

Abstract
The metabolic pathways of methane formation vary with environmental conditions, but whether this can also be linked to changes in the active archaeal community structure remains uncertain. Here, we show that the suppression of aceticlastic methanogenesis by methyl fluoride (CH3F) caused surprisingly little differences in community composition of active methanogenic archaea from a rice field soil. By measuring the natural abundances of carbon isotopes we found that the effective dose for a 90% inhibition of aceticlastic methanogenesis in anoxic paddy soil incubations was <0.75% CH3F (v/v). The construction of clone libraries as well as t-RFLP analysis revealed that the active community, as indicated by mcrA transcripts (encoding the α subunit of methyl-coenzyme M reductase, a key enzyme for methanogenesis), remained stable over a wide range of CH3F concentrations and represented only a subset of the methanogenic community. More precisely, Methanocellaceae were of minor importance, but Methanosarcinaceae dominated the active population, even when CH3F inhibition only allowed for aceticlastic methanogenesis. In addition, we detected mcrA gene fragments of a so far unrecognised phylogenetic cluster. Transcription of this phylotype at methyl fluoride concentrations suppressing aceticlastic methanogenesis suggests that the respective organisms perform hydrogenotrophic methanogenesis. Hence, the application of CH3F combined with transcript analysis is not only a useful tool to measure and assign in situ acetate usage, but also to explore substrate usage by as yet uncultivated methanogens.
Introduction
Methanogenesis is the dominating terminal process in anoxic freshwater habitats like sediments and flooded soils. In rice fields, most labile organic carbon is derived from plant material, and carbohydrates are the primary source for anaerobes resulting eventually in acetate and H2 + CO2 as most important methanogenic precursors [1]. The theoretical ratio of acetate : H2 + CO2 usage equals 2 : 1 [2]. However, depending on the exact oxidation state of labile organic carbon, but also on competing microbial processes, this ratio may vary. Hence, the fraction of methane produced via acetate is an important variable in understanding what controls mineralization in anoxic environments.
The amount of acetate-derived methanogenesis can be assessed with CH3F (methyl fluoride, fluoromethane), a specific inhibitor for aceticlastic methanogenesis. When applied for the first time in microbial ecology, CH3F was assumed to be a specific inhibitor for methane oxidation and ammonium oxidation [3], [4]. While providing direct access to processes, inhibitor experiments may be misleading, if specificity is confined to certain conditions [5]. Indeed, CH3F turned out to be an efficient inhibitor of methane and ammonium monooxygenases. However, it soon became evident that it may also inhibit methanogenesis [6], [7]. In anoxic incubations treated with CH3F, approximately as much acetate accumulates as methane is lacking compared to untreated controls. Selectivity of CH3F for suppression of aceticlastic methanogenesis was further validated in pure culture studies demonstrating that 1% v/v inhibited growth of and methanogenesis by pure cultures of aceticlastic Methanosaeta and Methanosarcina. Other microbes, homoacetogenic, sulfate reducing and fermentative bacteria, and a methanogenic mixed culture based on hydrogen syntrophy, were not inhibited [7]. In Methanosarcina barkeri, which is able to use acetate and H2 + CO2 simultaneously, only acetate utilization was suppressed, when both acetate and hydrogen were supplied [7]. However, pure cultures are not necessarily representative for yet uncultured populations, and many operational taxonomic units (OTUs) have been designated to a phylogenetic clade and named from environmental sequence information alone. Hence, some populations may show a behavior different from that found in pure cultures.
Another approach to determine methanogenic pathways uses isotopic signatures; for review see [8]. In short, methanogenesis from H2 + CO2 discriminates stronger against isotopically heavier carbon than does aceticlastic methanogenesis [8], [9]. This difference can be used to calculate the contribution of these two methanogenic pathways, provided the respective isotopic fractionation factors are known [8], [10], [11]. Indeed, combining the application of CH3F with the analysis of isotopic signatures revealed the expected patterns [12].
The methanogenic community in rice fields mainly consists of versatile Methanosarcinaceae and strictly acetotrophic Methanosaetaceae, as well as of hydrogenotrophic Methanomicrobiales, Methanobacteriales, and Methanocellales; the latter were formerly known as rice cluster I [1], [8], [12]–[14]. Rice paddy soil is found to be compartmented into two habitats: rhizosphere and bulk soil. Methanogenic communities on rice roots are dominated by Methanocellales, with hydrogenotrophic methanogenesis contributing 60–80% to total methane production [15]–[17]. The influence of rice cultivars was found to be minor [18]. In bulk soil however, methane is mainly derived from acetate (50–83%), and Methanosarcinaceae are the prevailing methanogens [19], [20]. The community structure of methanogens remains rather stable even under dry-wet cycles [21]. In summary, cell numbers fluctuate with management [21], but methanogenic communities in paddy fields of different geographical origin are highly related [22].
Here, we re-visit the inhibition of aceticlastic methanogenesis in a paddy soil asking not only how specifically CH3F inhibits aceticlastic methanogenesis, but also for the response of different methanogenic archaea to this inhibitor. We studied the dose-response relationship of methanogenesis as a function of CH3F concentration by combining process measurements with isotopic data and molecular analyses targeting the mcrA gene (encoding the subunit A of methyl coenzyme M reductase, a protein characteristic and essential for methanogenesis [23]). Since quite often only a minor fraction of a methanogenic community is metabolically active [16], [25], [26], we aimed at both the mcrA gene (community) and the respective mRNA (active community), as mcrA transcripts have been shown to be directly connected to energy metabolism and methanogenesis [24].
