

Abstract
It remains difficult to obtain high-resolution atomic force microscopy images of HIV-1 integrase bound to DNA in a dimeric or tetrameric fashion. We therefore constructed specific target DNAs to assess HIV-1 integrase binding and purified the complex by dialysis prior to analysis. Our resulting atomic force microscopy analyses indicated precise size of binding human immunodeficiency virus type 1 (HIV-1) recombinant integrase in a tetrameric manner, inducing formation of a loop-like or figure-eight-like secondary structure in the target DNA. Our findings regarding the target DNA secondary structure provide new insights into the intermediate states of retroviral integration.
Introduction
Retroviruses integrate their genome into the genome of host cells in a process catalyzed by the enzyme integrase, which binds the 5′- and 3′- termini of retroviral long terminal repeats (LTRs) and integrates them into the host cell's DNA. Elucidating the underlying mechanism of integration will necessitate that we first obtain a more complete understanding of the transitional states involved in the binding of HIV-1 integrase to target DNA. Hare et al. recently reported a crystal structure of prototype foamy virus integrase complexed with its cognate DNA [1] that is suggestive of a tetrameric structure. HIV-1 integrase forms stable tetramers and associates with the transcriptional coactivator LEDGF/p75, which is an essential cofactor for HIV integration [2], [3]. Inhibitors have been developed to target tetrameric integrases [4], [5]. To date, no convincing visualizations of integrase tetramerization have been reported, perhaps due to difficulties in generating a tetramer stable enough to visualize.
In an effort to visualize integration intermediate states, we previously prepared target DNAs rich in 5′-CA and 5′-TG dinucleotides recognized by HIV-1 integrase [14] and successfully detected the formation of integrase oligomers using atomic force microscopy (AFM). This and other studies provided data that greatly advanced our understanding of the intermediate states of retroviral integration [6], [7], [8]. The results of recent studies of the binding of nucleoproteins to retroviral cDNA ends suggest that integrase forms oligomers at the ends of the retroviral genome [9], [10].
In the present study, we used a purified recombinant integrase produced in Sf9 insect cells for formation of the integrase-DNA complex. Kotova et al. estimated that the number of integrase molecules that bind to individual strands varies significantly [9], suggesting that evaluations based on the number of molecules binding to target DNA are more error prone than evaluations based upon measurement of the volume of DNA-integrase complexes. In studies involving AFM observations, background noise often impedes accurately measuring the intensity of the objective peaks. In the present study we therefore employed dialysis to reduce the background noise sufficiently to resolve the oligomeric structure of DNA-integrase complexes using AFM. Our study also revealed that oligomeric integrase digests the target DNA at the oligomeric integrase-binding site with secondary structure formation. Our AFM data indicated that recombinant HIV-1 integrase forms a tetrameric structure in complex with target DNA. These results will enhance current understanding of the molecular mechanism of retroviral integration.
Materials and Methods
Incubation of integrase with target DNA
Recombinant HIV-1 integrase was kindly provided by Dr. Tomokazu Yoshinaga, Shionogi Institute for Medical Science, Japan, who previously described the HIV-1 integration signal sequence [11]. This integrase was prepared free of nucleases. The target DNA consisted of approximately 4.0 kb of xkyk : (5′-GTGGAGGGCAGT-3′) k(5′-ACTGGGAAGGGAC-3′)k or x′ky′k : (5′-GAGGAGGGTAGC -3′)k(5′-GCTACCCTCCTC-3′)k, where 160<k<170 and the italicized portions of the sequence (CAGT and ACTG) represent sequence that originated from the LTR end of the HIV-1 provirus. In x′ky′k, CAGT and ACTG have been replaced. A 10-µL volume of solution containing 10 ng of target DNA was combined with 50 ng of recombinant integrase in 10 µL of binding buffer and incubated for 0 to 20 min at 30°C to prevent excess digestion by endonuclease activity that would occur during incubation at 37°C. The binding buffer was comprised of 25 mM MnCl2, 80 mM potassium glutamate, 10 mM mercaptoethanol, 10% DMSO, and 35 mM MOPS (pH 7.2).
