

Abstract
Highly virulent pantropic canine coronavirus (CCoV) strains belonging to subtype IIa were recently identified in dogs. To assess the distribution of such strains in Europe, tissue samples were collected from 354 dogs that had died after displaying systemic disease in France (n = 92), Hungary (n = 75), Italy (n = 69), Greece (n = 87), The Netherlands (n = 27), Belgium (n = 4), and Bulgaria (n = 1). A total of 124 animals tested positive for CCoV, with 33 of them displaying the virus in extraintestinal tissues. Twenty-four CCoV strains (19.35% of the CCoV-positive dogs) detected in internal organs were characterized as subtype IIa and consequently assumed to be pantropic CCoVs. Sequence and phylogenetic analyses of the 5′ end of the spike protein gene showed that pantropic CCoV strains are closely related to each other, with the exception of two divergent French viruses that clustered with enteric strains.
INTRODUCTION
Coronaviruses (CoVs; order Nidovirales, family Coronaviridae) are positive-sense RNA viruses commonly associated with mild infections in birds and mammals. The family Coronaviridae is now organized into two subfamilies, Coronavirinae and Torovirinae, with the former including four different genera, Alphacoronavirus, Betacoronavirus, Gammacoronavirus, and Deltacoronavirus. Canine coronavirus (CCoV) belongs to the genus Alphacoronavirus and forms a unique species, Alphacoronavirus 1, along with feline coronaviruses (FCoVs), transmissible gastroenteritis virus of swine (TGEV), and its derivative, porcine respiratory coronavirus (PRCoV) (1, 2).
CCoVs are paradigmatic of the CoV genetic evolution and complexity (3, 4). In addition to the known genotypes, CCoV types I (CCoV-I) and II (CCoV-II) (5), which share up to 96% of nucleotide sequence identity in the viral genome but are highly divergent in the spike (S) protein gene (6), CCoV subtypes and biotypes have been more recently identified (3, 4). Detection of TGEV/CCoV recombinant strains led to the classification of CCoV-II into two subtypes, including the classical (CCoV-IIa) and recombinant (CCoV-IIb) subtypes, respectively (7, 8). A hypervirulent CCoV-IIa strain, designated pantropic CCoV, was isolated from dead pups at a pet shop in Italy in 2005 (9). The virus, strain CB/05, was associated with severe clinical signs and postmortem lesions. Experimental infections of dogs reproduced the disease, with the severity varying with the age and immune status of the infected animals (10, 11) but invariably leading to long-term lymphopenia (12).
Natural outbreaks of pantropic CCoV infection have been reported in France and Belgium (13), Greece (14), and Italy (15).
The aim of the present study is to report the detection of pantropic CCoV in some European countries.
MATERIALS AND METHODS
Sample collection.
A total of 354 carcasses of dogs that died after displaying systemic disease consisting of fever, leukopenia, depression, enteritis, respiratory distress, and/or neurological signs were sampled from 2009 to 2011 (Table 1). Cases were admitted to the study if they showed two or more of these clinical signs. Dogs for sample collection were recruited in animal shelters, breeding kennels, and academic or private veterinary clinics of different European countries (Table 1), including Italy (n = 69), France (n = 92), Greece (n = 87), Hungary (n = 75), The Netherlands (n = 27), Belgium (n = 4), and Bulgaria (n = 1). When possible, the entire carcass was sent to the different laboratories, but on several occasions, only tissue samples that were collected from the gut (or a rectal swab), lung, liver, spleen, kidney, tonsil, thymus, and/or other tissues were available. A total of 2,129 canine tissue samples were analyzed. Samples were screened by the different laboratories, and results were confirmed by the Infectious Diseases Unit of the Department of Veterinary Medicine of Bari, where further molecular investigations were conducted.
Table 1.