Materials and Methods
One kg bulk soil was sampled in spring 2008 from a rice field in the delta region of River Yangtze (Zhejiang Province, China) representing one of the major rice growing areas of the world. The particular field had been used for wetland rice production for about 2000 years [27]–[29]. Ten grams air-dried soil were mixed with ten milliliters oxygen-free distilled water in 26-ml pressure tubes. Tubes were capped with butyl rubber stoppers and flushed with N2 for ten minutes. Different amounts of CH3F corresponding to initial concentrations of 0.2, 0.3, 0.4, 0.6, 0.79, 0.99, 1.19, 1.57, 1.96, 2.72 and 3.85% were injected by syringe in two tubes each. Another three tubes did not receive CH3F serving as control, and three were sampled immediately as primary soil material. Water, tubes, and stoppers had been sterilized. The tubes were incubated for14 days in the dark at 25°C. Methane, carbon dioxide and methyl fluoride in the headspace were measured repeatedly after sampling with a 0.25-ml pressure-lok syringe (Valco Instruments, USA) on a GC-FID (SRI-8610, SRI Instruments, USA). Only endpoint measurements are shown here. Quantification of lactate, formate, acetate, propionate, ethanol and butyrate were performed by analyzing filtered (ReZist, 0.2 µm PTFE, Schleicher and Schuell, Germany) pore water samples after 14 days of incubation by HPLC (SRI Instruments, USA).
Methane produced from carbon dioxide (mCO2) was measured under inhibition of aceticlastic methanogenesis (≥0.75% CH3F, see below), while methane produced from acetate (macetate) was calculated from the balance to total methane produced in controls without inhibitor: macetate = mtotal−mCO2.
Carbon isotopic signatures in methane and acetate were measured as described elsewhere [30]. 13C signatures are given in δ-notation referring to the respective standard material, Vienna Pee Dee Belemnite (VPDB) [8].
Total nucleic acids were extracted as described elsewhere [31]. For tRFLP analysis, mcrA gene fragments were obtained with primers ME1/ME2 [32], where the forward primer was labeled with FAM. PCR conditions were: initial denaturing at 94°C for 5 minutes, 35 cycles of 30 s at 94°C, 45 s at 55°C, 1.5 min at 72°C, and a final extension at 72°C for 5 min. Amplicons were digested with SAU96I and analyzed on a capillary sequencer (3130 Genetic Analyzer, Applied Biosystems). For reverse transcriptase PCR (RT-PCR), 5 µl sample were treated with DNA-free DNase (Qiagen) followed by exonuclease treatment (mRNA-ONLY Prokaryotic mRNA Isolation Kit, Epicentre Technologies) and cleaning (RNAeasy Mini Kit, Qiagen) according to manufacturers' instructions. Reverse transcription and amplification was performed in one step combining reverse transcription (Reverse Transcription System, Promega, Germany) with 30 PCR cycles at conditions as described above, but without a FAM-label on primer ME1.
In tRFLP analysis measured fragment size may deviate from real (in silico) size. Different factors have been claimed to be responsible for size shifts [33], [34], but a detailed residual analysis was lacking so far. Residuals, the difference between real and estimated size, were calculated by running a FAM-labeled size standard as ‘sample’ against a ROX-labeled size standard. Both standards were purchased from Eurogentec (Germany). The ‘fragment’ size of the FAM-labeled standard was calculated with the built-in software using a third order polynomial as calibration function. Even if the calibration curve gave nearly perfect fit, residuals showed a considerable non-linearity being best described by a fifth order polynomial (Figure 1; intercept = 16.67359, a = −0.3238648, b = 1.831838e-3, c = −3.81772e-06, d = 3.17735e-09, e = −8.61187e-13). This polynomial was used to correct measured TRF size making it comparable to in-silico fragment size.
Figure 1. Residuals, the difference between real and estimated size, of a FAM-labeled size standard used as ‘sample’ in t-RFLP analysis.
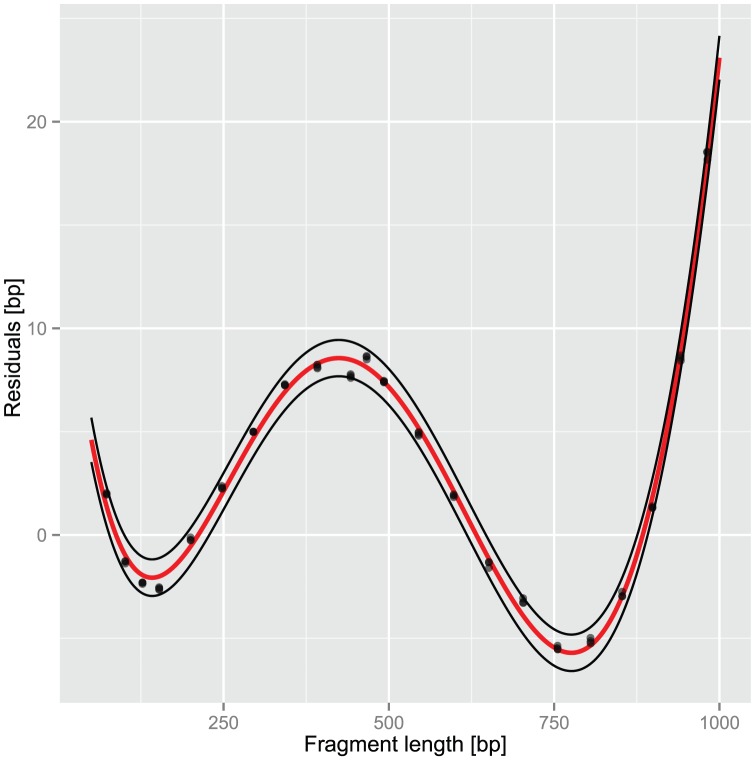
Data from three replicate runs are shown. Fit: fifth order polynomial, red line; 95% prediction intervals: black lines.
