the Burmese in other countries. For the non-American-derived Burmese, diseases endangering the general health of the breed are hypokalemia [20], [25], diabetes mellitus [40] and an orofacial pain syndrome [41], [42]. The American-derived Burmese are highly associated with a craniofacial defect [43] and flat-chested kitten syndrome [44] is found in both breed populations. Amongst cat breeds, both the USA and non-USA Burmese populations present the highest linkage disequilibrium within cats (data not shown), likely due to geographical isolations and population bottlenecks. Due to these suspected population differences, USA-derived Burmese were not used as controls. In addition, 27,152 SNPs (43.1%) were excluded from the analysis due to MAFs <0.05, suggesting low genetic diversity in the breed, which was previously suggested by microsatellite analyses [45]. All the non-American Burmese clustered to some extent, indicating that some population sub-structure is present, which was supported by a genomic inflation factor of 1.8. The genomic inflation was reduced after each sample pruning, and got as low as 1.09 after excluding 34 samples. However, the lower samples size reduces power in detecting the association. When the samples size drops as low as 34, the SNP previously detecting the association is no longer the most significant association and the regional candidate would not be identified. To reduce genomic inflation, sib-pairs from a litter would be more ideal, however, as with most disease ascertainment, cases were collected over a long period and access to normal sibs or parents was not usually feasible.
The haplotype analysis of the 2.6 Mb region surrounding the most significant SNP association identified a 1.45 Mb block, with the most common haplotype having a frequency of >75% within the cases and ∼16% in the controls, supporting the region containing KCNH4 and WNK4 as strongly associated with the disease. The two related random bred cats were excluded from the association analyses, both were outliers in the MDS analyses and both had a completely different genetic background surrounding the associated SNPs as determined by the haplotype analysis. However, the random bred cats were homozygous for the identified hypokalemia mutation in the Burmese breed suggesting a mutation that precedes breed information. By fine mapping, a common short haplotype block with the controls would probably be identified.
Within ∼1.45 Mb of the highest associated SNP, two viable candidate genes, KCNH4 and WNK4, were identified. In KCNH4, no polymorphisms were identified between cases and controls, suggesting no differences between the affected and wild type alleles. However, sequencing of KCNH4 was not completed, hence the gene cannot be excluded as responsible for the disease. Sequencing of the second candidate gene, WNK4 identified a c.2899C>T nonsense mutation in the coding sequence. To confirm the association of the c.2899C>T mutation with phenotype, a multi-generational pedigree segregating for the disease was genotyped. All the affected cats tested homozygous for the premature stop codon, while the unaffected Burmese were either wildtype or heterozygous, supporting the c.2899C>T mutation as likely responsible for the disease. All identified cases in the study were homozygous for the mutation, while control Burmese were either heterozygous or wildtype. All available parents of the affected cats were heterozygous for the identified mutation. The genotyping was extended to a limited random bred population from South Asian and other South Asian - derived breeds, such as Bombay and Tonkinese, which use or have used Burmese in their breeding programs. The mutation was not identified within these cats. The initial estimate of the mutation frequency within the non-USA derived Burmese population, as well as in the breeds that use Burmese in outcross programs, is 14.9%. The frequency was obtained using data from a service laboratory from cats submitted for hypokalemia genetic testing, hence likely suspected to be at risk for disease, thus a positive bias in the data is expected. The genetic test will greatly assist breeders to avoid mating that could generate affected cats and identify carrier cats that could be excluded from the breeding program.
Point mutations in WNK4 have been associated in humans with the dominant disorder familial hyperkalemic hypertension and characterized by dysfunction in renal Na+, K+, and Cl− homeostasis [46]–[52]. Wilson et al. [50] originally identified WNK4 as a serine-threonine protein kinase that is expressed virtually exclusively in the kidney [53], localizing to the distal convoluted tubule and the cortical collecting duct [54], adjacent segments of the distal nephron that play a key role in salt, water, potassium, and pH homeostasis [50]. WNK4 is a member of the WNK kinases that have recently become a major focus of investigation because they comprise a previously unrecognized signaling pathway. The pathway appears to be essential for normal development, regulation of arterial pressure, normal electrolyte balance and for sensory nerve function, however the mechanisms by which WNK kinases regulate these processes are poorly understood [48], [49].