Dialysis of the integrase-DNA complex
The integrase-DNA complex was dialyzed in order to reduce the background noise and eliminate artificial aggregation to obtain convincing images. Following incubation, 1.0 mL of PBS was added to the integrase-target DNA solution and the mixture was then gently injected into a dialysis bag (Slide-A-Lyzer Dialysis Cassettes, 20K MWCO, Pierce Biotechnology, Rockford, IL) and dialyzed against PBS for 48 h at 4°C in order to remove the MOPS and Mn2+. The PBS was exchanged six times every 2 hour. Finally, the dialysis product was gently withdrawn from the bag and analyzed using AFM or digested with ScaI (New England Biolabs, Ipswich, MA). This digestion was performed in order to prevent the formation of complicated DNA structures resulting from intersecting of the target DNA or supercoiled structure. We frequently observed such structures, which made it difficult to distinguish the monomeric integrase or artifacts in AFM observations due to heterogeneous fixation on the background mica.
Atomic force microscopy
A droplet of a solution containing target DNA and 10 mM spermidine was then placed on a freshly cleaved piece of mica (30–50 mm). Spermidine was used to aid the adsorption of DNA molecules onto the mica surface. After 5 min, the adsorbed DNA was washed with water and dried with N2 gas. The DNA molecules were analyzed in air at room temperature using an NVB100 atomic force microscope (Olympus Optical Co., Ltd., Tokyo, Japan; AFM controller and software: Nanoscope IIIa, Digital Instruments, Veeco, Camerillo, CA) operated in the tapping mode [12]. The integrase volume was calculated by
where Δx, Δy, and Δz denote the sizes of the three-dimensional axis. Details are described in the Results section below. These molecular size values were determined using Jmol, an open-source Java viewer for chemical structures in 3D (http://www.jmol.org/) on the basis of 3D X-ray structure analysis.
Electropherogram
Electrophoresis was performed following incubation of 10 ng of target DNA with 50 ng of integrase for various periods ranging from 0 to 20 min. The resulting signal intensities were transformed using Multigauge software (Fujifilm, Tokyo, Japan).
Statistical analysis
Statistical significance was assessed by unpaired t-tests using SPSS software (SPSS, Chicago, IL). P-values <0.01 were considered statistically significant.
Results
Binding of integrase to an artificial repeat sequence
xk yk or x′k y′k was incubated with recombinant HIV-1 integrase in a reaction buffer containing 40 mM MnCl2. Samples containing integrase and target DNA were dialyzed prior to analysis in order to obtain high-resolution images. Without this purification, high background was observed. After dialysis, AFM analysis indicated that HIV-1 recombinant integrase bound to the target DNA xkyk. The target DNA formed a twin-like globular structure. Well-defined loop-like (Figures 1A, 1B) or figure-eight-like structures (Figure 1C) were observed around the oligomeric integrase. The presence of free monomeric and oligomeric integrase indicated that integrase does not bind to target DNA incubated prior to dialysis in the absence of MnCl2 (Figure 1D). Binding of oligomeric integrase to x′ky′k was less observed in the presence of MnCl2 than xkyk (Figure 1E, See figure 2).
Figure 1. AFM observation of integrase-target DNA complexes.
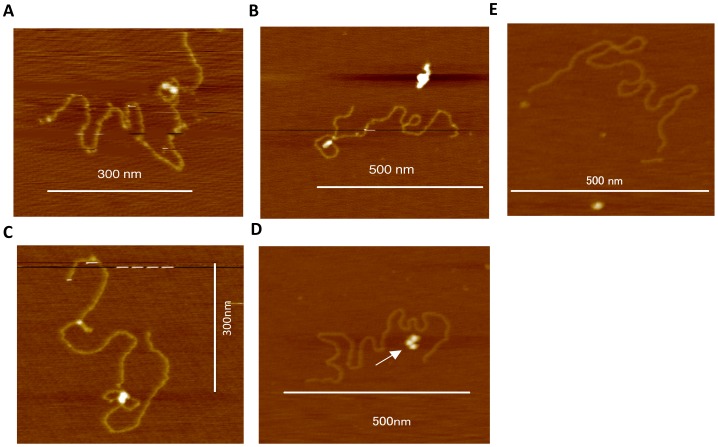
(A, B) Target DNA xkyk bound to HIV-1 integrase forming a loop-like structure or (C) a figure-eight-like structure. (D) DNA showing no HIV-1 integrase binding, where the arrow indicates unbound integrase. (E) Control experiment using x′ky′k including a ScaI digestion site AGTGACT. Bars indicate scale.
