

Abstract
Myocardial ischemia needs an alternative treatment such as gene therapy for the direct protection of cardiomyocytes against necrosis or apoptosis and to prevent the development of myocardial fibrosis and cardiac dysfunction. Despite the utility of gene therapy, its therapeutic use is limited due to inadequate transfection in cardiomyocytes and difficulty in directing to ischemic myocardium. Here, we present a polymeric gene carrier that is capable of targeting ischemic myocardium, resulting in high localization within the ischemic zone of the left ventricle (LV) of a ischemia/reperfusion (I/R) rat model upon systemic administration. Cystamine bisacrylamide-diamino hexane (CD) polymer was modified with the ischemic myocardium-targeted peptide (IMTP) and D-9-arginine (9R) for dual effects of the homing to ischemic myocardium and enhanced transfection efficiency with minimized polymer use. Conjugation of IMTP and 9R to CD led to an increase in transfection under hypoxia and significantly reduced the amount of polymer required for high transfection. Finally, we confirmed targeting of IMTP-CD-9R/DNA polyplex to ischemic myocardium and enhanced gene expression in LV of the I/R rat after tail vein injection. This study provides a clue that gene therapy for the treatment of myocardial ischemia can be achieved by using homing peptide-guided gene delivery systems.
Keywords: Ischemic myocardium targeting, Targeted gene delivery, Myocardial ischemia, Gene therapy, Homing peptide
1. Introduction
Myocardial ischemia is one of the leading causes of death worldwide. This disease is typically caused by reduced blood flow, resulting in a decrease in the nutrient and oxygen supply.1, 2 The main injuries of myocardial ischemia include cardiomyocyte hypertrophy, apoptotic myocyte loss, progressive collagen replacement, and enlargement of the left ventricle (LV).3 Conventional pharmacological treatments for this disease are insufficient to prevent the remodeling of the LV and terminate the progress of heart failure. Attempts to develop more effective agents working directly to the ischemic heart have considered gene therapy because it has benefits of direct effects on cardiomyocytes against necrosis or apoptosis and thereby preventing the development of myocardial fibrosis and cardiac dysfunction. Unfortunately, gene therapy for the treatment of myocardial ischemia shows limited efficacy due to absence of sufficient gene carrier targeting to the disease area and difficulty in transfection to the myocardium.4
Systemic administration of polyplexes targeted to the ischemic myocardium affords significant advantages over local injection. First, targeted polyplexes can be administered immediately following reperfusion via IV injection and do not require a separate procedure with accompanying risks to deliver the polyplex locally. Second, systemic administration allows for easy follow-on dosing to maximize the gene therapy benefit. Primary cardiomyocyte-targeted polymeric gene carrier developed by our group enhanced gene transfection in cardiomyocytes using a homing peptide.5 However, since it may direct gene to whole heart tissues when administered systemically, but not only ischemic region, future challenges still remain.6 Recently, a peptide sequence that has high specificity to ischemic myocardium was identified by in vivo phage display in ischemia/reperfusion (I/R) rat.7 In vivo phage display for high-throughput screening referred to as in vivo biopanning capacitates to screen a specific peptide that homes organs or tissues in a living animal.8 This technique is useful for the development of targeted therapeutics, imaging agents, and diagnostic markers in various diseases.9, 10 Nevertheless, only a few studies have reported targeted delivery of therapeutics by using the diseased tissue-targeted peptides and their main objectives are the targeting to tumor or blood vasculature.11 We demonstrated that the ischemic myocardium-targeted peptide (IMTP) could direct the gene to the ischemic heart and increase the accumulation and transfection of gene in the ischemic myocardium.
Reducible cationic polymer is widely used for gene transfection due to its capability to condense nucleic acid, rapid cellular uptake, and intracellular reducibility to facilitate release of gene.12 Cystamine bisacrylamide-diamino hexane (CD) was chosen as a backbone for the modification with IMTP and D-9-arginine (9R).13 Since 9R has been known as the most effective peptide for protein transduction as well as gene transfection, the attachment of 9R to CD would increase the transfection efficiency and decrease amount of the polymer required for sufficient transfection.14, 15 According to our hypothesis, we prepared eight types of IMTP-CD-9R (IMTP and 9R-conjugated CD) polymers that contain different molar ratios of IMTP and 9R to CD and compared transfection efficiencies of those polymers. The optimum ratio between IMTP and 9R was decided by comparing transfection efficiency and cytotoxicity, and the best modification showed higher transfection than did PEI. Although various polymeric gene carriers have shown the evidence of a great potency for cardiovascular medicine, gene therapy for the disease still has challenges due to the difficulty in ischemic heart targeting.16 In this study, we clearly demonstrated the targeting of IMTP-CD-9R/DNA polyplex to and the increase in gene expression in the LV of the I/R rats after a systemic injection.
2. Materials and Methods
2.1 Materials
CD (Mw=3.5 kDa) was prepared as described in previously published paper.13 IMTP (Ac-CSTSMLKAC-COOH, cyclic form, Mw=943.2 Da), scrambled IMTP (Ac-CSKTALSMC-COOH), and D-9R (Crrrrrrrrr, Mw=1526.8 Da; C: L-cysteine and r: D-arginine) were synthesized and purified by the Health Sciences Center core research facilities at the University of Utah (Salt Lake City, UT). Poly(ethylenimine) (PEI; branched form, Mw=25 kDa), 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT), Fluoreshield mounting solution were purchased from Sigma-Aldrich (St. Louis, MO). EDC, sulfo-NHS, SPDP and Slide-A-Lyzer Dialysis Cassettes (3.5 k MWCO) were purchased from Pierce (Thermo Fisher Scientific Inc.; Rockford, IL). Cy5.5-NHS ester was from Amersham Biosciences (GE Healthcare; Piscataway, NJ). All cell culture products including fetal bovine serum (FBS), Dulbecco’s phosphate buffered saline (DPBS), and Dulbecco’s modified Eagle’s medium (DMEM) were obtained from Invitrogen (GibcoBRL; Carlsbad, CA). The Luciferase assay system and reporter lysis buffer were from Promega (Madison, WI). Wheat Germ Agglutinin, Alexa Fluor® 594 Conjugate, and 5-(Aminomethyl) Fluorescein-Hydrochloride were from Invitrogen (Molecular Probes, Inc.; Eugene, OR).