The WNK4 gene in humans is 16 Kb, contains 19 exons [50] and the protein is characterized by a kinase domain, an auto inhibitory autophosphorylation domain that lies in close proximity to the C-terminal end, two coiled-coils domains and proline-rich sequences [46]. The truncated protein in affected Burmese lacks the C-terminal coiled-coil domain, and is missing the highly conserved Akt1/SGK phosphorylation site at position S1169 [54]. Wang et al [55] demonstrated that truncated WNK4 constructs lacking the C terminus had no kinase activity toward themselves (autophosphorylation), hence WNK4 is constantly active. Studies suggest that when WNK4 is over-expressed, blood pressure is lower and when challenged with a low potassium diet, serum K+ is also reduced [56]. To date, studies of blood pressure in affected Burmese cats with hypokalemia have not been undertaken; it would therefore be informative to determine if cats are hypotensive during episodes of hypokalemia. Morphological studies in mice show that extra WNK4 copies reduced the abundance of the thiazide-sensitive Na-Cl cotransporter (NCC) in the kidney cortex [47], [57], [58]. The interaction between WNK4 and NCC, through its kinase domain, is suggested to result in high lysosomal degradation of NCC. When WNK4 is inhibited by autophosphorylation, NCC transfers to the membrane and Na+ and Cl− re-absorption activity is initiated. The deranged physiology seen in cats with BHP may be similar to the loss of function of NCC in humans [59] and mice [60], [61]. In hypokalemic cats, the WNK4 protein should be constantly active since the autophosphorylation site is missing and the protein constantly acts as an inhibitor for the NCC channel. When the NCC channel is not present on the cell surface, sodium gets reabsorbed by the epithelial channel (ENaC) [62], leaving a lumen Cl− negative potential that causes a secretion of K+ by the ATP-sensitive epithelial K+ channel (ROMK) channel [63]. The presumptive role of the C-terminal region of WNK in the cat can be proven and elucidated by functional studies using truncated transcripts in the future.
Materials and Methods
Animal Screening and Clinical Description
Thirty-eight cats with a diagnosis of hypokalemia were ascertained ( Table 1 , Figure 1 ). Most cases were diagnosed by the authors or by close colleagues; however, some were obtained by solicitation and submitted by owners who had received a BHP diagnosis for their cat from their primary care veterinarian. Burmese cats were verified as having BHP based on consistent clinical signs, seen in temporal association with serum potassium less than 3.0 mmol/l, and usually in concert with elevated creatine kinase (CK) activity in serum or plasma. Furthermore, all cats suspected of having hypokalemic polymyopathy had an obvious improvement in muscle strength with oral potassium supplementation (using potassium gluconate [4 mmol every 12 hours] or potassium chloride [3.5 mmol every 12 hours]). Unaffected cats (n = 34) had no clinical signs or owner/breeder reported evidence for BHP. No IACUC approvals were obtained for this study since samples were retained from patients as part of their primary care for disease diagnosis. Standards of veterinary care appropriate for an individual cat’s health and conditions were determined by the primary care veterinarian.
Genotyping
Thirty-eight cases (36 Burmese and 2 random bred cats) and 34 control Burmese were submitted to genotyping analysis. Whole blood samples were collected from affected cats as part of establishing their clinical diagnoses. Buccal swabs were donated for some affected and unaffected cats by Burmese breeders and owners. Moreover, a pedigree of cats segregating for hypokalemia was developed from a sub-set of submitted samples (four were included in the GWAS) ( Figure 4 ). Additional unaffected cats were selected from an available database of non-USA origin/domiciled Burmese cats. Genomic DNA was isolated from blood and buccal swab samples using the DNAeasy Kit (Qiagen, Valencia, CA) and concentrated using the Genomic DNA clean & Concentrator Kit (Zymo Research, Irvine, CA) when necessary. Quality and quantity of DNA was confirmed by visualization with agarose gel electrophoresis and by optical density using an UV exposure. Approximately 600 ng of genomic DNA was submitted to Neogene, Inc. (Lincoln, NE, USA) for genotyping on the illumina Infinium Feline 63K iSelect DNA array (illumina, Inc., San Diego, CA).
Genome Wide Association Study
SNP genotyping rate and minor allele frequency (MAF) was calculated using PLINK [64]. SNPs with a MAF <5%, genotyping rate <90%, and individuals genotyped for <90% of SNPs were excluded from further analyses. A classic multi-dimensional scaling (MDS) with 2 dimensions was performed on 56,811 SNPs in PLINK to evaluate population substructure within cases and controls. Inflation of p-values was evaluated by calculating the genomic inflation factor (λ) and was assessed with a quantile-quantile plot (Q-Q plot). A case-control association analysis was performed and corrected with 50,000 t-max permutations (-mperm 50,000). T-max permuted p-values were considered genome-wide significant at p<0.05. A Manhattan plot of the results was generated using HAPLOVIEW [65]. Linkage disequilibrium (LD) and haplotypes were determined considering SNPs from position 71,917,552 to position 74,507,499 of cat chromosome E1 and presented as plots produced by HAPLOVIEW [65]. The P (proportion of identical by state – IBS) for each individual was calculated using PLINK. To evaluate and reduce λ several strategies were applied, including, (i) exclusion of individuals with a proportion of IBD >0.3, (ii) selection of samples tightly clustered from the MDS plot and (iii) selection for each case to the closest control using the values from the MDS dimensions (Figure S1). After each correction, the number of cases and controls included in each analysis, as well as the p-value of the highest associated SNP, was evaluated and reported in Table S1.
