

Abstract
Objective: An imbalance in CD4+CD25+ regulatory T (Treg) cells and Th17 cells has been found to correlate to occurrence of acute coronary syndrome [ACS, including unstable angina (UA) and acute myocardial infarction (AMI)]. However, the mechanisms of Th17/Treg imbalance in ACS patients are still unclear. The purpose of this study is to investigate the possibility of differences in sensitivity of Th17 and Tregs to Fas-mediated apoptosis which could lead to Th17/Treg imbalance in ACS patients. Methods: We examined the apoptosis of Th17 and Treg cells, apoptosis-related Fas/Fas ligand(FasL) pathway, and inflammatory markers in patients with AMI, UA, stable angina (SA) and controls by Flow cytometry and ELISA. Then we analysed the correlation of inflammatory markers and sFasL to Treg apoptosis, and the effect of anti-FasL antibody on Treg apoptois in vitro. Results: Our study demonstrated that apoptotic Tregs, Fas and FasL expression, Caspase-3 activity of Tregs were significantly higher in ACS patients than those in NCA and SA patients (all P < 0.05). The percentage of apoptotic Tregs is positively correlated with the levels of inflammatory markers and sFasL. In vitro incubation of peripheral blood mononuclear cells from ACS patients with anti-FasL antibody resulted in a markedly reduction of apoptotic Treg cells. However, there were no significant differences in apoptotic Th17 cells and in Fas and FasL expression for Th17 cells between the four groups (all P >0.05). Conclusions: Tregs, but not Th17 cells, become apoptotic through Fas/FasL pathway, which contributed to reduction of Tregs leading to an imbalance between Th17 and Treg cells. This could be the mechanism underlying Th17/Treg imbalance and occurrence of ACS.
Keywords: Apoptosis, Fas, Fas ligand, T helper 17, regulatory T cells, acute coronary syndrome
Introduction
Atherosclerosis (AS) is a chronic inflammatory disease in which immune response plays a vital role. Various immune cells, particularly CD4+ T-helper cells (Th), participate in pathogenesis of atherogenesis [1,2]. It has been demonstrated that Th cells play an important role in coronary plaque rupture resulting in occurence of acute coronary syndrome (ACS), including unstable angina (UA) and acute myocardial infarction (AMI) [3].
Recently, CD4+CD25+ regulatory T cells (Treg) and Th17 cells have been described as two new subsets in Th cells. Treg cells have important effects on the maintenance of immune homeostasis by contact-dependent suppression and the ability to release anti-inflammatory cytokines, such as interleukin (IL)-10 and transforming growth factor (TGF)-β [4]. Th17 cells exert an important role in the pathogenesis of many autoimmune diseases and inflammatory conditions [5].
Previous studies from our group and others, both Treg cells and Th17 cells have shown to be involved in pathogenesis of atherosclerosis and ACS, and Th17/Treg imbalance appear to be critical in the development of AS, especially ACS [6,7]. However, few studies have explored the mechanisms of Th17/Treg imbalance in AS and ACS patients.
Apoptosis is an active process of cell death involving the sequential activation of a series of caspases after appropriate stimulation [8]. Apoptosis can also be induced by receptormediated or mitochondrial-mediated pathways [9]. Fas and Fas ligand (FasL) are important proapoptotic proteins, which belong to the TNFR family [10], and play essential roles in many human autoimmune diseases [11]. Activation-induced cell death (AICD) is a major mechanism of peripheral T cell apoptosis in which Fas/FasL pathway plays an important role [12,13].
Human Treg have been shown to express Fas and FasL [14]. Whether Treg are susceptible to Fas-mediated apoptosis in disease is controversial [15-17]. Recent studies have shown that Th17 cells are less sensitive than Th1 cells to Fas-mediated apoptosis [18]. The role of the Fas/Fas ligand mediated apoptosis in Th17/Treg imbalance is unknown. In this study, we examined the apoptosis of Th17 and Treg cells and their sensitivity to Fas/FasL pathway in ACS patients, and determined the differences in sensitivity of Th17 and Tregs to Fas-mediated apoptosis which may contribute to Th17/Treg imbalance and ACS occurrence.
Methods
Patients
The study was approved by the institutional review boards of Anhui Medical University and conforms with the declaration of Helsinki. All patients gave written informed consent prior enrolment into the study.
Patients at Anhui provincial hospital who underwent diagnostic catheterization between July 2008 and June 2010 (105 males and 60 females) were examined. Patients were classified into four groups: Group 1, AMI patients (23 males and 15 females, mean age 57.8±16.7 years). Myocardial infarction was confirmed by definite (>2mm) ST-segment elevations in at least two consecutive leads and significant rise of creatine kinase MB and troponin I levels. Group 2, UA patients (24 male and 16 female, mean age 63.9±12.4). UA was defined as chest pain at rest accompanied by definite ischaemic electrocardiographic changes (ST-segment changes and/or T-wave inversions). Group 3, SA patients (28 male and 14 female, mean age 60.6±11.8). SA was diagnosed by typical exertional chest discomfort associated with downsloping or horizontal ST-segment depression >1mm in an exercise test. Group 4, subjects with normal coronary arteries (NCA). Control subjects were selected on the basis of a recent angiography showing NCA (30 male and 15 female, mean age 56.7±15.9). Patients with ACS and stable angina had similar extent of coronary atherosclerosis. There were no evident differences between the four groups with regard to age.