2.2 Synthesis of IMTP-CD-9R
IMTP was first conjugated with CD backbone through an amide bond by EDC/NHS reaction. To prevent the inter-molecular crosslinking between IMTP molecules, N-terminal-blocked Ac-CSTSMLKAC-COOH peptide was synthesized and used for the IMTP-CD conjugation. IMTP and CD were dissolved in PBS (pH 7.4). For the IMTP activation, EDC was added directly to the IMTP solution at a final concentration of 2.5 mM. After 30 minutes, sulfo-NHS was added to the reaction, which yields 575 mM; then incubate 2 hours at room temperature. The activated IMTP was added to the CD solution at various molar ratios. The mixture was reacted for overnight at room temperature. To separate the IMTP-CD from excess EDC/NHS and unreacted IMTP and CD, the final resultant was dialyzed against water using a Slide-A-Lyzer Dialysis Cassette (3.5 kDa MWCO) and then freeze-dried. The 9R was conjugated to the IMTP-CD polymer using SPDP, a heterobifunctional crosslinker. SPDP-mediated crosslinking between the 9R peptide and the IMTP-CD was performed according to the manufacturer’s protocol. The feeding molar ratios used to prepare different types of IMTP-CD-9R are shown in Table 1. The unreacted impurities were removed by a dialysis against water using the Slide-A-Lyzer Dialysis Cassette (3.5 kDa MWCO). The lyophilized IMTP-CD-9R was kept at -20°C until use. The synthesis was confirmed by 1H-NMR.
Table 1.
Characteristics of IMTP-CD-9R
| Reaction | Feeding ratio (mol/mol) | Substitution ratio (mol/mol) | ||||
|---|---|---|---|---|---|---|
| IMTP | CD | 9R | IMTP | CD | 9R | |
| Rx1 | 0.625 | 10 | 3 | 1 | 10 | 1.5 |
| Rx2 | 1.25 | 10 | 3 | 2 | 10 | 1 |
| Rx3 | 2.5 | 10 | 3 | 2.5 | 10 | 1 |
| Rx4 | 5 | 10 | 3 | 3 | 10 | 1 |
| Rx5 | 1.25 | 10 | 3.5 | 2 | 10 | 2 |
| Rx6 | 1.25 | 10 | 4.5 | 2 | 10 | 3 |
| Rx7 | 1.25 | 10 | 5.5 | 2 | 10 | 3.5 |
| Rx8 | 1.25 | 10 | 6.5 | 2 | 10 | 4 |
2.3 Gel retardation
IMTP-CD-9R polymer dissolved in PBS was mixed with 1 μg pSV-Luci at various weight ratios. The mixtures were incubated for 20 minutes at room temperature and electrophoresed on 1% (w/v) agarose gel for 30 minutes at 100 V in 0.5% TBE buffer solution. Ratios were calculated as weight ratios of IMTP-CD-9R to the pSV-Luci for all data. To observe reducibility of the polyplex, the IMTP-CD-9R polyplex prepared at weight ratio of 4/1 (Rx7/pSV-Luci) was incubated in the presence of various concentrations of DTT. The samples were reacted under gentle stirring for 2 hours at 37 °C and then electrophoresed as described above.
2.4 DNA stability in serum
To test DNA stability in serum after forming polyplex with IMTP-CD-9R, the Rx7/pSV-Luci polyplexes were prepared at weight ratios of 2.0 and 4.0. After 20 min of incubation, equal volume of rat serum was added to the samples and then incubated at 37°C under shaking at 150 rpm. After 1 or 2 hours of incubation, heparin in PBS (pH 7.4, 0.3 mol/l NaCl) containing 10 mmol/l EDTA was added to the samples at a final volume ratio of 1:1 for disassembling the Rx7/pSV-Luci polyplex. After 1 hour, the mixtures were electrophoresed on 1% (w/v) agarose gel for 30 minutes at 100 V in 0.5% TBE buffer solution.
2.5 Polyplex characterization
Mean diameters and zeta potentials of the polyplexes prepared at various weight ratios of the IMTP-CD-9R to pSV-Luci were measured using DLS (Zetasizer-Nano ZS; Malvern Instruments, Worcestershire, UK). Four micrograms of pSV-Luci were mixed with the Rx7 at weight ratios of 0.5, 1.0. 2.0, 3.0, and 4.0 in 1 ml PBS (pH 7.4). To see how the polyplex behaves under reductive environments, DTT was added to the polyplex prepared at weight ratio of 4/1 at a final concentration of 5 mM after 47.5 minutes of incubation, and the size and zeta potential changes were observed for 127.5 minutes with a 2.5 minutes interval. At 170 minutes post-polyplex formation, DTT was added again at a final concentration of 10 mM, and DLS was performed. To see reducibility of the polyplex, DTT was added to the samples at various concentrations after polyplex formation and then incubated at 37°C under shaking at 150 rpm. After 120 minutes of incubation, the samples were electrophoresed as described above.
2.6 Cell culture
DMEM containing 10% FBS and 1% penicillin-streptomycin was used for H9C2 cells, rat cardiomyocytes. H9C2 cells were seeded on 24-well plates at a density of 2.0 × 104 cells/well when cell confluence reached 80%. Cells were cultured under hypoxia, normoxia, or hypoxia followed by normoxia conditions. The seeded cells were incubated under the hypoxia condition composed of 94% NO2, 5% CO2, and 1% O2, normoxia states, or mixed condition of hypoxia and normoxia (12 + 12 hours).