KCNH4 Genomic Analyses
The genomic analyses of KCNH4 were conducted on genomic DNA from five cats including two cases and two controls from the Burmese breed and a random bred cat. Partial CDS of KCNH4 is publicly available (http://ensembl.org) and can be found on GeneScaffold_2463∶456,252-473,070. Missing exonic sequence was retrieved by aligning the individual human exons for KCNH4 (NM_012285.2) in the trace archive databases for Felis catus – WGS (http://blast.ncbi.nlm.nih.gov). Primers were designed in both UTRs and intronic regions, flanking the exons. Primers were tested for efficient product amplification on a DNA Engine Gradient Cycler (MJ Research, GMI, Ramsey, MN) and the final PCR magnesium concentrations, annealing temperatures, and amplicon sizes for each primer pair are shown in Table S5. PCR and thermocycling conditions were conducted as previously described [66]. The PCR products were purified with ExoSap (USB, Cleveland, OH) per the manufacturer’s recommendations and directly sequenced using the BigDye terminator Sequencing Kit v3.1 (Applied Biosystems, Foster City, CA). Sequences were verified and aligned using the software sequencer version 4.10 (Gene Codes Corp., Ann Arbor, MI).
WNK4 Genomic and mRNA Analyses
The genomic analysis of WNK4 was conducted as described for KCNH4. WNK4 was not annotated in the available cat genome assembly (Dec. 2008 NHGRI/GTB V17e/felCat4) and no sequence was publicly available. The gene was identified by comparison to the homologous region on human chromosome 17 corresponding to the associated cat region [67], [68]. Cat exonic sequences were retrieved from the cat trace archive using the individual exons of human WNK4 (NM_032387.4). The final PCR magnesium concentrations, annealing temperatures, and amplicon sizes for each primer pair are shown in Table S5. The PCR cycle was as follow: initial denaturation at 94°C for 4 min followed by 35 cycles as described: 94°C×30 sec, 65°C×30 sec, 72°C×30 sec and final extension at 72°C for 20 min.
Total RNA was extracted using the PAXgene™ (Qiagen) from whole blood from a control random bred cat and four affected Burmese cats from the UK. Complementary DNA templates were synthesized using SuperScript III (Invitrogen, Carlsband, CA) by reverse transcription of 1 µg of total RNA with gene specific primers (Table S5) and PolyT to obtain partial 5′ UTR, CDS and full 3′ UTR. Each cDNA sample was subjected to PCR using primers (10 µM each) combined as follow: 5utrF - 5utrR, F1 - R1, F2 - R2, F3 - R3, F4 - R4, F5 - R5, F6 - R6, F7 - R7, F8 - polyT. The PCR conditions were: 1.5 mM Mg, 2 µl of cDNA in a total volume of 20 µl. The PCR cycle was: initial denaturation at 94°C for 4 sec followed by a first set of 5 cycles at: 94°C×30 sec, 70°C×30 sec, 72°C×30 sec, 5 additional cycles at: 94°C×30 sec, 68°C×30 sec, 72°C×1 min and a set of 30 additional cycles at: 94°C×30 sec, 68°C×30 sec, 60°C×1 min and a final extension at 72°C for 20 min. The PCR products with appropriate lengths were purified using the ExoSap (USB) enzyme per manufacturer’s recommendations. Purified genomic products were directly sequenced in both directions using BigDye Terminator Sequencing Kit v3.1 (Applied Biosystems), purified with Illustra Sephadex G-50 (Ge Healthcare, Piscataway, NJ) according to manufacturer’s recommendations and electrophoretically separated on an ABI 3730 DNA analyzer (Applied Biosystems).
WNK4 Mutation Genotyping
To implicate the identified mutation as causative for hypokalemia, a pedigree segregating for hypokalemia with 35 available cats, was genotyped by direct sequencing ( Figure 4 ). Relationships within the pedigrees were confirmed with a published microsatellite-based parentage panel available for the cat (data not shown) [28]. Additional cats were also genotyped by direct sequencing to support correlation with disease, including 16 USA-derived Burmese, 16 unrelated unaffected, 34 affected GWAS Burmese (Asia, Europe, Australia), 2 affected random bred GWAS cats, 10 Tonkinese, 13 Bombay and 10 random bred cats ( Table 1 ). Primers for the direct sequencing genotyping are presented in Table S5.