Figure 2. Number of integrase molecules bound in target DNA and modified DNA complexes.
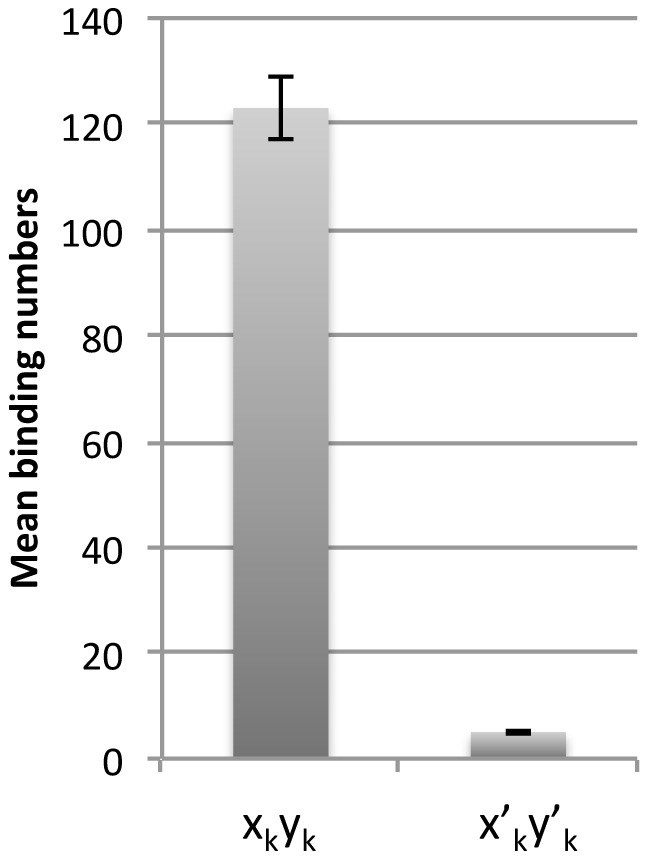
Mean number of oligomeric integrases binding to the target DNA or modified target DNA strands (per one hundred strands) (mean ± s.d.). Results were determined from 5 independent experiments.
Modification of target DNA significantly represses integrase binding
The target DNA x′ky′k was also incubated with recombinant HIV-1 integrase in a reaction buffer containing 40 mM MnCl2. Subsequent AFM analysis indicated that significantly fewer HIV-1 recombinant integrase molecules bound to the x′ky′k target DNA compared to the xkyk target DNA (5 molecules bound to x′ky′k strands vs. 123 molecules bound to xkyk strands, P<0.01) (Figure 2). These data indicate that the focal nucleotide sequence has a significant influence on integrase binding.
Integrase binds to target DNA in a tetrameric fashion
The objective molecular size was evaluated based on peak intensity using an AFM controller and software. Representative trace data are shown in Figure 3A [15], [16]. According to the protein database (PDB), the size of the x-, y-, and z-axes of the objective twin-like structure should be about Δx≈5, Δy≈5, and Δz≈5 nm. The diameter corresponds to the long axis of the monomeric integrase structure obtained from X-ray data [15], [16]. Our data revealed mean sizes of 5.8±0.3, 6.1±0.1, and 3.2±0.2 nm for Δx, Δy, and Δz, respectively (Figure 3A, B). Taking into account that fixation of the protein on the mica surface reduced the thickness to 60% of its actual value, the thickness (Δz) of the oligomeric integrase applied to the surface of mica corresponds to 3.2 nm/0.6 = 5.3 nm, implying that one integrase molecule was piled atop another [17], [18], [19]. The present Δx and Δy data indicate that the majority of the complexes contained dimeric integrase. The mean volume of the oligomeric integrase was 112±12 nm3 assuming an elliptical structure (twin-like structures; n = 123 DNA strands; Figure 3C). The volume distribution corresponds to a molecular mass of between 62 and 65 kDa [9]. Because the mass of monomeric integrase is 32 kDa, our data indicate that the majority of the complexes contained dimeric integrase. Therefore, the twin-like structure definitely binds to the target DNA in quadruplets.
