

Abstract
Background
Living cells constantly sense and adapt to redox shifts by the induction of genes whose products act to maintain the cellular redox environment. In the eukaryote Saccharomyces cerevisiae, while stationary cells possess a degree of constitutive resistance towards oxidants, treatment of exponential phase cultures with sub-lethal stresses can lead to the transient induction of protection against subsequent lethal oxidant conditions. The sensors of oxidative stress and the corresponding transcription factors that activate gene expression under these conditions have not yet been completely identified.
Results
We report the role of SOD1, SOD2 and TPS1 genes (which encode the cytoplasmic Cu/Zn-superoxide dismutase, the mitochondrial Mn-isoform and trehalose-6-phosphate synthase, respectively) in the development of resistance to oxidative stress. In all experimental conditions, the cultures were divided into two parts, one was immediately submitted to severe stress (namely: exposure to H2O2, heat shock or ethanol stress) while the other was initially adapted to 40°C for 60 min. The deficiency in trehalose synthesis did not impair the acquisition of tolerance to H2O2, but this disaccharide played an essential role in tolerance against heat and ethanol stresses. We also verified that the presence of only one Sodp isoform was sufficient to improve cellular resistance to 5 mM H2O2. On the other hand, while the lack of Sod2p caused high cell sensitivity to ethanol and heat shock, the absence of Sod1p seemed to be beneficial to the process of acquisition of tolerance to these adverse conditions. The increase in oxidation-dependent fluorescence of crude extracts of sod1 mutant cells upon incubation at 40°C was approximately 2-fold higher than in sod2 and control strain extracts. Furthermore, in Western blots, we observed that sod mutants showed a different pattern of Hsp104p and Hsp26p expression also different from that in their control strain.
Conclusions
Trehalose seemed not to be essential in the acquisition of tolerance to H2O2 stress, but its absence was strongly felt under water stress conditions such as heat and alcoholic stresses. On the other hand, Sod1p could be involved in the control of ROS production; these reactive molecules could signal the induction of genes implicated within cell tolerance to heat and ethanol. The effects of this deletion needs further investigation.
Background
Since molecular oxygen appeared on earth, certain organisms have developed mechanisms to cope with this molecule. While for these organisms, molecular oxygen turns out to be essential for life, it can also be dangerous for maintenance of cellular viability. The superoxide anion (O2-), hydrogen peroxide (H2O2) and the hydroxyl radical (OH) are the most important reactive oxygen species (ROS) produced by cells. They are frequently formed via metal catalyzed reactions as a result of normal cellular metabolism, such as respiration and β-oxidation of fatty acids, as well as in inflammatory processes and heat, ethanol and chemical stresses [1]. ROS cause oxidative damage of nucleic acids, lipids, proteins, carbohydrates and other cellular components [2]. Moreover, some of the products of oxidative damage to lipids and sugars react readily with proteins and nucleic acids.
In response to the destructive nature of ROS, aerobically growing organisms have evolved multiple defense mechanisms to protect and prevent damage of their cellular components. The primary biological defense against oxidative damage includes protective proteins that remove ROS or sequester metal ions, while secondary defenses consist of enzymes that remove and repair the products of oxidatively damaged components [3]. The anti-oxidant enzyme superoxide dismutase is involved in the conversion of superoxide anion to dioxygen and hydrogen peroxide, which is further degraded by catalase or peroxidases [4]. Saccharomyces cerevisiae, the preferred model for studies of this stress in eukaryotic cells, possesses two ìsoforms, the cytoplasmic Cu/Zn and the mitochondrial Mn superoxide dismutase which have been shown to play an important role in protection of yeast against oxygen toxicity [5]. In addition to higher sensitivity and incapacity of growing on respiratory carbon sources, several metabolic defects have been associated with SOD deficiency, contributing to oxygen-dependent auxotrophy for lysine and methionine [6].
On the other hand, trehalose, one of the major reserve carbohydrates encountered in yeast cells, has been usually correlated with protection against stressful conditions. For example, high intracellular levels of trehalose have been shown to play an important role in stabilizing dry membranes in anhydrobiotic organisms, leading to resistance to dehydration [7]. The trehalose content of exponentially growing cells of Saccharomyces cerevisiae rapidly increases in response to temperature shifts which leads to acquisition of tolerance to heat, osmotic and ethanol stresses [8]. Yeast cells synthesize trehalose in a two-step reaction catalyzed by a complex involving the enzymes trehalose-6-phosphate synthase, encoded by TPS1, and trehalose-6-phosphate phosphatase, encoded by TPS2 and a third component encoded by TSL1, whose function is yet not well understood [9]. Mutants lacking either TPS1 or TPS2 are heat shock sensitive. Furthermore, mutants deleted in neutral trehalase (NTH1), the enzyme which hydrolyses trehalose, maintain acquired thermotolerance for longer periods than do wild type cells [10].
Other candidates as stress protectants are the heat shock proteins (Hsps), which have their synthesis induced in response to heat shock, among other adverse conditions. These proteins act as chaperones and seem to repair damaged proteins produced by stress. In yeast, trehalose and Hsp104p appear to play different biochemical functions: in vitro, evidence suggests that the disaccharide acts as a protectant by stabilizing proteins and preventing their inactivation during adverse conditions, whereas Hsp104p repairs denatured proteins by resolubilizing the insoluble aggregates [11]. It was observed that mutants lacking both Hsp104p and trehalose are extremely thermosensitive even at moderate temperatures, while single mutants display little if any heat shock sensitivity, suggesting that these two factors somehow cooperate towards heat resistance [12]. In a recent review, Singer and Lindquist [13] raised the hypothesis that the stabilization of denaturated proteins by trehalose could interfere with their posterior reactivation and therefore, unless trehalose is rapidly degraded during the recovery from stress, this sugar could impair the ability of Hsps to repair stress-induced damage to cellular proteins swiftly.
