

Abstract
The human gut microbiota is increasingly recognized as a player in colorectal cancer (CRC). While particular imbalances in the gut microbiota have been linked to colorectal adenomas and cancer, no specific bacterium has been identified as a risk factor. Recent studies have reported a high abundance of Fusobacterium in CRC subjects compared to normal subjects, but this observation has not been reported for adenomas, CRC precursors. We assessed the abundance of Fusobacterium species in the normal rectal mucosa of subjects with (n = 48) and without adenomas (n = 67). We also confirmed previous reports on Fusobacterium and CRC in 10 CRC tumor tissues and 9 matching normal tissues by pyrosequencing. We extracted DNA from rectal mucosal biopsies and measured bacterial levels by quantitative PCR of the 16S ribosomal RNA gene. Local cytokine gene expression was also determined in mucosal biopsies from adenoma cases and controls by quantitative PCR. The mean log abundance of Fusobacterium or cytokine gene expression between cases and controls was compared by t-test. Logistic regression was used to compare tertiles of Fusobacterium abundance. Adenoma subjects had a significantly higher abundance of Fusobacterium species compared to controls (p = 0.01). Compared to the lowest tertile, subjects with high abundance of Fusobacterium were significantly more likely to have adenomas (OR 3.66, 95% CI 1.37–9.74, p-trend 0.005). Cases but not controls had a significant positive correlation between local cytokine gene expression and Fusobacterium abundance. Among cases, the correlation for local TNF-α and Fusobacterium was r = 0.33, p = 0.06 while it was 0.44, p = 0.01 for Fusobacterium and IL-10. These results support a link between the abundance of Fusobacterium in colonic mucosa and adenomas and suggest a possible role for mucosal inflammation in this process.
Introduction
The human intestinal microbiota inhabits a complex and diverse environment populated by hundreds of different bacterial species. The number of bacterial cells in the gut exceeds all other eukaryotic cells in the human body by a factor of 10 [1], [2]. These bacteria are regulated in the gut by the mucosal immune system, which is made up of a complex network of functions and immune responses aimed at maintaining a cooperative system between the intestinal microbiota and the host [1]. In a healthy gut these bacteria maintain homeostasis with the host. However, when an imbalance, or bacterial dysbiosis, occurs in the gut, the host may experience inflammation and a loss of barrier function [3], [4]. Bacterial dysbioses have been linked to several diseases including ulcerative colitis, Crohn’s disease [5]–[7] and colorectal cancer (CRC) [8], [9]. Current research is focused on identifying key players in this imbalance as well as their specific contribution to colorectal carcinogenesis.
No single bacterial species has been identified as a risk factor for CRC, but recent studies report an increase in the abundance of Fusobacterium in human colorectal tumors compared to controls [8], [10], [11]. These studies suggest that Fusobacterium may be associated with the later stages of CRC, but it is unknown if they play a role in the early stages of colorectal carcinogenesis. While the causes of colorectal cancer are not fully known, it is becoming increasingly clear that the gut microbiota provide an important contribution [12].
We evaluated whether Fusobacterium nucleatum in normal rectal mucosal biopsies was associated with colorectal adenomas. We also examined a potential association between local inflammation and abundance of Fusobacterium in adenoma cases and non-adenoma controls. We found that Fusobacterium was more abundant in adenoma cases than controls. We observed significant positive correlation between IL-10 and TNF-α gene expression and abundance of Fusobacterium species in cases. Validation experiments were performed using CRC tissue and matching normal tissue to confirm previous reports of an association between CRC and Fusobacterium.
Results
Studies in Normal Rectal Mucosa of Adenoma and Non-adenoma Subjects
Fusobacterium abundance is higher in adenoma cases compared to controls
We evaluated Fusobacterium species in normal mucosal biopsies from 115 subjects, 48 cases and 67 controls by qPCR. Subject characteristics are shown in Table 1. All subjects were similar in age with cases having a mean age of 56.38±0.92, and controls 55.90±0.88 years. There were no significant differences between adenoma cases and non-adenoma controls for several risk factors evaluated including alcohol intake, caloric intake, waist-hip ratio, body mass index and total fat intake. Abundance of Fusobacterium species was significantly higher in adenoma cases compared to controls (mean log copy number and standard error, cases, 8.44±0.38; controls 7.40±0.22 p = 0.01) (Fig. 1). Compared to those with low abundance of Fusobacterium, those with high abundance of Fusobacterium were more likely to be adenoma cases (p-trend = 0.005) (Table 2). We also assessed the correlation between Fusobacterium abundance and the frequency and size (small, medium, large) of adenomas among cases. There was no significant correlation between Fusobacterium species and adenoma size (Table S1) or number of adenomas (r = −0.08, p = 0.57).
Table 1. Characteristics of Study Participants.
| Characteristic | Case (n = 48) | Control (n = 67) | P-value |
| Age (years, mean, se) | 56.38±0.92 | 55.90±0.88 | 0.71 |
| Waist-Hip ratio(mean, se) | 0.94±0.01 | 0.91±0.01 | 0.14 |
| Body Mass Index(kg/m2, mean, se) | 27.40±0.61 | 27.04±0.66 | 0.70 |
| Alcohol Intake(g/day, mean, se) | 12.65±1.94 | 21.17±8.88 | 0.41 |
| Calories(kcal/day, mean, se) | 2108.70±114.78 | 2140.38±144.0 | 0.87 |
| Total Fat intake(g/day, mean, se) | 82.36±5.31 | 79.36±4.78 | 0.67 |
| Red meat intake(oz/day, mean, se) | 1.59±0.17 | 1.36±0.14 | 0.30 |
| Dietary Fiber(g/day, mean, se) | 23.03±1.28 | 25.58±1.76 | 0.27 |
Figure 1. Abundance of Fusobacterium in rectal mucosal biopsies from adenoma cases and non-adenoma controls.