Frequency Estimation
Owners submitted buccal swab samples of cats (n = 268) to determine status of the hypokalemia mutation to Langford Veterinary Services, UK (Table S4). Cats were genotyped for the WNK4 mutation to establish the allele frequency in the breeding population and to further confirm association with disease. Briefly, DNA was isolated from buccal swabs as previously described [69]. The WNK4 mutation was amplified by PCR; 12.5 µl 2× GoTaq PCR Master Mix (Promega, UK), 200 mM each Hypo For and Rev primers (Table S5) and 2µl gDNA in a total volume of 25 µl using a Bio-Rad MJ Mini (Life Science, Hertfordshire, UK) as described: 95°C for 2 min, 38 cycles of 95°C×15 sec, 58°C×30 sec by 35 cycles. Amplicons were subjected to pyrosequencing using sequencing primer Hypo Seq on a PyroMark Q24 (Qiagen) using Pyro Gold reagents as described by the manufacturer.
Supporting Information
MDS plots illustrating the samples included in each case-control association analyses of cats with hypokalemia. MDS plots showing a. the distribution of all the genotyped samples, b. the samples included in the case-control after exclusion of the related Burmese cats (p_hat >0.3), c. the distribution of tightly clustered Burmese samples, d. allowing more diversity than the cluster illustrated in (c), e. the closest control for each case, and f. the samples remaining after exclusion of related Burmese and outliers.
(TIF)
Manhattan plot of Burmese GWAS for hypokalemia. After correcting for multiple testing (50,000 permutations), five SNPs (four on chromosome E1 and one on chromosome C2) retained significant association (p<0.05; strongest SNP association, cat chromosome E1 position 73,054,644). The dashed line indicates genome wide significance.
(TIF)
Haplotype analyses of cats in hypokalemia GWAS. Presented is the area within ∼2.5 Mb of the highest associated SNP on chromosome E2 in the Burmese cases and controls. a. Cases haplotype block of the region surrounding the highest hit in the Burmese. b. From the highest hit, a block extending for ∼1.45 Mb (9 SNPs) was identified c. Cases haplotype frequencies, the red rectangle indicates the haplotype most frequently found within the affected cats. d. Controls haplotype block of the same region surrounding the highest hit within the cases. e. Figure representing the same block presented in (b) within the controls. f. Controls haplotype frequency, the red rectangle indicates the frequency across the controls of the most common haplotype identified within all the cases.
(TIF)
Haplotype within the ∼2.5 Mb on chromosome E1. ∼2.5 Mb are represented across all cases, controls and the two random bred cats excluded from the association study. The SNP in green across all samples represents the highest associated SNP with the disease. The orange bar represents the haplotype block shown in supplementary figure 2b and the relative position of the 2 candidate genes within the block is presented.
(PDF)
Evaluation of improved design to reduce genomic inflation (λ) in Burmese hypokalemia.
(DOC)
Details of seven highest associated SNPs for Burmese hypokalemia GWAS.
(DOC)
SNP analyses of KCNH4 in cats with and without hypokalemia.
(DOC)
Genotypes and frequencies of WNK4 SNP in cats submitted for genetic testing.
(DOC)
KCNH4 and WNK4 PCR primers for analysis of hypokalemia in cats.
(DOC)
Acknowledgments
We appreciate the solicitation for samples in Australia via The Centre for Veterinary Education through their Control and Therapy Series newsletter. Fran Musca, Marcus Gunew, Vic Menrath, Rhett Marshall and Steve Metcalf assisted by providing blood specimens from affected cats they had diagnosed, while John Culvenor was especially helpful in being both a veterinarian and an owner of an affected cat. Fran Musca assembled an unpublished series of cases, which was presented at the Meeting of the Australian College of Veterinary Scientists. We would like to thank the many Burmese owners and breeders who so generously contributed samples and time to this study, especially Claire Lovell, Tiina Räsänen, Anne Marie Bjelke, Irmgard Tormann, Susan van Keulen, Marja Virtanen, Anna Virtue, and Elizabeth Ward. The authors would like to thank several members of the Lyons’ laboratory for technical assistance, including Hasan Alhaddad, Robert Grahn, Grace Lan, and Carlyn B. Peterson.