Gene libraries for archaeal mcrA sequences were constructed using cDNA from the control samples and from samples incubated under 3.85% methyl fluoride, as well as DNA from the primary soil material. (RT)-PCR products were ligated into pGEM-T vector plasmids (Promega, Germany) and transformed into Escherichia coli competent cells JM109 (Promega, Germany) according to the manufactures' instructions. The sequences were assembled with SeqManII (DNASTAR) and compared with sequences available in the GenBank database using the BLAST network service to determine the approximate phylogenetic affiliations. Alignment and phylogenetic analysis of the mcrA sequences from 69 DNA- and 91 mRNA-derived clones was done with ARB [35]. OTUs were defined by the average neighbor algorithm at 5% amino acid sequence divergence level; representative sequences for these OTUs were determined using mothur ver. 1.19.3 [36]. Sequence data have been submitted to GenBank under accession numbers JQ283291-JQ283438.
Statistical analysis was done in R ver. 3.12.2 [37]. Dose-response models were fitted using package drc, ver. 2.2-1 [38]. Constrained correspondence analysis (CCA) and non-metric multidimensional scaling (NMDS) were done with package vegan ver. 2.1-0 [39], and a multivariate regression tree (MRT) was fitted with package mvpart ver. 1.4-0 [40]. Graphics were produced with package ggplot2 [41].
Results and Discussion
Metabolites and isotopic signatures
With increasing CH3F concentration, acetate accumulated while methane accumulation was reduced accordingly (Figure 2A) resulting in a highly significant negative correlation (r = 0.7, P = 0.0002). No other fermentation products, in particular not formate, propionate, butyrate, or ethanol, did accumulate (data not shown). Along with the reduction of methanogenesis, both the δ 13C values of methane and acetate decreased (Figure 2B). The shift in δ 13C-CH4 by about −20‰ VPDB between control (0% CH3F) and incubations receiving ≥0.75% CH3F is in accordance with a shift from mixed substrate usage to H2 + CO2 dependent methanogenesis [17], [42]. Correspondingly, the relatively heavy carbon isotopic signature of −10‰ in acetate from control incubations implies that lighter acetate was preferentially consumed, thus enriching the remaining acetate in 13C. With increasing CH3F concentration, δ13Cacetate continuously decreased until values stabilized around −23‰, as known for acetate derived from fermentation of organic matter in rice fields [12]. Thereby we can exclude that homoacetogenesis was an important process in the incubations, as otherwise the isotopic signature of acetate should have been substantially lower [43].
Figure 2. Accumulation of acetate and methane (A), and the respective δ13C signatures in ‰ VPDB (B) depending on initial concentrations of methyl fluoride; δ13Cacetate is the combined signature for both C-atoms.
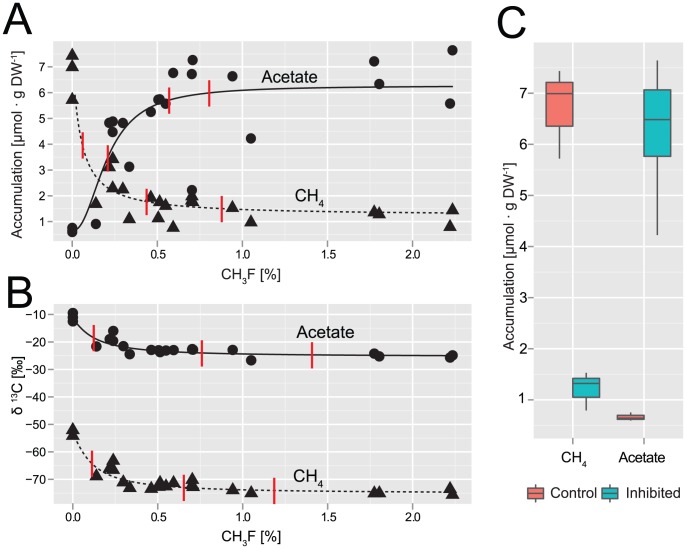
Data are endpoint measurements and not corrected for initial concentrations. The fitted dose-response curves follow a log-logistic model with the parameters ED50 (effective dose for 50% inhibition), upper limit, and slope, while the lower limit was fixed to the respective averages for 0% CH3F. ED50, ED90, and ED95 are marked by red lines. (C) Box-plot summarizing accumulation of methane and acetate in control (n = 3) and in samples with CH3F≥0.75%, n = 6) after 14 days of anoxic incubation.
All fitted dose-response curves have ED90 (effective dose for 90% inhibition) concentrations of <0.75% CH3F. The dose-response curves for acetate and methane accumulation even showed ED99 concentrations of <1%. The higher ED99 for the isotopic signatures may be due to the rather gentle slope of the respective curves (Figure 2B).
If only aceticlastic methanogenesis was inhibited while acetogenesis proceeded, the sums of methane and acetate in control and fully inhibited samples (assumed at ≥0.75% CH3F) should be equal. Indeed, no significant difference was found (Figure 2 C; two sample t-test, p = 0.87). On basis of the results of the different dose-response curves we conclude additionally that above 0.75% CH3F virtually no acetate was consumed. Furthermore, our data does not indicate an effect on residual, hydrogenotrophic methanogenesis. In a previous experiment, hydrogenotrophic methanogenesis was found unaffected even at 4% CH3F [6: in a hypersaline microbial mat from Solar Lake, Sinai]. However, in two incubations at elevated CH3F concentrations (2.7 and 2.9%) not included in the dose-response fits, the amount of acetate produced was about 50% higher than the corresponding methane deficit. Methanogenesis and isotopic signatures, on the other hand, were not affected. Similar disproportionate acetate values have been reported before [44] and perhaps, these imbalances are caused by substrate heterogeneities, not by effects on methanogenesis.