qPCR results show that Fusobacterium is more abundant in cases than controls.
Table 2. Association between Fusobacterium abundance and colorectal adenomas.
| Categories* | Case (n = 48) | Control (n = 67) | OR (95% CI)** |
| Tertile 1 | 8 | 23 | Reference |
| Tertile 2 | 12 | 22 | 1.57 (0.54–4.57) |
| Tertile 3 | 28 | 22 | 3.66 (1.37–9.74) |
| P trend |
The abundance of Fusobacterium among control subjects were used to generate tertile cut-off. The lowest tertile of Fusobacterium abundance was considered as the reference.
Odds ratio and 95% confidence interval.
Compared to subjects with a low copy number, subjects with high abundance of Fusobacterium are more likely to be adenoma cases than controls.
Localization of Fusobacterium in colonic mucosal by FISH analysis
We Observed that Fusobacterium was over-represented in adenoma cases compared to non-adenoma controls, therefore, we performed histological evaluation by Fluorescence in situ Hybridization (FISH) using a Fusobacterium-specific probe to localize Fusobacterium in colonic mucosal tissue sections (Fig. 2a and 2b). The results show that Fusobacterium was localized in the mucus layer above the epithelium as well as within the colonic crypts. A general bacterial probe was also used as a positive control (Fig. 2c). Results confirm the presence of bacteria in the mucus layer.
Figure 2. Representative fluorescence in situ hybridization targeting Fusobacterium sp. in colorectal mucosal biopsy sections using bacterial 16S rRNA probes.

Fig. 2A–B are composite images of Cy3 and DAPI views of sections hybridized with a Fusobacterium-specific probe. Fusobacterium species is localized within the mucus layer of colorectal sections (A) 20X and 40X. Fusobacterium species is localized within the crypts of colorectal section (B) 20X and 40X. Fig. 2C (20X and 40X) is a positive control and shows sections stained with general bacteria probe (Eub 388). General bacteria, including most Eubacteria species, are localized to the mucus layer above the epithelium. White arrows point to bacteria either in mucus layer above the colonic epithelium or within the crypt.
There is a significant positive correlation between Fusobacterium species abundance and local inflammation in adenoma cases
Correlation of local inflammatory cytokine gene expression and Fusobacterium species abundance was analyzed separately for adenoma cases and non-adenoma controls. Analysis of cytokines IL-6, IL-10, IL-12, IL-17 and TNF-α and Fusobacterium was observed to have a significant positive correlation with local inflammation in cases, but not controls (Fig. 3). A significant positive correlation was found between abundance of Fusobacterium species and IL-10 (r = 0.443 p = 0.01). The correlation for TNF-α (r = 0.335 p = 0.06) was borderline significant. Although the correlations for IL-6 and IL-17 were positive, they did not reach statistical significance.
Figure 3. Correlations between Fusobacterium abundance and local cytokine gene expression in adenoma cases and non-adenoma controls.

