

Abstract
We analyzed published downcore microfossil records from 150 studies and reinterpreted them from an ecological degradation perspective to address the following critical but still imperfectly answered questions: (1) How is the timing of human-induced degradation of marine ecosystems different among regions? (2) What are the dominant causes of human-induced marine ecological degradation? (3) How can we better document natural variability and thereby avoid the problem of shifting baselines of comparison as degradation progresses over time? The results indicated that: (1) ecological degradation in marine systems began significantly earlier in Europe and North America (∼1800s) compared with Asia (post-1900) due to earlier industrialization in European and North American countries, (2) ecological degradation accelerated globally in the late 20th century due to post-World War II economic growth, (3) recovery from the degraded state in late 20th century following various restoration efforts and environmental regulations occurred only in limited localities. Although complex in detail, typical signs of ecological degradation were diversity decline, dramatic changes in total abundance, decrease in benthic and/or sensitive species, and increase in planktic, resistant, toxic, and/or introduced species. The predominant cause of degradation detected in these microfossil records was nutrient enrichment and the resulting symptoms of eutrophication, including hypoxia. Other causes also played considerable roles in some areas, including severe metal pollution around mining sites, water acidification by acidic wastewater, and salinity changes from construction of causeways, dikes, and channels, deforestation, and land clearance. Microfossils enable reconstruction of the ecological history of the past 102–103 years or even more, and, in conjunction with statistical modeling approaches using independent proxy records of climate and human-induced environmental changes, future research will enable workers to better address Shifting Baseline Syndrome and separate anthropogenic impacts from background natural variability.
Keywords: Eutrophication, hypoxia, marine ecosystems, microfossils, pollution, species diversity
Introduction
What are the dominant causes and mechanisms of the deterioration of marine organisms, communities, and ecosystems by anthropogenic activities (hereafter marine ecological degradation)? How is timing of marine ecological degradation different among regions and how can we understand changes in such systems in the face of shifting understanding of natural states and variability (Pauly 1995; Knowlton and Jackson 2008)? These critical questions are still answered imperfectly. Marine ecological degradation is a serious concern both because of the changes in marine systems themselves, and because of potential consequences for human populations as ecosystem services historically provided by marine systems are lost (Jackson et al. 2001; Worm et al. 2006; Palumbi et al. 2009). Numerous lines of evidence have shown that marine ecological degradations, for example, biodiversity loss, population collapse, invasion of exotic species, and various faunal/floral changes, are now apparent in most, if not all, marine ecosystems (Lotze et al. 2006; Halpern et al. 2008; Jackson 2008). Causes include increased nutrient loads resulting in zones of low dissolved oxygen and other symptoms of eutrophication (Kemp et al. 2005; Diaz and Rosenberg 2008; Breitburg et al. 2009), fisheries exploitation (Jackson 2001; Jackson et al. 2001; Worm et al. 2006), physical habitat destruction (Jackson 2001; Jackson et al. 2001; Coleman and Williams 2002), ocean acidification (Orr et al. 2005; Hoegh-Guldberg et al. 2007; Fabry et al. 2008), pollution (Shahidul Islam and Tanaka 2004), land clearance/modification (Lotze et al. 2005; Airoldi and Beck 2007; Willard and Cronin 2007), and global warming (Hoegh-Guldberg and Bruno 2010).
Meta-analyses have detailed the general historical process of marine ecological degradation for the last 3000 years using fisheries, ecological, historical, archeological, and paleontological records, and have revealed rapid biodiversity and population loss starting at ∼1800, the onset of industrialization (Lotze et al. 2006; Worm et al. 2006). A similar approach was also used for coral reef ecosystems, suggesting even earlier timing of the inception of ecological degradation (Pandolfi et al. 2003). From these studies, we have a general idea of the current status and the historical process of marine ecological degradation. However, higher temporal resolution analyses of the long-term history of marine ecological degradation are scarce for the past few centuries and millennia, mainly because of the difficulty of obtaining long-term, continuous ecological records. This difficulty is due to a number of factors: (1) biological monitoring usually covers no more than the past 20 years (Borja et al. 2010), which is too short a time period to understand the long-term history of human-caused ecological degradation; (2) fisheries harvest records covering hundreds of years (Lotze and Milewski 2004) are available for only a few commercial fishes and may be artificially and politically biased; and (3) historical and archeological records are often exceedingly fragmentary in time and space. Furthermore, data are usually available only from waters of the coasts of North America, Europe, Australia, and New Zealand (e.g., Worm et al. 2006, 2009).
The fossil record has the potential to provide some essential information that is absent in these other records (Jablonski et al. 2003, 2006). Kidwell has recently shown the utility of comparing live and dead (sub-fossil) molluscan assemblages to detect changes in relative abundance attributable to environmental degradation (Kidwell 2007). Several researchers have also emphasized the importance of a paleoecological approach (Gorham et al. 2001) and integrated it with fisheries, ecological, historical, and archeological records (Jackson et al. 2001; Worm et al. 2006). However, the geological records of most fossil groups lack sufficient time resolution to elucidate a detailed history of marine ecological degradation during past centuries.
Marine microfossils include protists (foraminifera and radiolaria), microscopic algae (diatoms, dinoflagellates, and coccolithophores), and crustaceans (ostracods). These organisms are abundantly and continuously preserved as fossils in cores with potentially continuous sedimentary records, and so provide excellent temporal resolution and long-term ecological data for the past hundreds to thousands of years. Various radiometric dating methods can provide robust chronologies for sediment cores. Furthermore, each group of microfossils is composed of many species with various habitat and ecological preferences and well-known taxonomy. Thus, they are useful proxies or model organisms to infer states of whole marine ecosystems in the past (Thomas and Gooday 1996; Cronin and Raymo 1997; Yasuhara and Cronin 2008; Yasuhara et al. 2008). Although several researchers have shown the importance of microfossils in understanding marine ecological degradation (Tikkanen et al. 1997; Willard and Cronin 2007; Yasuhara et al. 2007; Tsujimoto et al. 2008), many studies focused on microfossil-based reconstruction of pollution history (Sen Gupta et al. 1996; Clarke et al. 2006; Gooday et al. 2009) rather than reconstruction of the biotic response to pollution. This situation is similar to that in paleolimnology. As Smol (2009) pointed out, paleolimnological data can typically be used in two important ways: (1) to use the information preserved in lake sediments to reconstruct past population of specific organisms or groups of organisms that comprise important components of aquatic ecosystems; (2) to use it to reconstruct past environmental conditions such as pH and oxygen contents. Most paleolimnological studies are typically the latter (Smol 2009). The same is true for marine paleoecology. In the present paper, we focus on the former perspective – biotic response, not paleoenvironmental reconstruction – and use the word “paleoecology” only for studies that use the (microfossil) information preserved in marine sediments to reconstruct past population of specific organisms or groups of organisms that comprise important components of marine ecosystems (cf. Smol 2009).
Microfossils can be useful to sort out the relative importance of climate changes and human-induced environmental changes. Periods of ecological degradation overlap with several serious climatic events including the Little Ice Age. With sufficiently detailed records, it may be possible to evaluate the relative importance of climate and human-induced environmental changes on marine ecosystems, including the degree to which human-induced post-20th century global warming has had a significant effect on marine ecosystems.
Numerous lines of evidences suggest regional differences in the timing of the inception of marine ecological degradation. For example, when one compares a review paper based on Asian records (Pauly and Thia-Eng 1988) with those based on North American and European records (Jackson et al. 2001; Worm et al. 2006), a clear difference in timing is apparent, with the degradation of Asian ecosystems occurring later on average. Furthermore, most human-induced environmental problems first appeared in Europe before spreading to other regions (Huettmann 2012), but these qualitative patterns are yet to be tested statistically on a global scale. Previous case studies are largely restricted to developed countries because long-term monitoring and robust fishery, historical, and archeological data are usually available only from North America, Europe, and Australia. Microfossil records can help to broaden this coverage, however, because one well-chosen sediment core allows for the reconstruction of hundred years of ecosystem history, even in the complete absence of local biological monitoring.
Here, we review and analyze published microfossil records from 150 studies and synthesize them in order to understand the spatial and temporal context of ecological degradation. Our objective is to use microfossil records to answer critical questions about marine degradation listed above, and to:
examine long-term global trends of marine ecological degradation, focusing on evidence on the timing of inception (i.e., start of degradation), acceleration (i.e., increasing severity of degradation), and ecosystem recovery;
provide an overview of the nature of marine ecological degradation evident in microfossil assemblages;
review regional trends in marine ecological degradation; and
discuss problems, solutions, and future outlook of micropaleontological analyses to investigate marine ecological degradation.
Methods
We compiled marine (in a broad sense, including deep sea, pelagic, shallow marine, estuarine, and brackish-water lake) microfossil records in dated sediment cores (hereafter downcore microfossil records) that preserved evidence of ecological degradation (Figs. 15; Online Table S1, Fig. S1, and Supplementary Text S1; see “Nature of Marine Ecological Degradation: Paleoecological Information from Microfossils” section below). These records were mainly composed of data on foraminifera, diatoms, dinoflagellate cysts, and ostracods (Figs S2–3; Table S1). Diatoms and dinoflagellates include both planktic and benthic species. Although foraminifera and ostracods also have both planktic and benthic species, with the exception of one planktic foraminiferal study on global warming (see Table S1 and Global warming section), our database includes only benthic taxa. Planktic foraminifera are rare in shallow, coastal, and/or brackish-water environments where marine ecological degradations are prominent. Planktic ostracods are rarely preserved as fossils because of their weakly calcified carapaces (Schellenberg 2007).
Figure 1.
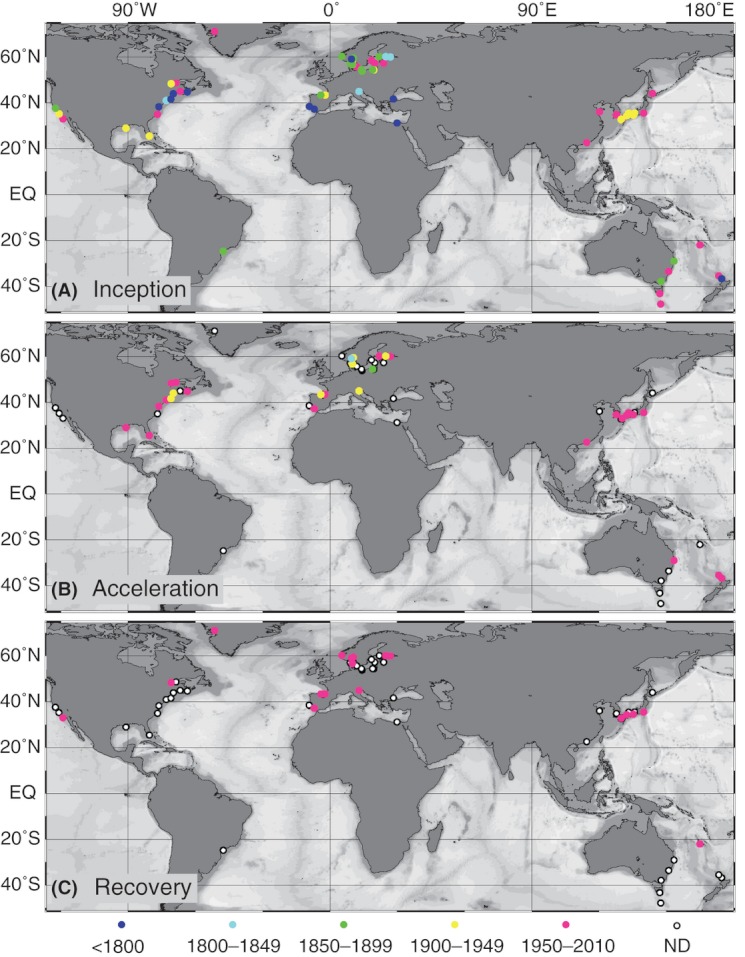
Global distribution of marine ecological degradations in downcore microfossil records. Complete data are found in Table S1. Distribution of (A) inception, (B) acceleration, and (C) recovery ages [five categories of <1800 (blue), 1800–1849 (light blue), 1850–1899 (green), 1900–1949 (yellow), and 1950–2010 (pink)] of marine ecological degradation detected in microfossil records. ND (open circle): not detected.
Figure 5.
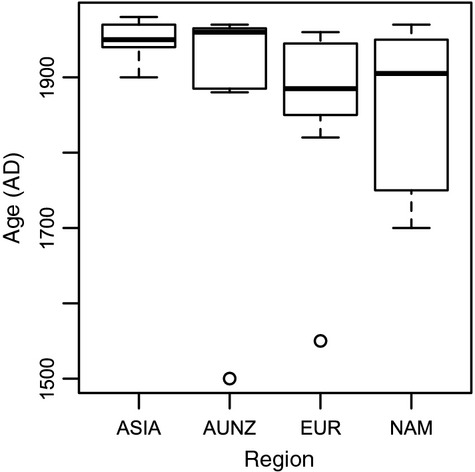
Boxplots showing regional differences of the inception dates of marine ecological degradation. ASIA: Asia; AUNZ: Australia and New Zealand; EUR: Europe; NAM: North America. Asian inception dates are significantly later than European (W = 261, P = 0.0008) and North American (W = 144, P = 0.0105) inception dates (but not significantly different from AUNZ inception dates: W = 36, P = 0.47). Only inception dates >1000 AD were used.
Most studies compiled here were conducted during or after the late 1990s (Fig. S1). A few pioneering studies go back to 1960–1970s, but their chronologies are often less reliable (Risdal 1963; Schafer 1970) because radiometric dating was not as widely available at that time, and thus are not included here. Microfossil records included in our analyses cover up to the last ∼10,000 years, but most of them cover only the last 200–300 years (Fig. 2). To evaluate regional differences in the timing of marine ecological degradation, we compared European, North American, Australia/New Zealand, and Asian records using Mann–Whitney tests. Datasets used for this paper are deposited at Dryad (http://dx.doi.org/10.5061/dryad.2t604).
Figure 2.
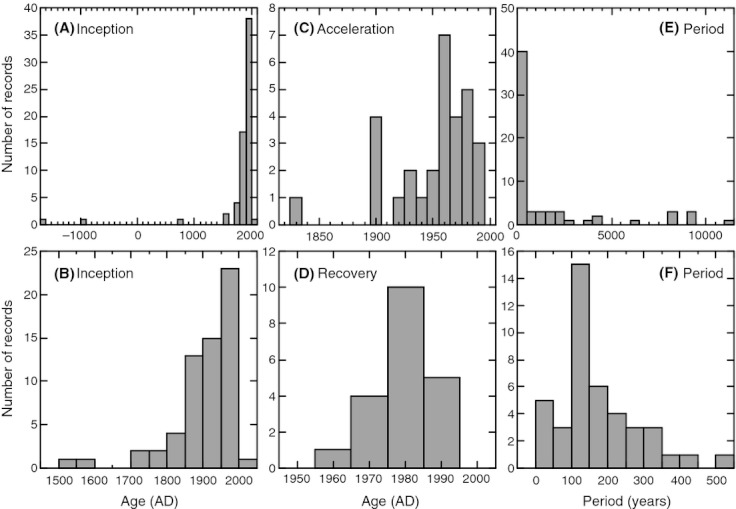
Age distribution of microfossil-based marine ecological degradation records. Histograms showing distribution of (A; B for closeup) inception, (C) acceleration, and (D) recovery ages of marine ecological degradations and of (E; F for closeup) covering period of microfossil records. Data from Table S1.
Global trends and regional differences of marine ecological degradation in microfossil records
Distribution of microfossil records
In this review of microfossil records of ecological degradation, we focus on the timing of three events: the inception (i.e., start) of degradation, its acceleration (i.e., increasing severity of degradation), and when observed, recovery from a degraded state. Ideally, we would establish statistical means to define these events and apply them to the compiled case studies, but this approach is not feasible here because the raw data are not available for the vast majority of these studies. Accordingly, we take the more practical route of identifying these events through visual interpretation of published figures, with the identifications all performed by the lead author in order to maximize consistency. Care was taken in identifying inception of degradation as when biotic changes exceeded natural, background fluctuations. Ages of recovery were interpreted as the initiation of changes in community parameters returning toward prehistorical or pre-impacted states. Although this approach is necessarily somewhat subjective, it allows for comprehensive use of published information.
Evidence for marine ecological degradation in microfossil records was mostly found in shallow, coastal environments (Fig. 1; Table S1). Signs of ecological degradation were concentrated in North America, Europe, and East Asia (Fig. 1; Table S1) because strong human footprints, including hypoxic dead zones, are concentrated there (Diaz and Rosenberg 2008; Halpern et al. 2008), and because there are few studies in developing countries in Africa, Central and South America, and Southeast and West Asia. The temporal patterns of inception and acceleration of degradation and ecosystem recovery are illustrated in Figures 1–5 to characterize global trends.
