

Abstract
Brain-derived neurotrophic factor (BDNF) has been associated with regulation of body weight and appetite. The goal of this study was to examine the interactions of a functional variant (rs6265) in the BDNF gene with dietary intake for obesity traits in the Boston Puerto Rican Health Study. BDNF rs6265 was genotyped in 1147 Puerto Rican adults and examined for association with obesity-related traits. Men (n = 242) with the GG genotype had higher BMI (P = 0.009), waist circumference (P = 0.002), hip (P = 0.002), and weight (P = 0.03) than GA or AA carriers (n = 94). They had twice the risk of being overweight (BMI ≥ 25) relative to GA or AA carriers (OR = 2.08, CI = 1.02–4.23, and P = 0.043). Interactions between rs6265 and polyunsaturated fatty acids (PUFA) intake were associated with BMI, hip, and weight, and n-3 : n-6 PUFA ratio with waist circumference in men. In contrast, women (n = 595) with the GG genotype had significantly lower BMI (P = 0.009), hip (P = 0.029), and weight (P = 0.027) than GA or AA carriers (n = 216). Women with the GG genotype were 50% less likely to be overweight compared to GA or AA carriers (OR = 0.05, CI = 0.27–0.91, and P = 0.024). In summary, BDNF rs6265 is differentially associated with obesity risk by sex and interacts with PUFA intake influencing obesity traits in Boston Puerto Rican men.
1. Introduction
Obesity is one of the fastest-growing health problems in the United States and worldwide [1]. The pathology of obesity is complex due to multiple genetic and environmental factors and their interactions [2]. BDNF, one member of the neurotrophin family of proteins, contributes to a specific type of memory inhibition function and suppresses food intake through hippocampal signaling [3, 4]. Heterozygous BDNF +/− mice with decreased BDNF expression and increased food intake exhibited hyperphagic behavior and obese phenotypes [5], while mice with BDNF knockout in the postnatal brain showed increased glucose and cholesterol concentrations [6]. Furthermore, hypothalamic overexpression of BDNF is one kind of genetic regulator, converting white fat into brown fat in adipose tissue, leading to energy dissipation and a lean phenotype [7]. Interestingly, plasma BDNF has been shown to be lower in humans with obesity and type 2 diabetes [8]. Also, a mutation in TrkB, the receptor for BDNF, was shown to be associated with an obese phenotype [9]. In addition, Yu et al. [10] found that decreased hippocampal BDNF and TrkB expression increased the risk of high-fat diet-induced obesity. Together, these findings support an important role for BDNF in energy metabolism and food intake regulation.
Recently, a missense mutation (rs6265), Val/Met (GTG/ATG), in the gene encoding BDNF was identified in a patient population with obesity [11]. BDNF (rs6265) has been associated with eating disorders [12] and obesity in several populations [13–15]. As the Met variant (A allele) was shown to be associated with abnormal intracellular packaging of the precursor of BDNF and decreased mature BDNF production in cells [16, 17], the carriers of one or two Met alleles had lower plasma BDNF than Val/Val carriers (i.e., GG homozygotes). However, rs6265 genotypes display inconsistent associations with BMI among populations. Women with Met/Met (AA) genotype had lower mean BMI than those with Val/Met (GA) or Val/Val (GG) genotypes in a general population (n = 481) [14], whereas women carrying the G allele (Val) had lower BMI compared to non-G carriers in an obese Caucasian population [13]. Furthermore, whether BDNF variants interact with environmental factors affecting obesity traits is still unknown. The Boston Puerto Rican Health Study population has demonstrated a high prevalence of obesity, diabetes, and hypertension [18]. Our aim in this study was to investigate the association between the functional BDNF variant and obesity traits and potential interactions with dietary intake for obesity traits.
2. Methods and Procedures
2.1. Study Subjects
The study population comprised 1340 Puerto Ricans (age 45–75 years), who were self-identified as Puerto Ricans living in the greater Boston metropolitan area. Participants with complete genotype and phenotypes of interest were included in this study. Demographic, dietary, medical, and social data were collected via questionnaires and biochemical analysis of blood samples that were drawn from participants. Participants were recruited by the investigators from the Boston Puerto Rican Center for Population Health, and Health Disparities to participate in a longitudinal study on stress, nutrition, health and aging—the Boston Puerto Rican Health Study (BPRHS, [18]). (http://www.northeastern.edu/cphhd/). Detailed study methods and population characteristics have been described previously [19–21]. Written informed consent was obtained from each participant and the study protocols were approved by the Institutional Review Board at Tufts University.
2.2. Dietary Assessment
The food frequency questionnaire (FFQ) was developed specifically for the Boston Puerto Rican population, originally based on the National Cancer Institute/Block FFQ. Data from the Hispanic Health and Nutrition Examination Survey were added to the NCI/Block instrument and validated through comparison with dietary recalls in Puerto Rican adults [20]. The food groups and portion sizes of the Puerto Rican population were expanded, as they differed significantly from those of the general US population. Our questionnaire captured the intakes reported in 24 h recalls more accurately for total nutrients and in ranking of individuals relative to the original NCI/Block FFQ [20, 21]. Nutrient intake profiles are estimated with NDSR, the Minnesota Nutrient Data System for Research. Intakes of total fat, total sugar, saturated fatty acids, monounsaturated fatty acids (MUFA), and PUFA are presented as percentage of total daily energy intake.
2.3. DNA Isolation and Genotyping
Genomic DNA was isolated from buffy coats of peripheral blood using QIAamp DNA Blood Mini kit (Qiagen, Hilden, Germany) according to the vendor's recommended protocol. The BDNF (rs6265) polymorphism was genotyped with the Applied Biosystems TaqMan SNP genotyping system (Applied Biosystems, Foster City, CA, USA).