Survey for pantropic CCoV in European countries
| Country | No. of dogs sampled | No. CCoV positive |
No. of dogs CCoV-IIa positive | |
|---|---|---|---|---|
| Dogs | Internal organs | |||
| France | 92 | 57 | 11 | 7 |
| Hungary | 75 | 18 | 7 | 7 |
| Italy | 69 | 12 | 7 | 6 |
| Greece | 87 | 22 | 3 | 1 |
| The Netherlands | 27 | 12 | 4 | 2 |
| Belgium | 4 | 3 | 1 | 1 |
| Bulgaria | 1 | 0 | 0 | 0 |
| Total | 354 | 124 | 33 | 24 |
RNA extraction.
Tissues were homogenized (10%, wt/vol) in Dulbecco's modified Eagle's medium (DMEM) and subsequently clarified by centrifuging at 2,500 × g for 10 min. One hundred forty microliters of the supernatants was then used for RNA extraction by means of a QIAamp viral RNA minikit (Qiagen S.p.A., Milan, Italy), following the manufacturer's protocol, and the RNA templates were stored at −70°C until their use.
CCoV RNA detection, quantification, genotyping, and subtyping.
All RNA extracts were subjected to a previously established TaqMan-based real-time reverse transcription-PCR (RT-PCR) assay for rapid detection and quantification of CCoV RNA (16), with minor modifications. Briefly, a one-step method was adopted using Platinum quantitative PCR SuperMix-UDG (Invitrogen srl, Milan, Italy) and a 50-μl mixture of the following: 25 μl of master mix, 300 nM primers CCoV-F and CCoV-R, 200 nM probe CCoV-Pb (Table 2), and 10 μl of template RNA. Duplicates of log10 dilutions of standard RNA were analyzed simultaneously in order to obtain a standard curve for absolute quantification (16). The thermal profile consisted of incubation with uracil-DNA glycosylase (UDG) at 50°C for 2 min and activation of Platinum Taq DNA polymerase at 95°C for 2 min, followed by 45 cycles of denaturation at 95°C for 15 s, annealing at 48°C for 30 s, and extension at 60°C for 30 s.
Table 2.
Oligonucleotides used for detection and characterization of CCoV strains
| Test | Primer/probe | Reference or source | Sequence (5′–3′)d | Sense | Position | Amplicon size (bp) | Specificity |
|---|---|---|---|---|---|---|---|
| Real-time RT-PCR | CCoV-F | 16 | TTGATCGTTTTTATAACGGTTCTACAA | + | 6585–6611a | 99 | CCoV-I/II |
| CCoV-R | AATGGGCCATAATAGCCACATAAT | − | 6660–6683a | ||||
| CCoV-Pb | FAM-ACCTCAATTTAGCTGGTTCGTGTATGGCATT-TAMRA | + | 6620–6650a | ||||
| Real-time RT-PCR | CCoVI-F | 17 | CGTTAGTGCACTTGGAAGAAGCT | + | 478–499b | 111 | CCoV-I |
| CCoVI-R | ACCAGCCATTTTAAATCCTTCA | − | 567–588b | ||||
| CCoVI-Pb | FAM-CCTCTTGAAGGTACACCAA-TAMRA | + | 508–526b | ||||
| Real-time RT-PCR | CCoVII-F | 17 | TAGTGCATTAGGAAGAAGCT | + | 6878–6897a | 105 | CCoV-II |
| CCoVII-R | AGCAATTTTGAACCCTTC | − | 6966–6982a | ||||
| CCoVII-Pb | FAM-CCTCTTGAAGGTGTGCC-TAMRA | + | 6906–6922a | ||||
| RT-PCR | 20179 | 8 | GGCTCTATCACATAACTCAGTCCTAG | + | 320–345a | 758 (CCoV-IIa) | CCoV-I/II |
| 12531–12556c | |||||||
| INS-R-dg | GCTGTAACATAKTCRTCATTCCAC | − | 1054–1077a | 499 (CCoV-IIb) | CCoV-IIa | ||
| 174-268 | CAACATGTAACCTTTGTCTGTGATCTGC | − | 13002–13029c | CCoV-IIb | |||
| RT-PCR | VNVF | This study | ACTTGTGTGTATAGGTTTTGGTGA | + | 5157–5180a | 530 (intact genes) | CCoV-I/II |
| VNVR | TAAGTGTCATTGATACAATCTTAAACA | − | 5660–5686a |
Oligonucleotide position in reference to the sequence of CCoV-I strain 259/01 (GenBank accession number AF502583).