2.7 In vitro transfection
After 24 hours post-seeding, the culture media were replaced with the plain media containing IMTP-CD-9R/pSV-Luci or PEI/pSV-Luci polyplexes prepared by mixing 1 Sg pSV-Luci with different amounts of IMTP-CD-9R or 1 Sg PEI. The culture media were replaced with the complete media after 4 hours of incubation. At 48 hours post-transfection, cells were washed with PBS and lysed with 100 Sl reporter lysis buffer according to the manufacturer’s instruction. The cells were harvested and centrifuged for 30 seconds at 13,000 rpm. Luciferase RLU of the cells was measured using a 96-well plate luminometer, and the results were expressed as RLU/mg protein determined by micro BCA assay kit with a BSA standard. Cell viabilities were determined by MTT assay.
2.8 Competition assay
After 24 hours of cell seeding, the culture media were replaced with the plain media containing IMTP to make final concentrations of 50, 100, and 200 μM. The Rx7/pSV-Luci polyplex prepared at weight ratio of 4/1 were treated to the cells 2 hours after the IMTP treatment. After additional 4 hours of incubation, the culture media were changed with the complete media and then the cells were cultured for 44 hours. The luciferase assay and MTT assay were performed as described above.
2.9 Ex vivo imaging
Animal experiments were approved by the University of Utah’s Institutional Animal Care and Use Committee and followed the guidelines provided by the National Institutes of Health in Guide for the Care and Use of Laboratory Animals. To prepare ischemia/reperfusion injury rat model, male Sprague-Dawley rats were anesthetized with 4% isoflurane, intubated and kept under 2% isoflurane for maintaining anesthesia. A thoracotomy was performed to expose the heart in the 4th or 5th intercostal space. The left anterior descending coronary artery (LAD) was occluded for 30 minutes by suture threading through a small piece of polyethylene tubing to serve as a reversible snare occluding. After 20 minutes of reperfusion, Cy5.5-labeled IMTP-CD-9R (Rx7)/pSV-Luci polyplex prepared at a weight ratio of 4/1 was administered through the tail vein at a DNA dose of 50 μg. Rats with the polyplex injection and without surgery were prepared as a control group. Following 20 minutes circulation, the hearts were isolated and washed with PBS three times. In addition, the heart was sectioned to clearly observe location of the polyplex in the heart tissue. The distribution of the Cy5.5-labeled polyplex was visualized and analyzed using the FMT 2500 quantitative tomography system (VisEn Medical, Boston, MA).
2.10 Targeted gene expression
Preparation of the animal model and injection of the IMTP-CD-9R/HO-1 polyplexes were performed following the method described above. After 3 days of injection, the rats were sacrificed under the lethal concentration of isoflurane, and then the main organs including heart (liver, kidney, spleen, and lung) were isolated and sectioned through short axis. LV and RV in the third heart section from apex and the other organs were separately homogenized in the total tissue protein extraction buffer (Invitrogen, Camarillo, CA) according to the manufacturer’s protocol. Human HO-1 expression and total protein concentration were determined using HO-1 ELISA kit (R&D Systems, Minneapolis, MN) and BCA protein assay kit (Pierce, Rockford, IL), respectively.
3. Results
3.1 Synthesis of IMTP-CD-9R
CD, a cationic reducible polymer, was modified with IMTP and 9R for the ischemic myocardium targeting and enhancing gene transfection, respectively, and the modification was confirmed and quantitatively analyzed by the 1H-NMR peaks of D-arginine (around 3.2 – 3.0 ppm: the protons of γ and δ carbons in the side chain) in D-9R and L-lysine (around 2.0 – 1.8 ppm: the protons of β, γ, and δ carbons in the side chain) in IMTP. Table 1 shows the feeding molar ratios and the substitution molar ratios of IMTP and 9R to CD and the resulting polymers were referred to as Rx1 – Rx8. The amounts of EDC/NHS and 9R were fixed for the synthesis of Rx1 – Rx4 and the constant concentrations of IMTP and SPDP were used to prepare the Rx5 – Rx8. The molecular weight of Rx7 was determined to be ~11 kDa. Various modifications were made by the reactions as shown in Table 1.
3.2 Characteristics of IMTP-CD-9R/DNA polyplex
The charge density of CD might be increased by 9R conjugation, which is one of the most effective cationic peptides for gene transfection and protein transduction. The conjugate is therefore expected to improve the DNA condensation ability of CD, regardless of the presence of IMTP. Gel retardation assay was performed to evaluate the capability of IMTP-CD-9R for DNA condensation. DNA migration was completely retarded at weight ratio of 1.0 in Rx1, 2, 5, 6, and 7 and DNA was partially and fully condensed at weight ratios of 1.0 and 1.5, respectively, in Rx3, 4, and 8 (Fig. 1a). There are no significant differences in DNA condensation ability among Rx1 – Rx8 polymers and all of the polymers are capable of condensing DNA. Naked DNA and condensed DNA by Rx7 were incubated with rat serum and the polyplexes were dissociated by adding heparin. Fig. 1b shows rapid degradation of naked DNA after 1 hour of exposure to the serum and clear DNA bands appeared at the same position as the control after dissociation of Rx7/DNA polyplexes. Thus, Rx7 is able to protect DNA even at weight ratio of 2 and 2 hours of incubation in the presence of serum. The mean diameter and the zeta potential of Rx7/DNA polyplex increased significantly with increasing weight ratios up to 2 and slight changes in the size and the zeta potential were observed up to 4 (Fig. 1d). The zeta potential was positive at the weight ratio of 1 and reached ~40 mV at the ratio of 4. Continuous measurement of the size and the zeta potential reveals that Rx7/DNA polyplex maintains its nano-structure and positive zeta potential for 6 hours of observation (data not shown).
Figure 1.


