

Abstract
Objective
The adipokine vaspin (visceral adipose tissue derived serine protease inhibitor, serpinA12) follows a meal-related diurnal variation in humans and intracerebroventricular vaspin administration leads to acutely reduced food intake in db/db mice. We therefore hypothesized that vaspin may play a role in human eating behaviour.
Materials and Methods
We measured serum vaspin concentrations in 548 subjects from a self-contained population of Sorbs (Germany) who underwent detailed metabolic testing including eating behaviour assessments using the three-factor eating questionnaire. In addition, genetic variation within vaspin was assessed by genotyping 28 single nucleotide polymorphisms (SNPs) in all study subjects.
Results
Serum vaspin concentrations correlated positively with restraint, disinhibition and hunger (all P<0.05), although the correlations did not withstand further adjustments for age, gender and BMI (all P>0.05). Independent of observed correlations, genetic variants in vaspin were associated with serum vaspin levels but showed no significant association with any of the eating behaviour phenotypes after accounting for multiple testing (P≥0.05 after adjusting for age, gender and BMI).
Conclusion
Our data suggest that serum vaspin concentrations might modulate human eating behaviour, which does not seem to be affected by common genetic variation in vaspin.
Introduction
The visceral adipose tissue derived serine protease inhibitor (vaspin, serpinA12) is an adipokine potentially linking obesity, type 2 diabetes (T2D) and insulin resistance (IR). Vaspin was isolated from visceral adipose tissue of the Otsuka Long-Evans Tokushima fatty (OLETF) rat, an experimental model for T2D [1] and was postulated to have insulin sensitizing effects in state of obesity [2]. Although adipose tissue and liver have been favoured as main target organs of vaspin action [2], [3], other organs can not be excluded either. For instance, along with original studies by Hida et al. (2005) showing presence of vaspin in visceral adipose tissue [2], we detected vaspin in the hypothalamus of db/db and C57BL/6 mice as well as in the cerebrospinal fluid and in the stomach of healthy individuals [4]. Moreover, intracerebroventricular vaspin injection resulted in acutely reduced food intake and in sustained improvement of glucose concentrations in db/db mice. In line with this, circulating vaspin levels followed a meal-related diurnal variation in humans, similar to that seen for ghrelin [5] and further supporting the so far unrecognized role of vaspin in the regulation of food intake. Interestingly, variation in vaspin concentrations has a strong genetic component as implicated by recent associations between serum vaspin and genetic variants in the vaspin locus [6]–[8].
Here, we hypothesized that serum vaspin might be involved in the regulation of human eating behaviour. We therefore tested whether serum vaspin concentrations correlate with human eating behaviour factors assessed by the widely used Three-Factor Eating Questionnaire (TFEQ) [9]. To rule out any unknown confounders in a potential relationship between eating behaviour and circulating vaspin, we further analyzed the associations between human eating behaviour factors and vaspin genetic variants.
Materials and Methods
Study Subjects and the Three-Factor Eating Questionnaire
A population of the Sorbs from Eastern Germany had been extensively phenotyped for a wide range of metabolic traits as described elsewhere [10]–[12]. In total, 618 Sorbian individuals completed the German version of the TFEQ [9] measuring three different eating behaviour factors affecting food intake: restraint, disinhibition and hunger. Restraint defines the extent of individual cognitive control to limit food intake in order to control body weight (21 items) whereas disinhibition measures the loss of cognitive control resulting in overeating (16 items). Hunger represents perception of hunger or the need of food (14 items). To avoid any confounding by T2D or its treatment, subjects with T2D were excluded from the study (definition of T2D according to ADA criteria [13]). The self-reported frequency of depression in the study population is 10.8% and 7.8% of the subjects were on antidepressant medication. Only one subject (0.2%) suffered from schizophrenia but this subject was on medication. Finally, the study included 548 subjects without T2D who gave written informed consent and whose main characteristics are presented in Table 1. The study has been approved by the ethics committee of the University of Leipzig.