Funding Statement
This work was supported by funding from the National Center for Research Resources R24 RR016094 and is currently supported by the Office of Research Infrastructure Programs/OD R24OD010928, the Cat Health Network grant D12FE-508, and the Center for Companion Animal Health, School of Veterinary Medicine, University of California, Davis. Richard Malik is supported by the Valentine Charlton Bequest from the University of Sydney. Support for the development of the Illumina Infinium Feline 63K iSelect DNA array was provided by the Morris Animal Foundation (http://www.morrisanimalfoundation.org) via a donation from Hill’s Pet Food, Inc. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Lehmann-Horn F, Jurkat-Rott K (1999) Voltage-gated ion channels and hereditary disease. Physiol Rev 79: 1317–1372. [DOI] [PubMed] [Google Scholar]
- 2.Schaefer TJ, Wolford RW (2005) Disorders of potassium. Emerg Med Clin North Am 23: 723–747, viii–ix. [DOI] [PubMed]
- 3. Layzer RB (1982) Periodic paralysis and the sodium-potassium pump. Ann Neurol 11: 547–552. [DOI] [PubMed] [Google Scholar]
- 4.Gabow PA, Peterson LN (1986) Disorders of Potassium Metabolism in Renal Electrolyte Disorders. Boston: Little Brown: 207–250.
- 5. Sterns RH, Cox M, Feig PU, Singer I (1981) Internal Potassium-Balance and the Control of the Plasma Potassium Concentration. Medicine 60: 339–354. [DOI] [PubMed] [Google Scholar]
- 6. Thier SO (1986) Potassium Physiology. American Journal of Medicine 80: 3–7. [DOI] [PubMed] [Google Scholar]
- 7. Schaer M (1982) Disorders of potassium metabolism. Vet Clin North Am Small Anim Pract 12: 399–409. [DOI] [PubMed] [Google Scholar]
- 8. Dow SW, Fettman MJ, Lecouteur RA, Hamar DW (1987) Potassium-Depletion in Cats - Renal and Dietary Influences. J Am Vet Med Assoc 191: 1569–1575. [PubMed] [Google Scholar]
- 9. Dow SW, Fettman MJ, Curtis CR, LeCouteur RA (1989) Hypokalemia in cats: 186 cases (1984–1987). J Am Vet Med Assoc 194: 1604–1608. [PubMed] [Google Scholar]
- 10. Fettman MJ (1989) Feline kaliopenic polymyopathy/nephropathy syndrome. Vet Clin North Am Small Anim Pract 19: 415–432. [DOI] [PubMed] [Google Scholar]
- 11. Gamstorp I (1957) Adynamia episodica hereditaria. Acta Genet Stat Med 7: 325–328. [DOI] [PubMed] [Google Scholar]
- 12. Bradley WG, Taylor R, Rice DR, Hausmanowa-Petruzewicz I, Adelman LS, et al. (1990) Progressive myopathy in hyperkalemic periodic paralysis. Arch Neurol 47: 1013–1017. [DOI] [PubMed] [Google Scholar]
- 13. Fontaine B, Khurana TS, Hoffman EP, Bruns GA, Haines JL, et al. (1990) Hyperkalemic periodic paralysis and the adult muscle sodium channel alpha-subunit gene. Science 250: 1000–1002. [DOI] [PubMed] [Google Scholar]
- 14. Rojas CV, Wang JZ, Schwartz LS, Hoffman EP, Powell BR, et al. (1991) A Met-to-Val mutation in the skeletal muscle Na+ channel alpha-subunit in hyperkalaemic periodic paralysis. Nature 354: 387–389. [DOI] [PubMed] [Google Scholar]
- 15. Naylor JM (1994) Equine hyperkalemic periodic paralysis: review and implications. Can Vet J 35: 279–285. [PMC free article] [PubMed] [Google Scholar]
- 16. Aleman M (2008) A review of equine muscle disorders. Neuromuscul Disord 18: 277–287. [DOI] [PubMed] [Google Scholar]
- 17. Rudolph JA, Spier SJ, Byrns G, Rojas CV, Bernoco D, et al. (1992) Periodic paralysis in quarter horses: a sodium channel mutation disseminated by selective breeding. Nat Genet 2: 144–147. [DOI] [PubMed] [Google Scholar]
- 18. Spier SJ, Carlson GP, Holliday TA, Cardinet GH, 3rd, Pickar JG (1990) Hyperkalemic periodic paralysis in horses. J Am Vet Med Assoc 197: 1009–1017. [PubMed] [Google Scholar]
- 19. Finsterer J (2008) Primary periodic paralyses. Acta Neurologica Scandinavica 117: 145–158. [DOI] [PubMed] [Google Scholar]
- 20. Blaxter A, Lievesley P, Gruffyddjones T, Wotton P (1986) Periodic Muscle Weakness in Burmese Kittens. Veterinary Record 118: 619–620. [DOI] [PubMed] [Google Scholar]
- 21. Jones BR, Gruffydd-Jones TJ (1990) Hypokalemia in the Cat. Cornell Veterinarian 80: 13–16. [PubMed] [Google Scholar]
- 22. Jones BR, Swinney GW, Alley MR (1988) Hypokalaemic myopathy in Burmese kittens. New Zealand Veterinary Journal 36: 150–151. [DOI] [PubMed] [Google Scholar]
- 23. Lantinga E, Kooistra HS, van Nes JJ (1998) [Periodic muscle weakness and cervical ventroflexion caused by hypokalemia in a Burmese cat]. Tijdschr Diergeneeskd 123: 435–437. [PubMed] [Google Scholar]
- 24. Mason KV (1988) Hereditary Potassium-Depletion in Burmese Cats. Journal of the American Animal Hospital Association 24: 481–481. [Google Scholar]
- 25.Schunk KL (1984) Feline Polymyopathy. Proc 2nd Annual Forum Am Coll Vet Int Med: 197–200.