Characteristics of IMTP-CD-9R/DNA polyplex. (a) DNA condensation: DNA was incubated with eight types of IMTP-CD-9R at different weight ratios for 20 minutes in PBS (pH 7.4). (b) DNA stability in serum: Rx7/DNA polyplexes prepared at ratios of 2 and 4 were incubated in the presence of 50% rat serum for 1 and 2 hours at 37°C and then heparin was added to the samples for polyplex dissociation. (c) Polyplex reducibility: Rx7/DNA polyplex was incubated with increasing concentrations of DTT for 2 hours. (d) Size and zeta potential, and (e) size and zeta potential changes in the presence of DTT. Size and zeta potential of Rx7/DNA polyplex was measured and the effect of DTT treatment on the size and zeta potential was observed using DLS. Ratios are indicated as w/w, and data represent mean ± SD (n=5).
Since CD is one of reducible polymers responding to glutathione, the behavior of Rx7/DNA polyplex in the presence of DTT was investigated by gel retardation assay and measuring changes in the size and the zeta potential. Rx7/DNA polyplex incubated with increasing concentrations of DTT released no DNA and showed weak DNA intensities in the wells on agarose gel, whereas no intensity was observed in the lane running only polyplex incubated without DTT (Fig. 1c). Observation of changes in the mean diameter and the zeta potential demonstrates how the polyplex behaves under the reducing environment (Fig. 1e). The polyplex size retained the steady state for 50 minutes of measurement, initiated to increase 10 minutes after adding DTT, and reached the plateau at ~350 nm in diameter. An additional DTT treatment at 170 minutes post-polyplex formation had no effect on the polyplex size under the test condition. The zeta potentials recorded before the first and the second DTT addition and at the end of the size measurements were +34 ± 1.5, −45 ± 3.5, and −46 ± 3.6 mV, respectively.
3.3 In vitro transfection using Rx1 – Rx8
The eight types of IMTP-CD-9R polymers may have different transfection efficiencies due to the contents of 9R they contain. In order to compare the transfection efficiency, eight types of IMTP-CD-9R/pLuci polyplexes were transfected to the cells that were cultured under hypoxia. The difference between Rx1 – Rx4 is mainly content of IMTP and that in Rx5 – Rx8 is portion of 9R peptide. Fig. 2a that shows the effect of IMTP on transfection efficiency reveals that Rx2 is the most effective modification among the four types of polymers. The optimum molar ratio of IMTP to CD was decided based on the transfection results at the feeding ratio of 1.25. Additional four types of IMTP-CD-9R were prepared to contain different ratios of 9R to CD with the constant feeding ratio of IMTP. As shown in Figs. 2b,c, Rx7 and Rx8 expressed luciferase higher than did PEI in all weight ratios and showed negligible cytotoxicity at weight ratio of 4 and a ratio-dependent cytotoxicity at and above the ratio 6. Since IMTP-CD-9R is designed to target ischemic myocardium, the transfection may increase under hypoxic condition compared to normoxia. Rx7/pLuci polyplexes were transfected to the cells incubated under the hypoxia, the hypoxia + normoxia, and the normoxic condition. The reporter gene expressions of Rx7/pLuci polyplex under the normoxia and the hypoxic + normoxic conditions were determined to be ~50% and ~80% relative to the hypoxia, respectively (Fig. 2d). On the other hand, PEI, a non-targeting polymer, showed highest transfection in the normoxic condition. This result suggests that IMTP-CD-9R has high specificity to the hypoxic cardiomyocytes rather than the normoxic cardiomyocytes.
Figure 2.

Determination of the best IMTP-CD-9R among the polymers. (a) Transfection efficiencies of Rx1 – Rx4 and (b) Rx5 – Rx8, (c) cytotoxicity of Rx5 – Rx8, and (d) transfection patterns under different conditions. H9C2 cells were incubated under the hypoxic condition for 24 hours and IMTP-CD-9R/pLuci polyplexes were transfected to the cells. Luciferase activity was measured 48 hours after the transfection and the cell viability was determined by MTT assay. The cells were cultured under 24 hours of normoxia, hypoxia, and hypoxia + normoxia (12 + 12 hours) prior to the transfection. Transfection efficiency was converted as relative values to PEI and data represent mean ± SD (n=4, triplicates). The numbers in figure legend indicate w/w ratio of IMTP-CD-9R/DNA.
3.4 Optimization of transfection condition for Rx7
Rx7 was selected as the best modification for ischemic myocardium-targeted gene delivery among eight types of IMTP-CD-9R in terms of the high transfection efficiency, the ischemia-targeting ability, and the low cytotoxicity. Scrambled IMTP and 9R-conjugated Rx7 (sRx7) was prepared and used as a negative control. Optimum weight ratio of Rx7 and pLuci needs to be determined to maximize transfection and minimize cytotoxicity. Ratios of 4 – 10 were already examined and it was proved that hypoxia condition led to increased transfection; therefore, weight ratios ranging from 1 to 4 were tested. As shown in Fig. 3a, the transfection was over 300% relative to PEI at the ratio of 4 under the hypoxic condition, whereas it was less than 100% under the normoxia and sRx7 showed ~200% in hypoxic cells. The transfection efficiency decreased in Rx7 weight ratio-dependent manner and no transfection was observed at the ratio of 1. The weight ratios of 1 – 4 were non-toxic to the cells under both hypoxia and normoxia (Fig. 3b). Taken together, we decided to use Rx7 at the weight ratio of 4 for all subsequent experiments.
Figure 3.

Optimum transfection condition. (a) Ratio-dependent transfection efficiency and (b) cell viability. Rx7/pLuci polyplexes prepared at different weight ratios and sRx7/pLuci polyplex prepared at weight ratio 4 were transfected to H9C2 cells that were pre-cultured under hypoxia or normoxia. Luciferase activity was calculated as percent values relative to PEI. Cytotoxicity was determined by MTT assay and expressed as relative cell viability compared to the control. Data represent mean ± SD (n=4, triplicates; *p < 0.01).