Table 1. Main characteristics of the study subjects (N = 548).
| female | male | total | |
| N | 346 | 202 | 548 |
| age (years) | 45±15 | 47±16 | 46±16 |
| BMI (kg/m2) | 25.7±4.9 | 26.7±3.3 | 26.1±4.4 |
| serum vaspin (ng/ml) | 0.78±3.03 | 0.33±2.57 | 0.57±3.09 |
| restraint score | 8.9±5.1 | 6.2±4.0 | 7.9±4.9 |
| disinhibition score | 4.7±3.2 | 3.7±2.5 | 4.3±3.0 |
| hunger score | 4.2±2.9 | 3.5±2.6 | 3.9±2.8 |
| lipid-lowering drugs | 7.8% | 9.4% | 8.4% |
| beta blockers | 11.6% | 13.4% | 12.2% |
| ACE inhibitors | 4.6% | 9.9% | 6.6% |
| angiotensin 2 receptor inhibitors | 5.5% | 5.4% | 5.5% |
| calcium channel blockers | 2.6% | 3.5% | 2.9% |
| antithyroid drugs | 1.2% | 0.5% | 0.7% |
| levothyroxine | 14.2% | 2.0% | 9.7% |
Means±standard deviation are given.
Measurement of Serum Vaspin
Blood samples were taken after an overnight fast and serum was separated by centrifugation. Using the enzyme-linked immunosorbent assay kit by AdipoGen (Seoul, Korea) serum vaspin concentrations were determined according to the manufactureŕs protocol [14]. The assay has a sensitivity of 12 pg/ml and the intra- and inter- assay coefficients of variance were 1.3% to 3.8% and 3.3% to 9.1%, respectively.
Genotyping of Vaspin Genetic Variants
28 representative single nucleotide polymorphisms (SNPs) have been previously genotyped to completely cover the common genetic variation within vaspin [8]. Briefly, the KASPar Assay system was used according to the manufactureŕs protocol (KBioscience Ltd, Hoddesdon, United Kingdom) and detection was done using the ABI PRISM 7500 Sequence Detecting System. To assess genotyping reproducibility, a random ∼5% selection of samples were re-genotyped for all SNPs; all genotypes matched initial designated genotypes.
Statistical Analysis
Statistical analysis was done using the PASW statistics version 20.0.1 (SPSS, Inc.; Chicago, IL). All non-normally distributed variables were logarithmically transformed to approximate normal distribution. Linear regression analysis was used to analyse the relationship of serum vaspin and eating behaviour and to assess a potential association between genetic variants and eating behaviour. Gender-genotype interaction as well as gender-vaspin-interaction was analyzed via linear regression analysis. The presented P-values were calculated in additive mode of inheritance and were adjusted for age, gender and body-mass-index (BMI). P-values <0.05 but not reaching statistical significance levels according to Bonferroni correction for multiple testing (required P<0.002 considering the 28 SNPs) were considered to be of nominal statistical significance. P-values are provided without the Bonferroni correction and effect directions are standardized to the positive strand and the minor allele.
Power calculation was done using the Quanto 1.2.4 computer program [15].
Results
Correlation of Serum Vaspin and Eating Behaviour
Serum vaspin concentrations positively correlated with restraint (P = 0.004), disinhibition (P = 0.004) and hunger (P = 0.010) (Figure 1; Table 2). After adjusting for age and BMI the correlation with restraint (P = 4.3×10−4) and disinhibition (P = 0.013) remained significant, but this did not withstand further adjustment for gender (Table 2). It is of note that medication did not affect the initially observed relationship between eating behaviour and circulating vaspin in any of the tested multivariate linear models (Table 2). Moreover, the data remained materially unchanged even after excluding subjects on medication from the analyses (data not shown).