2.4. Statistical Analysis
Statistic analysis was performed using SAS 9.2 software (SAS Institute, Cary, NC, USA). Associations between the BDNF variant and obesity traits (BMI, waist and hip circumference, and weight) were assessed using general linear models. For dichotomous characteristics, such as overweight (BMI ≥ 25), obesity (BMI ≥ 30), and abdominal obesity (waist circumference ≥102 cm in men, ≥88 cm in women), we used logistic regression models. Sex differences in demographic characteristics were examined using t-test. BDNF genotypes of individuals were treated as an independent variable. Associations between BDNF rs6265 and obesity traits or dietary intake were examined using general linear models after adjustment for potential confounders, including age, smoking status, alcohol use, sex, education, medication use for depression, and physical activity, and population admixture (see below). The mean values for intakes of total energy, protein, SFA, PUFA, and sugar were used as cutoffs to dichotomize these variables. Interaction between genotype and dietary intake (as dichotomized or continuous variables) was tested together with genotype and dietary intake as main effects in the same models. Differences between groups were considered significant at P ≤ 0.05.
2.4.1. Population Admixture
The presence of genetic subgroups or substructure in a population may lead to spurious associations. Thus, population admixture was estimated using Principal Component Analysis (PCA) based on 100 ancestry informative markers [22]. All analyses were adjusted for the estimated population admixture using the first major principal component in linear regression models [22].
2.4.2. Calculation of Independent Variables
To correct for multiple testing using Bonferroni test, as obesity traits or dietary intakes are highly correlated, we calculated the number of independent variables represented by obesity-related traits or dietary intakes based on their correlation matrix using a program called MatSpD [23]: http://gump.qimr.edu.au/general/daleN/matSpD/.
3. Results
3.1. BDNF Variant and Clinical Characteristics of the BPRHS Population
Characteristics of the participants in the BPRHS population and BDNF rs6265 genotype frequencies are shown in Table 1. The frequencies of GG, GA, and AA were 0.73, 0.25, and 0.02, respectively, which did not deviate from Hardy-Weinberg equilibrium expectation (P = 0.67). Men and women had similar mean age (≈57 y), but men had lower prevalence of diabetes, overweight, and obesity than women. Women had higher mean body mass index (BMI, kg/m2) and hip circumference (cm) than men, whereas men had higher body weight than women. The daily total energy intake, total protein intake, total fat intake, SFA intake, and MUFA intake were higher in men than in women (P < 0.05). Similarly, smoking and drinking alcohol were more common in men than women (34.2% versus 20.7; 76.2% versus 35.5%, resp.).
Table 1.
Population characteristic of the BPRHS participants.
| Characteristics | Men (n = 395) | Women (n = 945) |
|---|---|---|
| Mean ± s.d. | Mean ± s.d. | |
| Age, y | 56.7 ± 7.9 | 57.4 ± 7.5 |
| BMI, kg/m2 | 29.5 ± 5.0 | 32.9 ± 7.0* |
| Waist, cm | 102 ± 14.1 | 102 ± 15.4 |
| Hip, cm | 104 ± 11.0 | 111 ± 14.5* |
| Weight, kg | 83.2 ± 16.7 | 78.7 ± 17.5* |
| Physical activity score | 32.5 ± 5.8 | 31.1 ± 4.0* |
| Food intake, g/d | 3977 ± 1747 | 3392 ± 1478* |
| Energy intake, kcal/d | 2789 ± 1458 | 2112 ± 1076* |
| Total protein, g/d | 113 ± 57.6 | 88.2 ± 44.9* |
| Total fat, g/d | 102 ± 58.2 | 75.7 ± 43.1* |
| Sugar, % of energy | 18.6 ± 7.8 | 21.2 ± 8.3* |
| SFA, % of energy | 9.92 ± 2.55 | 9.50 ± 2.29* |
| MUFA, % of energy | 11.3 ± 2.16 | 11.0 ± 2.14* |
| PUFA, % of energy | 8.85 ± 2.02 | 8.72 ± 2.09 |
| n-3 : n-6 PUFA ratio | 0.09 ± 0.02 | 0.10 ± 0.03 |
| Current smoker, n (%) | 135 (34.18) | 196 (20.74)* |
| Current drinker, n (%) | 201 (76.20) | 335 (35.45)* |
| Diabetes, n (%) | 153 (38.73) | 377 (39.89)* |
| Overweight, n (%) | 322 (81.52) | 840 (88.89)* |
| Obese, n (%) | 171 (43.29) | 585 (61.90)* |
| Abdominal obesity, n (%) | 174 (44.05) | 420 (44.44) |
| BDNFrs6265 GG | 241 | 595 |
| genotype GA | 86 | 203 |
| AA | 8 | 23 |
*Indicates significant difference (P ≤ 0.05) between men and women.
3.2. Association of BDNF Variant and Obesity and Related Traits
When both men and women were combined, there were no statistically significant associations between BDNF rs6265 and obesity, such as BMI, waist circumference, hip, or body weight (Table 2). However, we found significant interactions between sex and BDNF genotype for these traits (BMI : P = 0.012; waist circumference : P = 0.005; hip: P = 0.014; weight : P = 0.003). Based on evidence of heterogeneity, we examined the association between rs6265 and obesity traits by sex. Because of low frequency of the AA genotype and the carriers of one or two A alleles (Met allele) having lower plasma BDNF than GG (Val/Val) carriers [16, 17], we thus combined the carriers of the A allele (GA + AA) into one group to increase the statistical power in subsequent analyses. In men, BDNF rs6265 was significantly associated with BMI (P = 0.009), waist circumference (P = 0.002), hip (P = 0.002), and weight (P = 0.030). The GG carriers had higher BMI, waist circumference, hip, and weight than GA and AA carriers. In women, rs6265 was significantly associated with BMI (P = 0.009), hip (P = 0.029), and weight (P = 0.027). GG carriers had lower BMI than GA and AA carriers. As these four obesity-related traits are highly correlated, they represent two independent variables (see Section 2.4.2). Thus, after correcting for multiple testing using Bonferroni correction (P = 0.05/(2 × 2) = 0.0125 adjusted for correlated traits), the associations of rs6265 with BMI, waist, and weight in men, and BMI in women remain significant. However, the association between rs6265 and waist circumference did not reach statistical significance (P = 0.071).