- 26. Dow SW, Lecouteur RA, Fettman MJ, Spurgeon TL (1987) Potassium-Depletion in Cats - Hypokalemic Polymyopathy. J Am Vet Med Assoc 191: 1563–1568. [PubMed] [Google Scholar]
- 27. Stolze M, Lund C, Kresken JG, Saerg KJ (2001) Periodic hypokalemic polymyopathy in the Burmese cat. Kleintierpraxis 46: 517–518. [Google Scholar]
- 28. Lipinski MJ, Amigues Y, Blasi M, Broad TE, Cherbonnel C, et al. (2007) An international parentage and identification panel for the domestic cat (Felis catus). Animal Genetics 38: 371–377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Hubner CA, Jentsch TJ (2002) Ion channel diseases. Hum Mol Genet 11: 2435–2445. [DOI] [PubMed] [Google Scholar]
- 30. Venance SL, Cannon SC, Fialho D, Fontaine B, Hanna MG, et al. (2006) The primary periodic paralyses: diagnosis, pathogenesis and treatment. Brain 129: 8–17. [DOI] [PubMed] [Google Scholar]
- 31. Sielman ES, Sweeney RW, Whitlock RH, Reams RY (1997) Hypokalemia syndrome in dairy cows: 10 cases (1992–1996). J Am Vet Med Assoc 210: 240–243. [PubMed] [Google Scholar]
- 32. Steiss JE, Naylor JM (1986) Episodic muscle tremors in a quarter horse: resemblance to hyperkalemic periodic paralysis. Can Vet J 27: 332–335. [PMC free article] [PubMed] [Google Scholar]
- 33.Cox JH (1986) An episodic weakness in four horses associated with intermittent serum hyperkalemia and the similarity of the disease to hyperkalemic periodic paralysis in man. Proceedings of the annual convention of the American Association of Equine Practitioners: 383–391.
- 34.Smith SG, Black-Schaffer B, Lasater TE (1950) Potassium deficiency syndrome in the rat and the dog; a description of the muscle changes in the potassium-depleted dog. AMA Arch Pathol 49: 185–199, illust. [PubMed]
- 35. Harrington ML (1996) Suspect hypokalemic myopathy in a dog. Progress in Veterinary Neurology 7: 130–132. [Google Scholar]
- 36. Talbott JH (1941) Periodic paralysis - A clinical syndrome. Medicine 20: 85–143. [Google Scholar]
- 37. Holtzapple GE (1905) Periodic paralysis. J Am Med Assoc 45: 1224–1238. [Google Scholar]
- 38.Levitt LP, Rose LI, Dawson DM (1972) Hypokalemic Periodic Paralysis with Arrhythmia. New England Journal of Medicine 286: 253–&. [DOI] [PubMed]
- 39. Hopkins AL (1989) Sporadic Feline Hypokalemic Polymyopathy. Veterinary Record 125: 17–17. [DOI] [PubMed] [Google Scholar]
- 40. Lutz TA, Rand JS (1995) Pathogenesis of feline diabetes mellitus. Vet Clin North Am Small Anim Pract 25: 527–552. [DOI] [PubMed] [Google Scholar]
- 41. Roche GM (1994) Irritation from Erupting Teeth. Veterinary Record 134: 360–360. [DOI] [PubMed] [Google Scholar]
- 42. Heath S, Rusbridge C, Johnson N, Gunn-Moore D (2001) Orofacial pain syndrome in cats. Veterinary Record 149: 660–660. [PubMed] [Google Scholar]
- 43.Noden DM, Evans HE (1986) Inherited Homeotic Midfacial Malformations in Burmese Cats. Journal of Craniofacial Genetics and Developmental Biology: 249–266. [PubMed]
- 44. Sturgess CP, Waters L, Gruffydd-Jones TJ, Nott HMR, Earle KE (1997) Investigation of the association between whole blood and tissue taurine levels and the development of thoracic deformities in neonatal Burmese kittens. Veterinary Record 141: 566–570. [DOI] [PubMed] [Google Scholar]
- 45. Lipinski MJ, Froenicke L, Baysac KC, Billings NC, Leutenegger CM, et al. (2008) The ascent of cat breeds: genetic evaluations of breeds and worldwide random-bred populations. Genomics 91: 12–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. McCormick JA, Ellison DH (2011) The WNKs: Atypical Protein Kinases With Pleiotropic Actions. Physiol Rev 91: 177–219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Kahle KT, Wilson FH, Leng Q, Lalioti MD, O’Connell AD, et al. (2003) WNK4 regulates the balance between renal NaCl reabsorption and K+ secretion. Nature Genetics 35: 372–376. [DOI] [PubMed] [Google Scholar]