Inception of degradation
These data reveal that marine ecological degradation started in the 1800s in many regions of European and North American countries (Figs. 1–3), a pattern consistent with temporal patterns in industrialization and land clearing for agriculture in the 19th century, and with previous meta-analysis of fisheries, ecological, historical, archeological, and macrofossil data (Worm et al. 2006). The earliest degradation record extends back to ∼1500 BC (Figs. 1–3), however, and was caused by mining activity in the Tinto Estuary, Spain (Ruiz et al. 2009).
Figure 3.
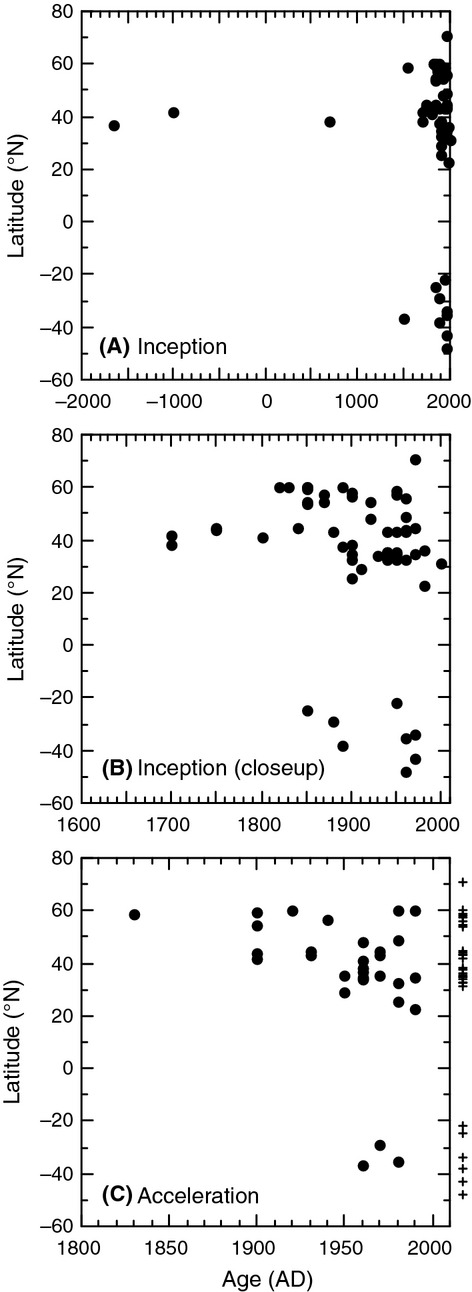
Latitudinal distribution of microfossil-based marine ecological degradation records. Latitude versus inception (A; B for closeup) and acceleration ages (C) of marine ecological degradation. Cross: locations lacking detectable acceleration. Data from Table S1.
Northern hemisphere data clearly show an earlier start of marine ecological degradation in higher latitudes where developed countries are concentrated (Figs. 3–4). The start age of marine ecological degradation shows a noisy but significant relationship with latitude (R2 = 0.07362, P = 0.0494; Fig. 4). In addition, onsets of degradation occurred significantly later in Asia compared with Europe (W = 261, P = 0.0008) and North America (W = 144, P = 0.0105) (Fig. 5: only data for the past 1000 years are used for Mann–Whitney test). In the Black Sea, downcore evidence of introduced species by exploration or merchant ships extends back to two or three thousand years ago (Marret et al. 2009). In Europe, eutrophication footprints on faunal/floral composition extend back to ∼1550 in a Norwegian fjord (Alve 2000) and to the mid-19th century in the Baltic Sea (Andrén 1999; Andrén et al. 1999). Initial ecological degradation in the 1700s in Chesapeake Bay in North America is indicated by disturbances to diatom flora and submerged aquatic vegetation associated with European land clearance (Brush and Hilgartner 2000; Willard and Cronin 2007; Brush 2009). In contrast, no marine ecological degradation was detected from pre-1900 sediments in Asia. Instead, microfossil evidence indicated that degradation in Asia started rapidly and became widespread in the early to mid-20th century (Fig. 1), most likely related to Asian industrialization (as early as ∼1900, about 100 years later than European and North American industrialization) and post-World War II economic growth. Typical examples are found in Japanese embayments (Yasuhara et al. 2007; Katsuki et al. 2008; Tsujimoto et al. 2008). It is plausible that there were earlier instances of marine ecological degradation in Asia because large-scale land-use changes are known in Asia even back to the Neolithic period (Zong et al. 2007) and many Asian countries have long histories of well-developed cities. However, industrialization has had the dominant effect on sedimentary records reviewed here, although future work on core records that are both highly resolved and of extended duration will be needed to investigate if human-induced marine ecological degradation is detectable in pre-industrial Asia. Nevertheless, by the mid- to late-20th century, microfossil evidence of marine ecological degradation is globally widespread, occurring even in polar Greenland (Elberling et al. 2003) and a remote Pacific island (Debenay and Fernandez 2009) (Figs. 1 and 3).
Figure 4.
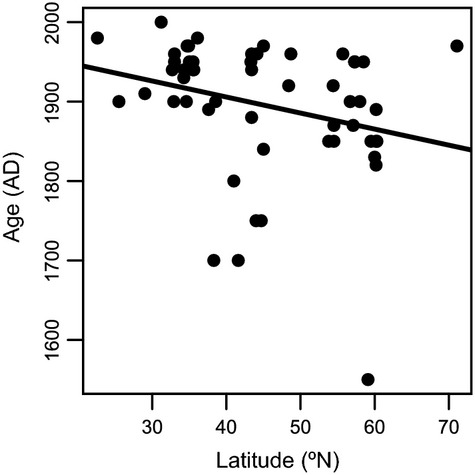
Relationship between latitude and the inception dates of marine ecological degradation in the northern hemisphere (R2 = 0.07362, P = 0.0494). Only inception dates >1000 AD were used.
Acceleration of degradation
In the mid- to late-20th century, marine ecological degradation accelerated in many areas (Figs. 1–3) in concert with rapid post-World War II economic and population growth, and increased fertilizer use. The results show that the earlier acceleration occurred in higher latitudes, especially in the northern hemisphere (Fig. 3) where developed countries and cities are concentrated (see Overview of Regional Trends section).
Ecosystem recovery
Alve (1995) presented a conceptual diagram that illustrates recovery from the degraded state following a similar (but reverse) trajectory as during the degradation process. However, the analysis of monitoring data in eutrophic systems by Duarte et al. (2009) indicates that degradation and recovery trajectories rarely coincide. Microfossil evidence indicates that relatively few sites show recovery from the degraded state to the predegradation state (e.g., abundance, diversity, and faunal or floral composition), and those recoveries that occurred were always during the late 20th century (Figs. 1 and 2) after various restoration efforts and environmental regulations were initiated. Furthermore, microfossil evidence indicates that complete recovery is very rare [see European and African waters section below for the Töölönlahti Bay study (Tikkanen et al. 1997) as one of the best examples of the ecosystem recovery]; instead post-recovery faunal or floral composition is usually different from predegradation faunal composition to varying degrees (McGann et al. 2003; Weckström 2006; Tsujimoto et al. 2008). Differences between degradation and recovery trajectories may occur because remediation of estuaries is usually slower than degradation (Hawkins et al. 2002), and because opportunistic species become the pioneer recolonizers under the post-degradation condition (Alve 1995). It is known that history of colonization (sequence of species arrival) influences community structure (Fukami and Morin 2003), and thus, “recovered” community is not necessarily same as predegradation community.
Causes of marine ecological degradation
The predominant cause of degradation detected in microfossil records was nutrient enrichment and the resulting symptoms of eutrophication including hypoxia (Fig. S4). Other causes also played considerable roles in some areas, including severe metal pollution around mining sites, acidification by acidic wastewater, and salinity changes from construction of causeways, dikes, and channels, deforestation, and land clearance (Table S1). These environmental causes of marine ecological degradation are likely more prominent in microfossil records than reported in other recent reviews on ecological degradation (Jackson et al. 2001; Jackson 2008), because other reviews have mainly focused on relatively large animals in which fisheries exploitation overwhelms other causes.
Nature of marine ecological degradation: paleoecological information from microfossils
Paleoecological signals (i.e., changes in microfossil assemblages) of marine ecological degradation can vary among systems, regions, and taxa (Table S1). There are, however, typical responses to particular stressors common among different locations in downcore microfossil records. Among these responses are changes in species diversity and total abundance, changes in abundance of both sensitive and tolerant species, increases in toxic algal bloom species, and the appearance or increase in invasive species.
Species diversity
A decline in species diversity is one of the typical signs of ecological degradation that can diminish ecosystem functioning (Cardinale et al. 2002; Worm et al. 2006; Danovaro et al. 2008). Species diversity is usually measured by the Shannon Index (H), species richness, or as the expected number of species in samples rarefied to n individuals E(Sn). These measures of diversity decrease in most of the microfossil records from systems with strong human influence on water clarity, nutrient loads and oxygen concentrations. For example, foraminiferal diversity decreased in Osaka Bay, foraminiferal and ostracod diversity decreased in the Gulf of Mexico, and diatom diversity decreased in Chesapeake Bay (Fig. 6) after the onset of intense human alteration of the environment. These diversity decreases most likely result from increasing prey production and resulting dominance of a few opportunistic species, local extinctions due to decreasing bottom-water oxygen content, and reduced light penetration to the benthos (Rabalais et al. 2007; Tsujimoto et al. 2008; Brush 2009). In the Gulf of Mexico, both foraminifera and ostracod diversities showed significant, negative relationships with increasing US fertilizer use, and in Osaka Bay, foraminiferal diversity showed a significant, negative relationship with COD (chemical oxygen demand) discharge (Fig. 7). Although a direct oxygen paleo-proxy (i.e., geochemical evidence for low-oxygen content) is rarely available for comparison, these indirect evidences strongly support the above-mentioned hypothesis.
Figure 6.
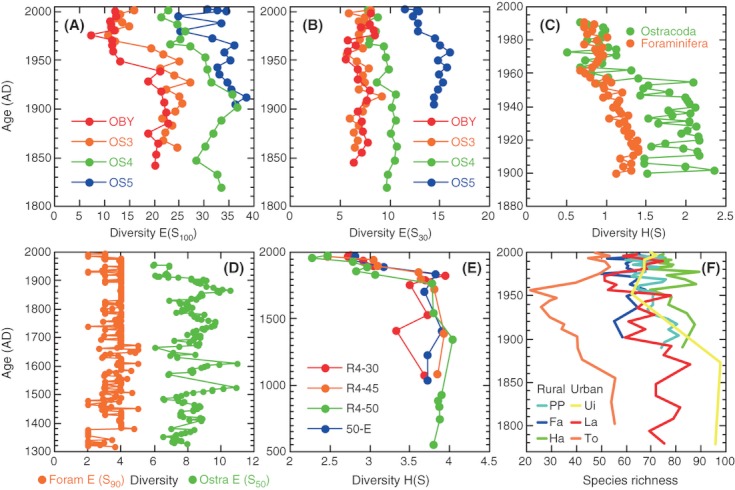
Long-term downcore trends of microfossil species diversity in representative regions from which high-resolution records are available. (A) Osaka Bay foraminiferal [data from Tsujimoto et al. (2008); OBY, OS3, OS4, and OS5: sediment core sites; cores OBY and OS3 were taken from inner part and cores OS4 and OS5 were taken from middle part of the bay], (B) Osaka Bay ostracod [data from Yasuhara et al. (2007)], (C) Gulf of Mexico foraminiferal (Blackwelder et al. 1996) and ostracod (Alvarez Zarikian et al. 2000), (D) Chesapeake Bay foraminiferal [data from Karlsen et al. (2000)] and ostracod [data from Cronin and Vann (2003)], (E) Chesapeake Bay diatom (Cooper 1995) (R4-30, R4-45, R4-50, and 50-E: sediment core sites), and (F) Baltic Sea diatom (Weckström et al. 2007) (PP, Fa, and Sa: rural sites; Ui, La, and To: urban sites) records. Species diversity shown by species richness, Shannon Index H(S), or the expected number of species in samples rarefied to n individuals E(Sn) depending on availability.
Figure 7.
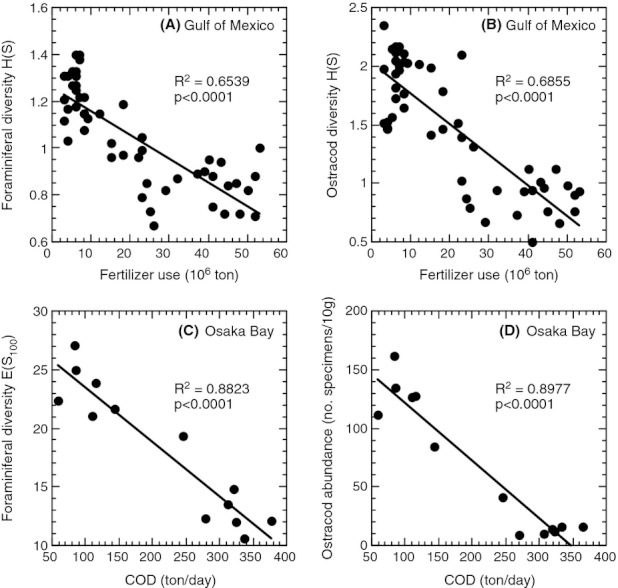
Relationship between eutrophication and species diversity and abundance. Correlations between (A) foraminiferal diversity H(S) in Gulf of Mexico core BC-10 and fertilizer use in USA, (B) ostracod diversity H(S) in Gulf of Mexico core BC-10 and fertilizer use in USA, (C) foraminiferal diversity E(S100) in Osaka Bay core OS3 and discharges of COD (chemical oxygen demand) from Osaka Prefecture, and (D) ostracod abundance (number of specimens per 10 g dry sediment) in Osaka Bay core OS3 and discharges of COD from Osaka Prefecture. Gulf of Mexico data from Alvarez Zarikian et al. (2000); Osaka Bay data from Yasuhara et al. (2007) and Tsujimoto et al. (2008).
There are also some exceptions in which species diversity does not show a decreasing trend. In Osaka Bay, ostracod diversity is almost unchanged, especially in the inner part of the bay (i.e., cores OBY and OS3: Fig. 6), even though abundance and faunal composition show dramatic changes caused by eutrophication and hypoxia (Yasuhara and Yamazaki 2005; Yasuhara et al. 2007). This stability of ostracod diversity may reflect a low number of local extinctions, but the reasons behind the persistence of these species are uncertain. There is also no significant change in diversity observed in the foraminifera and ostracod records of the Chesapeake Bay (Fig. 6), although the low diversity throughout the core is likely related to this brackish environment that is naturally stressful for marine benthic organisms due to low and fluctuating salinity and other environmental factors.
Recovery of species diversity by various restoration efforts and environmental regulations is also observed in microfossil records, but only in a few locations (Ellegaard et al. 2006; Weckström et al. 2007; Alve et al. 2009; Fig. 6). Instead, in most locations, diversities in the most recent sections of cores were significantly lower than those in pre-industrial or pre-urbanization core sections: For example, Mann–Whitney test shows that foraminiferal diversity is significantly (P < 0.05) higher in pre-urbanization (<1950) samples than in post-urbanization (>1950) samples in Osaka Bay cores (Fig. 6A), except core OS5 (P = 0.073) that has much smaller sample size and locality far from the source of anthropogenic nutrient input (the mouth of the Yodo River) compared with other cores.
Abundance
Marine ecological degradation is also detected as a decrease or increase in abundance, depending on the taxonomic group, habitat, and physiological tolerances of individual species (Fig. 8). In coastal ecosystems, anthropogenic causes of abundance changes in microfossil assemblages are mainly eutrophication and the resulting bottom-water hypoxia and/or anoxia (e.g., Cooper 1995; Yasuhara et al. 2007; Tsujimoto et al. 2008; Table S1). Plankton usually increases in response to an increasing nutrient and prey supply associated with eutrophication, a few benthic species that are resistant to low-oxygen conditions and low light penetration also increase for the same reason, and most benthic species that cannot tolerate low-oxygen conditions increase in the case of eutrophication without hypoxia or anoxia, but decrease where hypoxia or anoxia result from eutrophication. For example, total diatom and dinoflagellate abundances increase because of a substantial increase in planktic species; foraminiferal abundance increases because of a substantial increase in a few resistant species to hypoxic conditions; and ostracod abundance increases or decreases depending on oxygen concentrations (no ostracod species thrive in hypoxic conditions; the relative abundance of a few relatively resistant species increases but their absolute abundance still decreases). Absolute abundance information was rarely reported in the microfossil records that we compiled here, but the above-mentioned trends seem to be typical (Fig. 8; Table S1). Although quantitative proxies of eutrophication and/or oxygen content are not usually available for direct comparison with microfossil paleoecological data, a significant negative relationship was found between ostracod abundance in a hypoxic site of Osaka Bay and COD discharge from the Osaka Prefecture through the Yodo River to the bay (Fig. 7).