Table 2.
Association between BDNF rs6265 genotype and obesity traits in BPRHS.
| BDNF rs6265 | n a | BMI (kg/m2) | Waist (cm) | Weight (kg) | Hip (cm) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean ± s.e.b | P b | Mean ± s.e. | P | Mean ± s.e. | P | Mean ± s.e. | P | ||
| Total | |||||||||
| GG | 922 | 31.0 ± 0.3 | 0.213 | 101.9 ± 0.6 | 0.42 | 81.0 ± 0.7 | 0.677 | 107.4 ± 0.6 | 0.263 |
| GA | 313 | 31.8 ± 0.4 | 102.3 ± 1.0 | 81.7 ± 1.1 | 108.9 ± 0.9 | ||||
| AA | 24 | 30.1 ± 1.5 | 97.4 ± 3.6 | 78.5 ± 4.1 | 107.0 ± 3.3 | ||||
| Total | |||||||||
| GG | 922 | 31.0 ± 0.3 | 0.163 | 101.9 ± 0.6 | 0.927 | 81.0 ± 0.7 | 0.652 | 107.4 ± 0.6 | 0.103 |
| GA + AA | 337 | 31.7 ± 0.4 | 102.0 ± 1.0 | 81.5 ± 1.1 | 108.8 ± 0.9 | ||||
|
| |||||||||
| Women | |||||||||
| GG | 595 | 32.3 ± 0.3 | 0.009 | 101.1 ± 0.7 | 0.071 | 77.4 ± 0.8 | 0.029 | 110.3 ± 0.7 | 0.027 |
| GA + AA | 216 | 33.8 ± 0.5 | 103.4 ± 1.1 | 80.6 ± 1.3 | 113.4 ± 1.1 | ||||
| Men | |||||||||
| GG | 242 | 30.0 ± 0.4 | 0.009 | 103.7 ± 1.3 | 0.002 | 86.2 ± 1.5 | 0.002 | 105.8 ± 1.0 | 0.03 |
| GA + AA | 94 | 28.3 ± 0.6 | 97.7 ± 1.8 | 79.5 ± 2.1 | 102.1 ± 1.4 | ||||
a n: the sample size, bmeans and P values calculated by analysis of covariance using general linear models adjusted for age, smoking status, drinking status, sex, education, Medication use for depression, physical activity and population admixture.
To further characterize the risk of BDNF variant on obesity in the BPRHS population, we estimated the likelihood of being overweight, obese, or abdominally obese according to BDNF genotypes, using logistic regression models (Figure 1). When men and women were combined together, the risk of overweight, obese, and abdominal obesity of this population was not different between the GG carriers and GA + AA carriers (P = 0.319; P = 0.511; P = 0.655). When men and women were analyzed separately, however, men with the GG genotype had twice the risk of being overweight (OR = 2.08, CI: 1.02–4.23, and P = 0.043) relative to GA and AA carriers. In contrast, women with the GG genotype had half the risk of being overweight (OR = 0.50, CI: 0.27–0.91, and P = 0.024).
Figure 1.
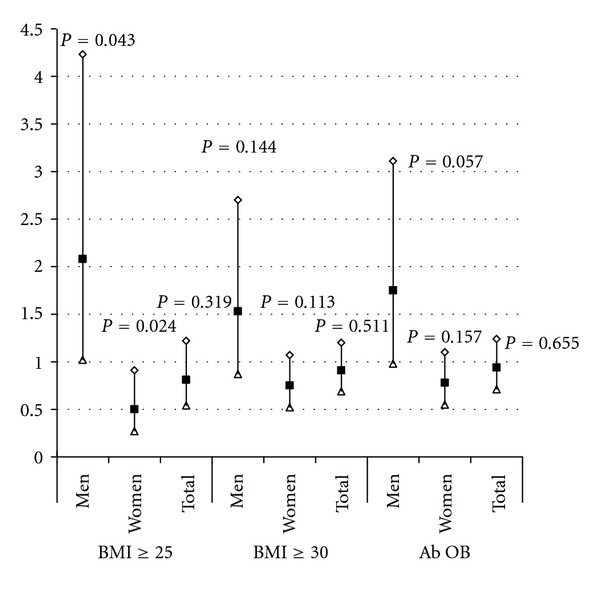
Association between BDNF genotype and overweight, obesity and abdominal obesity. P value was calculated using logistic regression models, adjusted for age, smoking status, drinking status, sex, education, medication use for depression, physical activity, and population admixture. OR = odds ratios, Lower = Lower 95% limit, Upper = Upper 95% limit, Ab OB = Abdominal obesity. For men, GG versus GA+AA, n = 242 versus 94; for women, GG versus GA + AA, n = 595 versus 216; for men and women combined, GG versus GA+AA, n = 922 versus 337 (n is the sample size).