While stationary yeast cells possess a significant degree of constitutive resistance towards oxidants, several results have demonstrated that treatment of exponential phase yeast cultures with sub-lethal stress treatments can lead to the transient induction of protection against subsequent lethal oxidant conditions [4]. However, the sensors of oxidative stress and the corresponding transcription factors that activate gene expression under these conditions have not yet been completely identified. With the aim of understanding the role of SOD and TPS1 genes in the development of resistance to oxidative stress, cell viability was analyzed under conditions which promote an increase in the levels of ROS: exposure to H2O2, heat shock and ethanol stress. We also analyzed the expression of two heat shock proteins, Hsp104p and Hsp26p, as well as, the level of intracellular oxidation under these oxidative stress conditions.
Results
Effect of SOD in stress tolerance
Survival of strains carrying SOD deletions was tested under three different stress conditions, which caused an increase of intracellular ROS concentration. Viability of Saccharomyces cerevisiae strains, exposed to H2O2, ethanol or heat stresses was analyzed in cells either grown up to the middle of the first exponential phase at 28°C or harvested at that stage and then exposed to an adaptive heat shock at 40°C/60 min. Sublethal stress conditions, such as a treatment at 40°C cause a set of metabolic alterations, which often lead to acquisition of tolerance against severe conditions. During this adaptive treatment activation of defense systems occurs, such as synthesis of trehalose, heat shock proteins and antioxidant enzymes, which are responsible, respectively, for stabilization of biomolecules, repair of denatured proteins and elimination of ROS [12;14]. According to Fig. 1A, after exposure of cells to a sublethal heat shock at 40°C we observed that the control and the SOD single mutants (sod1 and sod2) became 2–3 times more resistant, while the double mutant, deficient in both Sod1p and Sod2p, continued to show high sensitivity to H2O2. This result lead us to admit that the integrity of only one SOD gene would be sufficient to confer resistance to oxidative conditions caused by 5 mM hydrogen peroxide.
Figure 1.
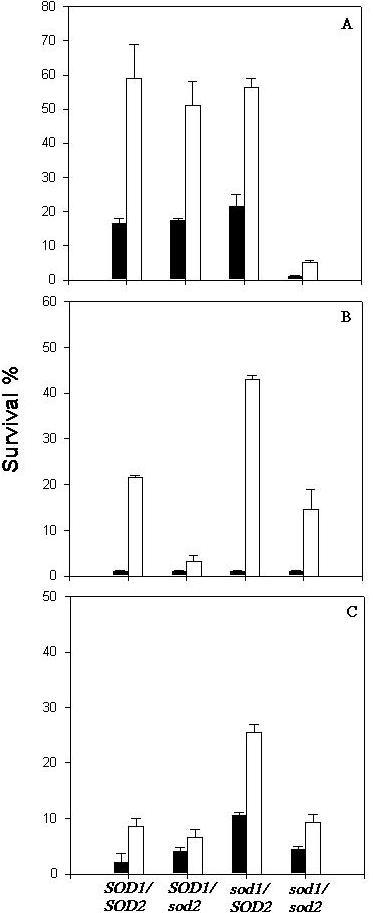
Effect of oxidative stress (A), heat shock (B) and ethanol stress (C) on cell viability. Control and sod mutant cells harvested in first exponential phase were directly stressed (black bars) or previously submitted to a shift of temperature from 28°C to 40°C/1 h and then exposed to stress conditions (white bars). The results represent the mean ± SE of three independent experiments.
Under other stress conditions, all strains, even the control, showed low rates of survival after direct exposure to 51°C for 8 min or 10% ethanol for 24 h, whereas cells pre-exposed to a mild heat treatment developed stress tolerance, except in the case of sod2 cells (Fig. 1B and 1C). In this mutant strain, cellular viability was considerably lower than that observed in pretreated cells of the control strain, suggesting that Sod2p has an important function in the acquisition of resistance to these stresses. On the other hand, the sod1 deletion did not impair the acquisition of tolerance to heat and ethanol after the pretreatment at 40°C. In fact, the survival rate of cells deleted in this isoform, under these conditions, was approximately 2-fold higher than that exhibited by the control strain (Fig. 1B and 1C). In addition, it can be noted in the same figure, that deficiency of both isoforms restored thermal and ethanol tolerance of preheated cells, showing levels of viability close to the control strain. The double deleted cells exhibited rates of survival which were, approximately, the difference between the rates of the sod1 and sod2 cells in all cases.
Intracellular oxidation
In order to measure the level of oxidation produced by shifting the cells from 28°C to 40°C, exponential cells were heated at 40°C for 60 minutes in the presence of the oxidant sensitive probe 2',7'-dichlorofluorescein. This probe is a molecule that can permeate the cell membrane by passive diffusion [15]. Once inside the cell, it becomes susceptible to the attack by radical species, producing a more fluorescent compound [16]. According to Table 1, while crude extracts from control and sod2 cells showed a 2- to 3- fold increase in fluorescence in response to incubation at 40°C, the rise in fluorescence produced by heating in sod1 cells was twice as high. However, the level of oxidation of exponential cells growing at 28°C was increased by a factor of 5 by sod1 deletion as compared to the other strains (results not shown). These results seem to reinforce the idea that an imbalance in the intracellular level of ROS may alter the stress response [17].