Oligonucleotide position in reference to the sequence of CCoV-IIa strain Insavc-1 (GenBank accession number D13096).
Oligonucleotide position in reference to the sequence of CCoV-IIb strain 174/06 (GenBank accession number EU856362).
FAM, 6-carboxyfluorescein; TAMRA, 6-carboxytetramethylrhodamine.
The detected CCoV strains were characterized by means of two distinct genotype-specific assays (17) performed by using Platinum quantitative PCR SuperMix-UDG (Invitrogen srl) and the following oligonucleotide sets (final concentrations, 600 and 200 nM for primers and probes, respectively): primer pair CCoVI-F/CCoVI-R and probe CCoVI-Pb for CCoV-I and primer pair CCoVII-F/CCoVII-R and probe CCoVII-Pb (Table 2) for CCoV-II. The thermal protocol was as described for CCoV detection, except for the use of different annealing temperatures (53°C and 48°C for CCoV-I and CCoV-II, respectively).
Samples that tested positive for CCoV-II were subjected to subtype-specific CCoV-IIa and CCoV-IIb gel-based RT-PCR assays targeting the spike (S) protein gene and using primer pair 20179/INS-R-dg or 20179/174-268 (8) (Table 2). Reactions were carried out with the SuperScript one-step RT-PCR for long templates (Invitrogen srl) and the following thermal protocol: reverse transcription at 50°C for 30 min, inactivation of SuperScript II reverse transcriptase at 94°C for 2 min, and 40 cycles of 94°C for 30 s, 55°C for 30 s, and 68°C for 30 s, with a final extension at 68°C for 10 min. The PCR products were detected by electrophoresis through a 1.5% agarose gel and visualization under UV light after bromide ethidium staining.
The presence of deletions in the accessory protein genes 3abc was searched for by using primers VNVF and VNVR (Table 2) and the same thermal protocol adopted for CCoV subtyping. These primers were designed to amplify a fragment encompassing the full-length 3abc region. Pantropic CCoV strains NA/9 (14) and CB/05 (9) were used as controls for intact and deleted genes, respectively.
Virus isolation.
Tissue samples containing discrete viral titers (not less than 104 copies of CCoV-IIa RNA μl−1 of template) were homogenized (10%, wt/vol) in DMEM together with antibiotics (penicillin, 5,000 IU/ml; streptomycin, 2,500 μg/ml; amphotericin B, 10 μg/ml) and inoculated into canine fibroma (A-72) cells. Infected cells were monitored daily for the occurrence of a cytopathic effect (CPE), and after 5 days of incubation, they were tested for CCoV antigen by an immunofluorescence (IF) assay using a monoclonal antibody targeting the Alphacoronavirus 1 nucleocapsid protein (courtesy of Gilles Chappuis, Merial, France).
Sequence analysis and phylogeny.
The RT-PCR products obtained with primer pair 20179/INS-R-dg from dogs that tested CCoV positive in one or more extraintestinal tissues (and thus assumed to be infected by pantropic CCoV) were subjected to direct sequencing at BaseClear B.V. (Leiden, The Netherlands). The sequences were manually edited and analyzed using the BioEdit software package (18) and the NCBI (http://www.ncbi.nlm.nih.gov) and EMBL (http://www.ebi.ac.uk) analysis tools. Phylogenetic and molecular evolutionary analyses were conducted using the Mega4.1 Beta program (19). Phylogenetic trees based on the 5′ end (411 to 414 nucleotides) of the S-protein gene were elaborated using both parsimony and neighbor-joining methods, supplying statistical support with bootstrapping of over 1,000 replicates. The distantly related Betacoronavirus canine respiratory coronavirus 240/05 (20, 21) was used as the outgroup.
Detection of other viral pathogens.