3.3. Interaction between Dietary Intake and BDNF Genotype Modulating Obesity Traits
To determine if dietary intake modulated the association between BDNF genotype and obesity traits, we examined interactions between rs6265 and dietary intake (See supplemental Tables 1, 2, 3 in Supplementary Material available online at doi: 10.1155/2012/102942). We found that, in men but not women, BDNF genotype displayed a significant interaction with PUFA intake in relation to BMI (P = 0.042), waist circumference (P = 0.018), and hip (P = 0.009). When PUFA intake was low (<8.76% of energy), men with the GG genotype had higher BMI (P = 0.025), waist circumference (P = 0.003) and hip (P = 0.005) than did A allele carriers, whereas women with GG genotype had similar BMI and waist circumference (P > 0.05) to those with GA genotype. When PUFA intake was high (≥8.76% of energy), there was no difference in BMI, waist circumference, or hip in men, regardless of BDNF genotype, whereas women with the GG genotype had lower BMI (P = 0.008) and hip (P = 0.006) than did A allele carriers. However, genotype by PUFA intake interaction on waist circumference was not significant (P = 0.987) in women. On the other hand, BDNF genotype displayed a significant interaction with SFA intake in relation to BMI (P = 0.045) in women, but not in men. When SFA intake was low (<9.63% of energy), there was no difference in BMI, waist, and hip circumference between GG and GA + AA carriers in men or women. When SFA intake was high (≥9.63% of energy), women with the GG genotype had lower BMI (P = 0.003), and hip (P = 0.010) than A allele carriers. While the interaction between BDNF genotype and N-3 PUFA or N-6 PUFA intake did not reach significance (data not shown), the interaction between BDNF genotype and N-3 : N-6 PUFA ratio is nominally significant for hip in men (P = 0.038), but not in women (P = 0.458). However, none of the interactions passed Bonferroni test of significance (P = 0.05/(1 × 2 × 10 × 2) = 0.001) after correction for multiple testings.
To further understand the interaction between dietary components and BDNF genotype in relation to obesity traits by sex, PUFA intake as a continuous variable was plotted against the predicted BMI by sex (Figure 2). As PUFA intake increased in men, GG carriers exhibited lower BMI, whereas the GA + AA carriers exhibited higher BMI (P for interaction = 0.002). In women, however, the interaction between BDNF genotype and PUFA for BMI was not significant (P for interaction = 0.131). For N-3 : N-6 PUFA ratio evaluated as a continuous variable (Figure 3), men with the GG genotype had greater waist circumference, but GA + AA carriers tended to have smaller waist, as N-3 : N-6 PUFA ratio increased (P = 0.013). However, in women, the predicted waist remains the same regardless of the genotype with greater ratio of n-3 : n-6 PUFA ratio (P = 0.780). As depicted in Figure 4, food intake (g/d) as a continuous variable was plotted against the predicted obesity measures after adjustment for confounding factors (Figure 4). For hip circumference in men, as food intake increased, GG carriers exhibited greater predicted hip circumference, whereas GA + AA subjects showed little increase in hip (P for interaction = 0.043). In women, the interaction between BDNF genotype and food intake for hip circumference was not significant (P = 0.198), although A allele carriers showed a trend of larger hip with increased total food intake.
Figure 2.
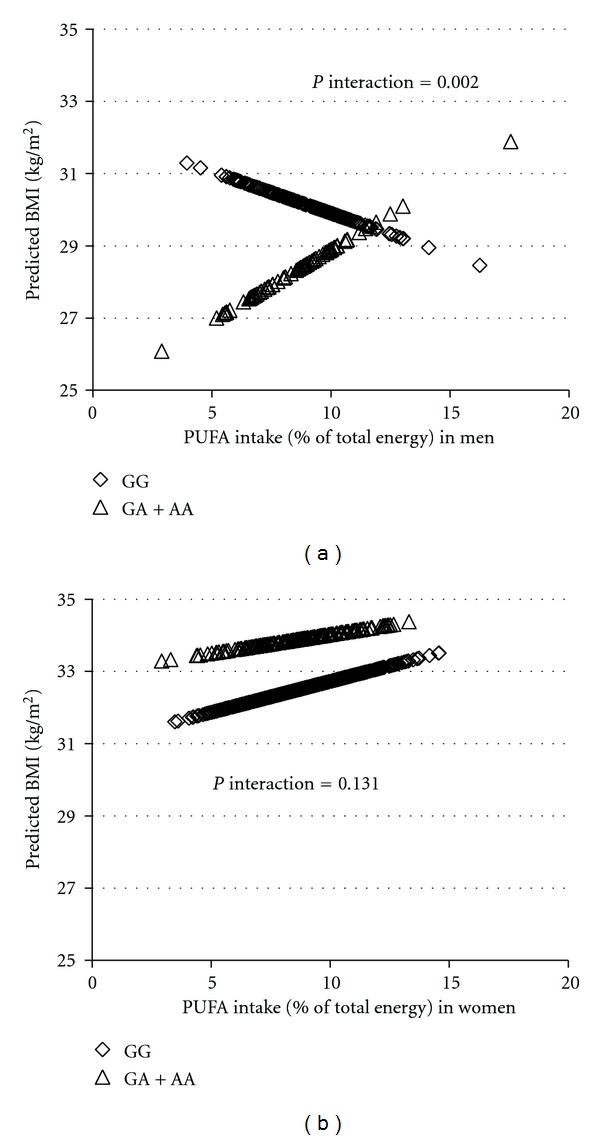
Interaction between BDNF genotypes and PUFA intake on BMI in the BPRHS. Lozenges represent the major allele GG homozygotes (n = 242 in men; n = 595 in women) and triangles the minor allele A carriers (AA homozygotes and GA heterozygotes combined together, n = 94 in men; women, n = 216 in women). Predicted BMI values for all subjects participants were plotted against their dietary PUFA intake expressed as the percentage of total energy intake. BMI values were calculated based on a regression model containing PUFA intake, BDNF genotypes, and potential confounders including age, smoking status, drinking status, sex, education, medication use for depression, physical activity, total energy, and population admixture.
Figure 3.