Figure 8.
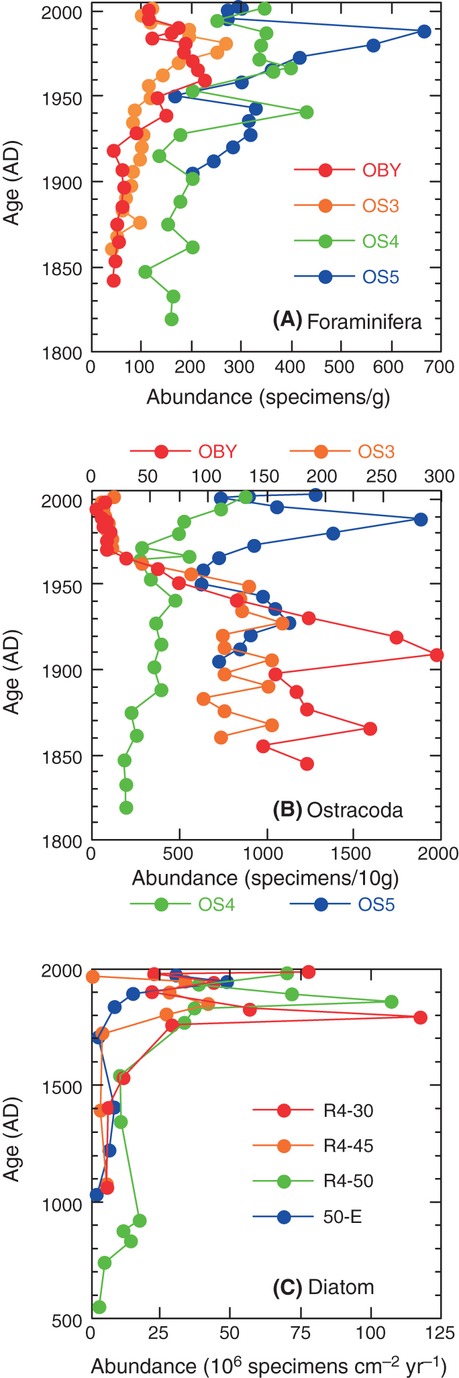
Long-term downcore trends of microfossil abundance in representative regions from which high-resolution records are available. (A) Osaka Bay foraminiferal (Tsujimoto et al. 2008) and (B) Osaka Bay ostracod (Yasuhara et al. 2007) records (OBY, OS3, OS4 and OS5: sediment core sites; cores OBY and OS3 were taken from inner part and cores OS4 and OS5 were taken from middle part of the bay) and (C) Chesapeake Bay diatom records (Cooper 1995) (R4-30, R4-45, R4-50, and 50-E: sediment core sites). In Osaka Bay, ostracode abundance decreases in the inner part of the bay but increases in the central part of the bay, because Osaka Bay hypoxia is restricted in the inner part of the bay (see Overview of Regional Trends section for mechanism of abundance change).
Taxon-specific responses
Benthic foraminifera
In benthic foraminifera, the tolerant genus Ammonia (here meaning tolerant of hypoxia/anoxia or preferring eutrophication and food-rich environment) increases and the sensitive genus Elphidium decreases with increasing eutrophication and resulting hypoxia/anoxia at various locations, including Osaka Bay (Tsujimoto et al. 2006, 2008), Gulf of Mexico (Sen Gupta et al. 1996; Sen Gupta and Platon 2006; Rabalais et al. 2007), Chesapeake Bay (Karlsen et al. 2000), San Francisco Bay (McGann 2008), Long Island Sound (Thomas et al. 2000), and the Bay of Biscay (Irabien et al. 2008). For example, in Osaka Bay, Ammonia starts to increase and Elphidium starts to decrease immediately after the industrial revolution in Japan at ∼1900 (Fig. 9). Agglutinated foraminiferal taxa are also known to be tolerant and increase in various regions (Alve 1996; Hayward et al. 2004; Tsujimoto et al. 2008). The tolerant species Eggerella advena and Trochammina hadai are also widely distributed and dominant in degraded ecosystems (McGann et al. 2003; Katsuki et al. 2008; Tsujimoto et al. 2008). Note that it is difficult to disentable the separate effects of oxygen depletion from other symptoms of eutrophication because we lack finely resolved environmental proxies for each.
Figure 9.
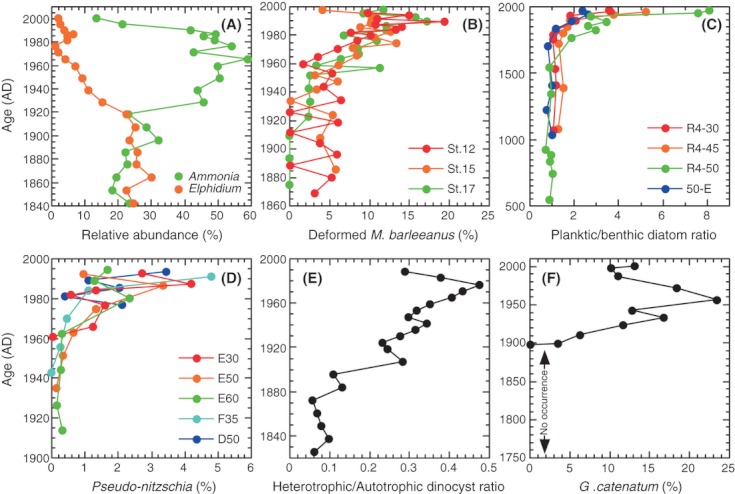
Representative downcore faunal and floral changes. Relative abundance of foraminiferal genera, Ammonia and Elphidium, in core OBY, Osaka Bay (Tsujimoto et al. 2008) (A); relative abundance of deformed specimens of foraminiferal species, Melonis barleeanus in sediment cores St. 12, St. 15, and St. 17 in a Greenlandic fjord (Elberling et al. 2003) (B); Planktic/benthic diatom ratio in Chesapeake Bay cores R4-30, R4-45, R4-50, and 50-E (Cooper 1995) (C); relative abundance of toxic diatom genus, Pseudo-nitzschia, in Gulf of Mexico cores E30, E50, E60, F35, and D50 (Parsons and Dortch 2002) (D); Heterotrophic/Autotrophic ratio of dinoflagellate cysts in the Adriatic Sea (Sangiorgi and Donders 2004) (E); and relative abundance of introduced dinoflagellate species, Gymnodinium catenatum, in Portuguese Margin, North Atlantic (Amorim and Dale 2006) (F).
Benthic Ostracoda
There is no universal response by particular benthic ostracod taxa to ecological degradation, probably in part because the limited dispersal abilities of these animals result in regionally differentiated faunas. There are, however, several tolerant species known in each region, for example, Loxoconcha sp. in the eastern coast of USA (Alvarez Zarikian et al. 2000; Cronin and Vann 2003) and Bicornucythere bisanensis in Japan (Irizuki et al. 2003; Yasuhara et al. 2003, 2007).
Diatoms and dinoflagellates
Eutrophic ecosystems are also characterized by the dominance of planktic diatoms and the decline of benthic diatoms (Clarke et al. 2006; Katsuki et al. 2009; Tuovinen et al. 2010), and by the increased dominance of heterotrophic dinoflagellates at the expense of autotrophic species (Dale 2000; Matsuoka 2001; Sangiorgi and Donders 2004), although a few exceptions are known (Chmura et al. 2004; Liu et al. 2008). In Chesapeake Bay, the planktic/benthic diatom ratio increased dramatically after European settlement and land clearance at ∼1700 (Cooper 1995) (Fig. 9). In the Adriatic Sea, the heterotrophic/autotrophic dinoflagellate cyst ratio increased beginning in ∼1900 due to a progressive increase in eutrophication (Sangiorgi and Donders 2004) (Fig. 9). An increase in Chaetoceros diatom cysts is also a typical signal of eutrophic ecosystems (Miller and Risberg 1990; Andrén 1999; Andrén et al. 1999).
Toxic bloom, introduced species, and deformity
Recent increases in toxic blooms and introduced species are recorded in microfossil records from several sites in the USA (McGann and Sloan 1996; McGann et al. 2000; Parsons and Dortch 2002), Europe (Amorim and Dale 2006; Marret et al. 2009), Australia (McMinn et al. 1997), and New Zealand (Irwin et al. 2003; Grenfell et al. 2007) (Fig. 10). For example, in the Gulf of Mexico, the toxic diatom genus Pseudo-nitzschia has rapidly increased since 1960 (Parsons and Dortch 2002) (Fig. 9) and the toxic dinoflagellate Gymnodinium catenatum has been introduced into the eastern Atlantic at ∼1900 via ballast water (Amorim and Dale 2006) (Fig. 9). Late-20th century introductions and blooms of this species are also known in Australia and New Zealand (McMinn et al. 1997; Irwin et al. 2003). On the western coast of the USA, introduction of the Japanese foraminiferal species Trochammina hadai is also known from ∼100 years ago (McGann 2008) and other benthic foraminiferal introductions are known in New Zealand (Grenfell et al. 2007). The earliest record of introduced microfossil species is from the southwestern Black Sea shelf at ∼1000 BC, possibly associated with early Greek exploration (Marret et al. 2009).
Figure 10.
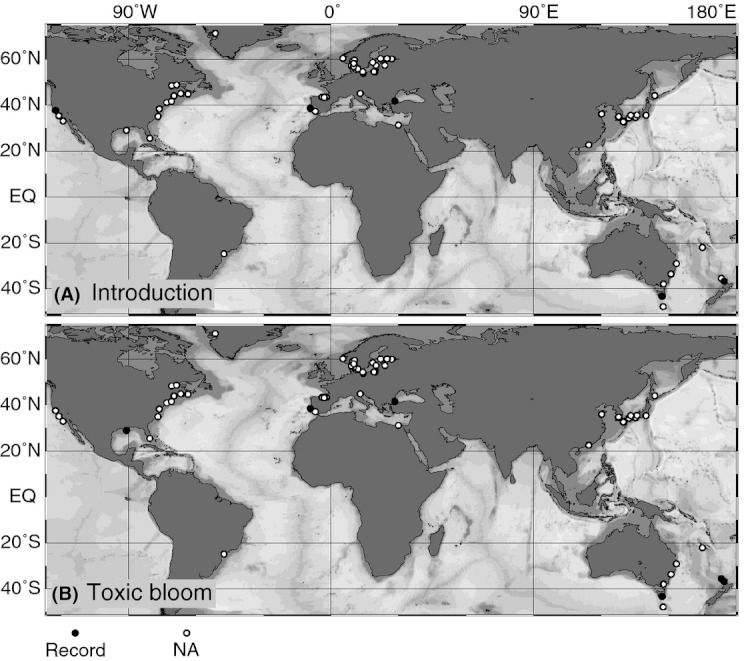
Global distribution of marine ecological degradation in downcore microfossil records. Distribution of (A) introduction and (B) toxic bloom events. NA: site with no introduction or toxic bloom evidence. Data from Table S1.
Deformed specimens of foraminifera also increase with increased human-induced stresses such as pollution by heavy metals (Elberling et al. 2003) and polychlorinated biphenyls (Scott et al. 2005) (Fig. 9) and hypoxia/anoxia (Karlsen et al. 2000), although the ultimate cause of foraminiferal deformity is still controversial (Polovodova and Schönfeld 2008; Martinez-Colon et al. 2009). In Aburatsubo Cove, Japan, specimens of ostracod Bicornucythere bisanensis with variant carapace ridges are relatively abundant (∼10–20%), which may be related to pollution because such variant ridges are never found in Holocene and Pliocene fossil specimens (Abe 1983).
Overview of regional trends
North American waters
In North America, the marine ecological degradation records are concentrated in the Atlantic Coast (e.g., Chesapeake Bay, Long Island Sound, and Canadian harbors), and Gulf of Mexico. In Chesapeake Bay, initial signs of ecological degradation are recognizable as an increase in total diatom abundance and in the diatom planktic/benthic ratio at around 1700, as a consequence of European settlement and deforestation and resulting increased river discharge and initial eutrophication (Cooper 1995; Cronin and Vann 2003). Later at ∼1800, diatom diversity started to decrease (Cooper 1995), because of further deforestation (Cronin and Vann 2003; Willard and Cronin 2007; Brush 2009) and perhaps increased human activity related to industrialization. However, the most drastic changes occurred ∼1960 due to urbanization and substantially increased population and fertilizer use (Cronin and Vann 2003; Willard and Cronin 2007; Brush 2009). The resulting eutrophication, hypoxia, and anoxia have become much more severe and widespread since this time, with an accelerated increase in the diatom planktic/benthic ratio and changes in the ostracod and foraminiferal fauna (Cooper 1995; Karlsen et al. 2000; Cronin and Vann 2003; Willard and Cronin 2007; Brush 2009).
A core drilled in the bottom of the deep channel provided a continuous high-resolution paleoecological record for the last 8000 years (Fig. S5) and clearly showed the unique nature of the mid–late 20th century ecosystem. The ostracod Loxoconcha sp. is solely dominant throughout most of the last 8000 years. Since this species is known to be common in hypoxic environments (Alvarez Zarikian et al. 2000),), the continuous dominance of this species suggests that the Chesapeake Bay deep benthic ecosystem has experienced hypoxia to some degree for at least the last 8000 years even without human influence. Currently, anoxia and hypoxia are widespread in the Chesapeake Bay. Cytheromorpha curta, another species that is known to be hypoxia-tolerant or opportunistic (Cronin and Vann 2003) and was never found in pre-1940 sediments, rapidly became dominant beginning in the mid-20th century, although Loxoconcha sp. is also still common (Fig. S5). This faunal change likely represents further ecological degradation. Ostracod and foraminiferal data in a shallower, channel edge core, which were taken in an area that currently experiences hypoxia, show a similar trend, in which Cytheromorpha curta and the foraminiferal species Ammonia parkinsoniana, rare or absent in pre-20th century sediments, became dominant in the mid–late 20th century (Karlsen et al. Cronin and Vann 2003). The relative abundance of deformed shells of Ammonia parkinsoniana also increased in this period. The relative abundance of Loxoconcha sp. showed multi-decadal–centennial oscillations ranging from 0 to 40% in pre-20th century sediments (Cronin and Vann 2003), suggesting that the shallower benthic ecosystem was only periodically hypoxic, in contrast with the deep channel [Similar patterns are also known in the Gulf of Mexico (Platon et al. 2005; Osterman et al. 2009)]. Currently, Loxoconcha sp. is widely dominant in the deep channel of the Chesapeake Bay (Cronin and Vann 2003).
Microfossils also provide evidence of post-1960s ecological degradation in the Gulf of Mexico (Blackwelder et al. 1996; Sen Gupta et al. 1996; Alvarez Zarikian et al. 2000; Osterman et al. 2005; Platon et al. 2005; Rabalais et al. 2007; Osterman et al. 2008, 2009). Ostracod and foraminiferal diversity rapidly decreased beginning in ∼1960 (Blackwelder et al. 1996; Alvarez Zarikian et al. 2000; Rabalais et al. 2007). At the same time, tolerant or opportunistic foraminiferal (e.g., Ammonia parkinsoniana), ostracod (e.g., Loxoconcha sp.), and diatom (e.g., Pseudo-nitzschia pseudodelicatissima,) species became dominant and other (sensitive) species (e.g., Elphidium spp. for foraminifera) collapsed (Sen Gupta et al. 1996; Alvarez Zarikian et al. 2000; Parsons and Dortch 2002; Osterman et al. 2005; Platon et al. 2005; Rabalais et al. 2007; Osterman et al. 2008, 2009). These faunal changes are associated with eutrophication and hypoxia resulting from population growth, urbanization, and increased use of fertilizers in the post-World War II era (Rabalais et al. 2002, 2007). Similar post-World War II ecological degradation is known from the Neuse and Pamlico River estuaries (Cooper et al. 2004), Long Island Sound (Thomas et al. 2004), Canadian harbors, estuaries, and fjords (Schafer et al. 1991; Scott et al. 1995, 2005; Thibodeau et al. 2006), and the western coast of North America of Southern California Bight (McGann et al. 2003; McGann 2009) and San Francisco Bay (McGann 2008).