Table 1.
Relation between the fluorescence of cells subjected to a heat treatment at 40°C and fluorescence of cells maintained at 28°C.
| Strain | Relative fluorescence |
| EG103 (SOD1/SOD2) | 2.3 ± 0.1 |
| EG110 (sod2) | 2.9 ± 0.3 |
| EG118 (sod1) | 5.4 ± 0.1 |
The results represent the mean ± SE of three independent experiments.
Effect of trehalose in stress tolerance
It has been shown that cells accumulate high levels of trehalose when submitted to a temperature shift from 28°C to 40°C, a condition that enables them to survive subsequent more drastic conditions [8;18]. Although the role of trehalose is well characterized under many stress conditions, up till now the function of this carbohydrate in protective response to oxidative stress is not conclusive [19; 20]. Due to the fact that the control and sod mutant strains under study accumulated very low levels of trehalose after the treatment at 40°C (Table 2), we decided to use a mutant strain deleted in the TPS1 gene (trehalose-6-phosphate synthase) and its parental strain, submitting them to the same oxidative stress conditions caused by 5 mM H2O2. Intracellular levels of trehalose can be seen in Table 2 and, as expected, the TPS1 control strain accumulated high levels of trehalose after mild heat pretreatment, in contrast to the tps1 mutant, which is not capable of synthesizing this carbohydrate. According to the results in Fig. 2, the tps1 deficiency did not impair the acquisition of tolerance to 5 mM H2O2, indicating that the disaccharide is not involved in the mechanism of protection against the damage caused by this type of stress. Furthermore, the tps1 mutant is shown to be as resistant as the SOD control strain (as shown in Fig. 1) and was even more tolerant than its parental strain JT2700, which accumulated considerable amounts of trehalose in response to the adaptive heat treatment. It is important to mention that both the tps1 mutant and its control strain showed similar levels of SOD activity (measured according to McCord and Fridovich [21]) as the SOD control strain (results not shown).
Table 2.
Intracellular levels of trehalose in cells growing exponentially on glucose subjected or not to a heat treatment at 40°C for 1 hr.
| Trehalose (mg/g cell) | ||
| Strain | 28°C | 40°C |
| EG103 (SOD1/SOD2) | 2.6 ± 0.5 | 12.0 ± 0.8 |
| EG110 (sod2) | 2.1 ± 0.3 | 8.8 ± 0.4 |
| EG118 (sod1) | 3.1 ± 0.6 | 9.6 ± 0.3 |
| EG133 (sod1/sod2) | 2.7 ± 0.4 | 9.8 ± 0.2 |
| JT2700 (TPS1) | 1.5 ± 0.5 | 24.7 ± 0,7 |
| JT9060 (tps1) | 0.8 ± 0.3 | 1.0 ± 0.2 |
The results represent the mean ± SE of three independent experiments.
Figure 2.
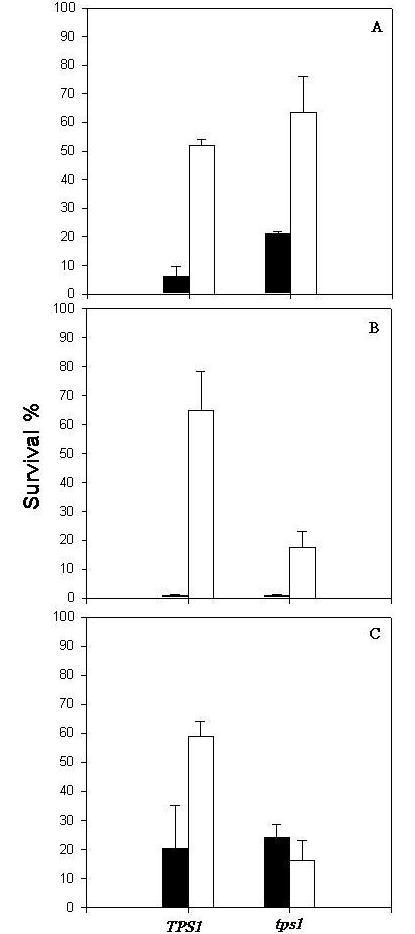
Effect of oxidative stress (A), heat shock (B) and ethanol stress (C) on cell viability. Control and tps1 mutant cells harvested in first exponential phase were directly stressed (black bars) or were previously submitted to a mild heat shock at 40°C/1 h and them exposed to stress conditions (white bars). The results represent the mean ± SE of three independent experiments.
On the other hand, trehalose, as expected, proved to be required for maintenance of cellular viability during heat and ethanol stresses. Induction of trehalose synthesis by the preheating treatment caused an enhancement of about 3.5-fold of survival of the control strain as compared to the tps1 mutant cells submitted to severe heat shock and permitted the acquisition of tolerance to ethanol stress only by the control strain JT2700 (Fig. 2B and 2C). In addition, tps1 mutant cells showed similar thermo and alcoholic tolerance rates to the SOD strains, which also show low levels of intracellular trehalose. These results indicate that trehalose is essential to protect yeast cells against heat and ethanol stress conditions, it seems, however, not to be recquired for protection against oxidative damage.