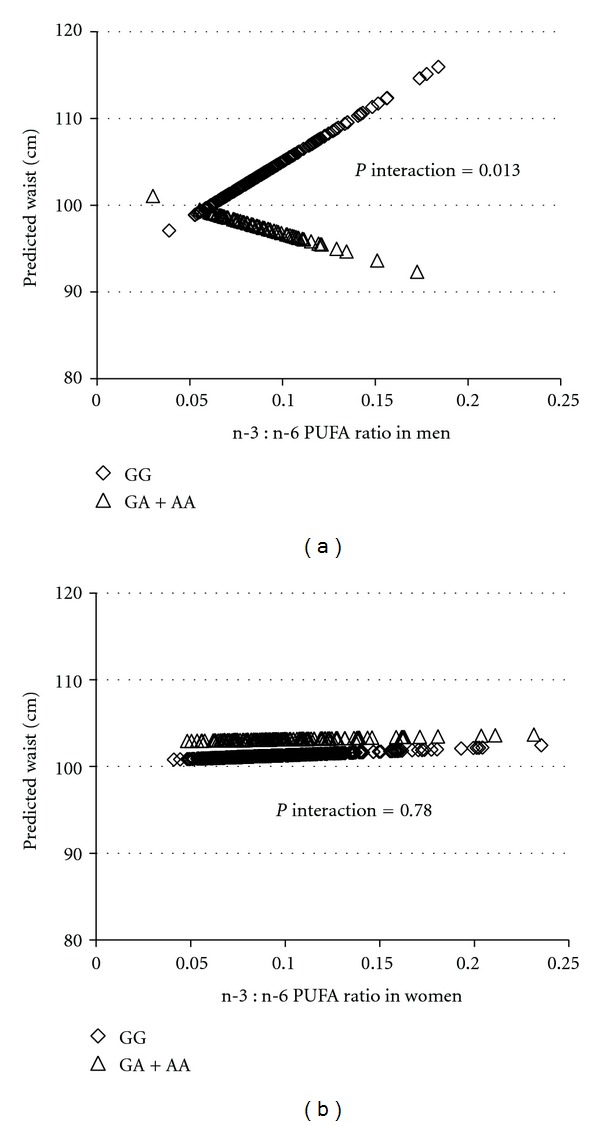
Interaction between BDNF genotype and n-3 : n-6 PUFA ratio on waist circumference in the BPRHS. Lozenges represent the major allele GG homozygotes (n = 240 in men; n = 585 in women) and triangles the minor allele A carriers (AA homozygotes and GA heterozygotes combined together, n = 92 in men; n = 215 in women). Predicted waist circumference values for all subjects were plotted against their n-3 : n-6 PUFA ratio. Waist values were calculated based on a regression model containing n-3 : n-6 PUFA ratio, BDNF genotypes, and the potential confounders including age, smoking status, drinking status, sex, education, medication use for depression, physical activity, total energy, and population admixture.
Figure 4.
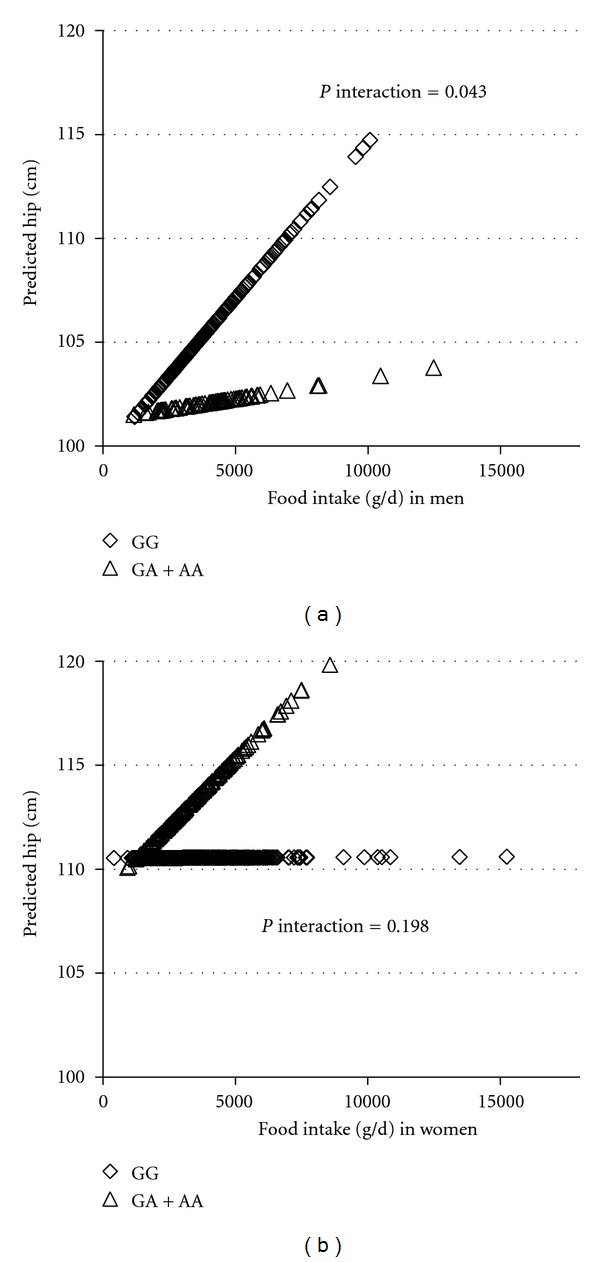
Interaction between BDNF genotype and food intake on hip in BPRHS. Lozenges represent the major allele GG homozygotes (n = 242 in men; n = 595 in women) and triangles the minor allele A carriers (AA homozygotes and GA heterozygotes combined together, n = 94 in men; n = 216 in women). Predicted hip values for all subjects were plotted against their food intake expressed as g/d. Hip values were calculated based on the regression model containing PUFA intake, BDNF genotypes, and the potential confounders including age, smoking status, drinking status, sex, education, medication use for depression, physical activity, and population admixture.
4. Discussion
4.1. Association of BDNF Genotype and Obesity Traits
This study demonstrated gender-specific associations between BDNF rs6265 and obesity traits in BPRHS. Men with the GG genotype had higher BMI, waist, hip, and weight than those with GA and AA genotypes, whereas women with the GG genotype had significantly lower BMI, hip, and weight than GA and AA carriers. In addition, BDNF rs6265 genotype interacted significantly with dietary PUFA and n-3 : n-6 PUFA ratio in relation to obesity traits of Boston Puerto Rican adults. When PUFA intake was low, men with the GG genotype had higher BMI and waist circumference than did A allele carriers. In contrast, when PUFA intake was high, there is no significant difference in BMI between genotypes.