Tissue samples of dogs that tested positive for pantropic CCoV were submitted to molecular detection of the main viral pathogens of dogs, such as reoviruses (22), rotaviruses (23), caliciviruses (24), canine parvovirus type 2 (CPV-2) (25, 26), canine adenoviruses type 1 and type 2 (27), canine distemper virus (28), canine minute virus (29), canid herpesvirus type 1 (30), and canine respiratory coronavirus (CRCoV) (31).
Nucleotide sequence accession numbers.
The nucleotide sequences of the analyzed pantropic strains were deposited in GenBank under accession numbers JQ929041 to JQ929046 (Italian strains), JF682842 (Greek strain), and JX269009 to JX269018 (strains from other countries).
RESULTS
Identification of pantropic CCoV strains.
CCoV RNA was detected in 124 (35.02%) out of 354 sampled dogs, with 33 animals testing CCoV positive in one or more extraintestinal tissues (Table 1). Twenty-four of these strains were characterized as CCoV-IIa (19.35% of the CCoV-positive dogs), whereas the remaining strains were recognized as TGEV/CCoV recombinant viruses (CCoV-IIb) that can occasionally spread to internal organs. The geographic distribution of the CCoV-IIa strains detected in extraintestinal tissues is reported in Table 1.
The viral RNA titers observed in the internal organs of the 24 dogs assumed to be infected by pantropic CCoV (CCoV-IIa positive in internal organs) were generally low, ranging from 1.20 × 101 (liver of an Italian dog) to 8.16 × 105 (lung of a Hungarian dog) copies μl−1 of template (Fig. 1). The highest median CCoV titers were detected in the bladder (1.32 × 104 copies μl−1 of template) and lymphoid tissues (2.77 × 103 and 2.12 × 103 copies μl−1 of template for the mesenteric lymph node and thymus, respectively) of infected dogs (Fig. 1).
Fig 1.

Viral RNA titers μl−1 of template detected in different tissues of dogs that tested positive for pantropic CCoV. Each symbol represents a clinical sample; bars indicate the median values. ln, lymph node.
Virus isolation attempts were not successful for any of the tissue samples displaying the highest copy numbers of viral RNA, as shown by the absence of a CPE and negative IF testing even after three consecutive passages on cell cultures.
Genetic characterization of pantropic CCoV strains.
By RT-PCR amplification of the accessory protein genes 3abc, none of the potential pantropic strains displayed any detectable deletion in this region. The 5′ end of the S-protein gene was determined for 17 out of the 24 pantropic strains, which were representative of the different geographic areas, including 4/7 French, 3/7 Hungarian, 6/6 Italian, 1/1 Greek, 2/2 Dutch, and 1/1 Belgian viruses. Sequence analysis showed the lack of specific genetic markers that were common to all obtained sequences. The mutation D125N, identified in both Italian strains currently sequenced (9, 15), was detected in 9 out of 17 viruses of the present survey (Fig. 2).
Fig 2.

Alignment of the N-terminal domain of the spike protein of pantropic and enteric CCoV-IIa strains. The residues displaying the D125N mutation are boxed.
Despite the absence of common markers, most pantropic viruses except two French strains (322/09 and 45/10-6) were closely related to each other, displaying nucleotide and amino acid identities ranging from 92% to 100% and from 90.5% to 100%, respectively. These viruses showed a close genetic relatedness to reference strains CB/05 and 450/07 (nucleotide and amino acid identities ranging from 94.4% to 97.5% and from 91.3% to 99.2%, respectively) (see Table S1 in the supplemental material). Strains 322/09 and 45/10-6 diverged from the pantropic group, showing the highest nucleotide (79.9%) and amino acid (78.9%) identities with Hungarian or Italian viruses, whereas they were more closely related to the enteric virus 03-3684 (nucleotide identity of 90% to 90.2%), which was recently detected in the United Kingdom (32).
By phylogeny conducted on the same S-protein gene region, most pantropic CCoV strains fell in the same cluster as Italian prototype viruses, which was separated from the clusters grouping enteric CCoV-IIa strains. Again, the French viruses 322/09 and 45/10-6 were found to diverge from other pantropic CCoVs, clustering with enteric strains (Fig. 3).