Ecosystem impacts of land clearance and deforestation is also found in other microfossil records in North American waters, for example, in New Bedford Harbor as an increase in autotrophic dinoflagellates (Chmura et al. 2004), in Merrymeeting Bay as an increase in diatom abundance (Köster et al. 2007), and in Halifax Harbour as an increase in foraminiferal and dinoflagellate abundance (Miller et al. 1982). Microfossil assemblages, such as foraminifera, diatom, and dinoflagellate cyst, also provide evidence of urbanization impacts appeared as early as ∼1900 in shallow-marine ecosystems adjacent to several cities, including New Bedford Harbor (Pospelova et al. 2002; Chmura et al. 2004; Scott et al. 2005) and Merrymeeting Bay (Köster et al. 2007).
European and African waters
In Europe, numerous studies have been conducted in the Baltic Sea, mainly using diatoms. In the urban sites of the Gulf of Finland, increases in the relative abundance of planktic diatoms reflect urbanization-induced eutrophication beginning early in 19th century (Weckstrom et al. 2004; Kauppila et al. 2005; Weckström 2006). After this initial degradation, species diversity started to decrease starting in the late 19th century, and planktic and eutrophication-indicator species are dominant throughout the 20th century (Weckstrom et al. 2004; Kauppila et al. 2005; Clarke et al. 2006; Weckström 2006; Weckström et al. 2007). A clear recovery of species diversity is observed since the 1970s related to the cessation of wastewater loading, although planktic diatoms are still dominant (Kauppila et al. 2005; Clarke et al. 2006; Weckström 2006; Weckström et al. 2007). In contrast, the ecological degradation observed in the rural sites of the gulf was minor, and is reflected in the microfossil record as an increasing relative abundance of planktic diatoms and a slight diatom diversity decrease (but not always clear); these changes occurred later, starting in the 1940s and accelerating in 1980s, mainly due to intensification of agriculture in Finland after the World War II (Weckström 2006; Weckström et al. 2007). Similar trends are found in other locations in the Baltic Sea (Miller and Risberg 1990; Grönlund 1993; Witkowski and Pampkowiak 1995; Korhola and Blom 1996; Tikkanen et al. 1997; Andrén 1999; Andrén et al. 1999, 2000; Jankowska et al. 2005; Witak et al. 2005; Witak and Jankowska 2005; Witak et al. 2006; Leśniewska and Witak 2008; Olli et al. 2008; Tuovinen et al. 2010). Töölönlahti Bay is one of the best examples of marine ecosystem recovery observed in the microfossil record (Tikkanen et al. 1997). Species diversity and faunal composition almost completely recovered to the 19th century condition after cessation of waste-water disposal in the 1960s (Tikkanen et al. 1997).
Intensive research has also been conducted in northern European fjords mainly using foraminifera and dinoflagellate cysts. Available evidence suggests that marine ecological degradation extends back to mid 16th century in this region. In Frierfjord, Norway, marine ecological degradation probably started at ∼1550 associated with initial eutrophication caused by saw mills (Alve 2000; Dale 2000). In Oslofjord, Norway, initial ecological degradation dates to 1850, as evidenced by a decrease in foraminiferal species diversity and an increase in the abundance of total dinoflagellates and Lingulodinium machaerophorum; this was caused by eutrophication associated with human population growth (Nagy and Alve 1987; Alve 1991; Dale and Fjellså 1994; Dale et al. 1999; Dale 2000; Alve et al. 2009). After that, from the early to mid 20th century, a rapid increase in the abundance of total foraminifera occurred, together with an increase in tolerant, opportunistic, and arenaceous foraminiferal species and a decrease in sensitive and calcareous foraminiferal species (Nagy and Alve 1987; Alve 1991; Alve et al. 2009). Ecosystem recovery began in the 1970s as a result of relocating much of the Oslo City's sewage treatment (Dale et al. 1999; Alve et al. 2009). Similar ecological degradation histories are known from microfossil records in other North European fjords (Moodley et al. 1993; Paetzel and Schrader 1995; Alve 1996; Hass 1997a,b; Thorsen and Dale 1997; Amsinck et al. 2003; Clarke et al. 2003; Ryves et al. 2004; Clarke et al. 2006; Ellegaard et al. 2006). In Limfjord and Vejlerne Nature Reserve, Denmark, remarkable faunal and floral changes were caused by salinity decrease due to land reclamation and closure of the fjord in the 1870s (Amsinck et al. 2003; Ryves et al. 2004).
In Europe, microfossils also record marine ecological degradation in Spanish estuaries caused by acidic wastewater, metal pollution, eutrophication, oxygen depletion, and increased sedimentation as a result of mining and smelting operations, chemical factories, petroleum refineries, industrialization, and land reclamation (González-Regalado et al. 1996; Ruiz Muñoz et al. 1997; Cearreta et al. 2000; Gonzalez-Regalado et al. 2001; Cearreta et al. 2002a,b; Pascual et al. 2002; Ruiz et al. 2004; Irabien et al. 2008; Ruiz et al. 2008, 2009). In Portugal, ecosystem changes in the Albufeira Lagoon were caused by salinity increase as a result of anthropogenic alterations of the barrier (Bao et al. 1999); whereas on the Atlantic Portuguese coast, changes were caused by urbanization-induced eutrophication and pollution (Amorim and Dale 2006; Bartels-Jónsdóttir et al. 2006, 2009). In the Adriatic Sea, changes were caused by eutrophication and anoxia as a result of human activities including agriculture, wastewater disposal, and diversion of river outflow (Puskaric et al. 1990; Barmawidjaja et al. 1995; Sangiorgi and Donders 2004).
There are no African data available, except for foraminiferal research along the Mediterranean coast of Egypt, where large faunal change was caused by causeway reconstruction ∼2000 years ago (Bernasconi et al. 2006).
Asian waters
In Asia, most late Holocene marine micropaleontological records are from Japan. For example, in Osaka Bay, microfossils indicate that marine ecological degradation started with the beginning of the Japanese industrialization period at ∼1900. Foraminiferal diversity and abundance started to decrease and increase, respectively, and a decline of sensitive species and increase in tolerant or opportunistic species also started at that time. At the same time ostracod faunal patterns changed in a similar fashion, and ostracod abundance decreased in the inner part the bay. During the Japanese high economic growth period of 1950s–1970s, these foraminiferal and ostracod faunal trends accelerated. The degradation of Osaka Bay was mainly caused by urbanization-induced eutrophication and resulting bottom-water hypoxia. In the middle part of the bay, ostracod abundance increased, especially during the mid–late 20th century, due to increased nutrient supply without hypoxia. In the inner part of Osaka Bay, the abundance of the opportunistic foraminiferal species decreased and the foraminiferal species diversity increased since the 1970s. This recovery was a result of environmental regulation, which led to a reduction in organic pollution loads and a decrease in food supply to the benthos. However, modern foraminiferal distribution is still dominated by the opportunistic species. In addition to foraminiferal and ostracod records, dinoflagellate microfossils are also abundant in Japanese and Korean embayments, and show eutrophication-induced increases in heterotrophic species during the early to mid 20th century (Matsuoka 1995; Kim and Matsuoka 1998; Matsuoka 1999; Matsuoka and Kim 1999; Matsuoka 2001; Kim et al. 2009; Matsuoka and Shin 2010; Shin et al. 2010).
There are also many data available from Japanese brackish lakes. In Lake Nakaumi, significant ecological degradation had occurred since the 1940s due to enhanced salinity variability and bottom-water hypoxia as a result of construction projects including land reclamation and dike construction, as well as human-induced eutrophication. Benthic diatom disappeared after the 1940s, red-tide diatom became dominant in 1950s -1960s, and foraminiferal and ostracod faunas had altered since 1940. In Lake Saroma, scallop farming since the 1960s has caused intensive eutrophication and bottom-water hypoxia and resulting ecological degradation, that is represented by increase and decrease in the planktic and benthic diatoms, respectively.
Elsewhere, microfossils recording 20th century eutrophication-induced marine ecological degradation have also been analyzed in Ariake and Isahaya Bays, Japan (Akimoto et al. 2004; Matsuoka 2004; Yokose et al. 2005); in Hiroshima Bay, Japan (Yasuhara et al. 2003); in Tokyo Bay, Japan (Matsumoto 1981, 1983; Matsumoto and Saito 1984; Ikeya 1995; Sato 1995; Toyoda and Kitazato 1995; Matsuoka 1999; Tanimura et al. 2003); Lake Kugushi, Japan (Nomura and Kawano 2011); Jiaozhou Bay, China (Liu et al. 2008); and Daya Bay, South China Sea (Wang et al. 2004).
Southern hemisphere waters
In the southern hemisphere, the few late Holocene marine microfossil studies conducted are concentrated in New Zealand and Australia. In Australia, salinity-change-induced ecological degradation and introduction of exotic species are recorded for the eastern coast, which were caused by construction and ballast water release, respectively, during the late 19th–20th century (McMinn et al. 1997; Saunders et al. 2007, 2008; Taffs et al. 2008; Saunders and Taffs 2009). New Zealand studies in the North Island harbors indicated that Polynesian forest clearance caused minor but initial ecological degradation starting at ∼1500 (Hayward et al. 2006). European land clearance since ∼1840 and late-20th-century urbanization had a much larger impact on the marine ecosystem (Hayward et al. 2004; Matthews et al. 2005; Hayward et al. 2006; Grenfell et al. 2007; Hayward et al. 2008). Details of North Island harbors' ecological degradation is complex and involves industrial and domestic sewage disposal, introduction of Asian date mussels and cordgrass, land clearance/deforestation, urbanization, and oyster farming (see, Hayward et al. 2004; Matthews et al. 2005; Hayward et al. 2006; Grenfell et al. 2007; Hayward et al. 2008).
In South America, the only available data are from the Atlantic coast of Brazil (de Mahiques et al. 2009). Foraminiferal faunal changes including increase in agglutinated species and decrease in calcareous species, species diversity, and abundance in that region were likely caused by salinity decrease and heavy metal pollution as a result of opening of an artificial channel and mining activities since the 1850s (de Mahiques et al. 2009).
Problems, solutions, and outlook of marine paleoecological approach based on microfossils
As reviewed in the present paper, microfossil paleoecology is useful for marine ecological degradation research. However, there are various limitations and concerns inherent to this approach. In this section, we discuss these issues and their possible solutions, along with the outlook for developing microfossil-based marine ecological degradation research in the future.
Climate versus human-induced environmental changes
Although our micropaleontological literature review strongly suggests that most instances of marine ecological degradation are caused by eutrophication and resulting hypoxia, many of the studies are qualitative and their interpretation is subject to some uncertainties. Furthermore, these studies generally do not take into account possible effects of climatic changes, including major events such as the Little Ice Age and 20th century global warming, although a few exceptions use statistical approaches to do so (e.g., Uthicke et al. 2012). Although changes in high-human footprint habitats such as urban embayments are not easily attributable to climate cooling and warming, the possibility that ecological degradation is related to climatic change merits further consideration, focusing on these two climatic events.
Little Ice Age
The Little Ice Age (AD ∼1400–1700) is the most recent major climatic cooling event (Mann et al. 2009). In the Baltic Sea, Andrén et al. (2000) reported a detectable influence of the Little Ice Age on fossil diatom assemblages. However, clear indications of the Little Ice Age (e.g., increase in cold water indicator species) are rarely detected in microfossil faunal and floral records. In fact, no other research in the Baltic Sea has detected a clear paleoecological footprint of the Little Ice Age. In the Chesapeake Bay, trace element analysis of ostracod shells clearly showed Little Ice Age cooling (Cronin et al. 2005; Willard and Cronin 2007; Cronin et al. 2010). Although the climate change was clearly present in geochemical records, no clear biotic response could be detected in marine microfossil communities (Karlsen et al. 2000; Cronin and Vann 2003). Furthermore, the magnitude of temperature decline during the Little Ice Age is relatively small (<1°C) (Mann et al. 2009) and thus it is unlikely that recent microfossil evidence of ecological degradation reviewed in the present paper is mainly caused by this cooling event.
Global warming
Recent research suggests that human-induced global warming negatively affects marine biodiversity (Harley 2011). However, ecological theories suggest positive relationship between temperature and biodiversity both on evolutionary and ecological time-scales (Allen et al. 2002; Currie et al. 2004; Yasuhara et al. 2012b). Thus, it is still largely uncertain how global warming affects marine ecosystems.
Furthermore, global warming since ∼1900 has increased temperatures by less than 1°C (Intergovernmental Panel on Climate Change 2007), which is small compared with seasonal and yearly fluctuations [although some habitats, such as coral reefs, may be particularly sensitive to warming (e.g., Pandolfi et al. 2011)]. Moreover, many of the ecosystem degradation events compiled here predate 20th century warming. In addition, the autoecology of affected taxa, and extrinsic information about land use and environmental change consistently suggest that eutrophication and the resulting hypoxia is the predominant cause of marine ecosystem degradation as reviewed in this article. Recent studies suggest that global-warming footprints on marine ecosystems are detectable from the systems much less influenced by human-induced nutrient enrichment, for example, pelagic, open ocean (Field et al. 2006; Spielhagen et al. 2011). Nevertheless, much of this evidence is qualitative in nature, and we discuss below how to improve this situation. Climatic change may also be related to eutrophication itself (Savage et al. 2010), and the connection between these two phenomena warrants further research.
Temporal resolution, time-averaging, and taphonomic bias
Robust chronology is essential for the accurate reconstruction of the past ecological changes from sedimentary records. There are various tools for inferring age-depth relationships within cores spanning hundreds to thousands of years. Radiometric dating methods, for example 210Pb, 137Cs, and 14C, are widely used on this time scale. Radiocarbon (14C) dating has ∼101–102 years error and thus does not have sufficient resolution for the past 100–200 years sediments, but it is useful for older sediments. In contrast, 210Pb and 137Cs methods are applicable only for the past ∼100–200 years sediments, because half-life of 210Pb is short and 137Cs method is based on detection of nuclear testing or accident events. Other dating methods including pollen biostratigraphy and tephrochronology are also often useful, depending on the regions and time intervals. Thus, usage of multiple complementary methods is important for reconstructing robust chronology throughout the past 102–103 years (e.g., Willard et al. 2003). Furthermore, selecting high sedimentation rate sites are important to obtain high time-resolution records. For further details of dating methods, see Cronin (2010).
Bioturbation and physical mixing combines fossils from different intervals of time and thereby places a practical limit on the temporal resolution of sediment records. Under most conditions, yearly resolution is not attainable, but decadal-scale resolution can be, as long as the sedimentation rate is high enough (e.g., Cronin et al. 2010). Fortuitously, bottom-water hypoxia tends to increase the temporal acuity of sedimentary records because few bioturbating animals can tolerate very low-oxygen levels. Thus, cores recording marine ecological degradation from this cause tend to record decadal signal well, especially during degraded periods.
Microfossil shells are small and thus can be transported by current after their death. Such postmortem transportation is more likely at sites dominated by coarser (i.e., sandy) sediments that indicate deposition under stronger currents than at muddy sites typical of low-energy environments. Although rarely applied, there are several ways proposed to detect such postmortem transport (e.g., see Whatley 1988 for ostracods).
Selective preservation is also a potential problem, especially in higher energy environments. Microfossil species with robust shells will be better preserved than those with thin, delicate shells. Shells of earlier juveniles tend to be thinner and more delicate, that is also a potential cause of selective preservation. However, microfossils are still generally very well preserved in sedimentary records compared with any other fossil groups. Preservation states of microfossils can be tested by, for example, fragmentation rate and clarity (Martin 1999; Dwyer et al. 2002; Sexton et al. 2006).
Time-averaging, selective preservation, and transport may render fossil assemblages biased samples of their source live communities, but intensive assessments show good fidelity between living organisms and the death assemblage of shells for mollusks (Kidwell 2001; Tomašových and Kidwell 2009). Such live-death fidelity is also known in microfossils (e.g., Kornicker 1964; Izuka and Kaesler 1986; Wollenburg and Kuhnt 2000). Time-averaging can even be an advantage in ecological research. Fossil samples typically represent more than 1 year (usually a few to tens of years, depending on sedimentation rate and sample thickness). This multi-year averaging means that variability at shorter time-scales (i.e., hourly, daily, and seasonal) can be eliminated (Olszewski 1999). This temporal smoothing is an advantage because, in the context of ecological degradation, such short-term variation is usually just noise around what is of much greater interest – shifts in the long-term state of the community. Substantial effort can be required to determine annual or multi-year average conditions when sampling modern biota (see Kidwell 2009).