BDNF plays an important role in the growth and maintenance of several neuronal systems [24]. In mice heterozygous for BDNF knockout, behavioral abnormalities such as intermale aggressiveness and cognitive deficiencies were accompanied by hyperphagia, hyperinsulinemia, high leptin levels, and significant weight gain [25]. These findings were confirmed and extended by Rios et al. [6] who reported that conditional deletion of BDNF in mice led to increased body weight by 80–150%, stimulated linear growth, and elevated serum of leptin, insulin, glucose, and cholesterol in blood. Similarly in humans, patients with chronic disorders such as obesity, type 2 diabetes, and dementia had low levels of plasma BDNF [26]. BDNF Val66Met SNP (rs6265), a functional variant, has been associated with obesity in several populations [11, 13]. In in vitro studies, the Met allele (A allele) was associated with abnormal intracellular packaging of the precursor of BDNF and decreased mature BDNF production [16, 17]. In population studies, carriers of one or two Met alleles had lower plasma BDNF, higher BMI, and were more likely obese than Val (G allele) carriers [27]. Thus, Met alleles may increase obesity risk through reduction of BDNF secretion. Our study tested the association between BDNF rs6265 and obesity (BMI, waist circumference, hip, and weight), but did not detect associations when men and women were combined. However, when analyses were stratified by sex, we observed that men with the GG genotype (Val/Val) had twice the risk of being overweight than GA and AA carriers, whereas women with the GG genotype had half the risk of being overweight relative to GA and AA carriers. This is in line with the result of Beckers et al. [13], who reported that women with G alleles had lower BMI than A carriers. Akkermann et al. [28] also reported that girls with G alleles had a tendency toward lower BMI than A carriers. However, Shugart et al. [29] reported that female European G allele carriers exhibited higher BMI than carriers of the A allele. Similarly, Gunstad et al. [14] found that healthy female AA carriers had lower BMI than GG and GA carriers, which differs from our results. Still, these observations suggest that sex affects the association between BDNF Val66Met and BMI [13, 30]. In addition, these inconsistent findings may be due to differences in dietary intake and other lifestyle factors, which could modify the associations between genotype and obesity traits.
4.2. Interaction between Diet Factors and BDNF Genotype on Obesity Traits
To examine the effect of dietary intake on BDNF association with obesity, we investigated interactions between BDNF genotype and dietary intakes. Previous studies in mice and humans demonstrated that BDNF was associated with the regulation of eating behavior and energy intake [11, 31]. In animal models, central and peripheral administration of BDNF decreased food intake, increased energy expenditure, ameliorated hyperglycemia, and improved serum lipid parameters in obese diabetic mice [32, 33]. Central infusion of BDNF increased serotonin (5-HT) turnover and dose-dependently suppressed appetite [34], whereas 5-HT turnover was reduced in heterozygous BDNF knock-out mice, causing excessive food intake and weight gain [5]. Further, food deprivation in mice reduced body weight and BDNF expression by 60% in the ventromedial hypothalamic nuclei, suggesting that nutritional state plays an important role in the expression of BDNF, and that BDNF may regulate appetite and body weight [35]. In our study, BDNF rs6265 variant was not associated with total food intake, total energy intake, protein intake, and PUFA intake per se (data not shown). However, BDNF rs6265 variant interacted with PUFA intake on BMI, N-3 : N-6 PUFA ratio on waist, and total food intake on hip circumference, particularly in men. In diet-induced obese mice, high fat [10] or n-3 PUFA deficient diets [36] decreased BDNF expression by over 30% in the hippocampus, whereas there was no difference in low fat diet. This implies that n-3 PUFA deficient diet or high fat diet reduces BDNF expression in the brain of mice, which leads to obesity-related phenotype. In addition, supplementation of fish oil can increase BDNF expression in the rat hippocampus and cortex [37]. In our study, high n-6 PUFA diet reflects n-3 PUFA deficient diet (n-3 : n-6 PUFA ratio is low). We found A allele carriers in men have increased BMI as PUFA intake is increased (Figure 2) and had decreased waist with increased n-3 : n-6 PUFA ratio (Figure 3). On the other hand, GG homozygous men went the opposite direction in BMI (with increased PUFA intake) or in waist (with increased n-3 : n-6 PUFA ratio). Thus, our observation that the association of BDNF genotype and obesity-related traits is modified by PUFA intakes is supported by the observations in rodents [10, 36, 37]. This may suggest that dietary n-3 PUFAs modify the association between BDNF genotype and obesity traits by regulation of BDNF expression in the brain, although this remains to be illustrated in humans. This could be one of the reasons that the association between BDNF rs6265 genotype and obesity is inconsistent among populations, as in these studies the role of gene-diet interaction was not considered. However, limitation may exist with our study. Although our FFQ was validated in terms of how it accurately captures dietary nutrient intake in general [20], n-3 or n-6 PUFA intake was not validated directly. Hence, it should be cautious when the results are generalized to other populations.
In summary, our study showed a significant association between the BDNF rs6265 variant and obesity traits in a sex-specific manner. BDNF genotype appears to interact with dietary components to influence obesity traits in the Boston Puerto Rican population, especially in men. The results support a role for BDNF in energy balance regulation and obesity. Knowledge of BDNF genotype and other genetic variants that are responsive to diet may improve our ability to reduce obesity through tailored dietary recommendations.
Supplementary Material
Supplemental Table 1: Interaction between BDNF rs6265 genotype and dietary factors on BMI in Puerto Rican adults.
Supplemental Table 2: Interaction between BDNF rs6265 genotype and nutrition factors on waist circumference in Puerto Rican adults.
Supplemental Table 3: Interaction between BDNF rs6265 genotype and nutrition factors on hip in BPRHS.
Conflict of Interests
There is no conflict of interests.