Assuming that an initial CH3F concentration of 0.75% inhibited aceticlastic methanogenesis, hydrogenotrophic methanogenesis contributed 18.3% to total methane production. The inhibitory concentration is within the range usually applied to rice field [2], [4], [6], [17], [44]–[49] and other wetland soils [50]–[53]. A decade ago, CH3F was thought to be a specific inhibitor for methane oxidation in general [3] and has been applied to chamber experiments quantifying methane oxidation from the difference between methane fluxes with and without CH3F (Table 1). Considering an ED50 of <0.25% CH3F for aceticlastic methanogenesis, these experiments may likely have underestimated the amount of methane oxidized due to co-inhibition of aceticlastic methanogenesis.
Table 1. Experiments quantifying methane oxidation from the difference between methane fluxes measured with and without CH3F.
| Reference | Year | Ecosystem | Biome, Ecozone | CH3F concentration |
| [45] | 1995 | Wetland rice | Temperate | 1% |
| [58] | 1997 | Wetland | Temperate | 1.5% |
| [47] | 1996 | Wetland rice | Tropics | 1.5, 3% |
| [59] | 1993 | Wetland rice, weeds | Subtropics | 1.5, 3% |
| [6] | 1996 | Wetland rice | Mediterranean | 0.7, 1.7, 3% |
| [60] | 2001 | Weed (Myriophyllum) | Temperate | 84–140 µM |
| [48] | 2001 | Wetland rice | Subtropical | 3% |
| [61] | 1996 | Weed (Sparganium) | Boreal | 3–4% |
| [62] | 1998 | Tundra wetland | Subarctic | 1% |
| [63] | 2000 | Wetland | Boreal | 1.5–3% |
The methanogenic community
Community composition (DNA-based) and transcripts were analyzed by t-RFLP analysis as well as by cloning of the mcrA gene fragments and transcripts. Results of the t-RFLP analysis of the mcrA gene (Figure 3) indicated a high relative abundance of versatile Methanosarcinaceae (tRF 126, 133, 652, 683) and hydrogenotrophic Methanobacteriales (tRF 126, 663, 752). In addition, Methanocellales (tRF 133) were found in all incubations. Two tRFs could not be separated further: an in silico analysis of mcrA sequences from the clone library revealed that tRF 133 occurred in Methanocellales, the Fen cluster, and Methanosarcinaceae, while tRF 126 comprised both Methanobacteriales and Methanosarcinaceae. Despite this, t-RFLP patterns showed a distinct separation between total and active community in all analyses applied: CCA (Figure 3A) and MRT igure 3B) demonstrated consistently that a homogenous, active community was found across the whole CH3F gradient applied. Furthermore, virtually the same separation was found with non-metric multidimensional scaling (NMDS; stress = 0.02, r2 linear = 0.99; ordination not shown). As found recently for methanogens [21] and other microbial guilds [54], the active community consisted only of a subset of the total. Most remarkable was here the nearly complete absence of restriction fragments indicative for Methanobacteriales mcrA transcripts.
Figure 3. Multivariate analysis of relative abundances of terminal restriction fragments (tRF).
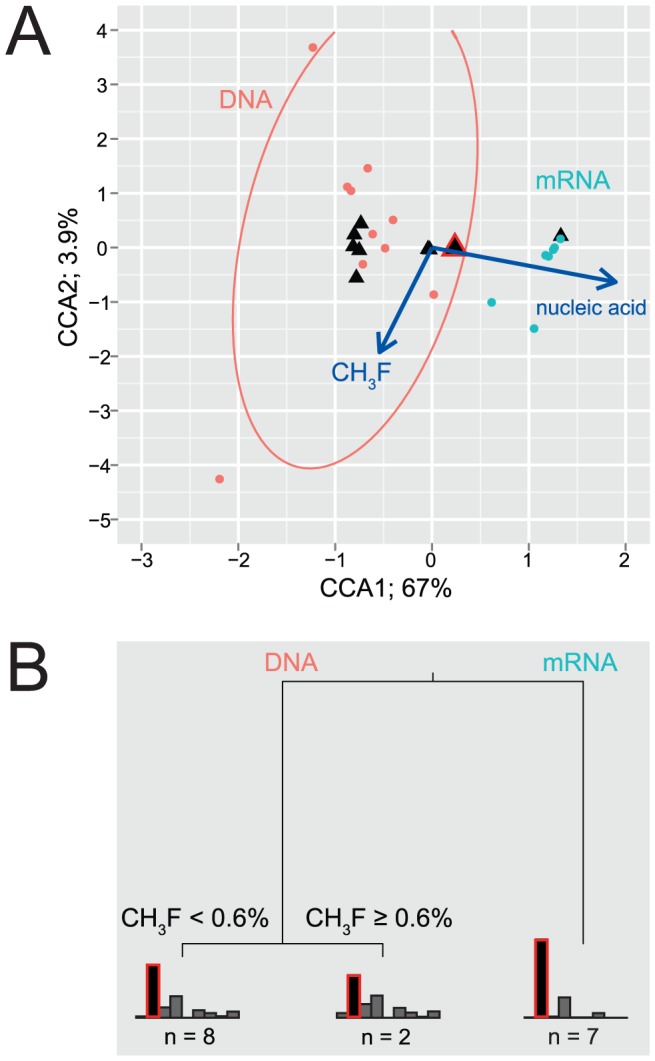
(A) Biplot of a constrained correspondence analysis (CCA). Two constraints were applied: CH3F concentration and the type of nucleic acid, i.e. DNA or mRNA. The CCA explains about 71% of overall variation, with CCA1 being the most important axis. The arrows indicate the direction in which constraints correlate with the ordination axes. Confidence ellipses (95%) surround the centers of DNA- and mRNA-derived communities, respectively. Closed circles represent the samples, and black triangles the different tRFs. The triangle surrounded by a red outline corresponds to tRF 133, the numerically dominant fragment. (B) Multivariate regression tree (MRT) based on squared Euclidean distances. The vertical spacing of the branches is proportional to the error in the fit; the first split reduces the error by 75%. The tree is pruned, i.e. the least important splits have been removed. Barplots at the leaves show the relative abundance of different tRFs; from left: 126, 133, 503, 648, 652, 663, 683, 743, and 752 bp. As in panel A, tRF 133 is marked by a red outline.