Hsps expression
The heat shock protein response is induced by heat shock and also by a large variety of different chemicals [22; 23]. Based on these findings, we decided to investigate the relationship between the expression of two Hsps, Hsp104p and Hsp26p and the acquisition of tolerance to H2O2, using cells impaired in the mechanism of antioxidant defense. Furthermore the results obtained might explain the anomalous behavior shown by the sod1 mutant (Fig. 1B and 1C). Cell-free extracts were prepared from exponentially growing cells subjected to oxidative stress, previously adapted or not at 40°C/60 min. The proteins were separated using SDS-PAGE (data not shown) and then transferred to a nitrocellulose membrane in order to test them with specific anti-Hsp104p and anti-Hsp26p antibodies.
As seen in Fig. 3, the control strains, SOD1/SOD2 and TPS1, and the tps1 mutant presented a similar pattern of induction of Hsp expression. While the induction of Hsp26p synthesis was induced in response to a mild heat shock treatment performed at 40°C, the expression of this Hsp was not increased when heat adapted cells were exposed to hydrogen peroxide. The absence of expression of Hsp26p at 28°C when 5 mM H2O2 was added, in contrast to the data published in the literature [22; 24], could be explained by the fact that under our conditions, cells were stressed with a high dose of hydrogen peroxide as compared to the 0.4 mM used in the reference. In our drastic environment, the cells might not be capable of developing a satisfactory stress response. However, we were able to identify the induction of expression of Hsp104p in exponentially growing cells submitted directly to 5 mM H2O2, and furthermore, an enhanced induction when cells were pretreated by a mild heat shock followed or not by an oxidative stress, which demonstrates that our experimental conditions did not lead to cell death.
Figure 3.
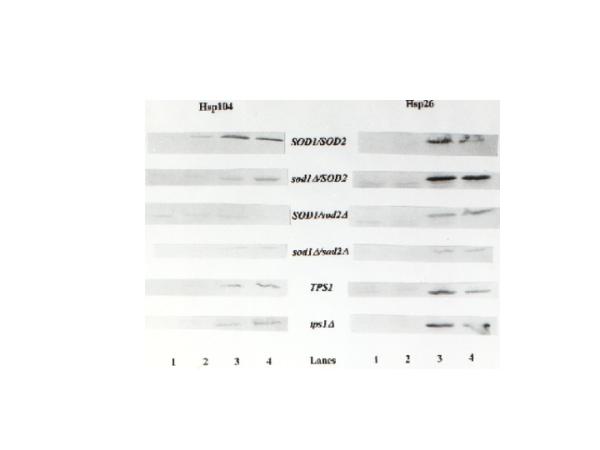
Induction of Hsp104 and Hsp26 expression in SOD and TPS1 strains. Total cellular proteins from strains EG103 (control strain), EG110 (sod2), EG118 (sod1), EG133 (sod1/sod2), JT2700 (control strain) and JT9060 (tps1) were analyzed by Western blot with anti-Hsp104 and anti-Hsp26 protein antibody. Lane1- extracts from cultures at exponential growth on glucose; Lane2- extracts from cultures at exponential growth on glucose directly submitted to 5 mM H2O2; Lane3- extracts from cultures exposed to a mild heat treatment at 40°C/1 h and Lane4- extract from cultures pre-exposed to 40°/1 h and then submitted to a lethal treatment with 5 mM H2O2. Each lane contain the same quantity of protein (50 μg of protein).
Curiously, according to Fig. 3 the SOD deletions affected the Hsp response. Mutants deficient in SOD genes showed a reduced level of Hsp104p synthesis when compared with their control strain. In relation to Hsp26p, while the sod2 and sod1/sod2 mutants presented a reduced level of Hsp expression, the strain lacking Sod1p showed an enhanced level of synthesis in response to stress. Nevertheless the H2O2 sensitive SOD double mutant revealed a different pattern of induction of Hsp104p expression and its synthesis was only induced in cells subjected to the adaptive heat treatment at 40°C. It is important to note that deficiency of the TPS1 gene did not affect the pattern of Hsp expression, indicating that the anomalous results presented by the sod strains are a consequence of a disturbance in the redox status of the cell.
Discussion
Aerobic organisms have to maintain a reduced cellular redox environment in the face of the pro-oxidative conditions characteristic of aerobic life. Besides the pernicious effect of an oxidative atmosphere, stress conditions, such as exposure to heavy metals, heat shock and alcoholic stress, are able to induce oxidative damage in cellular components by increasing ROS synthesis [2;14]. Living cells constantly sense and adapt to such redox shifts by the induction of genes whose products act to maintain the cellular redox environment [24]. From microrganisms to man, a primary defense against oxygen toxicity involves one or another form of the enzyme superoxide dismutase [25]. Null mutants of superoxide dismutase in Saccharomyces cerevisiae are associated with several biochemical defects, indicating that eukaryotic SOD genes may protect numerous metabolic enzymes against oxygen-induced damage [5;26].
Using Saccharomyces cerevisiae strains containing specific sod deletions, we verified that the Sodp isoforms seem to play different roles in the process of acquisition of tolerance to oxidative stress depending on the condition used to induce the synthesis of ROS. In the case of exposition to 5 mM H2O2, the presence of only one Sodp isoform was enough to induce cellular resistance. Considering the importance of Sodp in protecting cells exposed to an oxidative stress, these results could be explained by the fact that a deficiency in either SOD1 or SOD2 in Saccharomyces cerevisiae cells is overcome by an increase in the remaining Sodp activity when compared with the control strain [27]. Results obtained in this work with a SOD null mutant are consistent with this hypothesis, since 100% of exponentially growing cells were unable to form colonies after exposure to 5 mM H2O2 and only 3% of the heat adapted cells remained viable.