Acknowledgments
This work was supported by The Scientific and Technological Innovation Programs of Guangdong Academy of Agricultural Sciences, China (no. 2010-97), the National Institutes of Health/National Institute on Aging (Grant no. 5P01AG023394-02), the National Institutes of Health/National Heart, Lung, and Blood Institute (Grant nos. HL54776 and HL078885) and contracts 53-K06-5-10 and 58-1950-9-001 from the US Department of Agriculture Research Service. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture. The USDA is an equal opportunity provider and employer.
References
- 1.Christakis NA, Fowler JH. The spread of obesity in a large social network over 32 years. The New England Journal of Medicine. 2007;357(4):370–379. doi: 10.1056/NEJMsa066082. [DOI] [PubMed] [Google Scholar]
- 2.Maes HHM, Neale MC, Eaves LJ. Genetic and environmental factors in relative body weight and human adiposity. Behavior Genetics. 1997;27(4):325–351. doi: 10.1023/a:1025635913927. [DOI] [PubMed] [Google Scholar]
- 3.Monteleone P, Tortorella A, Martiadis V, Serritella C, Fuschino A, Maj M. Opposite changes in the serum brain-derived neurotrophic factor in anorexia nervosa and obesity. Psychosomatic Medicine. 2004;66(5):744–748. doi: 10.1097/01.psy.0000138119.12956.99. [DOI] [PubMed] [Google Scholar]
- 4.El-Gharbawy AH, Adler-Wailes DC, Mirch MC, et al. Serum brain-derived neurotrophic factor concentrations in lean and overweight children and adolescents. Journal of Clinical Endocrinology and Metabolism. 2006;91(9):3548–3552. doi: 10.1210/jc.2006-0658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lyons W, Mamounas LA, Ricaurte GA, et al. Brain-derived neurotrophic factor-deficient mice develop aggressiveness and hyperphagia in conjunction with brain serotonergic abnormalities. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(26):15239–15244. doi: 10.1073/pnas.96.26.15239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rios M, Fan G, Fekete C, et al. Conditional deletion of brain-derived neurotrophic factor in the postnatal brain leads to obesity and hyperactivity. Molecular Endocrinology. 2001;15(10):1748–1757. doi: 10.1210/mend.15.10.0706. [DOI] [PubMed] [Google Scholar]
- 7.Cao L, Choi EY, Liu XL, et al. White to brown fat phenotypic switch induced by genetic and environmental activation of a hypothalamic—adipocyte axis. Cell Metabolism. 2011;14:324–338. doi: 10.1016/j.cmet.2011.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Krabbe KS, Nielsen AR, Krogh-Madsen R, et al. Brain derived neurotrophic factor (BDNF) and type 2 diabetes. Diabetologia. 2007;50:431–438. doi: 10.1007/s00125-006-0537-4. [DOI] [PubMed] [Google Scholar]
- 9.Yeo GSH, Hung CCC, Rochford J, et al. A de novo mutation affecting human TrkB associated with severe obesity and developmental delay. Nature Neuroscience. 2004;7(11):1187–1189. doi: 10.1038/nn1336. [DOI] [PubMed] [Google Scholar]
- 10.Yu Y, Wang Q, Huang XF. Energy-restricted pair-feeding normalizes low levels of brain-derived neurotrophic factor/tyrosine kinase B mRNA expression in the hippocampus, but not ventromedial hypothalamic nucleus, in diet-induced obese mice. Neuroscience. 2009;160(2):295–306. doi: 10.1016/j.neuroscience.2009.01.078. [DOI] [PubMed] [Google Scholar]
- 11.Friedel S, Horro FF, Wermter AK, et al. Mutation screen of the brain derived neurotrophic factor gene (BDNF): identification of several genetic variants and association studies in patients with obesity, eating disorders, and attention-deficit/hyperactivity disorder. American Journal of Medical Genetics. 2005;132(1):96–99. doi: 10.1002/ajmg.b.30090. [DOI] [PubMed] [Google Scholar]
- 12.Ribasés M, Gratacòs M, Fernández-Aranda F, et al. Association of BDNF with anorexia, bulimia and age of onset of weight loss in six European populations. Human Molecular Genetics. 2004;13:1205–1212. doi: 10.1093/hmg/ddh137. [DOI] [PubMed] [Google Scholar]
- 13.Beckers S, Peeters A, Zegers D, Mertens I, Gaal LV, Van Hul W. Association of the BDNF Val66Met variation with obesity in women. Molecular Genetics and Metabolism. 2008;95(1-2):110–112. doi: 10.1016/j.ymgme.2008.06.008. [DOI] [PubMed] [Google Scholar]
- 14.Gunstad J, Schofield P, Paul RH, et al. BDNF Val66Met polymorphism is associated with body mass index in healthy adults. Neuropsychobiology. 2006;53(3):153–156. doi: 10.1159/000093341. [DOI] [PubMed] [Google Scholar]
- 15.Zhao J, Bradfield JP, Li M, et al. The role of obesity-associated loci identified in genome-wide association studies in the determination of pediatric BMI. Obesity. 2009;17(12):2254–2257. doi: 10.1038/oby.2009.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen ZY, Patel PD, Sant G, et al. Variant brain-derived neurotrophic factor (BDNF) (Met66) alters the intracellular trafficking and activity-dependent secretion of wild-type BDNF in neurosecretory cells and cortical neurons. Journal of Neuroscience. 2004;24(18):4401–4411. doi: 10.1523/JNEUROSCI.0348-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Egan MF, Kojima M, Callicott JH, et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell. 2003;112(2):257–269. doi: 10.1016/s0092-8674(03)00035-7. [DOI] [PubMed] [Google Scholar]
- 18.Tucker KL. Stress and nutrition in relation to excess development of chronic disease in Puerto Rican adults living in the Northeastern USA. Journal of Medical Investigation. 2005;52:252–258. doi: 10.2152/jmi.52.252. [DOI] [PubMed] [Google Scholar]