Figure 3. Distribution of the size of integrase-target DNA complexes.
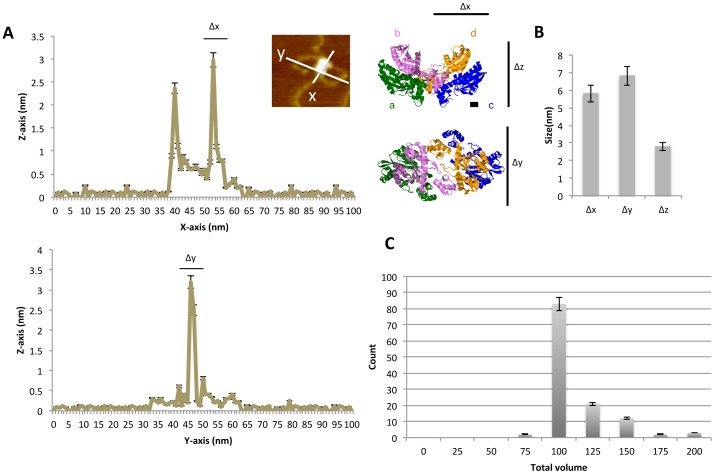
(A) Line graphs showing representative measurement of the diameter across a twin-like structure of integrase bound to the DNA strands along the x- and y-axes. The upper and lower graphs show the longer and shorter diameter of the twin-globule structure individually. The photo in the inset is identical to the photo shown in Figure 1C. To the right are 3D crystal structures of integrase obtained from X-ray data (PDB 1K6Y) [15]. Four integrase molecules associate in a tetrameric fashion in the DNA binding form. Monomers are represented by (a) green, (b) purple, (c) blue, and (d) orange. The diameter of the integrase dimer along the y-axis is approximately 5 nm, measured by the distance between two amino acids (E69 of c and I191 of d), represented in red. The thickness along the z-axis is approximately 3 nm, measured by the distance between Q62 of c and C65 of d, represented in red. These values were determined using Jmol. The scale bars are equivalent to approximately 1 nm. (B) Size of tetrameric integrase along the x-, y-, and z-axes (n = 123). (C) Histogram showing the distribution of the estimated volume (nm3) of the twin-like integrase structures bound to target DNA strands (n = 123).
Integrase binding generates target DNA secondary structures
To evaluate the effect of integrase binding on the target DNA structure, electrophoretic analysis was performed directly without digestion of the circular xkyk or x′ky′k DNA following the integrase binding reaction. In addition to the target DNA (i) and presumably supercoiled signal (iii), an upper signal (ii) and an evident mobility fluctuation of (iii) were observed (Figure 4A). This analysis showed that the relative areas associated with electrophoretic signals (ii) and (iii) were significantly higher in xkyk relative to x′ky′k (Figures 4B–D, *P<0.01), confirming that integrase binding to xkyk induces fluctuation in the DNA structure.
Figure 4. Electrophoretic analysis of circular DNA bound to integrase.