In relation to heat and ethanol stresses, only the single mutation sod2 prevented the acquisition of cell resistance against these adverse conditions. Heat and ethanol stresses have been associated with oxidative lesions by increasing the synthesis of oxygen free radicals. While a rise in temperature enhances O2 uptake, leading to an increase of superoxide reactivity that serves as stimulant for ROS production by the Fenton reaction and by polyamine oxidation [14;28], many of the toxic effects caused by ethanol seem to be due to the acetaldehyde produced during its metabolism [27]. Of the enzymatic defenses induced to counteract this increased ROS production, the enzyme superoxide dismutase is particularly important. Although a sublethal heat shock does not alter the activity of cytoplasmic Sod1p, this adaptive treatment increases the activity of the mitochondrial isoform Sod2p [27; 29]. By the dismutation of superoxide produced in excess within mitochondria of yeast cells under heat and ethanol stress conditions, Sod2p could prevent its reaction with biomolecules inside the mitochondria protecting them from oxidative damage. The H2O2 produced by the dismutation of superoxide catalyzed by Sod2p would be decomposed in a next step by cytochrome c peroxidase inside the mitochondria. The importance of the mitochondrial Sodp isoform is further reinforced by the increased ethanol tolerance of respiratory deficient petites, cells that have lost the capacity of producing ROS by the respiratory chain [14].
In contrast to the behavior of the sod2 mutant strain, Sod1p deficiency was beneficial for the development of cellular resistance against heat and ethanol stresses: sod1 cells acquired about 2-fold more tolerance than the control strain. At first sight these results seems to confirm the importance of Sod2p activity, since as previously mentioned, the deficiency in Sod1p is overcome by an almost 4-fold increase in the mitochondrial isoform activity [27]. However, the double mutant sod1/sod2 presented rates of survival after these stresses similar to those exhibited by the parental strain. A possible explanation for the intriguing behavior of sod1 cells would be that the deficiency in this isoform could lead to an imbalance of intracellular ROS production, generating a signal that triggers the transcriptional induction of genes whose products could be beneficial for acquisition of tolerance to heat and alcoholic stresses. Thus a cell may respond to a stimulus by generating ROS that diffuse to a target, react, and thereby transduce the signal. By using an oxidant-sensitive fluorescent probe, we demonstrated a heat-dependent intracellular oxidation. Furthermore, oxidative fluorescence induced by treatment at 40°C was increased by approximately 2-fold in sod1 cells compared to the control strain and to sod2 cells. In addition, we verified that a deficiency in SOD genes affected the transcriptional induction of Hsp genes, suggesting that this effect could be a consequence of an imbalance of intracellular ROS concentration. Many results from the literature support the idea that a redox perturbation of the cellular environment can activate the expression of critical stress responsive genes by the oxidation of glutathione, formation of non-native protein disulfide bonds and by the production of ROS that act as secondary messengers in signaling pathways in higher organisms [17; 30; 31]. It is important to keep in mind that despite the fact that some stress agents can induce similar protection systems, the responses produced by the cells to the various stress agents are not identical.
Another protection factor investigated, which could be implicated with cell tolerance against oxidative stress, was the disaccharide trehalose. According to the results obtained, trehalose seems not to be involved in protection against the damage caused by exposition of cells to 5 mM H2O2. In this type of stress, cells could develop tolerance in response to an adaptive treatment independently of their levels of trehalose. Recently, published data have shown that TPS1 is induced by a mild H2O2 treatment (0.4 mM H2O2), however, curiously, without trehalose accumulation [24]. These results suggest the existence of a recycled flux of this disaccharide, with the purpose of diverting carbohydrate metabolism to the generation of NADPH, the most important cellular reducing power [24]. On the other hand, acquisition of thermo and alcoholic tolerance seems to be dramatically affected by the capacity of cells to synthesize trehalose. We have previously shown that trehalose performs a protecting role under circumstances that involve a shock or an absolute water stress, such as heat and ethanolic shocks and dehydration [8;32]. This seems not be the case of the oxidative stress caused by exposition to 5 mM H2O2. Other factors are certainly involved in protecting cellular components during oxidation.
Expression of two Hsps, Hsp104p and Hsp26p, during oxidative stress conditions using specific antibodies was analyzed. We observed the induction of expression of both Hsp26p and Hsp104p in response to a moderate heat treatment, indicating that these Hsps could be involved with the adaptive response mechanism to protect cells from oxidative threat. Since some Hsps play essential roles in growth and metabolism, under non stressed conditions, by assisting protein biogenesis, oligomer assembly, traffic between organelles and selective proteolysis, we could speculate that these Hsps, besides their protective functions, might help to adapt metabolism to the needs of the oxidative stress response. The induction of synthesis of Hsp104p could also be seen when non adapted cells were directly exposed to 5 mM H2O2, although this pattern of expression was not observed with the SOD null mutant, which demonstrated an increased sensitivity to oxidative stress. These results seem to indicate that the expression of Hsp104p in cells at 28°C/5 mM H2O2 might be required for the recovery of certain damaged proteins produced by the cells under oxidative stress. Again, the sod1 mutant showed a different behavior when the expression of Hsp26p was considered.
The following conclusions can be drawn from the above results:
1) In cells of S. cerevisiae, acquisition of tolerance to oxidative stress caused by 5 mM H2O2 was not associated with intracellular levels of trehalose, although this disaccharide seemed to be very important in the development of cell resistance during stresses that involve loss of water, like heating and exposition to 10% ethanol.
2) The presence of only one Sodp isoform, either mitochondrial (Sod2p) or cytoplasmic (Sod1p), was sufficient for the mechanism of acquisition of tolerance to oxidative stress; the deletion of both Sodp isoforms caused the loss of the capacity of the cell to acquire tolerance to 5 mM H2O2.