No patient had been treated with anti-inflammatory drugs and/or immunosuppressive agents. None had thromboembolism, disseminated intravascular coagulation, advanced liver disease, renal failure, malignant disease, other inflammatory disease, chronic-immune-mediated disorders, valvular heart disease, atrial fibrillation or pacemaker.
Blood samples and cell preparation
Blood samples were obtained from all the patients in a fasting state on the following morning after admission. The time interval between symptom onset and blood sampling was less than 24 h in all cases. PB (5-10 ml) was collected from 45 NCA individuals and 120 patients. All samples were anti-coagulated with heparin and examined within 4 h. Peripheral blood mononuclear cells (PBMCs) were prepared by Ficoll density gradient for analysis of flow cytometry (FCM). Serum was obtained after centrifugation and stored at −80°C for the measurement of soluble Fas Ligand (sFasL), inflammation markers and other Biochemical indicators.
For the analysis of Th17, PBMCs were suspended at a density of 2×106 cells/ml in complete culture medium (RPMI 1640 supplemented with 100 U/ml penicillin and 100μg/ml streptomycin, 2mM glutamine and with 10% heat-inactivated fetal calf serum, Gibco BRL). The cell suspension was transferred to each well of 24-well plates. Cultures were stimulated with phorbol myristate acetate (PMA, 25 ng/ml) plus ionomycin (1μg/ml) for 4 h, in the presence of monensin (1.7μg/ml, all from Alexis Biochemicals, San Diego, CA) The incubator was set at 37 °C under a 5% CO2 environment. After culture, the contents of the well were transferred to 5ml sterile tubes. The cells were then centrifuged at 1200 rpm for 5 min. For the analysis of Treg, 100 μl of PBMCs (106) was added to tubes for further staining.
Cell staining for annexinV: apoptosis analysis in Treg and Th17 cells
Apoptotic cells were identified by staining for AnnexinV with a commercially available kit (Annexin V-FITC Apoptosis Detection Kit I, BD Pharmingen, San Diego, CA, USA). For Treg apoptosis analysis, aliquots (100μL) of mononuclear cells were incubated with 20μL of APCanti-CD4, PerCP-anti-CD25 and PE-cy7-anti-CD127 for 15 min at room temperature. Cells were then washed with PBS and suspended in 100μL binding buffer (BD Pharmingen). After incubation with 5μL FITC-AnnexinV for 15 min at room temperature, cells were resuspended in 400μL binding buffer and analyzed by flow cytometry (FCM) using FACS Aria II flow cytometer with BD FACSDiva Software (Becton Dickinson, San Jose, CA, USA).
For Th17 apoptosis analysis, the cells were incubated with APC-conjugated anti-CD4 (13B8.2 clone; Beckman Coulter-Immunotech, Marseille, France) at 4°C for 15 min. Cells were then washed with PBS and incubated with 5 μL FITC-AnnexinV for 15 min at room temperature. After fixation and permeabilization according to the manufacturer’s instructions, cells were stained with PerCP-conjugated anti-IL-17A (ebio64DEC17 clone, eBioscience).
In order to evaluate the influence of in vitro stimulation, fixation/permeabilization on cell apoptosis, CD4+ T cell apoptosis was examined among groups under pre-stimulation, poststimulation, and post-fixation/permeabilization before apoptosis analysis of Th17.
Fas and FasL expression in Treg and Th17 cells
As for Treg analysis, cell surface staining was performed by the use of isothiocyanate(FITC)-conjugated anti-Fas (DX2 clone, eBioscience, San Diego, CA, USA), phycoerythrin(PE)-conjugated anti-FasL (DOK-1 clone, eBioscience), APC-conjugated anti-CD4, peridinin chlorophy II protein (PerCP) conjugated anti-CD25 (B1.49.9 clone, eBioscience), PE-cy7-conjugated anti-CD127 (ebioRDR5 clone, eBioscience). Antibodies and appropriate isotype controls were incubated with cells for 30 min at room temperature in the dark. Cells were then washed in phosphate buffered solution (PBS).
For Th17 analysis, the cells were incubated with FITC-conjugated anti-Fas, PE-conjugated anti-FasL, APC-conjugated anti-CD4 at 4°C for 15 min, cells were fixed and permeabilized according to the manufacturer’s instructions after culture, then were stained with PerCP-conjugated anti-IL-17A (ebio64DEC17 clone, eBioscience) after fixation and permeabilization according to the manufacturer’s instructions. Stained cells were assessed by FCM. The frequency of Fas and FasL in Treg (CD4+CD25+CD127low)and Th17(CD4+IL17+)T cells was expressed as a percentage of Treg or Th17 by sequential gating on lymphocytes, CD4+ T cells, Treg or Th17 cells.
Measurement of caspase-3 activity in Treg cells
Intracellular caspase-3 activity in Treg cells was detected by active Caspase-3 apoptosis Kit (Becton Dickinson Pharmingen,). Purified PBMCs (1×106 cells/mL) were stained with APC-conjugated anti-CD4, PE-conjugated anti-CD25 (B1.49.9 clone, eBioscience), PerCP-conjugated anti-CD127(ebioRDR5 clone, eBioscience) in the dark at room temperature for 30 min, then cells were fixed and permeabilized according to the manufacturer’s instructions after washing in PBS. At last, cells were stained with FITC-conjugated monoclonal anti-active Caspase-3 antibody for 30 min, and FACS analysis was performed using the FACS Aria II.