Interpretation of paleoecological records: beyond narrative interpretation and toward more quantitative and statistical approach
In our review of these microfossil records, our interpretations about the inception and acceleration of degradation were necessarily qualitative because the raw data are not available for most studies, and because many of the original studies were essentially descriptive. However, several studies used multivariate analyses and visualization methods effectively.
Changes in faunal and floral composition are typical of marine ecological degradation. Such signs can be detected as changes in specific species or genera as reviewed in the Taxon-specific responses section. But, multivariate analyses such as cluster analysis, nonmetric multidimensional scaling (NMDS), and detrended correspondence analysis (DCA) are important for summarizing and detecting whole-assemblage composition changes (Murray 2000; Yasuhara et al. 2003; Tuovinen et al. 2010; Uthicke et al. 2012). For example, in the NMDS plots of Osaka Bay core OS3, pre-1950 and post-1950 samples segregate neatly, showing the transition from pre-urbanization to post-urbanization assemblages coincident with Japan's high economic growth and resulting eutrophication and hypoxia after World War II (Fig. 11).
Figure 11.
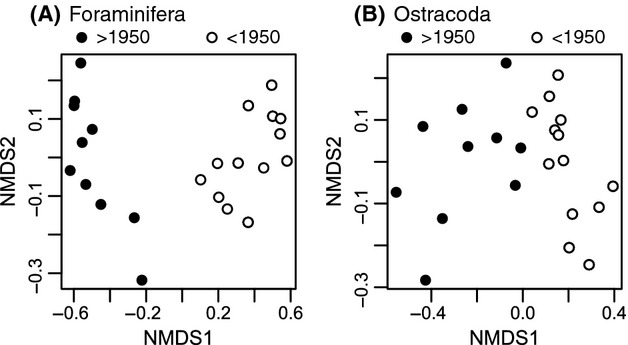
Ordination (NMDS) of microfossil relative abundance data in Osaka Bay core OS3. (A) foraminifera. (B) ostracods. Solid circles: post-1950 samples. Open circles: pre-1950 samples. Data from Tsujimoto et al. (2008) and Yasuhara et al. (2007).
Rank-abundance distributions can also show clear evidence of altered faunal composition (Scott et al. 2001). In Osaka Bay core OBY, rank-abundance distributions were remarkably different between pre- and post-urbanization (∼1950) faunas (Fig. 12). Pre-urbanization faunal composition has a relatively equitable distribution of abundances from dominant to rare species, with a long tail of rare species. In contrast, the post-urbanization fauna is dominated by only three resistant species at the expense of the rest of the fauna, with lower species richness overall. Although more richly informative than descriptive approaches, these more quantitative methods have been applied in only a minority of published studies.
Figure 12.
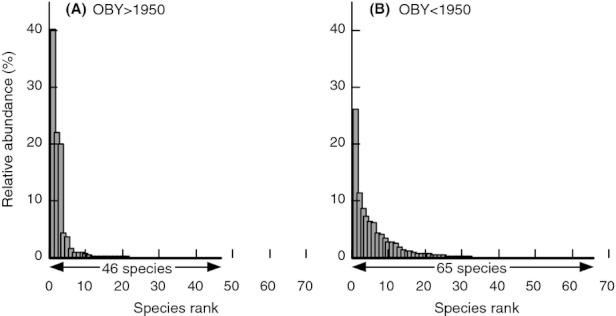
Rank-abundance distributions of foraminifera in Osaka Bay core OBY. (A) post-1950 assemblage (46 species in total). (B) pre-1950 assemblage (65 species in total). Data from Tsujimoto et al. (2008).
Ecosystem history research and microfossil-based reconstruction of environmental history may sometimes be two sides of the same coin. For example, we can interpret biodiversity decline and other biotic changes as ecological degradation, but the same information can be used as an indicator of pollution. This equivalence assumes a direct, immediate, and linear response between environmental changes and biotic response of microfossil communities. In the absence of independent proxies for environmental history, these assumptions cannot be tested. It is therefore an important priority for future work to document changes in assemblages jointly with geochemical and other independent proxies for environmental alteration (see e.g., Brush 2008). Moreover, it will be most productive to do so in a rigorous statistical framework, which is one aspect in which marine paleoecological research lags behind paleolimnology (e.g., Cottingham et al. 2000; Rusak et al. 2004; Tsugeki et al. 2009). Recently, paleoecological researchers have started to use multiple regression and other statistical modeling approaches to investigate climatic impact on deep-sea ecosystems. For example, recent work has compared microfossil species diversity with independent proxies for temperature, surface productivity and seasonality of productivity, and using multiple regression and model averaging (Hunt et al. 2005; Yasuhara et al. 2009, 2012a,c). However, such approaches have seldom been applied to understanding human-induced ecological degradation research in marine systems.
Independent evidence of environmental history can be obtained by a variety of means. There are well-established methods for reconstructing concentrations of various heavy metals and organic pollutants, oxygen content, temperature, salinity, and pH using core sediments (Cronin et al. 2005; Kemp et al. 2005; Yasuhara and Yamazaki 2005; Pelejero et al. 2010), and stable isotopes and trace elements of microfossil shells allow inference of various environmental parameters (Cronin 2010). These data will be critical in attempts to assess the relative importance of climate versus eutrophication, or organic pollution versus metal pollution, in affecting the marine organisms.
Other microfossil records indicative of ecological history
Sediment core records of fish scales are another rich archive of long-term ecological history. However, because fish population changes reconstructed from fish scale records tend to be interpreted as responses to natural climate changes (Field et al. 2009; Finney et al. 2010) we did not include these studies in this review of human-induced ecological degradation although future work may explore the relative importance of natural climate fluctuations versus overfishing through explicit statistical modeling. Similarly, our review excluded phytoplankton pigment records because of our focus on body fossils, rather than chemical remains. Pigment studies are rarer in marine systems compared with freshwater habitats (see Leavitt and Hodgson 2002), and they are concentrated in well-studied regions regarding microfossils such as Gulf of Mexico and Baltic Sea where their results have complemented those from more traditional microfossil studies (Reuss et al. 2005; Rabalais et al. 2007; Savage et al. 2010).
Unexplored ecosystems
Microfossil-based paleoecological studies of marine ecological degradation are still largely limited to mid-latitude estuaries and embayments. There is almost no such research in tropical coral reefs and mangroves, in polar environments, or in the deep sea, in spite of the high vulnerability of these habitats and public concern about them. Such studies on open shelf communities are also limited in number compared with those on estuary and embayment communities (Table S1).
Unexplored environmental issues
Ocean acidification is one of the most serious problems facing ocean life today, especially for organisms with calcified external skeletons (Orr et al. 2005; Hoegh-Guldberg et al. 2007; Fabry et al. 2008). Thus, calcareous microfossils such as foraminifera and ostracods are ideal for investigating the ecological impact of acidification over time. Micropaleontological research has revealed that glacial-interglacial changes in atmospheric carbon dioxide with the resulting changes in sea-water acidity have influenced marine calcification over the last 50,000 years (Barker and Elderfield 2002). However, culture experiments to determine the physiological effects of acidification on the organisms used in micropaleontological analyses have just begun (Spero et al. 1997; Kuroyanagi et al. 2009; Fujita et al. 2011).
Conclusions
In this review, we analyzed patterns in published microfossil records to better understand spatiotemporal trends of marine ecological degradation across multiple scales, and to illustrate the usefulness of microfossils in detecting ongoing ecological changes during the Anthropocene (Zalasiewicz et al. 2008). The results indicated that: (1) ecological degradation in marine systems began significantly earlier in Europe and North America (∼1800s) compared with Asia (post-1900) due to earlier industrialization in European and North American countries, (2) ecological degradation accelerated globally in the late 20th century due to post-World War II economic and population growth and exponential increase in use of chemical fertilizers, (3) recovery from the degraded state in late 20th century after restoration efforts and environmental regulations were implemented only in limited localities. The predominant cause of degradation detected in microfossil records was nutrient enrichment and the resulting symptoms of eutrophication including hypoxia.
Degradation of ocean ecosystems has been characterized as the collapse of large animals and the rise of microbes (Jackson 2001; Jackson et al. 2001; Jackson 2008), with many nonresistant species becoming ecologically extinct and replaced by a few opportunistic or tolerant species. Microfossil records show similar processes evidenced by high-density and low-diversity assemblages in eutrophic coastal areas worldwide (Alve 1995; Tsujimoto et al. 2008).
Downcore microfossil studies are also important for an improved understanding of the natural baseline, which is often difficult to reconstruct (Pauly 1995; Jackson et al. 2001; Knowlton and Jackson 2008). Although microfossil-based human-induced ecological degradation research has often focus solely on the last few hundreds of years, longer records from the past thousands of years are needed to better understand the severity of ecological degradation relative to pre-impact baselines.
Acknowledgments
We thank T. M. Cronin and C. A. Alvarez Zarikian for microfossil data; A. Moore and an associate editor for editing; and N. Knowlton, T. M. Cronin, and an anonymous reviewer for comments and discussion. Numerous colleagues helped us to access their papers. This work was supported by Smithsonian Marine Science Network Postdoctoral Fellowship and a Seed Funding Programme for Basic Research of the University of Hong Kong (project code: 201111159140) (to M. Y.).
Conflict of Interest
None declared.
Supporting Information
Additional Supporting Information may be found in the online version of this article at the publisher's web site:
Figure S1. Published age distribution of papers compiled in this review. Data from Table S1.
{kind=link}
Figure S2. Global distribution of marine ecological degradation in downcore microfossil records. (a) ostracod and (b) foraminiferal data sites (all of them are benthic, except one planktic foraminiferal data site from Santa Barbara Basin, USA: see Table S1). NA: site with no ostracod or foraminiferal data. Data from Table S1.
{kind=link}
Figure S3. Global distribution of marine ecological degradation in downcore microfossil records. (a) diatom and (b) dinoflagellate data sites. NA: site with no diatom or dinoflagellate data. Data from Table S1.
{kind=link}
Figure S4. Global distribution of marine ecological degradation caused by human-induced eutrophication. NA: site with no eutrophication evidence. Data from Table S1.
{kind=link}
Figure S5. Chesapeake Bay paleoecological records in the core MD99-2209/RD-98. Loxoconcha sp. relative abundance record (A) and its closeup for the last 200 years (B); Cytheromorpha curta relative abundance record (C) and its closeup (D); and conceptual figure explaining core locations of deep-channel site of MD99-2209/RD-98 (This figure) and shallow, channel edge site (E: see text for detail). Data from Cronin et al. (2005) and Cronin (unpublished data).
{kind=link}
Table S1. Dataset used for the present paper (see Supplementary Text for references). Ostracod: benthic ostracod; Foram: benthic foraminifera. Planktic foram: planktic foraminifera. Dino: dinoflagellate. ND: not detectable.
Supplementary Text S1. References cited in Table S1.
References
- Abe K. Population structure of Keijella bisanensis (Okubo) (Ostracoda, Crustacea) – An inquiry into how far the population structure will be preserved in the fossil record. J. Fac. Sci., Univ. Tokyo. Sec. II, Geol. Mineral. Geogr. Geophys. 1983;20:443–488. [Google Scholar]
- Airoldi L, Beck MW. Loss, status and trends for coastal marine habitat of Europe. Oceanogr. Mar. Biol. Annu. Rev. 2007;45:345–405. [Google Scholar]
- Akimoto K, Nakamura K, Kondo H, Ishiga H, Dozen K. Environmental reconstruction based on heavy metals, diatoms and benthic foraminifes in the Isahaya reclamation area, Nagasaki, Japan. Environ. Micropaleontol., Microbiol. and Meiobenthol. 2004;1:83–104. [Google Scholar]
- Allen AP, Brown JH, Gillooly JF. Global biodiversity, biochemical kinetics, and the energetic-equivalence rule. Science. 2002;297:1545–1548. doi: 10.1126/science.1072380. [DOI] [PubMed] [Google Scholar]
- Alvarez Zarikian CA, Blackwelder PL, Hood T, Nelsen TA, Featherstone C. 2000. pp. 896–905. Ostracods as indicators of natural and anthropogenically-induced changes in coastal marine environments. Coasts at the Millennium, Proceedings of the 17th International Conference of The Coastal Society, Portland, OR USA, July 9-12.