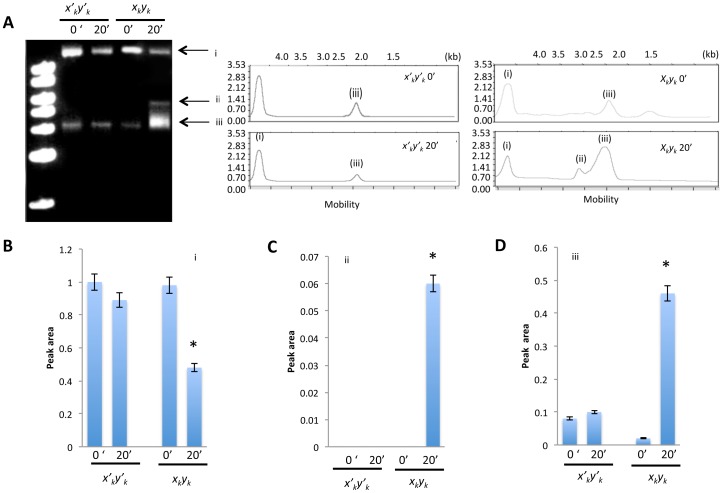
(A) Electrophoretic analysis of control and target DNA following incubation with integrase. Graphs show electropherograms of signals corresponding to bands (i)–(iii). Peak area of individual signals (B) (i), (C)(ii), and (D)(iii) was determined using Multigauge software. Control lane depicts the electrophoresis of the random-sequence control DNA.
Discussion
The results of the present study, which utilized target sequences originating from repeats of the HIV-1 cDNA ends, provide clear evidence that recombinant integrase forms a tetrameric complex on target DNA. A tetramer model of the HIV-1 integrase-DNA complex was previously constructed in an effort to predict the integrase residues that interact with the LTR termini [3], [13]. A repeat unit of target DNA was identical to the DNA for use in an in vitro integration assay in our previous study [14]. The target DNA was expected to maintain the integrase binding during the dialysis process. Sequence motif CAGT in the target DNA was observed in the end of HIV-1 ling terminal repeat, which was essential to HIV-1 integration [14], [20]. Indeed, this CAGT is a part of integration signal sequence that was reported by Yoshinaga et al [11]. In addition to this DNA sequence feature, the target DNA yielded a fluctuating secondary structure in the reaction buffer as shown in Figure 4, and this fluctuation contributes to the integration efficiency, probably because of the tension created by the secondary structure.
The present data clearly indicates that the twin-like structure binds to the target DNA in quadruplets, i.e., in a tetrameric fashion, which is consistent with the results of previous crosslinking experiments [9]. Our data also indicate that integrase induces a significant bend or fold in the target DNA, thereby producing a characteristic secondary structure. When the DNA was maintained in a circular structure the pattern of integrase-DNA binding was more complicated and it was frequently difficult to observe the binding complex. Therefore, we digested the complex in order to produce a more distinct secondary structure that would be easier to observe. By releasing the topological tension associated with the circular structure of the target DNA, we were able to reliably measure the volume of oligomeric integrase and thereby determine that the enzyme forms a tetrameric complex with the target. We found that dialyzing the integrase-DNA solution significantly reduced the background noise resulting from aggregation induced by MnCl2, thereby improving the resolution of subsequent images. This method enabled us to obtain high-resolution images of the structure of the integrase-DNA complex from which we were able to determine that dimeric integrase binds tightly to the target DNA in a symmetric fashion, producing a stable tetrameric configuration. Although our study is not applicable to in vivo integration studies, the data do provide new insights into how HIV-1 integrase interacts with host cell target DNA.
Acknowledgments
We are grateful to Masakazu Hatanaka and Tomokazu Yoshinaga for their helpful advice and insightful comments regarding the manuscript.