Fig 3.
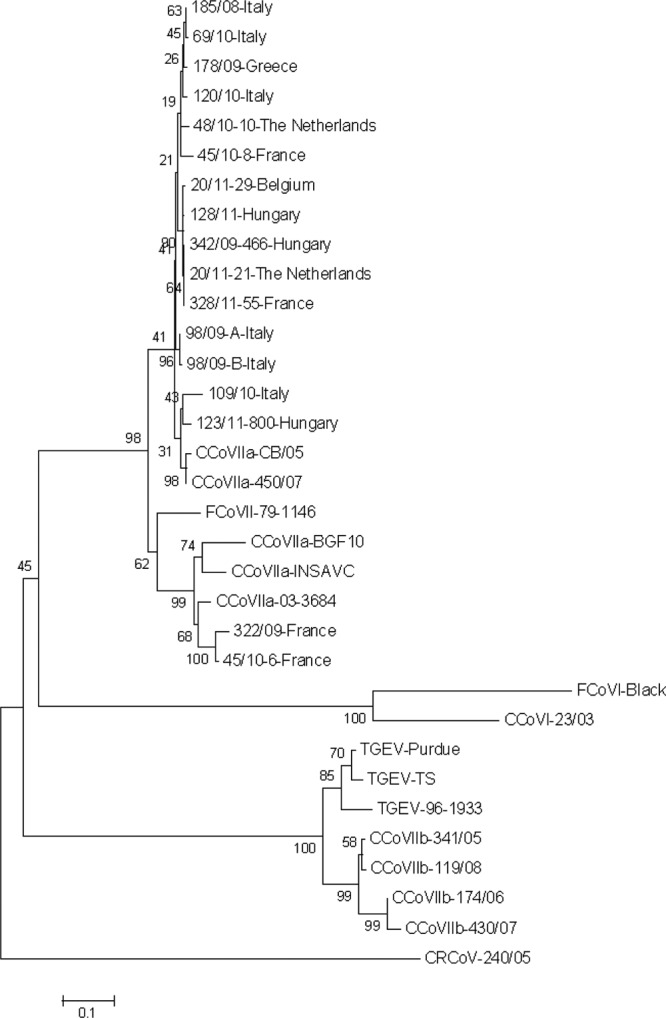
Phylogenetic analysis of pantropic CCoV. Neighbor-joining tree based on the partial 5′ end of the spike protein gene of Alphacoronavirus 1. For phylogenetic tree construction, the following coronavirus reference strains were used (GenBank accession numbers are reported in parentheses): CCoV-IIa pantropic strains CB/05 (DQ112226) and 450/07 (GU146061); CCoV-IIa enteric strains Insavc-1 (D13096), BGF10 (AY342160), and 03-3684 (FJ009113); CCoV-IIb 341/05 (EU856361), 174/06 (EU856362), 430/07 (EU924790), and 118/08 (EU924791); CCoV-I 23/03 (AY307021); feline coronavirus (FCoV) type I Black (EU186072); FCoV-II 79-1146 (NC_007025); and porcine transmissible gastroenteritis virus (TGEV) Purdue (NC_002306), TS (DQ201447), and 96-1933 (AF104420). The Betacoronavirus canine respiratory coronavirus (CRCoV) 240/05 (EU999954) was used as the outgroup. Statistical support was provided by bootstrapping over 1,000 replicates. The scale bars indicate the estimated numbers of nucleotide substitutions per site.
Detection of other pathogens in dogs infected by pantropic CCoV.
Most dogs that were infected by CCoV-IIa strains in their internal organs were found to be positive for CPV-2 (data not shown). An Italian dog was coinfected by CPV-2c and CRCoV. Only four dogs (one French, one Belgian, one Hungarian, one Italian) were shown to have single infections by pantropic CCoVs.