- Alve E. Foraminifera, climatic change, and pollution: a study of late Holocene sediments in Drammensfjord, southeast Norway. Holocene. 1991;1:243–261. [Google Scholar]
- Alve E. Benthic foraminiferal responses to estuarine pollution: a review. J. Foramin. Res. 1995;25:190–203. [Google Scholar]
- Alve E. Benthic foraminiferal evidence of environmental change in the Skagerrak over the past six decades. Nor. Geol. Unders. 1996;430:85–93. [Google Scholar]
- Alve E. Environmental stratigraphy: a case study reconstructing bottom water oxygen conditions in Frierfjord, Norway, over the past five centuries. In: Martin RE, editor. Environmental micropaleontology: the application of microfossils to environmental geology. New York: Kluwer Academic/Plenum Publishers; 2000. pp. 324–350. [Google Scholar]
- Alve E, Lepland A, Magnusson J, Backer-Owe K. Monitoring strategies for re-establishment of ecological reference conditions: possibilities and limitations. Mar. Pollut. Bull. 2009;59:297–310. doi: 10.1016/j.marpolbul.2009.08.011. [DOI] [PubMed] [Google Scholar]
- Amorim A, Dale B. Historical cyst record as evidence for the recent introduction of the dinoflagellate Gymnodinium catenatum in the north-eastern Atlantic. Afr. J. Mar. Sci. 2006;28:193–197. [Google Scholar]
- Amsinck SL, Jeppesen E, Ryves D. Cladoceran stratigraphy in two shallow brackish lakes with special reference to changes in salinity, macrophyte abundance and fish predation. J. Paleolimnol. 2003;29:495–507. [Google Scholar]
- Andrén E. Changes in the composition of the diatom flora during the last century indicate increased eutrophication of the Oder Estuary, south-western Baltic Sea. Estuar. Coast. Shelf Sci. 1999;48:665–676. [Google Scholar]
- Andrén E, Shimmield G, Brand T. Environmental changes of the last three centuries indicated by siliceous microfossil records from the southwestern Baltic Sea. Holocene. 1999;9:25–38. [Google Scholar]
- Andrén E, Andrén T, Kunzendorf H. Holocene history of the Baltic Sea as a background for assessing records of human impact in the sediments of the Gotland Basin. Holocene. 2000;10:687–702. [Google Scholar]
- Bao R, Andrade M, da Codnceição Freitas C. Separating eustatic from local environmental effects: a late-Holocene record of coastal change in Albufeira Lagoon, Portugal. Holocene. 1999;9:341–352. [Google Scholar]
- Barker S, Elderfield H. Foraminiferal calcification response to glacial-interglacial changes in atmospheric CO2. Science. 2002;297:833–836. doi: 10.1126/science.1072815. [DOI] [PubMed] [Google Scholar]
- Barmawidjaja DM, Vanderzwaan GJ, Jorissen FJ, Puskaric S. 150 years of eutrophication in the northern Adriatic Sea: Evidence from a benthic foraminiferal record. Mar. Geol. 1995;122:367–384. [Google Scholar]
- Bartels-Jónsdóttir HB, Knudsen KL, Abrantes F, Lebreiro S, Eiriksson J. Climate variability during the last 2000 years in the Tagus Prodelta, western Iberian Margin: Benthic foraminifera and stable isotopes. Mar. Micropaleontol. 2006;59:83–103. [Google Scholar]
- Bartels-Jónsdóttir HB, Voelker AHL, Knudsen KL, Abrantes F. Twentieth-century warming and hydrographical changes in the Tagus Prodelta, eastern North Atlantic. Holocene. 2009;19:369–380. [Google Scholar]
- Bernasconi MP, Melis R, Stanley DJ. Benthic biofacies to interpret Holocene environmental changes and human impact in Alexandria's Eastern Harbour, Egypt. Holocene. 2006;16:1163–1176. [Google Scholar]
- Blackwelder P, Hood T, Alvarez-Zarikian C, Nelsen TA, McKee B. Benthic foraminifera from the NECOP study area impacted by the Mississippi River plume and seasonal hypoxia. Quatern. Int. 1996;31:19–36. [Google Scholar]
- Borja Á, Dauer DDM, Elliott M, Simenstad CA. Medium- and long-term recovery of estuarine and coastal ecosystems: patterns, rates and restoration effectiveness. Estuaries Coasts. 2010;33:1249–1260. [Google Scholar]
- Breitburg DL, Hondorp DW, Davias LA, Diaz RJ. Hypoxia, nitrogen, and fisheries: integrating effects across local and global landscapes. Ann. Rev. Mar. Sci. 2009;1:329–349. doi: 10.1146/annurev.marine.010908.163754. [DOI] [PubMed] [Google Scholar]
- Brush GS. Historical land use, nitrogen, and coastal eutrophication: a paleoecological perspective. Estuaries Coasts. 2009;32:18–28. [Google Scholar]
- Brush GS, Hilgartner WB. Paleoecology of submerged macrophytes in the upper Chesapeake Bay. Ecol. Monogr. 2000;70:645–667. [Google Scholar]
- Cardinale BJ, Palmer MA, Collins SL. Species diversity enhances ecosystem functioning through interspecific facilitation. Nature. 2002;415:426–429. doi: 10.1038/415426a. [DOI] [PubMed] [Google Scholar]
- Cearreta A, Irabien MJ, Leorri E, Yusta I, Croudace IW, Cundy AB. Recent anthropogenic impacts on the Bilbao estuary, Northern Spain: geochemical and microfaunal evidence. Estuar. Coast. Shelf Sci. 2000;50:571–592. [Google Scholar]
- Cearreta A, Irabien MJ, Leorri E, Yusta I, Quintanilla A, Zabaleta A. Environmental transformation of the Bilbao estuary, N. Spain: microfaunal and geochemical proxies in the recent sedimentary record. Mar. Pollut. Bull. 2002a;44:487–503. doi: 10.1016/s0025-326x(01)00261-2. [DOI] [PubMed] [Google Scholar]
- Cearreta A, Irabien MJ, Ulibarri I, Yusta I, Croudace IW, Cundy AB. Recent salt marsh development and natural regeneration of reclaimed areas in the Plentzia Estuary, N. Spain. Estuar. Coast. Shelf Sci. 2002b;54:863–886. [Google Scholar]
- Chmura GL, Santos A, Pospelova V, Spasojevic Z, Lam R, Latimer JS. Response of three paleo-primary production proxy measures to development of an urban estuary. Sci. Total Environ. 2004;320:225–243. doi: 10.1016/j.scitotenv.2003.08.003. [DOI] [PubMed] [Google Scholar]
- Clarke A, Juggins S, Conley D. A 150-year reconstruction of the history of coastal eutrophication in Roskilde Fjord, Denmark. Mar. Pollut. Bull. 2003;46:1615–1629. doi: 10.1016/S0025-326X(03)00375-8. [DOI] [PubMed] [Google Scholar]
- Clarke AL, Weckstrom K, Conley DJ, Anderson NJ, Adser F, Andren E, et al. Long-term trends in eutrophication and nutrients in the coastal zone. Limnol. Oceanogr. 2006;51:385–397. [Google Scholar]
- Coleman FC, Williams SL. Overexploiting marine ecosystem engineers: potential consequences for biodiversity. Trends Ecol. Evol. 2002;117:40–44. [Google Scholar]
- Cooper SR. Chesapeake Bay watershed historical land use: impact on water quality and diatom communities. Ecol. Appl. 1995;5:703–723. [Google Scholar]
- Cooper SR, McGlothlin SK, Madritch M, Jones DL. Paleoecological evidence of human impacts on the Neuse and Pamlico Estuaries of North Carolina, USA. Estuaries. 2004;27:617–633. [Google Scholar]
- Cottingham KL, Rusak JA, Leavitt PR. Increased ecosystem variability and reduced predictability following fertilisation: evidence from palaeolimnology. Ecol. Lett. 2000;3:340–348. [Google Scholar]
- Cronin TM. Paleoclimates: understanding climate change past and present. New York: Columbia University Press; 2010. [Google Scholar]
- Cronin TM, Raymo ME. Orbital forcing of deep-sea benthic species diversity. Nature. 1997;385:624–627. [Google Scholar]
- Cronin TM, Vann CD. The sedimentary record of climatic and anthropogenic influence on the Patuxent estuary and Chesapeake Bay ecosystems. Estuaries. 2003;26:196–209. [Google Scholar]
- Cronin TM, Thunell R, Dwyer GS, Saenger C, Mann ME, Vann C, et al. Multiproxy evidence of Holocene climate variability from estuarine sediments, eastern North America. Paleoceanography. 2005;20:PA4006. doi: 10.1029/2005PA001145. [Google Scholar]
- Cronin TM, Hayo K, Thunell RC, Dwyer GS, Saenger C, Willard DA. The Medieval Climate Anomaly and Little Ice Age in Chesapeake Bay and the North Atlantic Ocean. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010;297:299–310. [Google Scholar]
- Currie DJ, Mittelbach GG, Cornell HV, Field R, Guegan JF, Hawkins BA, et al. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 2004;7:1121–1134. [Google Scholar]
- Dale B. Dinoflagellate cysts as indicators of cultural eutrophication and industrial pollution in coastal sediments. In: Martin RE, editor. Environmental micropaleontology: the application of microfossils to environmental geology. New York: Kluwer Academic/Plenum Publishers; 2000. pp. 305–321. [Google Scholar]
- Dale B, Fjellså A. Dinoflagellate cysts as paleoproductivity indicators: state of the art, potential and limits. In: Zahn R, Kaminski MA, Labeyrie L, Pederson TF, editors. Carbon cycling in the glacial ocean: constraints on the ocean's role in global change. Berlin: Springer-Verlag; 1994. pp. 521–537. [Google Scholar]
- Dale B, Thorsen TA, Fjellså A. Dinoflagellate cysts as indicators of cultural eutrophication in the Oslofjord, Norway. Estuar. Coast. Shelf Sci. 1999;48:371–382. [Google Scholar]
- Danovaro R, Gambi C, Dell'Anno A, Corinaldesi C, Fraschetti S, Vanreusel A, et al. Exponential decline of deep-sea ecosystem functioning linked to benthic biodiversity loss. Curr. Biol. 2008;18:1–8. doi: 10.1016/j.cub.2007.11.056. [DOI] [PubMed] [Google Scholar]
- Debenay JP, Fernandez JM. Benthic foraminifera records of complex anthropogenic environmental changes combined with geochemical data in a tropical bay of New Caledonia (SW Pacific) Mar. Pollut. Bull. 2009;59:311–322. doi: 10.1016/j.marpolbul.2009.09.014. [DOI] [PubMed] [Google Scholar]
- Diaz RJ, Rosenberg R. Spreading dead zones and consequences for marine ecosystems. Science. 2008;321:926–929. doi: 10.1126/science.1156401. [DOI] [PubMed] [Google Scholar]
- Duarte CM, Conley DJ, Carstensen J, Sanchez-Camacho M. Return to neverland: shifting baselines affect eutrophication restoration targets. Estuaries Coasts. 2009;32:29–36. [Google Scholar]
- Dwyer GS, Cronin TM, Baker PA. Trace elements in marine ostracodes. In: Holmes JA, Chivas AR, editors. The Ostracoda: applications in quaternary research. Washington, DC: American Geophysical Union; 2002. pp. 205–225. [Google Scholar]
- Elberling B, Knudsen KL, Kristensen PH, Asmund G. Applying foraminiferal stratigraphy as a biomarker for heavy metal contamination and mining impact in a fiord in West Greenland. Mar. Environ. Res. 2003;55:235–256. doi: 10.1016/s0141-1136(02)00219-2. [DOI] [PubMed] [Google Scholar]
- Ellegaard M, Clarke AL, Reuss N, Drew S, Weckström K, Juggins S, et al. Multi-proxy evidence of long-term changes in ecosystem structure in a Danish marine estuary, linked to increased nutrient loading. Estuar. Coast. Shelf Sci. 2006;68:567–578. [Google Scholar]
- Fabry VJ, Seibel BA, Feely RA, Orr JC. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES J. Mar. Sci. 2008;65:414–432. [Google Scholar]
- Field DB, Baumgartner TR, Charles CD, Ferreira-Bartrina V, Ohman MD. Planktonic foraminifera of the California Current reflect 20th-century warming. Science. 2006;311:63–66. doi: 10.1126/science.1116220. [DOI] [PubMed] [Google Scholar]
- Field DB, Baumgartner TR, Ferreira V, Gutiérrez D, Lozano-Montes H, Salvatteci R. Variability from scales in marine sediments and other historical records. In: Checkley D, Alheit J, Oozeki Y, Roy C, et al., editors. Climate change and small pelagic fish. Cambridge: Cambridge University Press; 2009. pp. 45–63. [Google Scholar]
- Finney BP, Alheit J, Emeis KC, Field DB, Gutiérrez D, Struck U. Paleoecological studies on variability in marine fish populations: a long-term perspective on the impacts of climatic change on marine ecosystems. J. Mar. Syst. 2010;79:316–326. [Google Scholar]
- Fujita K, Hikami M, Suzuki A, Kuroyanagi A, Sakai K, Kawahata H, et al. Effects of ocean acidification on calcification of symbiont-bearing reef foraminifers. Biogeosciences. 2011;8:2089–2098. [Google Scholar]
- Fukami T, Morin PJ. Productivity-biodiversity relationships depend on the history of community assembly. Nature. 2003;424:423–426. doi: 10.1038/nature01785. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Regalado ML, Ruiz F, Baceta JI, González-Regalado E, Munoz JM. Total benthic foraminifera assemblages in the southwestern Spanish estuaries. Geobios. 2001;34:39–51. [Google Scholar]
- González-Regalado ML, Ruiz F, Borrego J. Evolución de la distribución de los foraminíferos bentónicos en un medio contaminado: el estuario del río Odiel (Huelva, S.O. de España) Revista Española de Paleontología. 1996;11:1–10. [Google Scholar]
- Gooday AJ, Jorissen F, Levin LA, Middelburg JJ, Naqvi SWA, Rabalais NN, et al. Historical records of coastal eutrophication-induced hypoxia. Biogeosciences. 2009;6:1707–1745. [Google Scholar]
- Gorham E, Brush GS, Graumlich LJ, Rosenzweig ML, Johnson AH. The value of paleoecology as an aid to monitoring ecosystems and landscapes, chiefly with reference to North America. Environ. Rev. 2001;9:99–126. [Google Scholar]
- Grenfell HR, Hayward BW, Horrocks M. Foraminiferal record of ecological impact of deforestation and oyster farms, Mahurangi Harbour, New Zealand. Mar. Freshw. Res. 2007;58:475–491. [Google Scholar]
- Grönlund T. Diatoms in surface sediments of the Gotland Basin in the Baltic Sea. Hydrobiologia. 1993;269–270:235–242. [Google Scholar]
- Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, D'Agrosa C, et al. A global map of human impact on marine ecosystems. Science. 2008;319:948–952. doi: 10.1126/science.1149345. [DOI] [PubMed] [Google Scholar]
- Harley CDG. Climate change, keystone predation, and biodiversity loss. Science. 2011;334:1124–1127. doi: 10.1126/science.1210199. [DOI] [PubMed] [Google Scholar]
- Hass HC. The benthic foraminiferal response to late Holocene climate change over northern Europe. Grzybowsky Found. Spec. Publ. 1997a;5:199–216. [Google Scholar]
- Hass HC. Recent and subrecent agglutinated foraminifera in four box cores from the Skagerrak (NE North Sea) Grzybowsky Found. Spec. Publ. 1997b;5:217–226. [Google Scholar]
- Hawkins SJ, Gibbs PE, Pope ND, Burt GR, Chesman BS, Bray S, et al. Recovery of polluted ecosystems: the case for long-term studies. Mar. Environ. Res. 2002;54:215–222. doi: 10.1016/s0141-1136(02)00117-4. [DOI] [PubMed] [Google Scholar]
- Hayward BW, Grenfell HR, Nicholson K, Parker R, Wilmhurst J, Horrocks M, et al. Foraminiferal record of human impact on intertidal estuarine environments in New Zealand's largest city. Mar. Micropaleontol. 2004;53:37–66. [Google Scholar]
- Hayward BH, Grenfell HR, Sabaa AT, Morley MS, Horrocks M. Effect and timing of increased freshwater runoff into sheltered harbor environments around Auckland City, New Zeland. Estuaries Coasts. 2006;29:165–182. [Google Scholar]
- Hayward BH, Grenfell HR, Sabaa AT, Morley MS. Ecological impact of the introduction to New Zealand of Asian date mussels and cordgrass—The foraminiferal, ostracod and molluscan record. Estuaries Coasts. 2008;31:941–959. [Google Scholar]
- Hoegh-Guldberg O, Bruno JF. The impact of climate change on the world's marine ecosystems. Science. 2010;328:1523–1528. doi: 10.1126/science.1189930. [DOI] [PubMed] [Google Scholar]
- Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, et al. Coral reefs under rapid climate change and ocean acidification. Science. 2007;318:1737–1742. doi: 10.1126/science.1152509. [DOI] [PubMed] [Google Scholar]
- Huettmann F. From Europe to North America into the world and atmosphere: a short review of global footprints and their impacts and predictions. Environmentalist. 2012;32:289–295. [Google Scholar]
- Hunt G, Cronin TM, Roy K. Species–energy relationship in the deep sea: a test using the Quaternary fossil record. Ecol. Lett. 2005;8:739–747. [Google Scholar]
- Ikeya N. 1995. pp. 27–33. Ostracoda in sediment cores from Yokohama Port. Changes in marine organisms and environments at Yokohama Port: research on fossils in sediment core samples, Yokohama Environmental Research Institute, Report no. 11.