Detection of sFasL, blood Biochemistry and inflammatory markers
The levels of sFasL in serum were examined by enzyme-linked immunosorbent assay (ELISA). According to the manufacture’s instructions (sFasL ELISA kits, from R&D Systems), and measured at 450 nm on Biocell HT1 ELISA microplate reader. The minimal detectable concentrations were 2.66 pg/ml. Intra-assay and inter-assay coefficients of variation for ELISA were <5%. All samples were measured in duplicate.
Blood sugar and lipids were determined by the enzymatic method. Inflammatory factors, high sensitive C-reactive protein (hsCRP) and Lipoprotein (a) [Lp(a)] was measured by immunoturbidimetric method. All the assays were conducted on an Olympus AU2700 biochemical autoanalyzer (Olympus, Janpan).
Analysis of the effect of anti-FasL Ab on Treg apoptosis
PBMCs (1.0 × 106 cells/ml) from AMI subjects (n = 5) were incubated with culture medium (RPMI 1640 supplemented with 10% heat-inactivated fetal calf serum), including 2μg/ml anti-FasL (Mouse IgG1, San Diego, CA, USA) for 48 h in vitro. After incubation, the apoptosis of AnnexinV+ Treg cells was measured using FCM.
Statistical analysis
Values were expressed as mean ±standard deviation (SD) in the text and figures. Data were analyzed by using statistical software (SPSS 11.0, LEAD Technologies, Inc., Chicago, IL, USA). Statistical significance for the difference in different groups was assessed by one-way analysis of variance (ANOVA). If significance was found, Bonferroni test was performed for post-hoc analysis to detect the difference among groups when equal variance were assumed, while Dunnett’s C test were performed when equal variance were not assumed. Spearman’s correlation was used as a test of correlation between two continuous variables. A probable value of P < 0.05 was considered to be statistically significant.
Results
Patient characteristics
Table 1 shows the characteristics of patients. There were no significant differences in age, gender, coronary artery disease (CAD) extent, hypertension, smoking rate, obesity, fasting blood glucose (FBG), high-density lipoprotein-cholesterol (HDL-C), and very low-density lipoprotein-cholesterol (VLDL-C) concentrations among patients with AMI, UA and SA. The Levels of TC and TG in AMI and UA groups were significantly higher than those in SA group and NCA controls (P < 0.05, P < 0.01 respectively). Besides, the Levels of low density lipoprotein cholesterol (LDL-C) and FBG in AMI and UA groups were also significantly higher than those in NCA controls. (P < 0.05, P < 0.01 respectively).
Table 1.
Patient characteristics
| Item | NCA(n=45) | SA(n=42) | UA(n=40) | AMI(n=38) |
|---|---|---|---|---|
| Age | 56.7±15.9 | 60.6±11.8 | 63.9±12.4 | 57.8±16.7 |
| Sex (male/female) | 30/15 | 28/14 | 24/16 | 23/15 |
| CAD extent (n×vessels) | 0 | 2.0±0.7 | 1.9±0.5 | 2.1±0.9 |
| Hypertension n(%) | 18(40.0) | 19(45.2) | 21(52.5) | 19(50.0) |
| Smoking rate n (%) | 10(22.2) | 11(26.2) | 12(30.0) | 10 (26.3) |
| Obesity, n (%) | 7(15.6) | 8(19.0) | 7(17.5) | 6(15.8) |
| FBG (mmol/L) | 4.95±0.52 | 5.57±0.84* | 5.52±0.89* | 5.61±0.93* |
| TC (mmol/L) | 4.39±0.67 | 4.61±0.82 | 5.10±0.78#† | 5.26±0.89# ‡ |
| TG (mmol/L) | 1.14±0.45 | 1.25±0.57 | 1.68±0.59* † | 1.86±0.53# † |
| HDL-C(mmol/L) | 1.30±0.19 | 1.27±0.23 | 1.06±0.25 | 1.12±0.28 |
| LDL-C(mmol/L) | 2.51±0.38 | 2.70±0.41 | 3.03±0.59* | 3.27±0.74# † |
| VLDL-C(mmol/L) | 0.49±0.15 | 0.45±0.17 | 0.60±0.23 | 0.56±0.21 |
Values are expressed as mean±SD. AMI: acute myocardial infarction; UA: unstable angina; SA: stable angina; NCA: subjects with normal coronary arteries; CAD: coronary artery disease. FBG: fasting blood glucose; TC: total cholesterol; TG: total triglyceride; HDL-C: high-density lipoprotein cholesterol; LDL-C: low density lipoprotein cholesterol; VLDL-C: very low density lipoprotein-cholesterol.
P<0.05 vs. NCA;
P<0.01 vs. NCA;
P<0.05 vs. SA;
P<0.01 vs. SA.
Apoptosis in Treg and Th17 cells
As shown in Figure 1, the frequencies of CD4+ CD25+ CD127low Treg cells were significantly lower in the patients with AMI and UA as compared with the NCA group and SA patients (P < 0.01), while Treg frequencies in the SA groups were also markedly lower than those of the NCA group (P < 0.05). On the contrary, the frequencies of CD4+IL-17+ Th17 cells were evidently higher in patients with AMI and UA than those with NCA and SA (P < 0.01), while there was also obvious difference between the SA and NCA group (P < 0.01).
Figure 1.