- 19.Lai CQ, Tucker KL, Parnell LD, et al. PPARGC1A variation associated with DNA damage, diabetes, and cardiovascular diseases the boston puerto rican health study. Diabetes. 2008;57(4):809–816. doi: 10.2337/db07-1238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tucker KL, Bianchi LA, Maras J, Bermudez OI. Adaptation of a food frequency questionnaire to assess diets of Puerto Rican and non-Hispanic adults. American Journal of Epidemiology. 1998;148(5):507–518. doi: 10.1093/oxfordjournals.aje.a009676. [DOI] [PubMed] [Google Scholar]
- 21.Tucker KL, Mattei J, Noel SE, et al. The Boston Puerto Rican health study, a longitudinal cohort study on health disparities in Puerto Rican adults: challenges and opportunities. BMC Public Health. 2010;10, article 107 doi: 10.1186/1471-2458-10-107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lai CQ, Tucker KL, Choudhry S, et al. Population admixture associated with disease prevalence in the Boston Puerto Rican health study. Human Genetics. 2009;125(2):199–209. doi: 10.1007/s00439-008-0612-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li J, Ji L. Adjusting multiple testing in multilocus analyses using the eigenvalues of a correlation matrix. Heredity. 2005;95(3):221–227. doi: 10.1038/sj.hdy.6800717. [DOI] [PubMed] [Google Scholar]
- 24.Maisonpierre PC, Belluscio L, Friedman B, et al. NT-3, BDNF, and NGF in the developing rat nervous system: parallel as well as reciprocal patterns of expression. Neuron. 1990;5(4):501–509. doi: 10.1016/0896-6273(90)90089-x. [DOI] [PubMed] [Google Scholar]
- 25.Kernie SG, Liebl DJ, Parada LF. BDNF regulates eating behavior and locomotor activity in mice. The EMBO Journal. 2000;19(6):1290–1300. doi: 10.1093/emboj/19.6.1290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pedersen BK, Pedersen M, Krabbe KS, Bruunsgaard H, Matthews VB, Febbraio MA. Role of exercise-induced brain-derived neurotrophic factor production in the regulation of energy homeostasis in mammals. Experimental Physiology. 2009;94(12):1153–1160. doi: 10.1113/expphysiol.2009.048561. [DOI] [PubMed] [Google Scholar]
- 27.Zhang XY, Zhou DF, Wu GY, et al. BDNF levels and genotype are associated with antipsychotic-induced weight gain in patients with chronic schizophrenia. Neuropsychopharmacology. 2008;33(9):2200–2205. doi: 10.1038/sj.npp.1301619. [DOI] [PubMed] [Google Scholar]
- 28.Akkermann K, Hiio K, Villa I, Harro J. Food restriction leads to binge eating dependent upon the effect of the brain-derived neurotrophic factor Val66Met polymorphism. Psychiatry Research. 2011;185(1-2):39–43. doi: 10.1016/j.psychres.2010.04.024. [DOI] [PubMed] [Google Scholar]
- 29.Shugart YY, Chen L, Day INM, et al. Two British women studies replicated the association between the Val66Met polymorphism in the brain-derived neurotrophic factor (BDNF) and BMI. European Journal of Human Genetics. 2009;17(8):1050–1055. doi: 10.1038/ejhg.2008.272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Arija V, Ferrer-Barcala M, Aranda N, Canals J. BDNF Val66Met polymorphism, energy intake and BMI: a follow-up study in schoolchildren at risk of eating disorders. BMC Public Health. 2010;10, article 363 doi: 10.1186/1471-2458-10-363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.van Oostrom I, Franke B, Rijpkema M, et al. Interaction between BDNF Val66Met and childhood stressful life events is associated to affective memory bias in men but not women. Biological Psychology. 2012;89(1):214–219. doi: 10.1016/j.biopsycho.2011.10.012. [DOI] [PubMed] [Google Scholar]
- 32.Ono M, Itakura Y, Nonomura T, et al. Intermittent administration of brain-derived neurotrophic factor ameliorates glucose metabolism in obese diabetic mice. Metabolism. 2000;49(1):129–133. doi: 10.1016/s0026-0495(00)90988-0. [DOI] [PubMed] [Google Scholar]
- 33.Tonra JR, Ono M, Liu X, et al. Brain-derived neurotrophic factor improves blood glucose control and alleviates fasting hyperglycemia in C57BLKS-leprdb/leprdb mice. Diabetes. 1999;48(3):588–594. doi: 10.2337/diabetes.48.3.588. [DOI] [PubMed] [Google Scholar]
- 34.Pelleymounter MA, Cullen MJ, Wellman CL. Characteristics of BDNF-induced weight loss. Experimental Neurology. 1995;131(2):229–238. doi: 10.1016/0014-4886(95)90045-4. [DOI] [PubMed] [Google Scholar]
- 35.Xu B, Goulding EH, Zang K, et al. Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nature Neuroscience. 2003;6(7):736–742. doi: 10.1038/nn1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rapoport SI, Rao JS, Igarashi M. Brain metabolism of nutritionally essential polyunsaturated fatty acids depends on both the diet and the liver. Prostaglandins Leukotrienes and Essential Fatty Acids. 2007;77(5-6):251–261. doi: 10.1016/j.plefa.2007.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Vines A, Delattre AM, Lima MM, et al. The role of 5-HT1A receptors in fish oil-mediated increased BDNF expression in the rat hippocampus and cortex: a possible antidepressant mechanism. Neuropharmacology. 2012;62(1):184–191. doi: 10.1016/j.neuropharm.2011.06.017. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Table 1: Interaction between BDNF rs6265 genotype and dietary factors on BMI in Puerto Rican adults.
Supplemental Table 2: Interaction between BDNF rs6265 genotype and nutrition factors on waist circumference in Puerto Rican adults.
Supplemental Table 3: Interaction between BDNF rs6265 genotype and nutrition factors on hip in BPRHS.