- Intergovernmental Panel on Climate Change. 2007. Climate change 2007: the physical science basis. Summary for policymakers ( http://www.ipcc.ch)
- Irabien MJ, Cearreta A, Leorri E, Gómez J, Viguri J. A 130 year record of pollution in the Suances estuary (southern Bay of Biscay): implications for environmental management. Mar. Pollut. Bull. 2008;56:1719–1727. doi: 10.1016/j.marpolbul.2008.07.006. [DOI] [PubMed] [Google Scholar]
- Irizuki T, Nakamura Y, Takayasu K, Sakai S. Faunal changes in Ostracoda (Crustacea) in Lake Nakaumi, southwest Japan, over the last 40 years. Geosci. Rep. Shimane Univ. 2003;22:149–160. [Google Scholar]
- Irwin A, Mallegraeff GM, McMinn A, Harrison J, Heijnis H. Cyst and radionuclide evidence demonstrate historic Gymnodinium catenatum dinoflagellate populations in Manukau and Hokianga Harbours, New Zealand. Harmful Algae. 2003;2:61–74. [Google Scholar]
- Izuka SK, Kaesler RL. Biostratinomy of ostracode assemblages from a small reef flat in Maunalua Bay, Oahu, Hawaii. J. Paleontol. 1986;60:347–360. [Google Scholar]
- Jablonski D, Roy K, Valentine JW. Evolutionary macroecology and the fossil record. In: Blackburn TM, Gaston KJ, editors. Macroecology: concepts and consequences. Oxford: Blackwell Publishing; 2003. pp. 368–390. [Google Scholar]
- Jablonski D, Roy K, Valentine JW. Out of the tropics: evolutionary dynamics of the latitudinal diversity gradient. Science. 2006;314:102–106. doi: 10.1126/science.1130880. [DOI] [PubMed] [Google Scholar]
- Jackson JBC. What was natural in the coastal oceans? Proc. Natl. Acad. Sci. USA. 2001;98:5411–5418. doi: 10.1073/pnas.091092898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson JBC. Ecological extinction and evolution in the brave new ocean. Proc. Natl. Acad. Sci. USA. 2008;105:11458–11465. doi: 10.1073/pnas.0802812105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson JBC, Kirby MX, Berger WH, Bjorndal KA, Botsford LW, Bourque BJ, et al. Historical overfishing and the recent collapse of coastal ecosystems. Science. 2001;293:629–638. doi: 10.1126/science.1059199. [DOI] [PubMed] [Google Scholar]
- Jankowska D, Witak M, Huszczo D. Paleoecological changes of the Vistula Lagoon in the last 7,000 YBP based on diatom flora. Oceanol. Hydrobiol. Stud. 2005;34:109–129. [Google Scholar]
- Karlsen AW, Cronin TM, Ishman SE, Willard DA, Kerhin R, Holmes CW, et al. Historical trends in Chesapeake Bay dissolved oxygen based on benthic foraminifera from sediment cores. Estuaries. 2000;23:488–508. [Google Scholar]
- Katsuki K, Miyamoto Y, Yamada K, Takata H, Yamaguchi K, Nakayama D, et al. Eutrophication-induced changes in Lake Nakaumi, southwest Japan. J. Paleolimnol. 2008;40:1115–1125. [Google Scholar]
- Katsuki K, Seto K, Nomura R, Maekawa K, Khim BK. Effect of human activity on Lake Saroma (Japan) during the past 150 years: evidence by variation of diatom assemblages. Estuar. Coast. Shelf Sci. 2009;81:215–224. [Google Scholar]
- Kauppila P, Weckström K, Vaalgamaa S, Korhola A, Pitkänen H, Reuss N, et al. Tracing pollution and recovery using sediments in an urban estuary, northern Baltic Sea: are we far from ecological reference conditions? Mar. Ecol. Prog. Ser. 2005;290:35–53. [Google Scholar]
- Kemp WM, Boynton WR, Adolf JE, Boesch DF, Boicourt WC, Brush G, et al. Eutrophication of Chesapeake Bay: historical trends and ecological interactions. Mar. Ecol. Prog. Ser. 2005;303:1–29. [Google Scholar]
- Kidwell SM. Preservation of species abundance in marine death assemblages. Science. 2001;294:1091–1094. doi: 10.1126/science.1064539. [DOI] [PubMed] [Google Scholar]
- Kidwell SM. Discordance between living and death assemblages as evidence for anthropogenic ecological change. Proc. Natl. Acad. Sci. USA. 2007;104:17701–17706. doi: 10.1073/pnas.0707194104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kidwell SM. Evaluating human modification of shallow marine ecosystems: mismatch in composition of molluscan living and time-averaged death assemblages. Paleontol. Soc. Pap. 2009;15:113–139. [Google Scholar]
- Kim HS, Matsuoka K. Process of eutrophication estimated by dinoflagellate cyst assemblages in Omura Bay, Kyushu, West Japan. Bull. Plankton Soc. Jpn. 1998;45:133–147. [Google Scholar]
- Kim SY, Moon CH, Cho HJ, Lim DI. Dinoflagellate cysts in coastal sediments as indicators of eutrophication: a case of Gwangyang Bay, South Sea of Korea. Estuaries Coasts. 2009;32:1225–1233. [Google Scholar]
- Knowlton N, Jackson JBC. Shifting baselines, local impacts, and global change on coral reefs. PLoS Biol. 2008;6:215–220. doi: 10.1371/journal.pbio.0060054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korhola A, Blom T. Marked early 20th century pollution and the subsequent recovery of Töölö Bay, central Helsinki, as indicated by subfossil diatom assemblage changes. Hydrobiologia. 1996;341:169–179. [Google Scholar]
- Kornicker LS. A seasonal study of living Ostracoda in a Texas bay (Redfish Bay) adjoining the Gulf of Mexico. Pubblicazione della Stazione Zoologica di Napoli. 1964;33(supplemento):45–60. [Google Scholar]
- Köster D, Lichter J, Lea PD, Nurse A. Historical eutrophication in a river-estuary complex in mid-coast Maine. Ecol. Appl. 2007;17:765–778. doi: 10.1890/06-0815. [DOI] [PubMed] [Google Scholar]
- Kuroyanagi A, Kawahata H, Suzuki A, Fujita K, Irie T. Impacts of ocean acidification on large benthic foraminifers: results from laboratory experiments. Mar. Micropaleontol. 2009;73:190–195. [Google Scholar]
- Leavitt PR, Hodgson DA. Sedimentary pigments. Dev. Paleoenviron. Res. 2002;3:295–325. [Google Scholar]
- Leśniewska M, Witak M. Holocene diatom biostratigraphy of the SW Gulf of Gdańsk, Southern Baltic Sea (part III) Oceanol. Hydrobiol. Stud. 2008;37:35–52. [Google Scholar]
- Liu DY, Sun J, Zhang J, Liu GS. Response of the diatom flora in Jiaozhou Bay, China to environmental changes during the last century. Mar. Micropaleontol. 2008;66:279–290. [Google Scholar]
- Lotze HK, Milewski I. Two centuries of multiple human impacts and successive changes in a North Atlantic food web. Ecol. Appl. 2004;14:1428–1447. [Google Scholar]
- Lotze HK, Reise K, Worm B, Busch J, Van Beusekom M, Ehlers A, et al. Human transformations of the Wadden Sea ecosystem through time: a synthesis. Helgol. Mar. Res. 2005;59:84–95. [Google Scholar]
- Lotze HK, Lenihan HS, Bourque BJ, Bradbury RH, Cooke RG, Kay MC, et al. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science. 2006;312:1806–1809. doi: 10.1126/science.1128035. [DOI] [PubMed] [Google Scholar]
- de Mahiques MM, Burone L, Figueira RCL, Capellari AA, de Oliveira Lavenére-Wanderley B, Rogacheski CE, et al. Anthropogenic influences in a lagoonal environment: a multiproxy approach at the valo grande mouth, Cananéia-Iguape system (SE Brazil) Braz. J. Oceanogr. 2009;57:325–337. [Google Scholar]
- Mann ME, Zhang ZH, Rutherford S, Bradley RS, Hughes MK, Shindell D, et al. Global signatures and dynamical origins of the Little Ice Age and Medieval Climate Anomaly. Science. 2009;326:1256–1260. doi: 10.1126/science.1177303. [DOI] [PubMed] [Google Scholar]
- Marret F, Mudie P, Aksu A, Hiscott RN. A Holocene dinocyst record of a two-step transformation of the Neoeuxinian brackish water lake into the Black Sea. Quat. Int. 2009;197:72–86. [Google Scholar]
- Martin RE. Taphonomy: a process approach. Cambridge: Cambridge University Press; 1999. [Google Scholar]
- Martinez-Colon M, Hallock P, Green-Ruiz C. Potentially toxic elements and strategies for use of shallow water benthic foraminifers as bio-indicators: a review. J. Foramin. Res. 2009;39:278–299. [Google Scholar]
- Matsumoto E. Research on coastal marine pollution. Chishitsu News. 1981;319:52–58. [Google Scholar]
- Matsumoto E. Environmental changes recorded in sediments of coastal marine zone closed by big city. Mem. Geol. Soc. Jpn. 1983;23:91–95. [Google Scholar]
- Matsumoto E, Saito Y. Environmental changes of Tokyo Bay by man in comparison with environmental changes by nature during geological history. Bull. Geol. Surv. Jpn. 1984;35:243–260. [Google Scholar]
- Matsuoka K. 1995. pp. 45–61. Dinoflagellate cyst assemblage in core samples of Yokohama Port St. 1. Changes in marine organisms and environments at Yokohama Port: research on fossils in sediment core samples, Yokohama Environmental Research Institute, Report no. 116.
- Matsuoka K. Eutrophication process recorded in dinoflagellate cyst assemblages — a case of Yokohama Port, Tokyo Bay, Japan. Sci. Total Environ. 1999;231:17–35. doi: 10.1016/s0048-9697(99)00087-x. [DOI] [PubMed] [Google Scholar]
- Matsuoka K. Further evidence for a marine dinoflagellate cyst as an indicator of eutrophication in Yokohama Port, Tokyo Bay, Japan: comments on a discussion by B. Dale. Sci. Total Environ. 2001;264:221–233. doi: 10.1016/s0048-9697(00)00718-x. [DOI] [PubMed] [Google Scholar]
- Matsuoka K. Change in the aquatic environment of Isahaya Bay, Ariake Sound, West Japan: from the view point of dinoflagellate cyst assemblage. Bull. Coast. Oceanogr. 2004;42:55–59. [Google Scholar]
- Matsuoka K, Kim HS. Process of eutrophication in enclosed seas recorded in dinoflagellate cyst assemblages and sediments – the case in Nagasaki Bay, west Japan. Fossils (Kaseki) 1999;66:1–15. [Google Scholar]
- Matsuoka K, Shin HH. Environmental changes in the inner part of Ariake Sound, west Japan recorded in dinoflagellate cyst assemblages. In: Ishimatsu A, Lie H-J, editors. Coastal environmental and ecosystem issues of the East China sea. Tokyo and Nagasaki: TERRAPUB and Nagasaki University; 2010. pp. 111–120. [Google Scholar]
- Matthews A, Grenfell HR, Hayward BW, Horrocks M. Foraminiferal record of sewage outfall impacts on the inner Manukau Harbour, Auckland, New Zealand. NZ J. Mar. Freshw. Res. 2005;39:193–215. [Google Scholar]
- McGann M. High-resolution foraminiferal, isotopic, and trace element records from Holocene estuarine deposits of San Francisco Bay, California. J. Coastal Res. 2008;24:1092–1109. [Google Scholar]
- McGann M. Review of impacts of contaminated sediment on microfaunal communities in the Southern California Bight. Geol. Soc. Am. Spec. Pap. 2009;454:413–455. [Google Scholar]
- McGann M, Sloan D. Recent introduction of the foraminifer Trochammina hadai Uchio into San Francisco Bay, California, USA. Mar. Micropaleontol. 1996;28:1–3. [Google Scholar]
- McGann M, Sloan D, Cohen AN. Invasion by a Japanese marine microorganism in western North America. Hydrobiologia. 2000;421:25–30. [Google Scholar]
- McGann M, Alexander CR, Bay SM. Response of benthic foraminifers to sewage discharge and remediation in Santa Monica Bay, California. Mar. Environ. Res. 2003;56:299–342. doi: 10.1016/S0141-1136(02)00336-7. [DOI] [PubMed] [Google Scholar]
- McMinn A, Hallegraeff GM, Thomson P, Jenkinson AV, Heijnis H. Cyst and radionucleotide evidence for the recent introduction of the toxic dinoflagellate Gymnodinium catenatum into Tasmanian waters. Mar. Ecol. Prog. Ser. 1997;161:165–172. [Google Scholar]
- Miller U, Risberg J. Environmental changes, mainly eutrophication, as recorded by fossil siliceous micro-algae in two cores from the uppermost sediments of the north-western Baltic. Beiheft zur Nova Hedwigia. 1990;100:237–253. [Google Scholar]
- Miller AAL, Mudie PJ, Scott DB. Holocene history of Bedford Basin, Nova Scotia: foraminifera, dinoflagellate, and pollen records. Can. J. Earth Sci. 1982;19:2342–2367. [Google Scholar]
- Moodley L, Troelstra SR, Van Weering TCE. Benthic foraminiferal response to environmental change in the Skagerrak, northeastern North Sea. Sarsia. 1993;78:129–139. [Google Scholar]
- Murray JW. When does environmental variability become environmental change? The proxy record of benthic foraminifera. In: Martin RE, editor. Environmental micropaleontology: the application of microfossils to environmental geology. New York: Kluwer Academic/Plenum Publishers; 2000. pp. 7–37. [Google Scholar]
- Nagy J, Alve E. Temporal changes in foraminiferal faunas and impact of pollution in Sandebukta, Oslo Fjord. Mar. Micropaleontol. 1987;12:109–128. [Google Scholar]
- Nomura R, Kawano S. Foraminiferal assemblages response to anthropogenic influence and parallel to decadal sea-level changes over the last 70 years in Lake Kugushi, Fukui Prefecture, southwest Japan. Quat. Int. 2011;230:44–56. [Google Scholar]
- Olli K, Clarke A, Danielsson Å, Aigars J, Conley DJ, Tamminen T. Diatom stratigraphy and long-term dissolved silica concentrations in the Baltic Sea. J. Mar. Syst. 2008;73:284–299. [Google Scholar]
- Olszewski T. Taking advantage of time-averaging. Paleobiology. 1999;25:226–238. [Google Scholar]
- Orr JC, Fabry VJ, Aumont O, Bopp L, Doney SC, Feely RA, et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature. 2005;437:681–686. doi: 10.1038/nature04095. [DOI] [PubMed] [Google Scholar]
- Osterman LE, Poore RZ, Swarzenski PW, Turner RE. Reconstructing a 180 yr record of natural and anthropogenic induced low-oxygen conditions from Louisiana continental shelf sediments. Geology. 2005;33:329–332. [Google Scholar]
- Osterman LE, Poore RZ, Swarzenski PW. The last 1000 years of natural and anthropogenic low-oxygen bottom-water on the Louisiana shelf, Gulf of Mexico. Mar. Micropaleontol. 2008;66:291–303. [Google Scholar]
- Osterman LE, Poore RZ, Swarzenski PW, Senn DB, DiMarco SF. The 20th-century development and expansion of Louisiana shelf hypoxia, Gulf of Mexico. Geo-Mar. Lett. 2009;29:405–414. [Google Scholar]
- Paetzel M, Schrader H. Sewage history in the anoxic sediments of the fjord Nordåsvannet, western Norway (II): the origin of the sedimented organic matter fraction. Nor. Geol. Tidsskr. 1995;75:146–155. [Google Scholar]
- Palumbi SR, Sandifer PA, Allan JD, Beck MW, Fautin DG, Fogarty MJ, et al. Managing for ocean biodiversity to sustain marine ecosystem services. Front. Ecol. Environ. 2009;7:204–211. [Google Scholar]
- Pandolfi JM, Bradbury RH, Sala E, Hughes TP, Bjorndal KA, Cooke RG, et al. Global trajectories of the long-term decline of coral reef ecosystems. Science. 2003;301:955–958. doi: 10.1126/science.1085706. [DOI] [PubMed] [Google Scholar]
- Pandolfi JM, Connolly SR, Marshall DJ, Cohen AL. Projecting coral reef futures under global warming and ocean acidification. Science. 2011;333:418–422. doi: 10.1126/science.1204794. [DOI] [PubMed] [Google Scholar]
- Parsons ML, Dortch Q. Sedimentological evidence of an increase in Pseudo-nitzschia (Bacillariophyceae) abundance in response to coastal eutrophication. Limnol. Oceanogr. 2002;47:551–558. [Google Scholar]
- Pascual A, Rodriguez-Lazaro J, Weber O, Jouanneau JM. Late Holocene pollution in the Gernika estuary (southern Bay of Biscay) evidenced by the study of Foraminifera and Ostracoda. Hydrobiologia. 2002;475:477–491. [Google Scholar]
- Pauly D. Anecdotes and the shifting baseline syndrome of fisheries. Trends Ecol. Evol. 1995;10:430. doi: 10.1016/s0169-5347(00)89171-5. [DOI] [PubMed] [Google Scholar]
- Pauly D, Thia-Eng C. The overfishing of marine resources: socioeconomic background in Southeast Asia. Ambio. 1988;17:200–206. [Google Scholar]
- Pelejero C, Calvo E, Hoegh-Guldberg O. Paleo-perspectives on ocean acidification. Trends Ecol. Evol. 2010;25:332–344. doi: 10.1016/j.tree.2010.02.002. [DOI] [PubMed] [Google Scholar]
- Platon E, Sen Gupta BK, Rabalais NN, Turner RE. Effect of seasonal hypoxia on the benthic foraminiferal community of the Louisiana inner continental shelf: The 20th century record. Mar. Micropaleontol. 2005;54:263–283. [Google Scholar]
- Polovodova I, Schönfeld J. Foraminiferal test abnormalities in the western Baltic Sea. J. Foramin. Res. 2008;38:318–336. [Google Scholar]
- Pospelova V, Chmura GL, Boothman WS, Latimer JS. Dinoflagellate cyst records and human disturbance in two neighboring estuaries, New Bedford Harbor and Apponagansett Bay, Massachusetts (USA) Sci. Total Environ. 2002;298:81–102. doi: 10.1016/s0048-9697(02)00195-x. [DOI] [PubMed] [Google Scholar]
- Puskaric S, Berger GW, Jorissen FJ. Successive appearance of subfossil phytoplankton species in Holocene sediments of the northern Adriatic and its relation to the increased eutrophication pressure. Estuar. Coast. Shelf Sci. 1990;31:177–187. [Google Scholar]
- Rabalais NN, Turner RE, Wiseman WJ., Jr Gulf of Mexico, a.k.a. “The Dead Zone”. Annu. Rev. Ecol. Syst. 2002;33:235–263. [Google Scholar]
- Rabalais NN, Turner RE, Sen Gupta BK, Platon E, Parsons ML. Sediments tell the history of eutrophication and hypoxia in the northern Gulf of Mexico. Ecol. Appl. 2007;17:S129–S143. [Google Scholar]
- Reuss N, Conley DJ, Bianchi TS. Preservation conditions and the use of sediment pigments as a tool for recent ecological reconstruction in four northern European estuaries. Mar. Chem. 2005;95:283–302. [Google Scholar]
- Risdal D. Foraminiferfaunaen i en del sedimentkjerner fra indre Oslofjord. Nor. Geol. Unders. 1963;224:5–90. [Google Scholar]
- Ruiz Muñoz F, González-Regalado ML, Borrego Flores J, Morales JA. The response of ostracod assemblages to recent pollution and sedimentary processes in the Huelva Estuary, SW Spain. Sci. Total Environ. 1997;207:91–103. [Google Scholar]
- Ruiz F, González-Regalado ML, Borrego J, Abad M, Pendón JG. Ostracoda and Foraminifera as short-term tracers of environmental changes in very polluted areas: the Odiel Estuary (SW Spain) Environ. Pollut. 2004;129:49–61. doi: 10.1016/j.envpol.2003.09.024. [DOI] [PubMed] [Google Scholar]
- Ruiz F, Borrego J, González-Regalado ML, López González N, Carro B, Abad M. Impact of millennial mining activities on sediments and microfauna of the Tinto River estuary (SW Spain) Mar. Pollut. Bull. 2008;56:1258–1264. doi: 10.1016/j.marpolbul.2008.04.036. [DOI] [PubMed] [Google Scholar]
- Ruiz F, Borrego J, González-Regalado ML, López-González N, Carro B, Abad M. Interaction between sedimentary processes, historical pollution and microfauna in the Tinto Estuary (SW Spain) Environ. Geol. 2009;58:779–783. [Google Scholar]
- Rusak JA, Leavitt PR, McGowan S, Chen G, Olson O, Wunsam S. Millennial-scale relationships of diatom species richness and production in two prairie lakes. Limnol. Oceanogr. 2004;49:1290–1299. [Google Scholar]
- Ryves D, Clarke AL, Appleby PG, Amsinck SL, Jeppesen E, Landkildehus F, et al. Reconstructing the salinity and environment of the Limfjord and Vejlerne Nature Reserve, Denmark, using a diatom model for brackish lakes and fjords. Can. J. Fish. Aquat. Sci. 2004;61:1988–2006. [Google Scholar]
- Sangiorgi F, Donders TH. Reconstructing 150 years of eutrophication in the north-western Adriatic Sea (Italy) using dinoflagellate cysts, pollen and spores. Estuar. Coast. Shelf Sci. 2004;60:69–79. [Google Scholar]
- Sato H. 1995. pp. 63–76. Diatom assemblages in core samples from Yokohama Port. Changes in marine organisms and environments at Yokohama Port: research on fossils in sediment core samples, Yokohama Environmental Research Institute, Report no. 116.