Cloning and sequencing allowed further differentiation. The DNA-based library constructed from soil sampled at the beginning of the experiment was dominated by sequences affiliated to Methanocellales, Methanosarcinaceae and Methanobacteriales, but also by a few members of the Fen cluster and a so far uncharacterized cluster (Table 2). The latter (OTU 12; Table 2) were found in clones retrieved under CH3F suggesting a hydrogenotrophic mode of life. In accordance with our t-RFLP findings, only a minor fraction of this diversity could be retrieved from mRNA resulting in highly significant differences between DNA- and mRNA-based clone libraries (Table 2). Considering mRNA derived sequences as a proxy for group-specific activity, Methanobacteriales appeared to not produce methane at all. Similarly, Methanocellales seemed to have been much less important for methanogenesis than expected from their high dominance in the DNA-based clone library. With and without repression of aceticlastic methanogenesis, Methanosarcinaceae were the most active methanogens suggesting that they used acetate when possible, but shifted to H2 + CO2, if acetate usage was inhibited. This is in accordance with a previous experiment on Methanosarcina barkeri strain MS that was inhibited by CH3F when supplied with acetate, but not if grown on H2 + CO2 [7]. Methanosarcinaceae sequences detected here were affiliated to the type strain of Methanosarcina mazei (Figure 4) being able to use both these substrates, too [55]. It is intriguing that under CH3F inhibition, no Methanocellales-related sequences could be retrieved anymore from mRNA, resulting in a small yet still significant difference between the respective libraries (Table 2). While we cannot rule out a direct effect, shifting Methanosarcinaceae towards a hydrogenotrophic mode of life might also have changed competition for H2 resulting in an indirect effect on Methanocellales.
Table 2. Abundances of the 22 operational taxonomic units (OUTs) with a maximum intra-group distance of 5% (AA) in the clone library.
| OTU | Affiliation | TRF | Start, DNA | Control, mRNA | CH3F 3.85%, mRNA |
| 1 | Msarc | 139 | 15 | 40 | 42 |
| 2 | Mcell | 139 | 6 | 0 | 0 |
| 3 | Mcell | 138 | 6 | 0 | 0 |
| 4 | Mcell | 139 | 1 | 0 | 0 |
| 5 | Mcell | 139 | 5 | 0 | 0 |
| 6 | Mbac | 760 | 10 | 0 | 0 |
| 7 | Mbac | 131 | 3 | 0 | 0 |
| 8 | Mbac | 760 | 3 | 0 | 0 |
| 9 | Mbac | 666 | 1 | 0 | 0 |
| 10 | Mbac | 666 | 1 | 0 | 0 |
| 11 | Mcell | 138 | 10 | 3 | 0 |
| 12 | NN | 139 | 1 | 0 | 2 |
| 13 | Msaeta | 131 | 2 | 0 | 0 |
| 14 | Fen | 139 | 1 | 0 | 0 |
| 15 | Msarc | 139 | 1 | 0 | 0 |
| 16 | Mbac | 733 | 1 | 0 | 0 |
| 17 | Mbac | 760 | 1 | 0 | 0 |
| 18 | Mcell | 138 | 1 | 0 | 0 |
| 19 | Mcell | 138 | 0 | 1 | 0 |
| 20 | Msarc-like | 139 | 0 | 2 | 0 |
| 21 | Mcell | 139 | 0 | 0 | 1 |
| χ2 test,simulated p-values | Control, mRNA | CH3F, mRNA | |||
| Start, DNA | 0.0001 | 0.0001 | |||
| Control, mRNA | 0.05 | ||||
Clones were derived from samples taken before (‘start’, based on DNA) and after (‘control’ and 3.85% CH3F, based on transcripts) anoxic incubation for 14 days. OTU number and affiliation to families are given as in Figure 4. Msarc: Methanosarcinaceae, Mcell: Methanocellales, Mbac: Methanobacteriales, Msaeta = Methanosaetaceae, Fen = Fen cluster, Msarc-like = uncertain affiliation, but nearest to Methanosarcinaceae; NN = unknown cluster. Simulated p-values are from a Monte-Carlo simulation with 9999 replicates.
Figure 4. Neighbor-joining tree based on 147 deduced amino acid positions from 949 mcrA sequences.