3.5 Competition assay
The transfection increases by the interaction between IMTP and the ischemia-specific molecules on the cell surface. Thus, the transfection efficiency of Rx7 may decrease when it is transfected to the cells that are pre-incubated with free IMTP molecules due to the competitive binding to the unknown receptor on the cell surface. IMTP was pre-treated to the cells that were cultured under the hypoxia or the normoxia and Rx7/pLuci or PEI/pLuci polyplexes were transfected to the cells. Under the hypoxic condition, the transfection decreased dose-dependently in the presence of the free IMTP in the Rx7/pLuci transfection group, while PEI/pLuci showed no significant difference in the transfection (Fig. 4a). On the other hand, the competition was not observed in both Rx7 and PEI under the normoxia as shown in Fig. 4c. Concurrent determination of cytotoxicity in the presence of same concentrations of IMTP revealed that the results were not from the cytotoxicity of IMTP (Fig. 4b, d).
Figure 4.

Competition assay. Transfection efficiencies under (a) hypoxia and (c) normoxia and cell viability under (b) hypoxia and (d) normoxia in the presence of free IMTP. H9C2 cells were incubated under hypoxia or normoxia for 24 hours and then free IMTP was treated to the cells. After 2 hours, pLuci was transfected by IMTP-CD-9R or PEI and the luciferase activity and cell viability were measured at 48 hours post-transfection. Data represent mean ± SD (n=4, triplicates; *p < 0.01).
3.6 Targeting of IMTP-CD-9R/DNA polyplex to ischemic myocardium
Targeting of IMTP-CD-9R/DNA polyplex to the ischemic myocardium after systemic administration needs to be observed because myocardium is one of the hard-to-reach tissues among major organs. To observe the homing of the polyplex to ischemic myocardium, the I/R rat was prepared and IMTP-CD-9R was labeled with Cy5.5 dye. Cy5.5-IMTP-CD-9R/DNA polyplex was injected through tail vein after 30 minutes of ischemia and 20 minutes of reperfusion. The hearts were isolated 20 minutes after the polyplex administration. For the clear images, short-axis specimen sections with 2 mm thickness were performed and the specimens were observed using FMT 2500. Heart sections of the I/R and normal rats (Fig. 5) show the location of Cy5.5-IMTP-CD-9R/DNA polyplex (yellow dotted line). Significant fluorescence intensity was observed only in the LV wall as indicated in yellow region, whereas no fluorescence intensity was detected in the normal heart.
Figure 5.
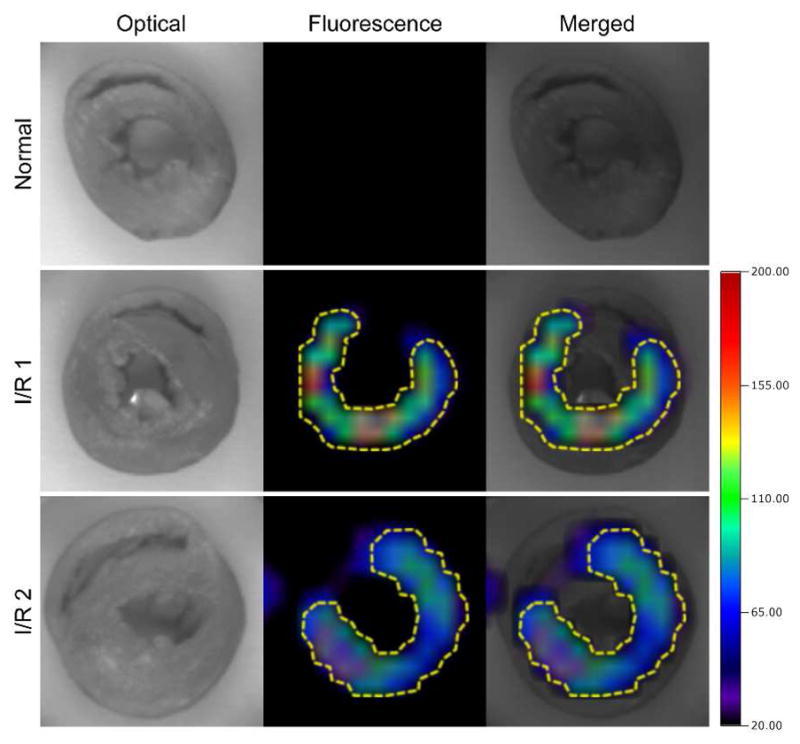
Targeting to the ischemic heart. Ex vivo distribution of IMTP-CD-9R/DNA polyplex in the heart. Cy5.5-labled IMTP-CD-9R/pLuci polyplex was injected to the I/R rats or normal rats and then circulated for 20 minutes. The hearts were isolated and perfused to remove the remaining bloods. I/R 1 and I/R 2 indicate two independent hearts of the rats. Distribution of the polyplexes were observed immediately, visualized and analyzed using FMT 2500.
3.7 Targeted gene expression after a systemic injection
The targeted gene expression was further investigated after a systemic injection of HO-1 plasmid as a model using IMTP-CD-9R because the ex vivo imaging demonstrated location of the polyplex, but not gene expression. The polyplexes of IMTP-CD-9R/HO-1 was injected intravenously to the I/R and the thoracotomy rats, and the HO-1 gene expression in the LV, the RV, and the other organs was determined to confirm the targeted gene expression. As shown in Fig. 6, HO-1 expression was ~2100 and ~1200 pg/mg in the LV and the RV of the I/R rats, respectively, and the thoracotomy rats showed no difference in HO-1 expression between the LV and the RV (~1200 pg/mg protein). The HO-1 expression was also observed in lung and liver, and spleen and kidney slightly expressed HO-1. I/R rats without the polyplex injection showed background level of HO-1 expression (cross-reactivity between human and rat), where is ~200 pg/mg tissue protein. The increase in HO-1 expression after systemic administration of IMTP-CD-9R/HO-1 polyplexes in the LV of the I/R rats is due to the targeting to ischemic myocardium.