3) While Sod2p activity seemed to play an important role in the acquisition of heat and ethanol tolerance, the deficiency of Sod1p appeared to be beneficial for the process of protection against these forms of stress.
4) The sod1 mutant showed a high level of intracellular oxidation, higher than control or sod2 strains, which could be associated with the increased capacity of acquisition of tolerance against heat and ethanol stresses shown by those cells.
5) Both Hsp26p and Hsp104p were induced during conditioning heat treatment, which enhanced resistence to 5 mM H2O2, suggesting that these proteins might play a role in the mechanism of accquisition of tolerance to oxidative stress.
Finally, our observations suggest that all forms of defense systems could operate synergistically, cooperating at the same time, at different levels, and functioning to produce a quick and efficient protection mechanism against stress conditions. Further investigations for the understanding of the behavior of the SOD1 deleted strain are underway.
Material and Methods
Yeast strains, media and growth conditions
Saccharomyces cerevisiae strains used in this work are listed in Table 3. Stocks of all strains were maintained on solid 2% YPD (1% yeast extract, 2% glucose, 2% peptone and 2% agar) in proper conditions to avoid the selection of petites or suppressors. For all experiments, cells were grown up to mid log phase (1.0 mg dry weight/mL) in liquid YED pH 5.2 medium (1.3% yeast extract, 2.0% glucose, 0.2% (NH4)2SO4 and 0.2% KH2PO4), using an orbital shaker, at 28°C and 160 rpm, with the ratio of flask volume/medium of 5/1.
Table 3.
Saccharomyces cerevisiae strains used in this study.
| Strain | Genotype | Source |
| EG103 | MAT α, leu2-3,112, his3-1, trp1-289a, ura3-52, GAL+ | 1 |
| EG110 | Like EG103 except sod2::TRP1 | 1 |
| EG118 | Like EG103 except sod1::URA3 | 1 |
| EG133 | Like EG103 except sod2::TRP1 sod1::URA3 | 1 |
| JT2700 | MATa, MAL1, ura3, leu2 | 2 |
| JT9060 | Like JT2700 except tps1::LEU2 | 2 |
1- E. B. Gralla and J. S. Valentine, Department of Chemistry and Biochemistry, University of California Los Angeles, California, USA. 2- J. M. Thevelein, Laboratorium voor Moleculaire Celbiologie, Instituut voor Plantkunde, Katholieke Universiteitte Leuven, Belgium
Stress conditions
Severe heat shock was performed by submitting 5 ml of the culture in first exponential phase to 51°C/8 min. The toxic effect caused by 10% (v/v) ethanol during 24 h, was evaluated by adding absolute ethanol to cultures in the same conditions previously described, and incubated at 28°C/160 rpm. Hydrogen peroxide sensitivity was determined by exposing cells to a lethal dose of 5 mM H2O2 during 60 min at 28°C/160 rpm. In all experimental conditions, the cultures were divided into two parts, one was immediately submitted to stress conditions, while the other was adapted at 40°C/60 min and then exposed to hydrogen peroxide, heat and ethanol stresses.
Viability and trehalose determinations
Cell viability was analyzed by plating in triplicate on solidified 2% YPD, after proper dilution. The plates were incubated at 28°C for 72 h and the colonies counted. Viability was determined before and after stress conditions, using cells adapted or not at 40°C/60 min. Tolerance was expressed as percentage of survival. Trehalose was obtained from 12 mg of cells (dry weight) by centrifugation, washed with distilled water, extracted with ethanol (1 mL) at 80°C [33] and determined by the enzymatic specific trehalase method [34].
Immunological detection of Hsps
Total protein extracts were obtained from 50 mg (dry weight) and fractionated in 10% SDS-PAGE with discontinuous buffer [35]. Thereafter, proteins were electrophoretically transferred to nitrocellulose membranes, incubated for 2 h with rabbit IgG purified anti-Hsps and a further 2 h with goat anti-rabbit IgG or IgM conjugated to horseradish peroxidase. Finally, the blots were developed with diaminobenzidine and H2O2 plus CoCl2 to enhance sensitivity. Antibody anti-Hsp104p was bought from ABR (Affinity BioReagents), while the antibody anti-Hsp26p was obtained from Dr. J. T. Silva [36].
Fluorescence assays
The oxidant-sensitive probe 2',7'-dichlorofluorescein was used to measure the levels of intracellular oxidation developed in response to the heat treatment at 40°C. Fluorescence was measured using a PTI (Photo Technology International) spectrofluorimeter set at an excitation wavelength of 504 nm and an emission wavelength of 524 nm [37]. As a control, fluorescence was recorded over 60 min at 40 °C without any cells and with cells previously killed (data not shown). A fresh 5 mM stock solution of dichlorofluorescein dissolved in ethanol was added to the culture (to a final concentration of 10 μM) and incubation at 28°C continued for 15 minutes to allow uptake of the probe. Half of the culture was heated at 40°C and the other was maintained at 28°C. After 60 minutes, 50 mg of cells (adapted or not at 40°C) were cooled on ice, harvested by centrifugation and washed twice with 50 mM phosphate buffer, pH 6.0. The pellets were resuspended in 500 μL of the same buffer and 1.5 g of glass beads were added. The samples were lysed by cycles of one minute agitation on a vortex mixer followed by one minute on ice, repeated 3 times. The supernatant solutions were obtained after centrifugation at 13,000 rpm for 5 min, diluted 6-fold with water and, then, fluorescence was measured. The results were expressed as the relation between the fluorescence of heated and non heated cells.