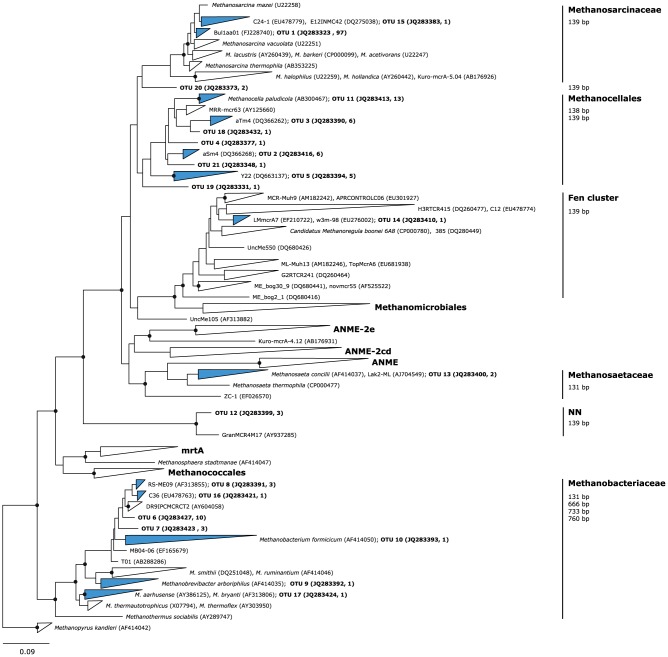
Phylogenetic nodes verified by a maximum likelihood tree are marked with closed circles. The outer branches of distinct clusters are collapsed, and those containing OTUs defined in this study are marked in blue. Only representative sequences for the OTUs have been incorporated into the tree and are depicted as ‘OTU name (accession number, number of sequences representing the OTU)’. Environmental clusters were labeled with two reference sequences showing maximum phylogenetic distance within the respective cluster, given as ‘name 1 (accession number 1), name 2 (accession number 2). The corresponding tRFs were calculated in silico using the TRiFLe package [64] and are given to the right. Scale bar: 0.09 changes per amino acid position. The outgroup is Methanopyrus kandleri.
Conclusion
While we found CH3F to act specifically on aceticlastic methanogenesis, the results obtained from the analysis of mcrA transcripts allow for relevant conclusions beyond this technical aspect. Community composition has often been regarded as a controlling factor for the flow of carbon and reductants through microbial communities. However, this experiment has shown how versatile Methanosarcinaceae are very well capable of delivering the same end-product under totally different conditions. This supports concepts developed to understand and predict the reaction of microbial communities to environmental changes [56], [57]. Furthermore, this experiment demonstrates how the sensible application of selective inhibitors can help detecting physiological traits of yet uncultivated microbes eventually supporting the design of cultivation strategies. Having found previously the same effect of CH3F on methanogenesis in a soil from an Italian rice field [6] more than 10,000 km apart from that in China let us trust that our findings are widely applicable.
Acknowledgments
The authors thank Zhi Hong Cao, Institute of Soil Science, CAS, Nanjing, (China), for providing the soil sample, and Peter Claus and Bellinda Schneider (MPI Marburg) for technical assistance. This is a contribution from the framework ‘Biogeochemistry of paddy soil evolution’. This publication is publication no. 5385 of the Netherlands Institute of Ecology.
Funding Statement
Work has been carried out at the Max Planck Institute for Terrestrial Microbiology, Marburg, Germany. AD's work on this project was funded by a grant of Deutsche Forschungsgemeinschaft to PF as part of the framework ‘Biogeochemistry of paddy soil evolution’, and further supported by Riks Laanbroek (NIOO-KNAW). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Conrad R, Frenzel P (2002) Flooded soils. In: Britton G, editor. New York: John Wiley & Sons. pp. 1316–1333.
- 2. Conrad R, Klose M (1999) How specific is the inhibition by methyl fluoride of acetoclastic methanogenesis in anoxic rice field soil? FEMS Microbiol Ecol 30: 47–56. [Google Scholar]
- 3. Oremland RS, Culbertson CW (1992) Evaluation of methyl fluoride and dimethyl ether as inhibitors for aerobic methane oxidation. Appl Environ Microbiol 58: 2983–2992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Bodelier PLE, Frenzel P (1999) Contribution of methanotrophic and nitrifying bacteria to CH4 and NH4 + oxidation in the rice rhizosphere using new methods for discrimination. Appl Environ Microbiol 65: 1826–1833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Oremland RS, Capone DG (1988) Use of “specific” inhibitors in biogeochemistry and microbial ecology. Adv Microb Ecol 10: 285–383. [Google Scholar]
- 6. Frenzel P, Bosse U (1996) Methyl fluoride, an inhibitor of methane oxidation and methane production. FEMS Microbiol Ecol 21: 25–36. [Google Scholar]
- 7. Janssen PH, Frenzel P (1997) Inhibition of methanogenesis by methyl fluoride: studies on pure and defined mixed cultures of anaerobic bacteria and archaea. Appl Environ Microbiol 63: 4552–4557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Conrad R (2005) Quantification of methanogenic pathways using stable carbon isotopic signatures: a review and a proposal. Org Geochem 36: 739–752. [Google Scholar]
- 9. Whiticar MJ, Faber E, Schoell M (1986) Biogenic methane formation in marine and freshwater environments: CO2 reduction vs. acetate fermentation - Isotope evidence. Geochim Cosmochim Acta 50: 693–709. [Google Scholar]
- 10. Krüger M, Eller G, Conrad R, Frenzel P (2002) Seasonal variation in pathways of CH4 production and in CH4 oxidation in rice fields determined by stable carbon isotopes and specific inhibitors. Global Change Biol 8: 265–280. [Google Scholar]
- 11. Conrad R, Chan OC, Claus P, Casper P (2007) Characterization of methanogenic Archaea and stable isotope fractionation during methane production in the profundal sediment of an oligotrophic lake (Lake Stechlin, Germany). Limnol Oceanogr 52: 1393–1406. [Google Scholar]
- 12. Penning H, Conrad R (2007) Quantification of carbon flow from stable isotope fractionation in rice field soils with different organic matter content. Org Geochem 38: 2058–2069. [Google Scholar]
- 13. Krüger M, Frenzel P, Kemnitz D, Conrad R (2005) Activity, structure and dynamics of the methanogenic archaeal community in a flooded Italian rice field. FEMS Microbiol Ecol 51: 323–331. [DOI] [PubMed] [Google Scholar]