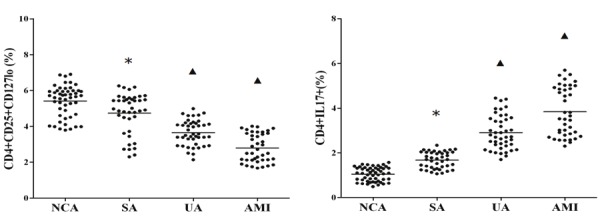
The frequencies of CD4+CD25+CD127low Treg and CD4+IL-17+Th17 cells in ACS patients. *P<0.01 vs. NCA; ▲P<0.01 vs. NCA and SA.
In addition, AnnexinV+ apoptotic Tregs in AMI and UA groups were significantly higher than those in NCA and SA patients (P < 0.01), Apoptotic Tregs in SA group were significantly higher than those in NCA patients, while Apoptotic Tregs in AMI group were also strikingly higher than those in UA patients (P < 0.05). Moreover, There were no significant differences in apoptotic Th17 cells between the four groups (all P>0.05, Figure 2A-C).
Figure 2.
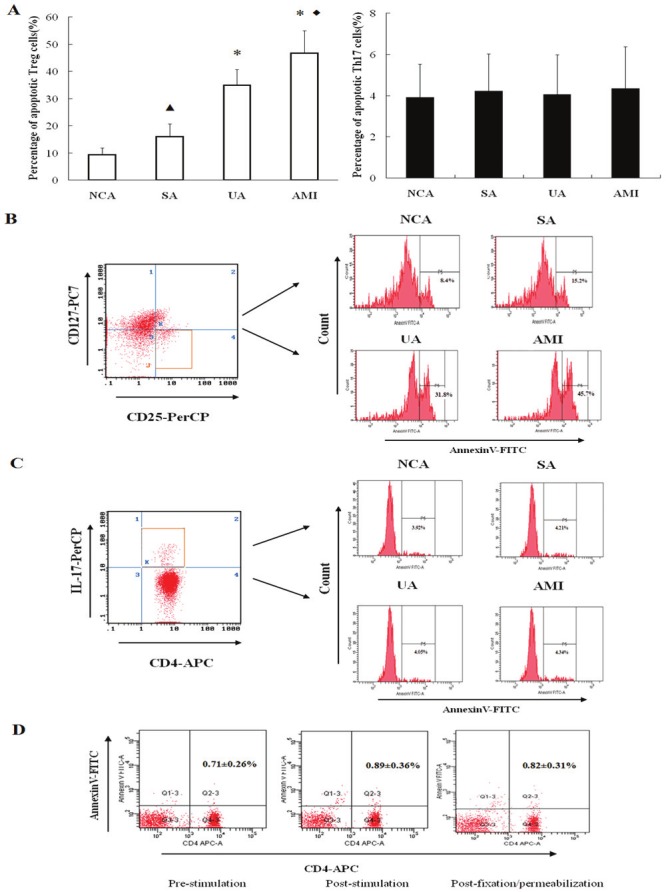
Apoptosis of Treg and Th17 cells in each group. A. Comparison of apoptotic Treg and Th17 cells in the four groups. *P<0.01 vs. NCA and SA, ▲P<0.05 vs. NCA, ♦P<0.05 vs. UA. B. Flow cytometry plots for apoptotic Treg cells (gated by CD4+CD25+CD127low cells) in each group. C. Flow cytometry plots for apoptotic Th17 cells (gated by CD4+IL17+ cells) in each group. D. CD4+T cells apoptosis among groups at pre-stimulation, post-stimulation, and post- fixation/permeabilization.
In the control test of apoptosis analysis in Th17 cells, CD4+ T cells apoptosis was not significantly increased in the post-stimulation group or the post-fixation/permeabilization group in comparison to the pre-stimulation group (P > 0.05, Figure 2D).
Expression of Fas and FasL in Treg and Th17 cells.
As shown in Figure 3A, the levels of Fas and FasL expression for Tregs were markedly higher in the AMI and UA groups than those in the SA and NCA group (P < 0.01, P < 0.05 respectively). In addition, Fas expression for Treg cells in the SA group was significantly higher than that in the NCA group, and Fas and FasL expression for Treg cells in the AMI group was also significantly higher than those in the UA group (P < 0.01 respectively). At the same time, FasL expression for Tregs in SA and NCA groups was not significantly different (P > 0.05). Interestingly, there were no significant differences in Fas and FasL expression for Th17 cells among the four groups (all P > 0.05).
Figure 3.