DISCUSSION
Virulent CCoV strains have been sporadically reported in the last few decades (3, 4), but only in 2006, an outbreak of systemic, fatal infection caused by a pantropic CCoV strain was described (9). Further pantropic CCoV outbreaks were later reported in France and Belgium (13), Greece (14), and Italy (15).
The surveillance activity reported in the present study led to the detection of potential pantropic strains in 24 out of 354 dog carcasses analyzed. However, the identification of CPV-2 (and in one case also of CRCoV) in 20 of these dogs prevents any definitive association between pantropic CCoV and the death of the infected animals from being established. Nevertheless, the most recent studies have elucidated that usually pantropic CCoV does not cause the death of the infected dogs but is more frequently associated with subclinical infections, invariably causing severe leukopenia and decreases of all lymphocytic subsets, mainly CD4+ T cells (12). The induced leukopenia may be responsible for the more severe clinical course of concurrent infections. Therefore, a pantropic CCoV infection is likely to be undiagnosed when other (more common) pathogens are simultaneously present. Unfortunately, blood was not available from the dogs infected by pantropic CCoV during our survey, hence precluding the assessment of leukopenia.
The molecular investigations demonstrated a close genetic relatedness among the majority of detected pantropic strains but failed to identify common genetic signatures. None of the strains displayed deletions in the ORF3abc region, whereas the mutation D125N, which was detected in the S-protein gene of the first two Italian viruses, strains CB/05 and 450/07, and tentatively associated with strain virulence (15, 33), was shared by 9/17 pantropic CCoVs subjected to sequence analysis. Interestingly, two French viruses were more closely related to enteric CCoVs than to other pantropic viruses. These viruses were recovered from dogs housed in the same breeding kennel that displayed diarrhea, vomiting, and neurological disorders (one dog), and both dogs were coinfected with CPV-2b. Considering that the viral RNA was isolated from the spleen (the only tissue with discrete viral titers), these viruses could be true enteric strains that had spread from the intestine to the spleen through the lymphohematogenic route. Indeed, enteric CCoV strains with high virulence were previously described, and coinfection with parvovirus may have exacerbated the clinical symptoms, leading to the death of the animals (3, 4).
Currently, there are no diagnostic tests able to differentiate the pantropic from the enteric strains since they are strictly related at the genetic and antigenic level and no specific marker of pathogenicity has been detected so far. Thus, dogs are operationally defined as being infected by pantropic CCoV through detection of a CCoV-IIa strain in extraintestinal tissues. Even considering the poor homogeneity of the sampling (not all tissues of all carcasses were available for the analyses), it is noteworthy that in most positive tissues, only low viral titers were detected by real-time RT-PCR. This finding could be due to a long-term CCoV infection with a normal reduction of viral titers and exacerbation of the clinical course by infections with other pathogens as a consequence of profound immunosuppression.
Another striking finding is the lack of isolation on cell cultures of the detected strains, including those having high viral titers. A possible explanation is that in most cases clinical samples and/or carcasses had not been properly stored before the virus isolation attempts.
In conclusion, practitioners and scientists should be adequately alerted about the circulation of pantropic CCoV strains, which should be considered in the diagnostic algorithm of systemic infections. Adequate information will help improve surveillance in kennels and rescue shelters, thus expanding the knowledge about the epidemiology and pathogenic potential of pantropic CCoV.
Supplementary Material
ACKNOWLEDGMENT
This work was supported by grants from Pfizer Animal Health (Kalamazoo, MI).
Footnotes
Published ahead of print 24 October 2012
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JCM.02466-12.