- 14. Sakai S, Conrad R, Liesack W, Imachi H (2010) Methanocella arvoryzae sp. nov., a hydrogenotrophic methanogen isolated from rice field soil. IJSEM 60: 2918–2923. [DOI] [PubMed] [Google Scholar]
- 15. Chin KJ, Lueders T, Friedrich MW, Klose M, Conrad R (2004) Archaeal community structure and pathway of methane formation on rice roots. Microbiol Ecol 47: 59–67. [DOI] [PubMed] [Google Scholar]
- 16. Lu Y, Conrad R (2005) In situ stable isotope probing of methanogenic archaea in the rice rhizosphere. Science 309: 1088–1090. [DOI] [PubMed] [Google Scholar]
- 17. Penning H, Conrad R (2006) Effect of inhibition of acetoclastic methanogenesis on growth of archaeal populations in an anoxic model environment. Appl Environ Microbiol 72: 178–184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Conrad R, Klose M, Noll M, Kemnitz D, Bodelier PLE (2008) Soil type links microbial colonization of rice roots to methane emission. Global Change Biol 14: 657–669. [Google Scholar]
- 19. Großkopf R, Janssen PH, Liesack W (1998) Diversity and structure of the methanogenic community in anoxic rice paddy soil microcosms as examined by cultivation and direct 16S rRNA gene sequence retrieval. Appl Environ Microbiol 64: 960–969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wu L, Ma K, Li Q, Ke X, Lu Y (2009) Composition of archaeal community in a paddy field as affected by rice cultivar and N fertilizer. Microbiol Ecol 58: 819–826. [DOI] [PubMed] [Google Scholar]
- 21. Ma K, Conrad R, Lu Y (2012) Responses of methanogen mcrA genes and their transcripts to alternate dry wet cycle of paddy field soil. Appl Environ Microbiol 78: 445–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ramakrishnan B, Lueders T, Dunfield PF, Conrad R, Friedrich MW (2001) Archaeal community structures in rice soils from different geographical regions before and after initiation of methane production. FEMS Microbiol Ecol 37: 175–186. [Google Scholar]
- 23. Thauer RK (1998) Biochemistry of methanogenesis - a tribute to Marjory Stephenson. Microbiology 144: 2377–2406. [DOI] [PubMed] [Google Scholar]
- 24. Hennigan AN, Reeve JN (1994) mRNAs in the methanogenic archaeon Methanococcus vannielii: numbers, half-lives and processing. Mol Microbiol 11: 655–670. [DOI] [PubMed] [Google Scholar]
- 25. Kotiaho M, Fritze H, Merilä P, Juottonen H, Leppälä M, et al. (2010) Methanogen activity in relation to water table level in two boreal fens. Biol Fert Soils 46: 567–575. [Google Scholar]
- 26. Watanabe T, Kimura M, Asakawa S (2009) Distinct members of a stable methanogenic archaeal community transcribe mcrA genes under flooded and drained conditions in Japanese paddy field soil. Soil Biol Biochem 41: 276–285. [Google Scholar]
- 27. Wissing L, Kölbl A, Vogelsang V, Fu JR, Cao ZH, et al. (2011) Organic carbon accumulation in a 2000-year chronosequence of paddy soil evolution. Catena 87: 376–385. [Google Scholar]
- 28. Ho A, Lüke C, Frenzel P (2011) Ageing well: methane oxidation and methane oxidising bacteria along a chronosequence of 2000 years. Environ Microbiol Rep 3: 738–743. [DOI] [PubMed] [Google Scholar]
- 29. Cheng YQ, Yang LZ, Cao ZH, Ci E, Yin S (2009) Chronosequential changes of selected pedogenic properties in paddy soils as compared with non-paddy soils. Geoderma 151: 31–41. [Google Scholar]
- 30. Conrad R, Goevert D (2009) Effect of substrate concentration on carbon isotope fractionation during acetoclastic methanogenesis by Methanosarcina barkeri and M. acetivorans and in rice field soil. Appl Environ Microbiol 75: 2605–2612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Lueders T, Manefield M, Friedrich MW (2004) Enhanced sensitivity of DNA- and rRNA-based stable isotope probing by fractionation and quantitative analysis of isopycnic centrifugation gradients. Environ Microbiol 6: 73–78. [DOI] [PubMed] [Google Scholar]
- 32. Hales BA, Edwards C, Ritchie DA, Hall G, Pickup RW, et al. (1996) Isolation and identification of methanogen-specific DNA from blanket bog peat by PCR amplification and sequence analysis. Appl Environ Microbiol 62: 668–675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Bukovská P, Jelínková M, Hršelová H, Sýkorová Z, Gryndler M (2010) Terminal restriction fragment length measurement errors are affected mainly by fragment length, G-C nucleotide content and secondary structure melting point. J Microbiol Meth 82: 223–228. [DOI] [PubMed] [Google Scholar]
- 34. Kaplan CW, Kitts CL (2003) Variation between observed and true Terminal Restriction Fragment length is dependent on true TRF length and purine content. J Microbiol Meth 54: 121–125. [DOI] [PubMed] [Google Scholar]
- 35. Ludwig W, Strunk O, Westram R, Richter L, Meier H, et al. (2004) ARB: a software environment for sequence data. Nucleic Acids Res 32: 1363–1371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, et al. (2009) Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl Environ Microbiol 75: 7537–7541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.R Development Core Team (2011) R: A language and environment for statistical computing. 2.13.1 ed.