- 48. Paver WK, Pauline GJ (1964) Hypertension and Hyperpotassaemia without Renal Disease in a Young Male. Med J Aust 2: 305–306. [DOI] [PubMed] [Google Scholar]
- 49. Proctor G, Linas S (2006) Type 2 pseudohypoaldosteronism: new insights into renal potassium, sodium, and chloride handling. Am J Kidney Dis 48: 674–693. [DOI] [PubMed] [Google Scholar]
- 50. Wilson FH, Disse-Nicodeme S, Choate KA, Ishikawa K, Nelson-Willams C, et al. (2001) Human hypertension caused by mutations in WNK kinases. Science 293: 1107–1112. [DOI] [PubMed] [Google Scholar]
- 51. Mayan H, Munter G, Shaharabany M, Mouallem M, Pauzner R, et al. (2004) Hypercalciuria in familial hyperkalemia and hypertension accompanies hyperkalemia and precedes hypertension: Description of a large family with the Q565E WNK4 mutation. Journal of Clinical Endocrinology & Metabolism 89: 4025–4030. [DOI] [PubMed] [Google Scholar]
- 52. Golbang AP, Murthy M, Hamad A, Liu CH, Cope G, et al. (2005) A new kindred with Pseudohypoaldosteronism type II and a novel mutation (564D>H) in the acidic motif of the WNK4 gene. Hypertension 46: 295–300. [DOI] [PubMed] [Google Scholar]
- 53. Kahle KT, Gimenez I, Hassan H, Wilson FH, Wong RD, et al. (2004) WNK4 regulates apical and basolateral Cl- flux in extrarenal epithelia. Proc Natl Acad Sci U S A 101: 2064–2069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Ring AM, Leng Q, Rinehart J, Wilson FH, Kahle KT, et al. (2007) An SGK1 site in WNK4 regulates Na+ channel and K+ channel activity and has implications for aldosterone signaling and K+ homeostasis. Proc Natl Acad Sci U S A 104: 4025–4029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Wang ZH, Yang CL, Ellison DH (2004) Comparison of WNK4 and WNK1 kinase and inhibiting activities. Biochemical and Biophysical Research Communications 317: 939–944. [DOI] [PubMed] [Google Scholar]
- 56. Lalioti MD, Zhang JH, Volkman HM, Kahle KT, Hoffmann KE, et al. (2006) Wnk4 controls blood pressure and potassium homeostasis via regulation of mass and activity of the distal convoluted tubule. Nature Genetics 38: 1124–1132. [DOI] [PubMed] [Google Scholar]
- 57. Wilson FH, Kahle KT, Sabath E, Lalioti MD, Rapson AK, et al. (2003) Molecular pathogenesis of inherited hypertension with hyperkalemia: The Na-Cl cotransporter is inhibited by wild-type but not mutant WNK4. Proceedings of the National Academy of Sciences of the United States of America 100: 680–684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Yang CL, Angell J, Mitchell R, Ellison DH (2003) WNK kinases regulate thiazide-sensitive Na-Cl cotransport. Journal of Clinical Investigation 111: 1039–1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Simon DB, NelsonWilliams C, Bia MJ, Ellison D, Karet FE, et al. (1996) Gitelman’s variant of Bartter’s syndrome, inherited hypokalaemic alkalosis, is caused by mutations in the thiazide-sensitive Na-Cl cotransporter. Nature Genetics 12: 24–30. [DOI] [PubMed] [Google Scholar]
- 60. Schultheis PJ, Lorenz JN, Meneton P, Nieman ML, Riddle TM, et al. (1998) Phenotype resembling Gitelman’s syndrome in mice lacking the apical Na+-Cl- cotransporter of the distal convoluted tubule. Journal of Biological Chemistry 273: 29150–29155. [DOI] [PubMed] [Google Scholar]
- 61. Loffing J, Vallon V, Loffing-Cueni D, Aregger F, Richter K, et al. (2004) Altered renal distal tubule structure and renal Na+ and Ca2+ handling in a mouse model for Gitelman’s syndrome. Journal of the American Society of Nephrology 15: 2276–2288. [DOI] [PubMed] [Google Scholar]
- 62. Canessa CM, Schild L, Buell G, Thorens B, Gautschi I, et al. (1994) Amiloride-Sensitive Epithelial Na+ Channel Is Made of 3 Homologous Subunits. Nature 367: 463–467. [DOI] [PubMed] [Google Scholar]