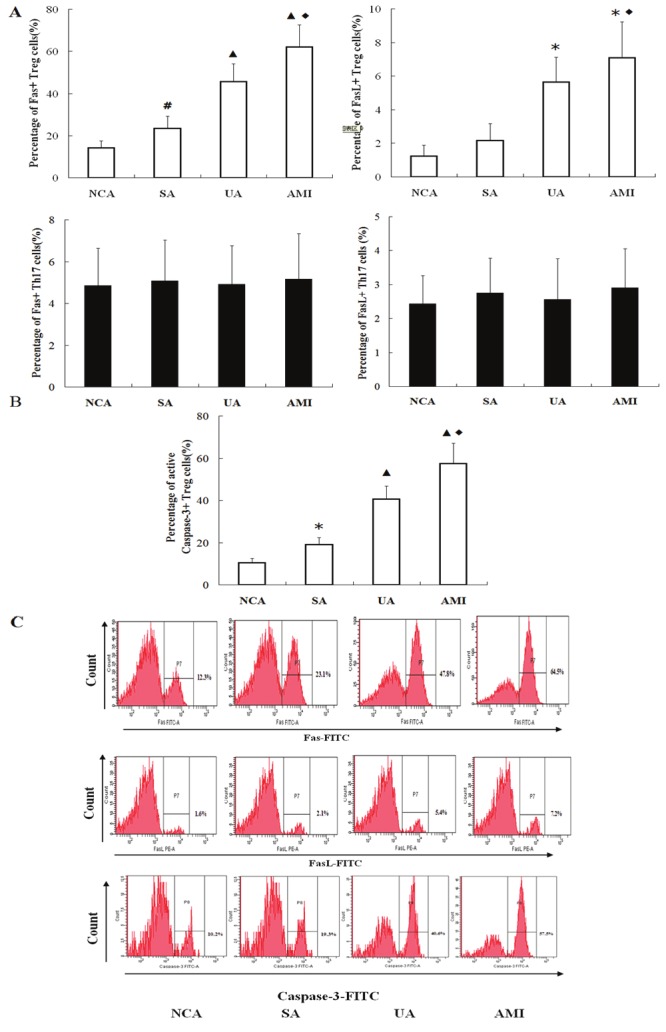
Expression of Fas, FasL and active Caspase-3 in Treg and Th17 cells for each group. A. Comparison of Fas and FasL in Tregs and Th17 among the four groups. ▲P<0.01 vs. NCA and SA, *P<0.05 vs. NCA and SA, #P<0.05 vs. NCA, ♦P<0.01 vs.UA. B. Comparison of active Caspase-3 in Tregs among the 4 groups. ▲P<0.01 vs. NCA and SA, ♦P<0.05 vs. UA, *P<0.05 vs. NCA. C. The flow cytometry plots about Fas, FasL and active Caspase-3expression in Tregs (gated by CD4+CD25+CD127low cells) from the 4 groups.
Caspase-3 activity in Treg cells
Caspase-3 activity of Treg cells in the UA and AMI groups was significantly higher than that in NCA and SA groups (P < 0.01, P < 0.05 respectively); In addition, Caspase-3 activity for Treg cells in the AMI group was significantly higher than that in the UA group, and in the SA group caspase-3 activity was also markedly higher than that in NCA group (both P < 0.05) (Figure 3B-C).
Correlation of Treg apoptosis to inflammatory markers and sFasL
The concentrations of hsCRP, Lp(a) and sFasL in the AMI and UA groups was significantly increased compared with SA and NCA groups (P < 0.01 for all). In addition, all the three indicators were significantly increased in SA group compared to the NCA group (P < 0.05) (Figure 4A).
Figure 4.
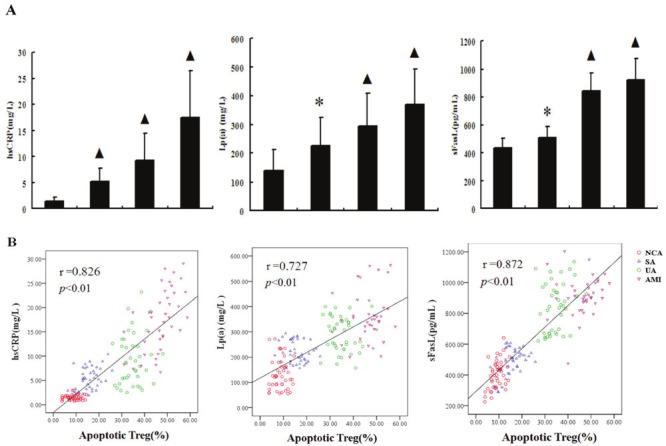
Correlation of Treg apoptosis to inflammatory markers and sFasL. A. Comparison of CRP, Lp(a) and sFasL among the four groups. ▲P<0.01 vs. NCA and SA, *P<0.05 vs. NCA. B. Spearman correlation of Treg apoptosis to the levels of inflammatory marker and sFasL.
For the four groups, inflammatory markers and sFasL had significant correlation with Treg apoptosis. The apoptotic rate of Tregs positively correlated with the concentrations of hsCRP, Lp(a) and sFasL (P < 0.01 and r=0.826,0.727, 0.872 respectively, Figure 4B).
Anti-FasL antibody reduce Treg apoptosis in ACS patients in vitro
AnnexinV+ apoptotic Treg cells in UA and AMI patients incubated with anti-FasL were markedly decreased in comparison to the group without incubation with anti-FasL after 48 h incubation in vitro (both P < 0.05). Apoptotic Treg cells in NCA and SA patients incubated with anti-FasL were slightly decreased, but this is no significant between anti-FasL group and non-anti-FasL group (both P > 0.05, Figure 5).
Figure 5.
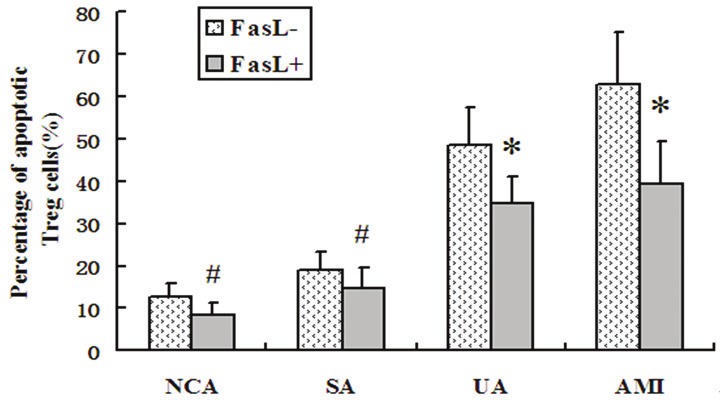
Treg apoptosis in the four groups in vitro treatment of PBMC with anti-FasL Ab for 48h. *P<0.05 vs. no-anti-FasL group, #P>0.05 vs. no-anti-FasL group.