Acknowledgments
Acknowledgments
We would like to thank Dr. Rogério Meneghini and Dr. Edith Gralla for providing the SOD strains as well as Prof. Ricardo Choloub and Ph.D student Vania Gomes Leal for the use of PTI spectrofluorimeter. This work was supported by grants from CNPq and FAPESP (n°1996/1405-7).
Contributor Information
Marcos D Pereira, Email: marcosdp@iq.ufrj.br.
Elis CA Eleutherio, Email: eliscael@iq.ufrj.br.
Anita D Panek, Email: anita@iq.ufrj.br.
References
- Fridovich I. Oxygen and Living Processes. Springer Verlag, New York/Heidelberg/Berlim. 1981.
- Valentine JS, Wertz DL, Lyons TJ, Liou L, Goto JJ, Gralla EB. The dark side of dioxygen biochemistry. Curr Opin Chem Biol. 1998;2:253–262. doi: 10.1016/s1367-5931(98)80067-7. [DOI] [PubMed] [Google Scholar]
- Moradas-Ferreira P, Costa V, Piper P, Mager W. The molecular defenses against reactive oxygen species in yeast. Mol Microbiol. 1996;19:651–658. doi: 10.1046/j.1365-2958.1996.403940.x. [DOI] [PubMed] [Google Scholar]
- Jamieson DJ. Oxidative stress responses of the yeast Saccharomyces cerevisiae. Yeast. 1998;14:1511–1527. doi: 10.1002/(SICI)1097-0061(199812)14:16<1511::AID-YEA356>3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]
- Gralla EB, Valentine JS. Null mutants of Saccharomyces cerevisiae Cu,Zn-superoxide dismutase: characterization and spontaneous mutation rates. J Bacteriol. 1991;173:5918–5920. doi: 10.1128/jb.173.18.5918-5920.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Changand EC, Kosman DJ. O2-dependent methionine auxotrophy in Cu/Zn-superoxide dismutase-deficient mutants of Saccharomyces cerevisiae. J Bacteriol. 1990;172:1840–1845. doi: 10.1128/jb.172.4.1840-1845.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crowe JH, Crowe LM. Preservation of mammalian cells - learning nature's tricks. Nat Biotechnol. 2000;18:145–146. doi: 10.1038/72580. [DOI] [PubMed] [Google Scholar]
- Eleutherio ECA, Ribeiro MJS, Pereira MD, Maia FM, Panek AD. Effect of trehalose during stress in a heat-shock resistant mutant of Saccharomyces cerevisiae. Biochem Mol Biol Int. 1995;36:1217–1223. [PubMed] [Google Scholar]
- Vuorio OE, Kaikkinen N, Londesborough J. Cloning of two related genes encoding the 56-kDa and 123 kDa subunits of trehalose synthase from the yeast Saccharomyces cerevisiae. Eur J Biochem. 1993;261:848–861. doi: 10.1111/j.1432-1033.1993.tb18207.x. [DOI] [PubMed] [Google Scholar]
- De Virgilio C, Hottiger T, Dominguez J, Boller T, Wiemken A. The role of trehalose synthesis for the acquisition of thermotolerance in yeast I. Genetic evidence that trehalose is a thermoprotectant. Eur J Biochem. 1994;219:179–186. doi: 10.1111/j.1432-1033.1994.tb19928.x. [DOI] [PubMed] [Google Scholar]
- Parsell DA, Kowal AS, Singer MA, Lindquist S. Protein desagregation mediated by heat shock protein Hsp104. Nature. 1994;372:475–476. doi: 10.1038/372475a0. [DOI] [PubMed] [Google Scholar]
- Elliott B, Haltiwanger RS, Futcher B. Synergy between trehalose and Hsp104 for thermotolerance in Saccharomyces cerevisiae. Genetics. 1996;144:923–933. doi: 10.1093/genetics/144.3.923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singer MA, Lindquist S. Thermotolerance in S. cerevisiae : the Yin and Yang of trehalose. Trends Biotechnol. 1998;16:460–468. doi: 10.1016/s0167-7799(98)01251-7. [DOI] [PubMed] [Google Scholar]
- Piper P. The heat shock and ethanol stress responses of yeast exhibit extensive similarity and functional overlap. FEMS Microbiol Lett. 1995;134:121–127. doi: 10.1111/j.1574-6968.1995.tb07925.x. [DOI] [PubMed] [Google Scholar]
- Yurkow EJ, McKenzie MA. Characterization of hypoxia-dependent peroxide production in cultures of Saccharomyces cerevisiae using flow cytometry: a model for ischemic tissue destruction. Cytometry. 1993;14:287–293. doi: 10.1002/cyto.990140309. [DOI] [PubMed] [Google Scholar]
- Tsuchiya M, Suematsu M. In vivo visualization of oxygen radical-dependent photoemission. Methods and Enzimology, Academic Press, USA. 1994;233:128–140. doi: 10.1016/s0076-6879(94)33015-8. [DOI] [PubMed] [Google Scholar]
- Zou J, Salminen WF, Roberts SM, Voellmy R. Correlation between glutathione oxidation and trimerization of heat-shock-factor-1, an early step in stress induction of the Hsp response. Cell Stresses & Chaperones. 1998;3:130–141. doi: 10.1379/1466-1268(1998)003<0130:cbgoat>2.3.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hottiger T, Boller T, Wiemken A. Rapid changes of heat and desiccation tolerance correlated with changes of trehalose content in Saccharomyces cerevisiae cells subjected to temperatures shifts. FEBS Lett. 1987;220:113–115. doi: 10.1016/0014-5793(87)80886-4. [DOI] [PubMed] [Google Scholar]