REFERENCES
- 1. Carstens EB. 2010. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2009). Arch. Virol. 155: 133–146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Adams MJ, Carstens EB. 2012. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2012). Arch. Virol. 157: 1411–1422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Decaro N, Buonavoglia C. 2008. An update on canine coronaviruses: viral evolution and pathobiology. Vet. Microbiol. 132: 221–234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Decaro N, Buonavoglia C. 2011. Canine coronavirus: not only an enteric pathogen. Vet. Clin. North Am. Small Anim. Pract. 41: 1121–1132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Pratelli A, Decaro N, Tinelli A, Martella V, Elia G, Tempesta M, Cirone F, Buonavoglia C. 2004. Two genotypes of canine coronavirus simultaneously detected in fecal samples of dogs with diarrhea. J. Clin. Microbiol. 42: 1797–1799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Pratelli A, Martella V, Decaro N, Tinelli A, Camero M, Cirone F, Elia G, Cavalli A, Corrente M, Greco G, Buonavoglia D, Gentile M, Tempesta M, Buonavoglia C. 2003. Genetic diversity of a canine coronavirus detected in pups with diarrhoea in Italy. J. Virol. Methods 110: 9–17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Decaro N, Mari V, Campolo M, Lorusso A, Camero M, Elia G, Martella V, Cordioli P, Enjuanes L, Buonavoglia C. 2009. Recombinant canine coronaviruses related to transmissible gastroenteritis virus of swine are circulating in dogs. J. Virol. 83: 1532–1537 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Decaro N, Mari V, Elia G, Addie DD, Camero M, Lucente MS, Martella V, Buonavoglia C. 2010. Recombinant canine coronaviruses in dogs, Europe. Emerg. Infect. Dis. 16: 41–47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Buonavoglia C, Decaro N, Martella V, Elia G, Campolo M, Desario C, Castagnaro M, Tempesta M. 2006. Canine coronavirus highly pathogenic for dogs. Emerg. Infect. Dis. 12: 492–494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Decaro N, Campolo M, Lorusso A, Desario C, Mari V, Colaianni ML, Elia G, Martella V, Buonavoglia C. 2008. Experimental infection of dogs with a novel strain of canine coronavirus causing systemic disease and lymphopenia. Vet. Microbiol. 128: 253–560 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Decaro N, Elia G, Martella V, Campolo M, Mari V, Desario C, Lucente MS, Lorusso E, Kanellos T, Gibbons RH, Buonavoglia C. 2010. Immunity after natural exposure to enteric canine coronavirus does not provide complete protection against infection with the new pantropic CB/05 strain. Vaccine 28: 724–729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Marinaro M, Mari V, Bellacicco AL, Tarsitano E, Elia G, Losurdo M, Rezza G, Buonavoglia C, Decaro N. 2010. Prolonged depletion of circulating CD4+ T lymphocytes and acute monocytosis after pantropic canine coronavirus infection in dogs. Virus Res. 152: 73–78 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Zicola A, Jolly S, Mathijs E, Ziant D, Decaro N, Mari V, Thiry E. 2012. Fatal outbreaks in dogs associated with pantropic canine coronavirus in France and Belgium. J. Small Anim. Pract. 53: 297–300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Ntafis V, Xylouri E, Mari V, Papanastassopoulou M, Papaioannou N, Thomas A, Buonavoglia C, Decaro N. 2012. Molecular characterization of a canine coronavirus NA/09 strain detected in a dog's organs. Arch. Virol. 157: 171–175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Decaro N, Mari V, von Reitzenstein M, Lucente MS, Cirone F, Elia G, Martella V, King VL, Di Bello A, Varello K, Zhang S, Caramelli M, Buonavoglia C. 2012. A pantropic canine coronavirus genetically related to the prototype isolate CB/05. Vet. Microbiol. 159: 239–244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Decaro N, Pratelli A, Campolo M, Elia G, Martella V, Tempesta M, Buonavoglia C. 2004. Quantitation of canine coronavirus RNA in the faeces of dogs by TaqMan RT-PCR. J. Virol. Methods 119: 145–150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Decaro N, Martella V, Ricci D, Elia G, Desario C, Campolo M, Cavaliere N, Di Trani L, Tempesta M, Buonavoglia C. 2005. Genotype-specific fluorogenic RT-PCR assays for the detection and quantitation of canine coronavirus type I and type II RNA in faecal samples of dogs. J. Virol. Methods 130: 72–78 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. (Oxf.). 41: 95–98 [Google Scholar]