Figure 1. Correlations of eating behaviour phenotypes with serum vaspin concentrations (N = 548).
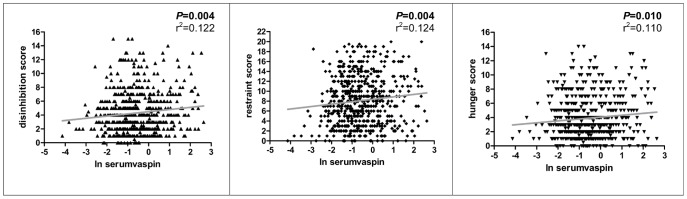
P-values were calculated using linear regression.
Table 2. Multivariate linear regression analyses of eating behaviour phenotypes with anthropometric parameters in the Sorbs (N = 548).
| restraint | disinhibition | hunger | ||||
| P-value | ß | P-value | ß | P-value | ß | |
| model 1 | ||||||
| serum vaspin | 0.004 | 0.536 | 0.004 | 0.325 | 0.010 | 0.276 |
| model 2 | ||||||
| serum vaspin | 0.001 | 0.629 | 0.045 | 0.224 | 0.082 | 0.183 |
| age | 0.001 | 0.044 | 3.7×10−9 | −0.048 | 8.3×10−9 | −0.044 |
| model 3 | ||||||
| serum vaspin | 0.001 | 0.608 | 0.001 | 0.379 | 0.009 | 0.284 |
| BMI | 1.5×10−4 | 4.897 | 7.2×10−6 | 3.584 | 0.678 | 0.314 |
| model 4 | ||||||
| serum vaspin | 0.004 | 0.525 | 0.003 | 0.336 | 0.008 | 0.284 |
| medication | 0.071 | 0.797 | 0.008 | −0.72 | 0.032 | −0.552 |
| model 5 | ||||||
| serum vaspin | 0.524 | 0.122 | 0.103 | 0.199 | 0.085 | 0.198 |
| gender | 6.8×10−9 | −2.639 | 0.005 | −0.808 | 0.063 | −0.5 |
| model 6 | ||||||
| serum vaspin | 4.3×10 −4 | 0.651 | 0.013 | 0.263 | 0.055 | 0.2 |
| age | 0.049 | 0.029 | 4.9×10−19 | −0.078 | 5.8×10−11 | −0.056 |
| BMI | 0.008 | 3.73 | 8.2×10−16 | 6.725 | 0.001 | 2.557 |
| model 7 | ||||||
| serum vaspin | 0.264 | 0.215 | 0.406 | 0.099 | 0.345 | 0.106 |
| age | 0.001 | 0.045 | 3.1×10−9 | −0.048 | 8.2×10−9 | −0.044 |
| gender | 4.4×10−9 | −2.646 | 0.004 | −0.801 | 0.06 | −0.491 |
| model 8 | ||||||
| serum vaspin | 0.359 | 0.173 | 0.053 | 0.232 | 0.078 | 0.203 |
| BMI | 4.5×10−6 | 5.777 | 1.1×10−6 | 3.882 | 0.529 | 0.477 |
| gender | 2.8×10−10 | −2.86 | 0.001 | −0.968 | 0.051 | −0.53 |
| model 9 | ||||||
| serum vaspin | 0.258 | 0.216 | 0.403 | 0.093 | 0.347 | 0.105 |
| age | 0.087 | 0.024 | 2.8×10−20 | −0.08 | 2.4×10−11 | −0.057 |
| gender | 4.8×10−10 | −2.82 | 2.9×10−5 | −1.097 | 0.018 | −0.619 |
| BMI | 0.001 | 4.786 | 9.8×10−18 | 7.136 | 0.001 | 2.788 |
| model 10 | ||||||
| serum vaspin | 0.001 | 0.647 | 0.009 | 0.279 | 0.056 | 0.202 |
| BMI | 0.009 | 3.732 | 4.5×10−16 | 6.856 | 0.002 | 2.56 |
| age | 0.088 | 0.027 | 3.7×10−15 | −0.074 | 1.7×10−9 | −0.055 |
| medication | 0.914 | 0.053 | 0.223 | −0.344 | 0.939 | −0.022 |
| model 11 | ||||||
| serum vaspin | 0.260 | 0.215 | 0.338 | 0.107 | 0.332 | 0.109 |
| age | 0.083 | 0.026 | 1.1×10−15 | −0.074 | 1.4×10−9 | −0.055 |
| gender | 2.1×10−10 | −2.905 | 1.3×10−5 | −1.156 | 0.018 | −0.627 |
| BMI | 3.6×10−4 | 4.981 | 2.5×10−18 | 7.352 | 0.001 | 2.829 |
| medication | 0.481 | −0.338 | 0.076 | −0.499 | 0.708 | −0.105 |
Data represent subjects without type 2 diabetes. P<0.05 (in bold) are presented without correction for multiple testing. BMI = body-mass index.