- Parrou JL, Teste MA, Francois J. Effects of various types of stress on the metabolism of reserve carbohydrates in Saccharomyces cerevisiae : genetic evidence for a stress-induced recycling of glycogen and trehalose. Microbiology. 1997;143:1891–1900. doi: 10.1099/00221287-143-6-1891. [DOI] [PubMed] [Google Scholar]
- Lewis JG, Learmonth RP, Attfield PV, Watson K. Stress co-tolerance and trehalose content in baking strains of Saccharomyces cerevisiae. J Ind Microbiol & Biotech. 1997;18:30–36. doi: 10.1038/sj.jim.2900347. [DOI] [PubMed] [Google Scholar]
- McCord JM, Fridovich I. Superoxide dismutase: an enzymic function for erythrocuprein. J Biol Chem. 1969;244:6049–6055. [PubMed] [Google Scholar]
- Jamieson DJ, Rivers SL, Stephen DWS. Analysis of Saccharomyces cerevisiae proteins induced by peroxide and superoxide stress. Microbiology. 1994;140:3277–3283. doi: 10.1099/13500872-140-12-3277. [DOI] [PubMed] [Google Scholar]
- Morimoto RI. Regulation of the heat shock transcriptional response: cross talk between a family of heat shock factors, molecular chaperones and negative regulators. Genes Dev. 1998;12:3788–3796. doi: 10.1101/gad.12.24.3788. [DOI] [PubMed] [Google Scholar]
- Godon C, Lagniel G, Lee J, Buhler JM, Kieffer S, Perrot M, Boucherie H, Toledano MB, Labarre J. The H2O2 stimulon in Saccharomyces cerevisiae. J Biol Chem. 1998;273:22480–22489. doi: 10.1074/jbc.273.35.22480. [DOI] [PubMed] [Google Scholar]
- Longo VD, Gralla EB, Valentine JS. Superoxide Dismutase activity is essential for stationary phase survival in Saccharomyces cerevisiae. J Biol Chem. 1996;271:12275–12280. doi: 10.1074/jbc.271.21.12275. [DOI] [PubMed] [Google Scholar]
- Tamai K, Gralla EB, Ellerby LM, Valentine JS. Yeast and mammalian methallothioneins functionally substitute for yeast copper-zinc superoxide dismutase. Proc Natl Acad Sci USA. 1993;90:8013–8017. doi: 10.1073/pnas.90.17.8013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costa V, Reis E, Quintanilha A, Moradas-Ferreira P. Acquisition of ethanol tolerance in Saccharomyces cerevisiae : the key role of the mitochondrial superoxide dismutase. Arch Biochem Biophys. 1993;300:608–614. doi: 10.1006/abbi.1993.1084. [DOI] [PubMed] [Google Scholar]
- Balasundaram D, Tabor CW, Tabor H. Sensitivity of polyamine-deficient S. cerevisiae to elevated temperatures. J Bacteriol. 1996;178:2721–2724. doi: 10.1128/jb.178.9.2721-2724.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costa V, Amorim MA, Quintanilha A, Moradas-Ferreira P. Mitochondrial superoxide dismutase is essential for ethanol tolerance of Saccharomyces cerevisiae in the post-diauxic phase. Microbiology. 1997;143:1649–1656. doi: 10.1099/00221287-143-5-1649. [DOI] [PubMed] [Google Scholar]
- McDufee AT, Senisterra G, Huntley S, et al. Proteins containing non-native disulfide bonds generated by oxidative stress can act as signals for the induction of the heat shock response. J Cell Physiol. 1997;171:143–151. doi: 10.1002/(SICI)1097-4652(199705)171:2<143::AID-JCP4>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- Polyak K, Xia Y, Zweier JL, Kinzler KW, Vogelstein B. A model for p53-induced apoptosis. Nature. 1997;389:300–305. doi: 10.1038/38525. [DOI] [PubMed] [Google Scholar]
- Eleutherio ECA, Araújo PS, Panek AD. Role of trehalose carrier in dehydration resistance of Saccharomyces cerevisiae. Biochim Biophys Acta. 1993;1156:263–266. doi: 10.1016/0304-4165(93)90040-f. [DOI] [PubMed] [Google Scholar]
- Ferreira JC, Paschoalin VMF, Panek AD, Trugo LC. Comparison of three different methods for trehalose determination in yeast extracts. Food Chem. 1997;60:251–254. [Google Scholar]
- Tourinho dos Santos CF, Bachinski N, Paschoalin VMF, Paiva CLA, Silva JT, Panek AD. Periplasmic trehalase from Escherichia coli -Characterization and mobilization on spherisorb. Braz J Med Biol Res. 1994;27:627–636. [PubMed] [Google Scholar]
- Laemmly UK. Cleavage of structural proteins during the assembly of the head of bacteriofage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Silva JT, Verícimo MA, Floriano WB, Dutra MB, Panek AD. On the Hsp26 of Saccharomyces cerevisiae. Biochem Mol Biol Int. 1994;3:211–220. [PubMed] [Google Scholar]
- Davidson JF, White B, Bissinger PH, Schiestl RH. Oxidative stress is involved in heat-induced cell death in Saccharomyces cerevisiae. Proc Natl Acad Sci USA. 1996;93:5116–5121. doi: 10.1073/pnas.93.10.5116. [DOI] [PMC free article] [PubMed] [Google Scholar]