Funding Statement
This work was supported by a Grant-in-Aid for Cancer Research from the Ministry of Education, Culture, Science and Technology, Japan, and by a grant from the Ministry of Health, Welfare, and Labor, Japan. The funding agencies had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hare S, Gupta SS, Valkov E, Engelman A, Cherepanov P (2010) Retroviral intasome assembly and inhibition of DNA strand transfer. Nature 464: 232–236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Liano M, Saenz DT, Meehan A, Wongthida P, Peretz M, et al. (2006) An essential role for LEDGF/p75 in HIV integration. Science 314: 461–464. [DOI] [PubMed] [Google Scholar]
- 3. Cherepanov P, Maertens G, Proost P, Devreese B, Van Beeumen J, et al. (2003) HIV-1 integrase forms stable tetramers and associates with LEDGF/p75 protein in human cells. J Biol Chem 278: 372–381. [DOI] [PubMed] [Google Scholar]
- 4. Bar-Magen T, Sloan RD, Donahue DA, Kuhl BD, Zabeida A, et al. (2010) Identification of Novel Mutations Responsible for Resistance to MK-2048, a Second-Generation HIV-1 Integrase inhibitor. J Virol 84: 9210–9216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Christ F, Voet A, Marchand A, Nicolet S, Desimmie BA, et al. (2010) Rational design of small-molecule inhibitors of the LEDGF/p75-integrase interaction and HIV replication. Nat Chem Biol 6: 442–448. [DOI] [PubMed] [Google Scholar]
- 6. Li M, Mizuuchi M, Burke TR Jr, Craigie R (2006) Retroviral DNA integration: Reaction pathway and critical intermediates. EMBO J 25: 1295–1304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Deprez E, Tauc P, Leh H, Mouscadet JF, Auclair C, et al. (2001) DNA binding induces dissociation of the multimeric form of HIV-1 integrase: a time-resolved fluorescence anisotropy study. Proc Natl Acad Sci U S A 98: 10090–10095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Craigie R (2002) Retroviral DNA integration. In Mobile DNA II, Craig NL, Craigie R, Gellert M, Lambowitz AM (eds) pp 613–630. Washington, DC: ASM Press
- 9. Kotova S, Li M, Dimitriadis EK, Craigie R (2010) Nucleoprotein intermediates in HIV-1 DNA integration visualized by atomic force microscopy. J Mol Biol 399: 491–500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Ren G, Gao K, Bushman FD, Yeager M (2007) Single-particle image reconstruction of a tetramer of HIV integrase bound to DNA. J Mol Biol 366: 286–294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Yoshinaga T, Fujiwara T (1995) Different roles of bases within the integration signal sequence of human immunodeficiency virus type 1 in vitro. J Virol 69: 3233–3236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Ueda M, Yoshikawa K (1996) Phase transition and phase segregation in a single double-stranded DNA molecule. Physical Review Letters 77: 2133–2136. [DOI] [PubMed] [Google Scholar]
- 13. Dolan J, Chen A, Weber IT, Harrison RW, Leis J (2009) Defining the DNA substrate binding sites on HIV-1 integrase. J Mol Biol 385: 568–579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Tsuruyama T, Nakai T, Hiratsuka T, Jin G, Nakamura T, et al. (2010) In vitro HIV-1 selective integration into the target sequence and decoy-effect of the modified sequence. PLoS One 5: e13841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Maertens GN, Hare S, Cherepanov P (2010) The mechanism of retroviral integration from X-ray structures of its key intermediates. Nature 468: 326–329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Wang JY, Ling H, Yang W, Craigie R (2001) Structure of a two-domain fragment of HIV-1 integrase: implications for domain organization in the intact protein. Embo Journal 20: 7333–7343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Kuznetsov VY, Ivanov YD, Bykov VA, Saunin SA, Fedorov IA, et al. (2002) Atomic force microscopy detection of molecular complexes in multiprotein P450cam containing monooxygenase system. Proteomics 2: 1699–1705. [DOI] [PubMed] [Google Scholar]
- 18. Ivanov YD, Frantsuzov PA, Zollner A, Medvedeva NV, Archakov AI, et al. (2011) Atomic force microscopy study of protein-protein interactions in the cytochrome cyp11a1 (p450scc)-containing steroid hydroxylase system. Nanoscale Research Letters 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Ohno H, Kobayashi T, Kabata R, Endo K, Iwasa T, et al. (2011) Synthetic RNA-protein complex shaped like an equilateral triangle. Nature Nanotechnology 6: 115–119. [DOI] [PubMed] [Google Scholar]
- 20.Tsuruyama T (2012) Intech publication. In: Deniz Ekinci, editor. HIV-1 selectively integrates into host DNA in vitro. Biochemistry. pp 306–320.