Association of Vaspin Genetic Variants and Eating Behaviour
To assess potential association of vaspin genetic variants with eating behaviour factors we applied linear regression models adjusted for age, gender and BMI. We observed nominal associations between disinhibition and two vaspin SNPs, rs3736806 (P = 0.025) and rs4905233 (P = 0.046; Table 3). Furthermore, rs8006968, rs17090987 and rs7152269 were nominally associated with hunger (P = 0.016, P = 0.038 and P = 0.053; Table 3). However, none of these associations withstood Bonferroni corrections for multiple testing (all P>0.002).
Table 3. Association of vaspin genetic variants with eating behaviour phenotypes (N = 548).
| restraint | disinhibition | hunger | ||||
| SNP | P-value | ß | P-value | ß | P-value | ß |
| rs17094919 C/T | 0.546 | −0.231 | 0.067 | 0.060 | 0.958 | −0.011 |
| rs17752900 C/T | 0.936 | −0.024 | 0.200 | −0.035 | 0.219 | 0.210 |
| rs76624128 T/C | 0.742 | −0.176 | 0.418 | 0.039 | 0.934 | −0.026 |
| rs74077748 G/T | 0.253 | −0.853 | 0.062 | 0.116 | 0.742 | −0.129 |
| rs8006968 T/A | 0.760 | −0.109 | 0.852 | 0.006 | 0.016 | −0.496 |
| rs17090987 C/G | 0.829 | −0.095 | 0.877 | −0.006 | 0.038 | −0.529 |
| rs1951017 T/C | 0.666 | 0.191 | 0.495 | 0.028 | 0.670 | 0.110 |
| rs17752833 C/T | 0.905 | −0.050 | 0.708 | 0.014 | 0.950 | 0.015 |
| rs7158068 T/C | 0.256 | −0.344 | 0.246 | 0.032 | 0.296 | −0.182 |
| rs10145558 A/G | 0.562 | −0.314 | 0.373 | 0.044 | 0.137 | −2.011 |
| rs1951007 T/C | 0.970 | 0.012 | 0.652 | 0.013 | 0.148 | −0.260 |
| rs1956709 A/G | 0.329 | −0.322 | 0.289 | 0.204 | 0.835 | −0.040 |
| rs8015166 T/C | 0.331 | −0.404 | 0.943 | 0.003 | 0.286 | −0.254 |
| rs10146894 A/G | 0.603 | −0.329 | 0.609 | 0.029 | 0.171 | 0.497 |
| rs12433651 A/G | 0.676 | 0.178 | 0.287 | 0.040 | 0.589 | 0.128 |
| rs11625995 A/T | 0.901 | −0.036 | 0.232 | 0.031 | 0.881 | −0.025 |
| rs11625941 T/A | 0.649 | 0.175 | 0.933 | 0.003 | 0.162 | 0.310 |
| rs4905211 G/A | 0.256 | −0.341 | 0.204 | 0.035 | 0.695 | −0.069 |
| rs1012808 A/G | 0.685 | 0.156 | 0.966 | 0.009 | 0.536 | −0.138 |
| rs2236240 G/A | 0.869 | −0.088 | 0.205 | 0.062 | 0.662 | −0.136 |
| rs7152296 G/A | 0.926 | −0.039 | 0.697 | −0.015 | 0.053 | −0.464 |
| rs2236241 T/C | 0.675 | 0.167 | 0.528 | 0.023 | 0.077 | 0.403 |
| rs2236242 A/T | 0.590 | 0.153 | 0.454 | −0.019 | 0.189 | 0.217 |
| rs1998207 C/A | 0.813 | −0.083 | 0.225 | 0.038 | 0.202 | 0.257 |
| rs3736806 A/G | 0.122 | −0.940 | 0.025 | 0.123 | 0.937 | −0.028 |
| rs4900233 G/A | 0.236 | −0.750 | 0.046 | 0.113 | 0.601 | −0.190 |
| rs3736803 G/A | 0.694 | −0.172 | 0.348 | −0.037 | 0.059 | −0.460 |
| rs11626701 G/A | 0.940 | −0.024 | 0.345 | 0.027 | 0.276 | −0.198 |
Data represent subjects without type 2 diabetes. P-values were calculated by linear regression analysis, after adjusting for age, gender and BMI in the additive mode of inheritance and are presented without correction for multiple testing. P-values <0.05 are in bold. ß (regression coefficient) is standardized to the minor allele. SNP = single nucleotide polymorphism; BMI = body mass index.