Figure 6.
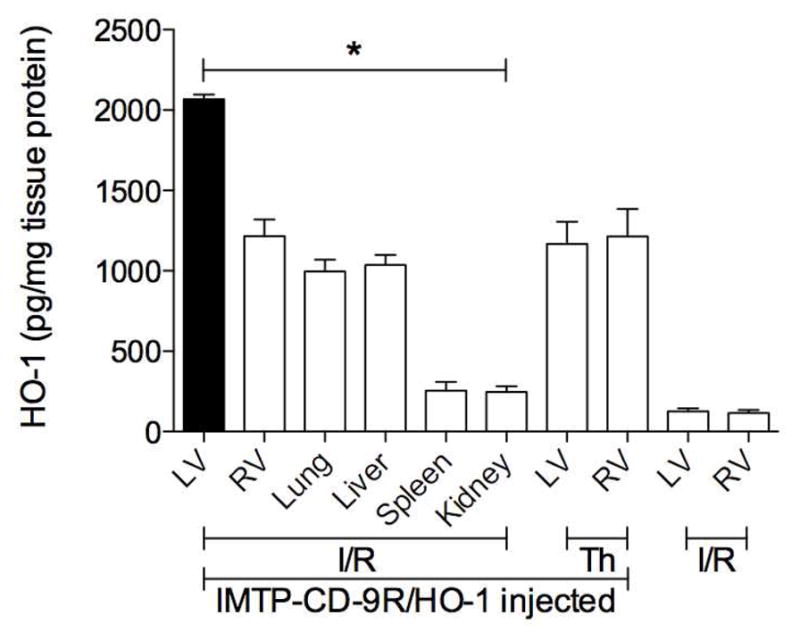
Targeted gene expression in LV of I/R rats. IMTP-CD-9R/HO-1 polyplex was injected intravenously to the I/R and the thoracotomy (Th) rats, and then the hearts were isolated, perfused, sectioned, and homogenized 3 days post-injection. The hearts of I/R rats without injection were prepared as a control. The HO-1 expression was determined using an ELISA kit and calculated as HO-1/mg total tissue protein. Data represent mean ± SD (n=6, *p < 0.01, one-way ANOVA).
4. Discussion
Biophysical properties of cationic polymer/DNA polyplex play important roles in transfection. Nano-sized aggregation, positive zeta potential, high stability in physiological condition, and rapid dissociation of polyplex in cells are main factors for enhancing transfection efficiency. Many of cationic polymers developed recently contain internal disulfide linkages that can be reduced by glutathione.17 Disulfide bond reduction facilitates dissociation of the interaction between cationic polymer and negatively charged nucleic acids, resulting in the improved transfection. On the other hand, a couple of papers reported that some reducible polymers did not release the nucleic acids upon disulfide bond reduction.18 CD backbone used here for conjugation of IMTP and 9R is one of the reducible polymers that release nucleic acids in the presence of DTT.19 However, no DNA release was observed upon reduction of IMTP-CD-9R. This is due, in part, to that the reduction initiates to separate the native 9R from CD, resulting in weak binding between the native 9R and DNA. According to the Table 1, the estimated ratio of 9R in Rx7 at weight ratio of 4 to DNA is ~0.6, where shows loose structure and negative zeta potential.14 The observation of changes in the size and the zeta potential in the presence of DTT explains why the reduction has no effect on the DNA release. The polyplex size was stable before the DTT treatment and it continuously increased and reached steady state after the first DTT treatment. The initial zeta potential of polyplex was positive and it changed to negative after the first DTT treatment. The second DTT adding did not affect the size and the zeta potential. The data represent that the first DTT treatment is sufficient to reduce the disulfide linkages in IMTP-CD-9R and the changes in size and zeta potential is not an artifact made by the DTT treatment. The size increase indicates that the compact aggregation converts to loose structure due to the weak rigidity and the negative zeta potential demonstrates that DNA exists in outer shell of the polyplex. Consequently, it is expected that IMTP-CD-9R/DNA polyplex is reduced by intracellular glutathione and they form loose structure, by which DNA exposes to cytosol, even though the complete dissociation does not occur.
One of the common characteristics of various PTDs is arginine-rich structure and it is well known that the peptide sequence containing six or more arginines leads to rapid internalization into cells.20, 21 Nevertheless, short peptide composed of arginine is insufficient to protect nucleic acids against degradation in serum or metabolism in vivo due to the low cooperative effect between the peptide and nucleic acids and inadequate number of free arginine for interaction with cell membrane.14, 22 Since only three or four arginines among octaarginine are open to contact with cell membrane when the peptide binds with nucleic acids, peptide composed of more arginines is expected more efficient. Thus, cellular uptake may be in direct proportion to the number of free arginine. It is assumed that the transfection efficiency increases as the content of 9R in IMTP-CD-9R increases. The transfection results prove that the key molecule to increase the transfection is the 9R peptide because conjugates of Rx5 – Rx8 are more efficient than Rx1 – Rx4 for gene transfection and they contain more 9R peptide than do Rx1 – Rx4. In addition, the positive relationship between the transfection and the content of 9R observed in Rx5 – Rx7 polymers supports that the 9R peptide affects the transfection efficiency primarily. There are negligible differences between Rx7 and Rx8; therefore, Rx7 is accepted as the best IMTP-CD-9R among the eight types of polymers in terms of the high transfection efficiency and the less amount of 9R required for preparing IMTP-CD-9R. Effect of IMTP, another factor affecting the cellular binding, on the transfection is moderate as observed in transfection using Rx1 – Rx4 because IMTP includes only one cationic amino acid that may have positive effects on the transfection.