Discussion
Treg and Th17 cells play important role in inflammation, and could participate in plaque destabilization and the onset of ACS. Our previous study demonstrated a significant increase of Th17 and related cytokines, and a decline of Treg and related cytokines in ACS patients compared to SA and NCA patients [6]. This abnormality could possibly cause Th17/Treg imbalance. In this study, we explored the mechanism underlying Th17/Treg imbalance in ACS patients by examining apoptosis and the Fas/Fas ligand pathway. ACS patients exhibited a remarkable increase in apoptotic rate, Fas/FasL expression and Caspase-3 activity of Tregs. The levels of hsCRP, Lp(a) and sFasL in ACS patients also increased. In addition, the apoptotic rate of Tregs was positively correlated with the concentrations of hsCRP, Lp(a) and sFasL in serum. At the same time, Treg apoptosis in ACS patients is decreased markedly by Anti-FasL Ab in vitro. However, there were no marked changes in apoptotic rate and Fas/FasL expression of Th17 cells among the four groups. Therefore, our findings demonstrated, for the first time, that apoptosis and Fas/Fas Ligand pathway contributes to the decrease of Tregs, not Th17, resulting in Th17/Treg imbalance and the onset of ACS.
Apoptosis plays an important part in lymphocyte development and regulation of immune response. Apoptosis disorders, both interrupted as well as accelerated apoptosis, were demonstrated as central defects in immune function, and could lead to a range of diseases [19]. Gritzapis et al [20] have demonstrated that the decrease in intratumor Tregs was due to apoptosis induced by cell contact with FasL+ Th. In other studies [18,21], Th17 cells are remarkably less susceptible to apoptosis than Th1. However, apoptosis of Treg and Th17 in ACS patients has not yet been reported.
In our study, ACS patients exhibited remarkable rise in apoptotic Tregs. Apoptotic Tregs in SA patients were significantly increased compared to those in NCA group, and those in AMI patients were also apparently elevated over those in UA patients. However, there were no significant differences in apoptotic Th17 cells between the four groups. Increased apoptosis of Treg cells may lead to relative reduction of Treg expression, and Th17/Treg imbalance. To exclude the effects of simulation and fixation/permeabilization on the results of Th17 apoptosis, CD4+ T cell apoptosis was examined pre-stimulation, post-stimulation, and post-fixation/permeabilization before Th17 apoptosis analysis. We found that the apoptotic rates of CD4+ T cells were not markedly altered in the three groups, then concluded that the process had no effects on Th17 apoptosis.
The Fas/FasL pathway plays an important role in T cell apoptosis [12,13]. Fas are activated by their natural ligands FasL. The combination of Fas and FasL on cell membrane as a trimeric complex triggers the activation cascade of caspases [22,23]. Caspases are a family of cysteine proteases in the downstream section of the Fas/FasL pathway that are necessary for the execution of apoptosis. Among them, caspase- 3 is considered to be a major executioner protease and plays a crucial role in the transduction of apoptotic signals. Activated caspase- 3 can interact with a large number of targets within an affected cell to bring about its disintegration and apoptotic bodies formation [24,25].
A study by Bossowska et al [26] has shown that a high expression of Fas and FasL on lymphocytes occurs in ACS patients. Furthermore, the results of Venet et al [27] indicate that Tregs act on monocytes by inhibiting their LPSinduced survival through a proapoptotic mechanism involving the Fas/FasL pathway. Other studies also suggest that the resistance of Th17 cells to is due to lower production of FasL, as compared to Th1 cells [18]. However, little was previously known about the sensitivity of Th17 and Treg to Fas-mediated apoptosis in ACS patients.
In our study, Treg cells in ACS patients were shown to be more sensitive to Fas-mediated apoptosis than those in NCA and SA groups, which suggested that Fas/FasL/caspase-3 pathway is activated in ACS patients. Moreover, expression of activated caspase-3 in Tregs was higher than apoptotic rates of Tregs for ACS patients, which is due to activation of caspase-3 prior to exposure of phosphatidylserine during apoptosis [28], so detection of caspase-3 activation in the cytoplasm may contribute to future prediction of apoptosis. Interestingly, the Fas/FasL pathway did not play a role in Th17 expression in ACS patients. By multiple analysis, Th17 cells expressed low level of Fas and FasL indicating no effect of Fas-mediated apoptosis on Th17 cells.
Here we found that Tregs, but not Th17 are sensitive to Fas-mediated apoptosis which may be the reason why Th17/Treg imbalance occurrs in ACS. We found that cell apoptosis and Fas-FasL apoptotic pathways could be responsible for the alteration of the fragile balance between Treg and Th17 cells, which leads to increases the risk of plaque rupture and ACS.
Apoptotic Tregs incubated with anti FasL were decreased in ACS patients incubated with anti-FasL. These results not only confirmed that Fas/FasL pathway is essential to Treg apoptosis, but also suggested that targeting Treg apoptosis may be a therapeutic strategy to maintain Th17/Treg balance in ACS, further leading to plaque stabilization. Consequently, blocking the Fas/FasL pathway and inhibiting Treg apoptotic processes could be beneficial to ACS. Treatments using anti-FasL, including caspase inhibitors, may show good results in terms of prevention and cure for ACS. However, this strategy needs to be verified in vivo by animal models.