We further analyzed the gender-SNP interaction for all 3 factors, which reached statistical significance with P-values up to 2.3×10−7. Similar to the analyses in the entire cohort, gender stratified analyses did not reveal any significant association between the SNPs and eating behaviour phenotypes (Table S1).
Discussion
In the present study we hypothesized that vaspin might play a role in the regulation of human eating behaviour. We found significant correlations between circulating vaspin concentrations and eating behaviour factors assessed by the German version of the Three-Factor Eating Questionnaire [9]. However genetic variants within the vaspin gene which associate with vaspin concentrations [8] did not show significant association with eating behaviour.
Vaspin was originally identified as an adipokine potentially linking obesity, insulin resistance and T2D [2]. Based on its strong homology with alpha1-antitrypsin it belongs to broadly distributed serpins, a protein superfamily of serine protease inhibitors of ∼500 genes, and is identical to serpinA12. So far, adipose tissue has been favoured as a target organ of vaspin action, as vaspin was isolated from visceral white adipose tissue of the OLETF rat and vaspin administration to obese mice led to the reversal of altered expression of genes relevant to insulin resistance [2]. However, very recent data by Nakatsuka et al. suggest that vaspin ameliorates ER stress in liver as a ligand for cell surface GRP78/MTJ-1 complex [3]. To further expand the list of organs relevant to vaspin’s action, our present study shows a relationship between vaspin and human eating behaviour. Even though this relationship is mere association and does not allow any conclusion with regards to causality, our data are in line with recent findings in mice, where intracerebroventricular vaspin administration resulted in acute reduction of food intake and sustained amelioration of plasma glucose [4]. Furthermore, vaspin injection into the arcuate nucleus of the hypothalamus in rats decreased food intake 24 h after vaspin administration, which seemed to be mediated by decreased gene expression of the orexigenic neuropeptide Y (NPY) and increased expression of the anorexigenic POMC (proopiomelanocortin), both involved in the central modulation of feeding [16]. It is of note that vaspin has been detected in the human cerebrospinal fluid as well and the mRNA expression of vaspin has been shown in the stomach of obese human subjects after gastric sleeve resection [4]. Altogether, correlations of vaspin with eating behaviour (hunger, disinhibition and restraint) suggest a previously unrecognized role of vaspin in the cognitive control of food intake. It needs to be mentioned that the three eating behaviour factors are not entirely independent from each other. The positive correlations between eating behaviour phenotypes and serum vaspin concentrations indicate that vaspin might potentiate hunger feeling, which would consequently lead to an increased food intake. Under normal physiologic conditions the increased food intake would be under higher cognitive control mirrored by a higher restraint score. However, losing this control would lead to a higher disinhibition and would consequently further promote increased food intake.