Despite of no significant effect of IMTP on the transfection, IMTP is capable of enhancing the transfection under the hypoxic condition. PEI, a non-hypoxia-responsible gene carrier, decreases the transgene expression in the order of the normoxia, the hypoxia + normoxia, and the hypoxia. On the other hand, IMTP-CD-9R increased the gene expression under the hypoxic condition, suggesting that IMTP might interact with the receptors on cell membrane rapidly and/or facilitate internalization of the polyplex. In addition, the competition assay further confirms the effects of the unknown interaction between IMTP and its receptors on the transfection. The transfection significantly reduced in the presence of free IMTP under the hypoxic condition, whereas no difference was observed under the normoxia. These results demonstrate that the hypoxia triggers the changes in the location of receptors or expression level of the receptors on the cell surface. The target receptor for IMTP and the mechanism of internalization in H9C2 cells are unclear because the in vivo phage display screened the peptide sequence, but not identified the target receptor.7 A recent report proving that the exposure of H9C2 cells to hypoxia changes location of protein expression indirectly supports that hypoxia may shift the unknown receptor for IMTP to the cell membrane or increase the expression level.23 Prohibitin, for example, an evolutionarily conserved and ubiquitously expressed protein, is involved in regulation of cell cycle and homeostatic mitochondrial functions. It mainly exists in inner mitochondrial membrane and cytoplasm; however, prohibitin is shifted to the cell membrane when cells are damaged.24, 25 Prohibitin on the cell membrane is known to protect H9C2 cells under hypoxic condition and one of the proteins that show different behaviors in the diseased cells.23 Optic Atrophy-1 (OPA-1) and Dynamin-1 like protein (DLP-1) are also accepted as examples of those protein types.26, 27 Since the sequence of IMTP is obtained from in vivo phage display, in vitro transfection is not enough to prove our hypothesis. We therefore confirmed the targeting ability of IMTP-CD-9R to the ischemic myocardium of the I/R rat after systemic administration. The I/R rat model is typically prepared by ligation of left anterior descending coronary (LAD), resulting in infarction in the LV.3 High fluorescence intensity in the LV of the I/R rat represents homing of IMTP-CD-9R/DNA polyplex to the ischemic myocardium and also visualizes the occurrence of infarction in the LV due to the capability of homing peptide for targeted delivery, imaging of disease area, and diagnosis.11 IMTP-CD-9R targets only the damaged LV without any accumulation in the RV and the normal myocardium. In addition, IMTP-CD-9R increased gene expression in the LV compared to the RV of the I/R rats, lung, liver, spleen, and kidney upon a systemic injection, while negligible difference between the LV and the RV was observed in the thoracotomy group. This result is consistent with the ex vivo imaging data and suggests that IMTP-CD-9R is capable of enhancing the gene expression in ischemic myocardium. Despite the success in the targeting to ischemic myocardium and the increased gene expression in the LV of the I/R rats, identification of the receptor recognized by IMTP, precise mechanism of the internalization, and effects of therapeutic gene on the infarction remains as future challenges.
We developed an ischemic myocardium-targeted gene delivery system by conjugation of IMTP to CD polymer and reduced the required amount of polymer for transfection by attaching 9R. The resulting polymer, IMTP-CD-9R, is non-toxic to the cells and effective for gene transfection under the hypoxic condition. Finally, IMTP-CD-9R is capable of targeting to ischemic myocardium and enhancing gene expression in the LV of the I/R rat upon an intravenous injection. This study demonstrates a proof of concept for the targeted gene delivery using cationic polymer and the homing peptide to the ischemic myocardium through systemic route. Although identification of the target receptor for IMTP, the molecular mechanism of internalization, and the in vivo therapeutic efficacy need to be further investigated, the present study provides a new direction of gene therapy for the treatment of myocardial ischemia.
Acknowledgments
This work was supported by NIH grants HL065477 (SW Kim). This work was supported by a grant from the National Research Foundation in Korea funded by the Ministry of Education, Science and Technology (M. Lee, 2012K001394).
References
- 1.McGinn AN, Nam HY, Ou M, Hu N, Straub CM, Yockman JW, Bull DA, Kim SW. Bioreducible polymer-transfected skeletal myoblasts for VEGF delivery to acutely ischemic myocardium. Biomaterials. 2011;32(3):942–9. doi: 10.1016/j.biomaterials.2010.09.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yockman JW, Choi D, Whitten MG, Chang CW, Kastenmeier A, Erickson H, Albanil A, Lee M, Kim SW, Bull DA. Polymeric gene delivery of ischemia-inducible VEGF significantly attenuates infarct size and apoptosis following myocardial infarct. Gene Ther. 2009;16(1):127–35. doi: 10.1038/gt.2008.146. [DOI] [PubMed] [Google Scholar]
- 3.Won YW, Kim JK, Cha MJ, Hwang KC, Choi D, Kim YH. Prolongation and enhancement of the anti-apoptotic effects of PTD-Hsp27 fusion proteins using an injectable thermo-reversible gel in a rat myocardial infarction model. J Control Release. 2010;144(2):181–9. doi: 10.1016/j.jconrel.2010.02.014. [DOI] [PubMed] [Google Scholar]