The Th17/Treg imbalance is related to the nature of an inflammatory response. The levels of hsCRP, Lp(a) and sFasL in ACS patients increased significantly, and their levels in SA patients also elevated in comparison to NCA group. Furthermore, there was a positive association between Treg apoptosis and the concentration of inflammation markers as well as sFasL, which also seemed to be linked directly to the degree of myocardial damage and plaque destabilization.
Conclusions
Our findings show that Tregs, but not Th17 cells, give rise to significant apoptosis in ACS patients. Treg apoptosis is mediated by Fas/FasL pathway which may cause imbalance between Th17 and Treg cells, leading to plaque destabilization and the onset of ACS. We need to further prove our conclusion in a large scale of the population, and ongoing efforts should be made to explore whether other pathways is related to Treg apoptosis and Th17/Treg imbalance. Our study provides a new insight into Th17/Treg imbalance, may be useful to devise novel strategies for research on the pathogenesis and treatment of ACS.
Acknowledgments
This study was supported by National Natural Science Foundation of China (No. 30971226), Natural Science Youth Foundation of Anhui province of China (No. 11040606Q08), Natural Science Key Foundation of Anhui Universities of China (No. KJ2011A166), and Anhui Medical University Research Foundation(No. 2010XKJ083).
We thank Dr. Helen Williams from Vascular Biology Research Centre, Department of Surgery, the University of Sydney at Westmead Hospital for her assistance with grammar and for her valuable comments on the manuscript.
Disclosure
The authors declare no conflict of interest.
References
- 1.Shoenfeld Y, Sherer Y, Harats D. Artherosclerosis as an infectious, inflammatory and autoimmune disease. Trends Immunol. 2001;22:293–5. doi: 10.1016/s1471-4906(01)01922-6. [DOI] [PubMed] [Google Scholar]
- 2.Zeman K. Atherosclerosis and infection? Vnitr Lek. 2006;52:823–6. [PubMed] [Google Scholar]
- 3.Hansson GK. Inflammation, atherosclerosis, and coronary artery disease. N Engl J Med. 2005;352:1685–95. doi: 10.1056/NEJMra043430. [DOI] [PubMed] [Google Scholar]
- 4.Sakaguchi S, Sakaguchi N, Shimizu J, Yamazaki S, Sakihama T, Itoh M, Kuniyasu Y, Nomura T, Toda M, Takahashi T. Immunologic tolerance maintained by CD25+CD4+regulatory T cells: their common role in controlling autoimmunity, umor immunity, and transplantation tolerance. Immunol Rev. 2001;182:18–32. doi: 10.1034/j.1600-065x.2001.1820102.x. [DOI] [PubMed] [Google Scholar]
- 5.Miossec P, Korn T, Kuchroo VK. Interleukin-17 and type 17 helper T cells. N Engl J Med. 2009;361:888–98. doi: 10.1056/NEJMra0707449. [DOI] [PubMed] [Google Scholar]
- 6.Li Q, Wang Y, Chen K, Zhou Q, Wei W, Wang Y, Wang Y. The role of oxidized low-density lipoprotein in breaking peripheral Th17/Treg balance in patients with acute coronary syndrome. Biochem Biophys Res Commun. 2010;394:836–42. doi: 10.1016/j.bbrc.2010.03.090. [DOI] [PubMed] [Google Scholar]
- 7.Cheng X, Liao YH, Ge H, Li B, Zhang J, Yuan J, Wang M, Liu Y, Guo Z, Chen J, Zhang J, Zhang L. The Th17/Treg imbalance in patients with acute coronary syndrome. Clin Immunol. 2008;127:89–97. doi: 10.1016/j.clim.2008.01.009. [DOI] [PubMed] [Google Scholar]
- 8.hompson CB. Apoptosis in the pathogenesis and treatment of disease. Science. 1995;267:1456–62. doi: 10.1126/science.7878464. [DOI] [PubMed] [Google Scholar]
- 9.Zhang H, Xu Q, Krajewski S, Krajewska M, Xie Z, Fuess S, Kitada S, Paulowski K, Godzik A, Reed JC. BAR: an apoptosis regulator at the intersection of caspases and Bcl-2 family proteins. Proc Natl Acad Sci. 2000;97:2597–602. doi: 10.1073/pnas.97.6.2597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Griffith TS, Brunner T, Fletcher SM, Green DR, Ferguson FA. Fas ligand-induced apoptosis as a mechanism of immune privilege. Science. 1995;270:1189–92. doi: 10.1126/science.270.5239.1189. [DOI] [PubMed] [Google Scholar]
- 11.Suvannavejh GC, Dal Canto MC, Matis LA, Miller SD. Fas-mediated apoptosis in clinical remissions of relapsing experimental autoimmune encephalomyelitis. J Clin Invest. 2000;105:223–31. doi: 10.1172/JCI8561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhang Y, Xu G, Zhang L, Roberts AI, Shi Y. Th17 cells undergo Fas-mediated activation-induced cell death independent of IFN-r. J Immunol. 2008;181:190–6. doi: 10.4049/jimmunol.181.1.190. [DOI] [PubMed] [Google Scholar]