- 19. Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24: 1596–1599 [DOI] [PubMed] [Google Scholar]
- 20. Decaro N, Desario C, Elia G, Mari V, Lucente MS, Cordioli P, Colaianni ML, Martella V, Buonavoglia C. 2007. Serological and molecular evidence that canine respiratory coronavirus is circulating in Italy. Vet. Microbiol. 121: 225–230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Lorusso A, Desario C, Mari V, Campolo M, Lorusso E, Elia G, Martella V, Buonavoglia C, Decaro N. 2009. Molecular characterization of a canine respiratory coronavirus strain detected in Italy. Virus Res. 141: 96–100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Leary TP, Erker JC, Chalmers ML, Cruz AT, Wetzel JD, Desai SM, Mushahwar IK, Dermody TS. 2002. Detection of mammalian reovirus RNA by using reverse transcription-PCR: sequence diversity within the λ3-encoding L1 gene. J. Clin. Microbiol. 40: 1368–1375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Gouvea V, Santos N, Timenetsky Mdo C. 1994. Identification of bovine and porcine rotavirus G types by PCR. J. Clin. Microbiol. 32: 1338–1340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Jiang X, Huang PW, Zhong WM, Farkas T, Cubitt DW, Matson DO. 1999. Design and evaluation of a primer pair that detects both Norwalk- and Sapporo-like caliciviruses by RT-PCR. J. Virol. Methods 83: 145–154 [DOI] [PubMed] [Google Scholar]
- 25. Decaro N, Elia G, Martella V, Desario C, Campolo M, Trani LD, Tarsitano E, Tempesta M, Buonavoglia C. 2005. A real-time PCR assay for rapid detection and quantitation of canine parvovirus type 2 DNA in the feces of dogs. Vet. Microbiol. 105: 19–28 [DOI] [PubMed] [Google Scholar]
- 26. Decaro N, Elia G, Campolo M, Desario C, Lucente MS, Bellacicco AL, Buonavoglia C. 2005. New approaches for the molecular characterization of canine parvovirus type 2 strains. J. Vet. Med. B Infect. Dis. Vet. Public Health 52: 316–319 [DOI] [PubMed] [Google Scholar]
- 27. Hu RL, Huang G, Qiu W, Zhong ZH, Xia XZ, Yin Z. 2001. Detection and differentiation of CAV-1 and CAV-2 by polymerase chain reaction. Vet. Res. Commun. 25: 77–84 [DOI] [PubMed] [Google Scholar]
- 28. Elia G, Decaro N, Martella V, Cirone F, Lucente MS, Lorusso E, Di Trani L, Buonavoglia C. 2006. Detection of canine distemper virus in dogs by real-time RT-PCR. J. Virol. Methods 136: 171–176 [DOI] [PubMed] [Google Scholar]
- 29. Decaro N, Altamura M, Pratelli A, Pepe M, Tinelli A, Casale D, Martella V, Tafaro A, Camero M, Elia G, Tempesta M, Jirillo E, Buonavoglia C. 2002. Evaluation of the innate immune response in pups during canine parvovirus type 1 infection. New Microbiol. 25: 291–298 [PubMed] [Google Scholar]
- 30. Decaro N, Amorisco F, Desario C, Lorusso E, Camero M, Bellacicco AL, Sciarretta R, Lucente MS, Martella V, Buonavoglia C. 2010. Development and validation of a real-time PCR assay for specific and sensitive detection of canid herpesvirus 1. J. Virol. Methods 169: 176–180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Decaro N, Elia G, Campolo M, Desario C, Mari V, Radogna A, Colaianni ML, Cirone F, Tempesta M, Buonavoglia C. 2008. Detection of bovine coronavirus using a TaqMan-based real-time RT-PCR assay. J. Virol. Methods 151: 167–171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Erles K, Brownlie J. 2009. Sequence analysis of divergent canine coronavirus strains present in a UK dog population. Virus Res. 141: 21–25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Decaro N, Martella V, Elia G, Campolo M, Desario C, Cirone F, Tempesta M, Buonavoglia C. 2007. Molecular characterisation of the virulent canine coronavirus CB/05 strain. Virus Res. 125: 54–60 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.