- 4.Ko YT, Hartner WC, Kale A, Torchilin VP. Gene delivery into ischemic myocardium by double-targeted lipoplexes with anti-myosin antibody and TAT peptide. Gene Ther. 2009;16(1):52–9. doi: 10.1038/gt.2008.135. [DOI] [PubMed] [Google Scholar]
- 5.Nam HY, McGinn A, Kim PH, Kim SW, Bull DA. Primary cardiomyocyte-targeted bioreducible polymer for efficient gene delivery to the myocardium. Biomaterials. 2010;31(31):8081–7. doi: 10.1016/j.biomaterials.2010.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.McGuire MJ, Samli KN, Johnston SA, Brown KC. In vitro selection of a peptide with high selectivity for cardiomyocytes in vivo. J Mol Biol. 2004;342(1):171–82. doi: 10.1016/j.jmb.2004.06.029. [DOI] [PubMed] [Google Scholar]
- 7.Kanki S, Jaalouk DE, Lee S, Yu AY, Gannon J, Lee RT. Identification of targeting peptides for ischemic myocardium by in vivo phage display. J Mol Cell Cardiol. 2011;50(5):841–8. doi: 10.1016/j.yjmcc.2011.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pasqualini R, Ruoslahti E. Organ targeting in vivo using phage display peptide libraries. Nature. 1996;380(6572):364–6. doi: 10.1038/380364a0. [DOI] [PubMed] [Google Scholar]
- 9.Arap W, Pasqualini R, Ruoslahti E. Cancer treatment by targeted drug delivery to tumor vasculature in a mouse model. Science. 1998;279(5349):377–80. doi: 10.1126/science.279.5349.377. [DOI] [PubMed] [Google Scholar]
- 10.Hajitou A, Pasqualini R, Arap W. Vascular targeting: recent advances and therapeutic perspectives. Trends Cardiovasc Med. 2006;16(3):80–8. doi: 10.1016/j.tcm.2006.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Laakkonen P, Vuorinen K. Homing peptides as targeted delivery vehicles. Integr Biol (Camb) 2010;2(7–8):326–37. doi: 10.1039/c0ib00013b. [DOI] [PubMed] [Google Scholar]
- 12.Won YW, Yoon SM, Lee KM, Kim YH. Poly(oligo-D-arginine) with internal disulfide linkages as a cytoplasm-sensitive carrier for siRNA delivery. Mol Ther. 2011;19(2):372–80. doi: 10.1038/mt.2010.242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ou M, Wang XL, Xu R, Chang CW, Bull DA, Kim SW. Novel biodegradable poly(disulfide amine)s for gene delivery with high efficiency and low cytotoxicity. Bioconjug Chem. 2008;19(3):626–33. doi: 10.1021/bc700397x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Won YW, Kim HA, Lee M, Kim YH. Reducible poly(oligo-D-arginine) for enhanced gene expression in mouse lung by intratracheal injection. Mol Ther. 2010;18(4):734–42. doi: 10.1038/mt.2009.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kim WJ, Christensen LV, Jo S, Yockman JW, Jeong JH, Kim YH, Kim SW. Cholesteryl oligoarginine delivering vascular endothelial growth factor siRNA effectively inhibits tumor growth in colon adenocarcinoma. Mol Ther. 2006;14(3):343–50. doi: 10.1016/j.ymthe.2006.03.022. [DOI] [PubMed] [Google Scholar]
- 16.Muller OJ, Katus HA, Bekeredjian R. Targeting the heart with gene therapy-optimized gene delivery methods. Cardiovasc Res. 2007;73(3):453–62. doi: 10.1016/j.cardiores.2006.09.021. [DOI] [PubMed] [Google Scholar]
- 17.Won YW, Lim KS, Kim YH. Intracellular organelle-targeted non-viral gene delivery systems. J Control Release. 2011;152(1):99–109. doi: 10.1016/j.jconrel.2011.01.013. [DOI] [PubMed] [Google Scholar]
- 18.Wetzer B, Byk G, Frederic M, Airiau M, Blanche F, Pitard B, Scherman D. Reducible cationic lipids for gene transfer. Biochem J. 2001;356(Pt 3):747–56. doi: 10.1042/0264-6021:3560747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nam HY, Kim J, Kim S, Yockman JW, Kim SW, Bull DA. Cell penetrating peptide conjugated bioreducible polymer for siRNA delivery. Biomaterials. 2011;32(22):5213–22. doi: 10.1016/j.biomaterials.2011.03.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mitchell DJ, Kim DT, Steinman L, Fathman CG, Rothbard JB. Polyarginine enters cells more efficiently than other polycationic homopolymers. J Pept Res. 2000;56(5):318–25. doi: 10.1034/j.1399-3011.2000.00723.x. [DOI] [PubMed] [Google Scholar]
- 21.Rothbard JB, Kreider E, VanDeusen CL, Wright L, Wylie BL, Wender PA. Arginine-rich molecular transporters for drug delivery: role of backbone spacing in cellular uptake. J Med Chem. 2002;45(17):3612–8. doi: 10.1021/jm0105676. [DOI] [PubMed] [Google Scholar]
- 22.Reschel T, Konak C, Oupicky D, Seymour LW, Ulbrich K. Physical properties and in vitro transfection efficiency of gene delivery vectors based on complexes of DNA with synthetic polycations. J Control Release. 2002;81(1–2):201–17. doi: 10.1016/s0168-3659(02)00045-7. [DOI] [PubMed] [Google Scholar]
- 23.Muraguchi T, Kawawa A, Kubota S. Prohibitin protects against hypoxia-induced H9c2 cardiomyocyte cell death. Biomed Res. 2010;31(2):113–22. doi: 10.2220/biomedres.31.113. [DOI] [PubMed] [Google Scholar]
- 24.Sharma A, Qadri A. Vi polysaccharide of Salmonella typhi targets the prohibitin family of molecules in intestinal epithelial cells and suppresses early inflammatory responses. Proc Natl Acad Sci U S A. 2004;101(50):17492–7. doi: 10.1073/pnas.0407536101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Patel N, Chatterjee SK, Vrbanac V, Chung I, Mu CJ, Olsen RR, Waghorne C, Zetter BR. Rescue of paclitaxel sensitivity by repression of Prohibitin1 in drug-resistant cancer cells. Proc Natl Acad Sci U S A. 2010;107(6):2503–8. doi: 10.1073/pnas.0910649107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chen L, Gong Q, Stice JP, Knowlton AA. Mitochondrial OPA1, apoptosis, and heart failure. Cardiovasc Res. 2009;84(1):91–9. doi: 10.1093/cvr/cvp181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hom J, Sheu SS. Morphological dynamics of mitochondria--a special emphasis on cardiac muscle cells. J Mol Cell Cardiol. 2009;46(6):811–20. doi: 10.1016/j.yjmcc.2009.02.023. [DOI] [PMC free article] [PubMed] [Google Scholar]