- 63. Ho K, Nichols CG, Lederer WJ, Lytton J, Vassilev PM, et al. (1993) Cloning and Expression of an Inwardly Rectifying Atp-Regulated Potassium Channel. Nature 362: 31–38. [DOI] [PubMed] [Google Scholar]
- 64. Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, et al. (2007) PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet 81: 559–575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Barrett JC, Fry B, Maller J, Daly MJ (2005) Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21: 263–265. [DOI] [PubMed] [Google Scholar]
- 66. Bighignoli B, Niini T, Grahn RA, Pedersen NC, Millon LV, et al. (2007) Cytidine monophospho-N-acetylneuraminic acid hydroxylase (CMAH) mutations associated with the domestic cat AB blood group. BMC Genet 8: 27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. O’Brien SJ, Cevario SJ, Martenson JS, Thompson MA, Nash WG, et al. (1997) Comparative gene mapping in the domestic cat (Felis catus). J Hered 88: 408–414. [DOI] [PubMed] [Google Scholar]
- 68. O’Brien SJ, Wienberg J, Lyons LA (1997) Comparative genomics: lessons from cats. Trends Genet 13: 393–399. [DOI] [PubMed] [Google Scholar]
- 69. Helps CR, Tasker S, Barr FJ, Wills SJ, Gruffydd-Jones TJ (2007) Detection of the single nucleotide polymorphism causing feline autosomal-dominant polycystic kidney disease in Persians from the UK using a novel real-time PCR assay. Molecular and Cellular Probes 21: 31–34. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
MDS plots illustrating the samples included in each case-control association analyses of cats with hypokalemia. MDS plots showing a. the distribution of all the genotyped samples, b. the samples included in the case-control after exclusion of the related Burmese cats (p_hat >0.3), c. the distribution of tightly clustered Burmese samples, d. allowing more diversity than the cluster illustrated in (c), e. the closest control for each case, and f. the samples remaining after exclusion of related Burmese and outliers.
(TIF)
Manhattan plot of Burmese GWAS for hypokalemia. After correcting for multiple testing (50,000 permutations), five SNPs (four on chromosome E1 and one on chromosome C2) retained significant association (p<0.05; strongest SNP association, cat chromosome E1 position 73,054,644). The dashed line indicates genome wide significance.
(TIF)
Haplotype analyses of cats in hypokalemia GWAS. Presented is the area within ∼2.5 Mb of the highest associated SNP on chromosome E2 in the Burmese cases and controls. a. Cases haplotype block of the region surrounding the highest hit in the Burmese. b. From the highest hit, a block extending for ∼1.45 Mb (9 SNPs) was identified c. Cases haplotype frequencies, the red rectangle indicates the haplotype most frequently found within the affected cats. d. Controls haplotype block of the same region surrounding the highest hit within the cases. e. Figure representing the same block presented in (b) within the controls. f. Controls haplotype frequency, the red rectangle indicates the frequency across the controls of the most common haplotype identified within all the cases.
(TIF)
Haplotype within the ∼2.5 Mb on chromosome E1. ∼2.5 Mb are represented across all cases, controls and the two random bred cats excluded from the association study. The SNP in green across all samples represents the highest associated SNP with the disease. The orange bar represents the haplotype block shown in supplementary figure 2b and the relative position of the 2 candidate genes within the block is presented.
(PDF)
Evaluation of improved design to reduce genomic inflation (λ) in Burmese hypokalemia.
(DOC)
Details of seven highest associated SNPs for Burmese hypokalemia GWAS.
(DOC)
SNP analyses of KCNH4 in cats with and without hypokalemia.
(DOC)
Genotypes and frequencies of WNK4 SNP in cats submitted for genetic testing.
(DOC)
KCNH4 and WNK4 PCR primers for analysis of hypokalemia in cats.
(DOC)