- 38. Ritz C, Streibig JC (2005) Bioassay analysis using R. J Stat Software 12: 1–22. [Google Scholar]
- 39.Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, et al.. (2011) vegan: Community Ecology Package. 2.2-0 ed.
- 40. De'Ath G (2002) Multivariate regression trees: a new technique for modeling species-environment relationships. Ecology 83: 1105–1117. [Google Scholar]
- 41.Wickham H (2009) ggplot2. Elegant graphics for data analysis; Gentleman R, Hornik K, Parmigiani G, editors. Dordrecht: Springer. 1–212 p.
- 42. Whiticar MJ (1999) Carbon and hydrogen isotope systematics of bacterial formation and oxidation of methane. Chem Geol 161: 291–314. [Google Scholar]
- 43. Gelwicks JT, Risatti JB, Hayes JM (1989) Carbon isotope effects associated with autotrophic acetogenesis. Org Geochem 14: 441–446. [DOI] [PubMed] [Google Scholar]
- 44. Conrad R, Klose M (2000) Selective inhibition of reactions involved in methanogenesis and fatty acid production on rice roots. FEMS Microbiol Ecol 34: 27–34. [DOI] [PubMed] [Google Scholar]
- 45. Banker BC, Kludze HK, Alford DP, DeLaune RD, Lindau CW (1995) Methane sources and sinks in paddy rice soils: relationship to emissions. Agricult Ecosys Environ 53: 243–251. [Google Scholar]
- 46. Bodelier PLE, Hahn AP, Arth I, Frenzel P (2000) Effects of ammonium-based fertilisation on microbial processes involved in methane emission from soils planted with rice. Biogeochemistry 51: 225–257. [Google Scholar]
- 47. Denier van der Gon HAC, Neue HU (1996) Oxidation of methane in the rhizosphere of rice plants. Biol Fert Soils 22: 359–366. [Google Scholar]
- 48. Jia ZJ, Cai ZC, Xu H, Li XP (2001) Effect of rice plants on CH4 production, transport, oxidation and emission in rice paddy soil. Plant Soil 230: 211–221. [Google Scholar]
- 49. Roy R, Klüber HD, Conrad R (1997) Early initiation of methane production in anoxic rice soil despite the presence of oxidants. FEMS Microbiol Ecol 24: 311–320. [Google Scholar]
- 50. Reiche M, Torburg G, Küsel K (2008) Competition of Fe(III) reduction and methanogenesis in an acidic fen. FEMS Microbiol Ecol 65: 88–101. [DOI] [PubMed] [Google Scholar]
- 51. Jerman V, Metje M, Mandic-Mulec I, Frenzel P (2009) Wetland restoration and methanogenesis: the activity of microbial populations and competition for substrates at different temperatures. Biogeosciences 6: 1127–1138. [Google Scholar]
- 52. Metje M, Frenzel P (2007) Methanogenesis and methanogenic pathways in a peat from subarctic permafrost. Environ Microbiol 9: 954–964. [DOI] [PubMed] [Google Scholar]
- 53. Metje M, Frenzel P (2005) Effect of temperature on anaerobic ethanol oxidation and methanogenesis in an acidic peat from a northern wetland. Appl Environ Microbiol 71: 8191–8200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Krause S, Lüke C, Frenzel P (2010) Succession of methanotrophs in oxygen-methane counter-gradients of flooded rice paddies. ISME J 4: 1603–1607. [DOI] [PubMed] [Google Scholar]
- 55.Liu Y (2010) Methanosarcinales. In: Timmis KN, editor. Handbook of Hydrocarbon and Lipid Microbiology. Berlin Heidelberg: Springer. pp. 595–604.
- 56. Comte J, Del Giorgio PA (2011) Composition influences the pathway but not the outcome of the metabolic response of bacterioplankton to resource shifts. PLoS One 6: e25266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Allison SD, Martiny JBH (2008) Resistance, resilience, and redundancy in microbial communities. Proc Natl Acad Sci U S A 105: 11512–11519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Boeckx P, van Cleemput O (1997) Methane emission from a freshwater wetland in Belgium. Soil Sci Soc America J 61: 1250–1256. [Google Scholar]
- 59. Epp MA, Chanton JP (1993) Rhizospheric methane oxidation determined via the methyl fluoride inhibition technique. J Geophys Res 98: 18413–18418. [Google Scholar]
- 60. Heilman MA, Carlton RG (2001) Methane oxidation associated with submersed vascular macrophytes and its impact on plant diffusive methane flux. Biogeochemistry 52: 207–224. [Google Scholar]
- 61. King GM (1996) In situ analyses of methane oxidation associated with the roots and rhizomes of a bur reed, Sparganium eurycarpum, in a Maine wetland. Appl Environ Microbiol 62: 4548–4555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Moosavi SC, Crill PM (1998) CH4 oxidation by tundra wetlands as measured by a selective inhibitor technique. J Geophys Res-Atmos 103: 29093–29106. [Google Scholar]
- 63. Popp TJ, Chanton JP, Whiting GJ, Grant N (2000) Evaluation of methane oxidation in the rhizosphere of a Carex dominated fen in north central Alberta, Canada. Biogeochemistry 51: 259–281. [Google Scholar]
- 64. Junier P, Junier T, Witzel KP (2008) TRiFLe, a program for in silico terminal restriction fragment length polymorphism analysis with user-defined sequence sets. Appl Environ Microbiol 74: 6452–6456. [DOI] [PMC free article] [PubMed] [Google Scholar]