- 13.Wei Y, Chen K, Sharp GC, Yagita H, Braley-Mullen H. Expression and regulation of Fas and Fas ligand on thyrocytes and infiltrating cells during induction and resolution of granulomatous experimental autoimmune thyroiditis. J Immunol. 2001;167:6678–86. doi: 10.4049/jimmunol.167.11.6678. [DOI] [PubMed] [Google Scholar]
- 14.Strauss L, Bergmann C, Szczepanski M, Gooding W, Johnson JT, Whiteside TL. A unique subset of CD4+CD25highFoxp3+ T cells secreting IL-10 and TGF-β mediates suppression in the tumor microenvironment. Clin Cancer Res. 2007;13:4345–54. doi: 10.1158/1078-0432.CCR-07-0472. [DOI] [PubMed] [Google Scholar]
- 15.Grossman WJ, Verbsky JW, Barchet W, Colonna M, Atkinson JP, Ley TJ. Human T regulatory cells can use the perforin pathway to cause autologous target cell death. Immunity. 2004;21:589–601. doi: 10.1016/j.immuni.2004.09.002. [DOI] [PubMed] [Google Scholar]
- 16.Janssens W, Carlier V, Wu B, VanderElst L, Jacquemin MG, Saint-Remy JM. CD4+CD25+ T cells lyse antigen-presenting B cells by Fas-Fas ligand interaction in an epitope-specific manner. J Immunol. 2003;171:4604–2. doi: 10.4049/jimmunol.171.9.4604. [DOI] [PubMed] [Google Scholar]
- 17.Fritzsching B, Oberle N, Eberhardt N, Quick S, Haas J, Wildemann B, Krammer PH, Suri-Payer E. In contrast to effector T cells, CD4+CD25+Foxp3+ regulatory T cells are highly susceptible to CD95 ligand- but not to TCRmediated cell death. J Immunol. 2005;175:32–6. doi: 10.4049/jimmunol.175.1.32. [DOI] [PubMed] [Google Scholar]
- 18.Fang Y, Yu S, Ellis JS, Sharav T, Braley-Mullen H. Comparison of sensitivity of Th1, Th2, and Th17 cells to Fas-mediated apoptosis. J Leukoc Biol. 2010;87:1019–28. doi: 10.1189/jlb.0509352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Szuster-Ciesielska A, Daniluk J, Bojarska-Junak A. Apoptosis of blood mononuclear cells in alcoholic liver cirrhosis. The influence of in vitro ethanol treatment and zinc supplementation. Toxicology. 2005;212:124–34. doi: 10.1016/j.tox.2005.04.009. [DOI] [PubMed] [Google Scholar]
- 20.Gritzapis AD, Voutsas IF, Lekka E, Papamichail M, Baxevanis CN. Peptide vaccination breaks tolerance to HER-2/neu by generating vaccinespecific FasL(+) CD4(+) T cells: first evidence for intratumor apoptotic regulatory T cells. Cancer Res. 2010;70:2686–96. doi: 10.1158/0008-5472.CAN-09-2517. [DOI] [PubMed] [Google Scholar]
- 21.Tan C, Ramaswamy M, Shi G, Vistica BP, Siegel RM, Gery I. Inflammation-inducing Th1 and Th17 cells differ in their expression patterns of apoptosis-related molecules. Cell Immunol. 2011;271:210–3. doi: 10.1016/j.cellimm.2011.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Krueger A, Fas SC, Baumann S, Krammer PH. The role of CD95 in the regulation of peripheral T2 cell apoptosis. Immunol Rev. 2003;193:58–69. doi: 10.1034/j.1600-065x.2003.00047.x. [DOI] [PubMed] [Google Scholar]
- 23.Brunner T, Wasem C, Torgler R, Cima I, Jakob S, Corazza N. Fas (CD95/Apo-1) ligand regulation in T cell homeostasis, cell-mediated cytotoxicity and immune pathology. Semin Immunol. 2003;15:167–76. doi: 10.1016/s1044-5323(03)00035-6. [DOI] [PubMed] [Google Scholar]
- 24.Creagh EM, Conroy H, Martin SJ. Caspase-activation pathways in apoptosis and immunity. Immunol Rev. 2003;193:10–21. doi: 10.1034/j.1600-065x.2003.00048.x. [DOI] [PubMed] [Google Scholar]
- 25.Riedl SJ, Shi Y. Molecular mechanisms of caspase regulation during apoptosis. Nat Rev Mol Cell Biol. 2004;5:897–907. doi: 10.1038/nrm1496. [DOI] [PubMed] [Google Scholar]
- 26.Bossowska A, Bossowski A, Galar B. Analysis of apoptotic markers Fas/FasL (CD95/CD95L) expression on the lymphocytes in patients with acute coronary syndrome. Kardiol Pol. 2007;65:883–89. [PubMed] [Google Scholar]
- 27.Venet F, Pachot A, Debard AL, Bohe J, Bienvenu J, Lepape A, Powell WS, Monneret G. Human CD4+CD25+ regulatory T lymphocytes inhibit lipopolysaccharide- induced monocyte survival through a Fas/Fas ligand-dependent mechanism. J Immunol. 2006;177:6540–7. doi: 10.4049/jimmunol.177.9.6540. [DOI] [PubMed] [Google Scholar]
- 28.Bonomini M, Dottori S, Amoroso L, Arduini A, Sirolli V. Increased platelet phosphatidylserine exposure and caspase activation in chronic uremia . J Thromb Haemost. 2004;2:1275–81. doi: 10.1111/j.1538-7836.2004.00837.x. [DOI] [PubMed] [Google Scholar]