Our findings deserve particular attention in the context of data by Jeong et al. who reported that serum vaspin concentrations raised up prior to an anticipated meal even if the study subjects remained fasted [5]. Altogether these studies support the potential neuronal function of vaspin and despite missing direct evidence one might hypothesize that vaspin degrades an anti-orexigenic factor.
It is of note however that the correlations can not be supported by the associations of genetic variants in vaspin with assessed factors of eating behaviour although we could recently show that vaspin polymorphisms were associated with circulating vaspin concentrations as well [8]. We are aware that due to the small sample size we had only limited statistical power to detect any SNP effects. Given minor allele frequencies ranging from 5–50%, we had 80% power (alpha = 0.05) to detect genotypic differences in restraint scores ranging from 11–25%, in disinhibition scores from 12–27% and hunger scores from 12–28%. Therefore, due to the lack of power, smaller effects could have been missed easily in our study. Although replication of our findings in independent cohorts would be desirable, identification of the causal variant(s) controlling circulating vaspin levels will still be inevitable to establish a causative link between serum vaspin, eating behaviour and genetic variation.
In conclusion, we show that circulating vaspin correlates with human eating behaviour, which seems to be unaffected by genetic variation within the vaspin gene. Our data suggest that the previously reported association between circulating vaspin and metabolic traits might be mediated by vaspin effects on human eating behaviour.
Supporting Information
Gender-stratified vaspin-SNP associations with eating behaviour phenotypes (N = 548).
(DOC)
Acknowledgments
We thank all those who participated in the studies. We would like to acknowledge excellent technical assistance by Beate Enigk and Manuela Prellberg.
Funding Statement
This work was supported by grants from the Federal Ministry of Education and Research (BMBF), Boehringer Ingelheim Foundation (to P.K. and D.S.), Integrated Research and Treatment Center IFB Adiposity Diseases, K7-38, K7-39 (to Y.B.) and K7-37 and K403 (to A.T.), Deutsche Forschungsgemeinschaft (DFG), the Clinical Research Group “Atherobesity” KFO 152 (projects BL 833/1-1 to M.B., and Stu192/6-1 (MS), KO 3512/1-1 to A.K., KO 3880/1-2 to P.K., M.B. and J.B.), from the German Diabetes Association (to D.S. and P.K.), from the DDS Foundation to Y.B., DHFD (Diabetes Hilfs- und Forschungsfonds Deutschland; to M.S., P.K., M.B., A.T.), and LIFE – Leipzig Research Center for Civilization Diseases, Universität Leipzig. LIFE is funded by means of the European Union, by the European Regional Development Fund (ERDF) and by means of the Free State of Saxony within the framework of the excellence initiative. IFB Adiposity Diseases is supported by the Federal Ministry of Education and Research (BMBF), Germany, FKZ: 01EO1001. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Kawano K, Hirashima T, Mori S, Saitoh Y, Kurosumi M, et al. (1992) Spontaneous Long-Term Hyperglycemic Rat with Diabetic Complications - Otsuka Long-Evans Tokushima Fatty (Oletf) Strain. Diabetes 41: 1422–1428. [DOI] [PubMed] [Google Scholar]