- Saunders KM, Taffs KH. Palaeoecology: a tool to improve the management of Australian estuaries. J. Environ. Manage. 2009;90:2730–2736. doi: 10.1016/j.jenvman.2009.03.001. [DOI] [PubMed] [Google Scholar]
- Saunders KM, McMinn A, Roberts D, Hodgson DA, Heijnis H. Recent human-induced salinity changes in Ramsar-listed Orielton Lagoon, south-east Tasmania, Australia: a new approach for coastal lagoon conservation and management. Aquat. Conserv. 2007;17:51–70. [Google Scholar]
- Saunders KM, Hodgson DA, Harrison J, McMinn A. Palaeoecological tools for improving the management of coastal ecosystems: a case study from Lake King (Gippsland Lakes) Australia. J. Paleolimnol. 2008;40:33–47. [Google Scholar]
- Savage C, Leavitt PR, Elmgren R. Effects of land use, urbanization, and climate variability on coastal eutrophication in the Baltic Sea. Limnol. Oceanogr. 2010;55:1033–1046. [Google Scholar]
- Schafer CT. Rome, Italy: 1970. pp. 1–14. Pollution and benthonic foraminifera species diversity in Long Island Sound. Food and Agricultural Organization Technical Conference on Marine Pollution and Its Effects on Living Resources and Fishing. [Google Scholar]
- Schafer CT, Collins ES, Smith JN. Relationship of Foraminifera and thecamoebian distributions to sediments contaminated by pulp mill effluent: Saguenay Fiord, Quebec, Canada. Mar. Micropaleontol. 1991;17:255–283. [Google Scholar]
- Schellenberg SA. Marine ostracods. In: Elias SA, editor. Encyclopedia of quaternary science. Amsterdam: Elsevier; 2007. pp. 2046–2062. [Google Scholar]
- Scott DB, Schafer CT, Honig C, Younger DC. Temporal variations of benthic foraminiferal assemblages under or near aquaculture operations: documentation of impact history. J. Foramin. Res. 1995;25:224–235. [Google Scholar]
- Scott DB, Medioli FS, Schafer CT. Monitoring in coastal environments using foraminifera and thecamoebian indicators. Cambridge: Cambridge University Press; 2001. [Google Scholar]
- Scott DB, Tobin R, Williamson M, Medioli FS, Latimer JS, Boothman WA, et al. Pollution monitoring in two north American estuaries: historical reconstructions using benthic foraminifera. J. Foramin. Res. 2005;35:65–82. [Google Scholar]
- Sen Gupta BK, Platon E. Tracking past sedimentary records of oxygen depletion in coastal waters: Use of the AmmoniaElphidium foraminiferal index. J. Coastal Res. 2006;3:1351–1355. [Google Scholar]
- Sen Gupta BK, Turner RE, Rabalais NN. Seasonal oxygen depletion in continental-shelf waters of Louisiana: historical record of benthic foraminifers. Geology. 1996;24:227–230. [Google Scholar]
- Sexton PF, Wilson PA, Pearson PN. Microstructural and geochemical perspectives on planktic foraminiferal preservation: “Glassy'' versus “Frosty''. Geochem. Geophys. Geosyst. 2006;7:Q12P19. doi: 10.1029/2006GC001291. [Google Scholar]
- Shahidul Islam M, Tanaka M. Impacts of pollution on coastal and marine ecosystems including coastal and marine fisheries and approach for management: a review and synthesis. Mar. Pollut. Bull. 2004;48:624–649. doi: 10.1016/j.marpolbul.2003.12.004. [DOI] [PubMed] [Google Scholar]
- Shin HH, Mizushima K, Oh SJ, Park JS, II, Noh H, Iwatani M, et al. Reconstruction of historical nutrient levels in Korean and Japanese coastal areas based on dinoflagellate cyst assemblages. Mar. Pollut. Bull. 2010;60:1243–1258. doi: 10.1016/j.marpolbul.2010.03.019. [DOI] [PubMed] [Google Scholar]
- Smol JP. Conservation biology and environmental change: a paleolimnological perspective. Paleontol. Soc. Pap. 2009;15:25–37. [Google Scholar]
- Spero HJ, Bijma J, Lea DW, Bemis BE. Effect of seawater carbonate concentration on foraminiferal carbon and oxygen isotopes. Nature. 1997;390:497–500. [Google Scholar]
- Spielhagen RF, Werner K, Sorensen SA, Zamelczyk K, Kandiano E, Budeus G, et al. Enhanced modern heat transfer to the Arctic by warm Atlantic Water. Science. 2011;331:450–453. doi: 10.1126/science.1197397. [DOI] [PubMed] [Google Scholar]
- Taffs KH, Farago LJ, Heijnis H, Jacobsen G. A diatom-based Holocene record of human impact from a coastal environment: Tuckean Swamp, eastern Australia. J. Paleolimnol. 2008;39:71–82. [Google Scholar]
- Tanimura Y, Kato M, Shimada C, Matsumoto E. A one-hundred-year succession of planktonic and tychopelagic diatoms from 20th century Tokyo Bay. Bull. Natl. Sci. Mus., Tokyo. Ser. C, Geol. Paleontol. 2003;29:1–8. [Google Scholar]
- Thibodeau B, Mucci A, de Vernal A. Recent eutrophication and consequent hypoxia in the bottom waters of the Lower St. Lawrence Estuary: micropaleontological and geochemical evidence. Mar. Geol. 2006;231:37–50. [Google Scholar]
- Thomas E, Gooday AJ. Cenozoic deep-sea benthic foraminifers: tracers for changes in oceanic productivity? Geology. 1996;24:355–358. [Google Scholar]
- Thomas E, Gapotchenko T, Varekamp JC, Mecray EL, Buchholtz ten Brink MR. Benthic foraminifera and environmental changes in Long Island Sound. J. Coastal Res. 2000;16:641–655. [Google Scholar]
- Thomas E, Abramson I, Varekamp JC, Buchholtz ten Brink MR. Groton, CT: 2004. pp. 87–91. Eutrophication of long island sound as traced by benthic foraminifera. Proceedings 6th Biennual Long Island Sound Meeting, October 2002. [Google Scholar]
- Thorsen TA, Dale B. Dinoflagellate cysts as indicators of pollution and past climate in a Norwegian fjord. Holocene. 1997;7:433–446. [Google Scholar]
- Tikkanen M, Korhola A, Seppä H, Virkanen J. A long-term record of human impacts on an urban ecosystem in the sediments of Töölönlahti Bay in Helsinki, Finland. Environ. Conserv. 1997;24:326–337. [Google Scholar]
- Tomašových A, Kidwell SM. Preservation of spatial and environmental gradients by death assemblages. Paleobiology. 2009;35:119–145. [Google Scholar]
- Toyoda K, Kitazato H. 1995. pp. 11–26. Paleoenvironmental changes of Yokohama Port since 1870 based on benthic foraminiferal fossils. Changes in marine organisms and environments at Yokohama Port: research on fossils in sediment core samples, Yokohama Environmental Research Institute, Report no. 116:
- Tsugeki NK, Ishida S, Urabe J. Sedimentary records of reduction in resting egg production of Daphnia galeata in Lake Biwa during the 20th century: a possible effect of winter warming. J. Paleolimnol. 2009;42:155–165. [Google Scholar]
- Tsujimoto A, Nomura R, Yasuhara M, Yamazaki H, Yoshikawa S. Impact of eutrophication on shallow marine benthic foraminifers over the last 150 years in Osaka Bay, Japan. Mar. Micropaleontol. 2006;60:258–268. [Google Scholar]
- Tsujimoto A, Yasuhara M, Nomura R, Yamazaki H, Sampei Y, Hirose K, et al. Development of modern benthic ecosystems in eutrophic coastal oceans: the foraminiferal record over the last 200 years, Osaka Bay, Japan. Mar. Micropaleontol. 2008;69:225–239. [Google Scholar]
- Tuovinen N, Weckström K, Virtasalo JJ. Assessment of recent eutrophication and climate influence in the Archipelago Sea based on the subfossil diatom record. J. Paleolimnol. 2010;44:95–108. [Google Scholar]
- Uthicke S, Patel F, Ditchburn R. Elevated land runoff after European settlement perturbs persistent foraminiferal assemblages on the Great Barrier Reef. Ecology. 2012;93:111–121. doi: 10.1890/11-0665.1. [DOI] [PubMed] [Google Scholar]
- Wang ZH, Matsuoka K, Qi YZ, Chen JF, Lu SH. Dinoflagellate cyst records in recent sediments from Daya Bay, South China Sea. Phycolog. Res. 2004;52:396–407. [Google Scholar]
- Weckström K. Assessing recent eutrophication in coastal waters of the Gulf of Finland (Baltic Sea) using subfossil diatoms. J. Paleolimnol. 2006;35:571–592. [Google Scholar]
- Weckström K, Juggins S, Korhola A. Quantifying background nutrient concentrations in coastal waters: a case study from an urban embayment of the Baltic Sea. Ambio. 2004;33:324–327. doi: 10.1579/0044-7447-33.6.324. [DOI] [PubMed] [Google Scholar]
- Weckström K, Korhola A, Weckstrom J. Impacts of eutrophication on diatom life forms and species richness in coastal waters of the Baltic Sea. Ambio. 2007;36:155–160. doi: 10.1579/0044-7447(2007)36[155:ioeodl]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Whatley RC. Population structure of ostracods: some general principles for the recognition of palaeoenvironments. In: Colin P, De Deccker JP, Peypouquet JP, editors. Ostracoda in the earth sciences. Amsterdam: Elsevier; 1988. pp. 245–256. [Google Scholar]
- Willard DA, Cronin TM. Paleoecology and ecosystem restoration: case studies from Chesapeake Bay and the Florida Everglades. Front. Ecol. Environ. 2007;5:491–498. [Google Scholar]
- Willard DA, Cronin TM, Verardo S. Late-Holocene climate and ecosystem history from Chesapeake Bay sediment cores, USA. Holocene. 2003;13:201–214. [Google Scholar]
- Witak M, Jankowska D. The Vistula Lagoon evolution based on diatom records. Baltica. 2005;18:68–76. [Google Scholar]
- Witak M, Boryn K, Mayer A. Holocene environmental changes recorded by diatom stratigraphy in the Vistula Lagoon. Oceanol. Hydrobiol. Stud. 2005;34:111–133. [Google Scholar]
- Witak M, Jankowska D, Piekarek-Jankowska H. Holocene diatom biostratigraphy of the SW Gulf of Gdańsk, Southern Baltic Sea (part I) Oceanol. Hydrobiol. Stud. 2006;35:307–329. [Google Scholar]
- Witkowski A, Pampkowiak J. Reconstructing the development of human impact from diatoms and 210Pb sediment dating (the Gulf of Gdansk-southern Baltic Sea) Geographia Polonica. 1995;65:63–78. [Google Scholar]
- Wollenburg JE, Kuhnt W. The response of benthic foraminifers to carbon flux and primary production in the Arctic Ocean. Mar. Micropaleontol. 2000;40:189–231. [Google Scholar]
- Worm B, Barbier EB, Beaumont N, Duffy JE, Folke C, Halpern BS, et al. Impacts of biodiversity loss on ocean ecosystem services. Science. 2006;314:787–790. doi: 10.1126/science.1132294. [DOI] [PubMed] [Google Scholar]
- Worm B, Hilborn R, Baum JK, Branch TA, Collie JS, Costello C, et al. Rebuilding global fisheries. Science. 2009;325:578–585. doi: 10.1126/science.1173146. [DOI] [PubMed] [Google Scholar]
- Yasuhara M, Cronin TM. Climatic influences on deep-sea ostracode (Crustacea) diversity for the last three million years. Ecology. 2008;89:S52–S65. doi: 10.1890/07-1021.1. [DOI] [PubMed] [Google Scholar]
- Yasuhara M, Yamazaki H. The impact of 150 years of anthropogenic pollution on the shallow marine ostracode fauna, Osaka Bay, Japan. Mar. Micropaleontol. 2005;55:63–74. [Google Scholar]
- Yasuhara M, Yamazaki H, Irizuki T, Yoshikawa S. Temporal changes of ostracode assemblages and anthropogenic pollution during the last 100 years, in sediment cores from Hiroshima Bay, Japan. Holocene. 2003;13:527–536. [Google Scholar]
- Yasuhara M, Yamazaki H, Tsujimoto A, Hirose K. The effect of long-term spatiotemporal variations in urbanization-induced eutrophication on a benthic ecosystem, Osaka Bay, Japan. Limnol. Oceanogr. 2007;52:1633–1644. [Google Scholar]
- Yasuhara M, Cronin TM, deMenocal PB, Okahashi H, Linsley BK. Abrupt climate change and collapse of deep-sea ecosystems. Proc. Natl. Acad. Sci. USA. 2008;105:1556–1560. doi: 10.1073/pnas.0705486105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuhara M, Hunt G, Cronin TM, Okahashi H. Temporal latitudinal-gradient dynamics and tropical instability of deep-sea species diversity. Proc. Natl. Acad. Sci. USA. 2009;106:21717–21720. doi: 10.1073/pnas.0910935106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuhara M, Hunt G, Cronin TM, Hokanishi N, Kawahata H, Tsujimoto A, et al. Climatic forcing of Quaternary deep-sea benthic communities in the North Pacific Ocean. Paleobiology. 2012a;38:162–179. [Google Scholar]
- Yasuhara M, Hunt G, Dowsett HJ, Robinson MM, Stoll DK. Latitudinal species diversity gradient of marine zooplankton for the last three million years. Ecol. Lett. 2012b;15:1174–1179. doi: 10.1111/j.1461-0248.2012.01828.x. [DOI] [PubMed] [Google Scholar]
- Yasuhara M, Hunt G, Arrigo G, van Dijken KR, Cronin TM, Wollenburg JE. Patterns and controlling factors of species diversity in the Arctic Ocean. J. Biogeogr. 2012c;39:2081–2088. [Google Scholar]
- Yokose H, Momoshima N, Matsuoka K, Hase Y, Honza E. Environmental assessments of Ariake Bay during the past 100 years based on marine sediments. J. Geogr. 2005;114:1–20. [Google Scholar]
- Zalasiewicz J, Williams M, Smith A, Barry TL, Coe AL, Bown PR, et al. Are we now living in the Anthropocene? GSA Today. 2008;18:4–8. [Google Scholar]
- Zong Y, Chen Z, Innes JB, Chen C, Wang Z, Wang H. Fire and flood management of coastal swamp enabled first rice paddy cultivation in east China. Nature. 2007;449:459–462. doi: 10.1038/nature06135. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.