- 2. Hida K, Wada J, Eguchi J, Zhang H, Baba M, et al. (2005) Visceral adipose tissue-derived serine protease inhibitor: A unique insulin-sensitizing adipocytokine in obesity. Proc Natl Acad Sci U S A 102: 10610–10615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nakatsuka A, Wada J, Iseda I, Teshigawara S, Higashio K, et al.. (2012) Vaspin Is an Adipokine Ameliorating ER Stress in Obesity as a Ligand for Cell-Surface GRP78/MTJ-1 Complex. Diabetes; DOI: 10.2337/db12–0232. [DOI] [PMC free article] [PubMed]
- 4. Klöting N, Kovacs P, Kern M, Heiker JT, Fasshauer M, et al. (2012) Central vaspin administration acutely reduces food intake and has sustained blood glucose-lowering effects. Diabetologia 54: 1819–1823. [DOI] [PubMed] [Google Scholar]
- 5. Jeong E, Youn BS, Kim DW, Kim EH, Park JW, et al. (2010) Circadian Rhythm of Serum Vaspin in Healthy Male Volunteers: Relation to Meals. J Clin Endocrinol Metab 95: 1869–1875. [DOI] [PubMed] [Google Scholar]
- 6.Teshigawara S, Wada J, Hida K, Nakatsuka A, Eguchi J, et al.. (2012) Serum Vaspin Concentrations Are Closely Related to Insulin Resistance, and rs77060950 at SERPINA12 Genetically Defines Distinct Group with Higher Serum Levels in Japanese Population. J Clin Endocrinol Metab 97:DOI: 10.1210/jc.2011–3297. [DOI] [PubMed]
- 7. Kempf K, Rose B, Illig T, Rathmann W, Strassburger K, et al. (2010) Vaspin (SERPINA12) Genotypes and Risk of Type 2 Diabetes: Results from the MONICA/KORA studies. Exp Clin Endocrinol Diabetes 118: 184–189. [DOI] [PubMed] [Google Scholar]
- 8.Breitfeld J, Tönjes A, Böttcher Y, Schleinitz D, Wiele N, et al.. (2012) Genetic variation in the vaspin gene affects circulating serum vaspin concentrations. Int J Obes (Lond) DOI: 10.1038/ijo.2012.133. [DOI] [PubMed]
- 9.Pudel V, Westenhöfer J (1989) Fragebogen zum Eßverhalten (FEV). Handanweisung. Göttingen: Hogrefe.
- 10. Tönjes A, Zeggini E, Kovacs P, Böttcher Y, Schleinitz D, et al. (2010) Association of FTO variants with BMI and fat mass in the self-contained population of Sorbs in Germany. Eur J Hum Genet 18: 104–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Tönjes A, Koriath M, Schleinitz D, Dietrich K, Böttcher Y, et al. (2009) Genetic variation in GPR133 is associated with height: genome wide association study in the self-contained population of Sorbs. Hum Mol Genet 18: 4662–4668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Veeramah KR, Tönjes A, Kovacs P, Gross A, Wegmann D, et al. (2011) Genetic variation in the Sorbs of eastern Germany in the context of broader European genetic diversity. Eur J Hum Genet 19: 995–1001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. ADA (2010) Standards of Medical Care in Diabetes-2010. Diabetes Care 33: S11–S61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Youn BS, Klöting N, Kratzsch J, Lee N, Park JW, et al. (2008) Serum vaspin concentrations in human obesity and type 2 diabetes. Diabetes 57: 372–377. [DOI] [PubMed] [Google Scholar]
- 15.Gauderman WJ, Morrison JM (2006) QUANTO 1.1: A computer program for power and sample size calculations for genetic-epidemiology studies, http://hydra.usc.edu/gxe.
- 16. Brunetti L, Di Nisio C, Recinella L, Chiavarolo A, Leone S, et al. (2011) Effects of vaspin, chemerin and omentin-1 on feeding behavior and hypothalamic peptide gene expression in the rat. Peptides 32: 1866–1871. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Gender-stratified vaspin-SNP associations with eating behaviour phenotypes (N = 548).
(DOC)