Results suggest a significant positive correlation between Fusobacterium abundance and local inflammation in cases but not controls. The Correlations were significant for IL-10 (r = 0.44, p = 0.01) and TNF-α (r = 0.33, p = 0.06). *p<0.05.
Confirmatory Studies in Colorectal Cancer
Pyrosequencing analysis of 16s rRNA gene in colorectal cancer (CRC) tissue and matched normal colonic tissue revealed higher Fusobacterium species abundance in CRC compared to normal tissue
Previous studies reported an association between Fusobacterium species and colorectal cancer [8], [10], [11]. We reproduced these results by conducting high-throughput pyrosequencing analysis on 19 matched samples, 10 CRC tissues and 9 non-malignant matched controls from adjacent mucosa. All subjects were Caucasian and predominantly female, with ages ranging from 37–78 years. High-throughput sequencing revealed differences in abundance and richness in CRC compared to normal tissue. We identified 13 phyla, 24 classes and 176 bacteria genera. Overall, Shannon diversity and richness were higher in the CRC samples than matched normal tissue. Abundance of individual bacteria varied between groups. We observed reduced abundance of Bacteroidetes in CRC tissue compared to normal colon tissue, however, the distribution of the phylum Fusobacteria was higher in CRC tissue. The results showed a higher abundance of Fusobacterium in the CRC tissue compared to normal tissue. (Fig. 4).
Figure 4. Log Abundance of Fusobacterium in matched normal colon and colorectal cancer tissue.

Fusobacterium abundance was evaluated in DNA samples from normal colon and tumor tissue by qPCR using Fusobacterium-specific primers. Results suggest that Fusobacterium is increased in colon cancer tissue compared to normal tissue (t-test p = 0.0005).
qPCR validation of Fusobacterium species in colorectal cancer patients
qPCR analysis of Fusobacterium species in 10 CRC and 9 matching normal control tissues revealed a significant increase in abundance among colorectal cancer tissue compared to normal tissue, confirming previously reported results of higher Fusobacterium abundance in CRC patients. We also evaluated the relationship between CRC characteristics such as tumor location, treatment and Fusobacterium abundance. We did not observe significant associations for most of the tumor characteristics; however, we observed higher abundance of Fusobacterium species in the sigmoid than right side tumor location (Table 3). We further validated the pyrosequencing results by qPCR and observed significantly positive correlation between the two methods (r = 0.76, p = 0.0001).
Table 3. Relationship between Fusobacterium and colorectal tumor characteristics.
| Variable | Fusobacterium(copy #, mean, se) | P-value |
| Tumor Location | ||
| Right | 1.82±0.13 | |
| Transverse | 1.94±0.09 | NS |
| Sigmoid | 2.21±0.31 | 0.04 Sigmoid vs. Right |
| Stage | ||
| T-2 | 1.83±0.29 | |
| T-3 | 1.98±0.11 | 0.56 |
| Adjuvant Therapy | ||
| No | 2.16±0.03 | 0.20 |
| Yes | 2.01±0.10 |
Discussion
The human gut microbiota has been shown to have a dynamic and observable impact on the human host [4], [13]. While many of these bacteria are commensal and facilitate the maintenance of a healthy and functioning gastrointestinal tract, current research has shown that interactions between the host and the bacteria colonizing the gut can contribute to various diseases including colorectal carcinogenesis [12]–[15]. In particular, bacterial dysbiosis in the gut has been implicated in colorectal neoplasia, although no specific bacteria or bacterial signatures have been identified for colorectal adenomas [8], [9]. We evaluated the abundance of Fusobacterium in relation to colorectal adenomas in a case-control study and found that compared to controls, cases had significantly higher abundance of Fusobacterium.
There has been a recent focus on Fusobacterium, Gram-negative bacteria that usually colonize the oral cavity [10], [16]. Several groups have identified Fusobacterium in tumors of patients with colorectal carcinoma [8], [10], [17]–[19] and reported that the tumor tissue was characterized by a higher abundance of Fusobacterium than that of the normal colon. These results suggest Fusobacterium as a potential biomarker for colorectal carcinogenesis. However, it is not known whether Fusobacterium is associated with adenomas, early precursors of CRC. We observed significant differences in bacterial abundance between adenoma versus non-adenoma subjects and found that there was a strong positive correlation between high abundance of Fusobacterium and the presence of colorectal adenomas (p = 0.01). In particular those with high levels of Fusobacterium had about three and half fold increased risk of adenomas. As a CRC precursor, adenomas have become increasingly important in the study of colorectal carcinogenesis. Our results suggest that the changes in gut microbiota are associated with the earliest stages of tumor development [20]. Our results for colorectal adenomas and increased Fusobacterium levels are similar to previously reported studies involving Fusobacterium and colorectal cancer [8], [10], [11]. We validated the previously reported association between Fusobacterium and colorectal carcinoma in a set of matched CRC tumor and normal human colon tissue samples. Using both pyrosequencing and qPCR analysis of the 16S bacterial rRNA gene we were able to successfully reproduce these published results. We found that among CRC tumors and matched controls, Fusobacterium abundance was significantly higher in tumor tissue based on both qPCR as well as pyrosequencing analysis, with a significant correlation between both methods (r = 0.76, p = 0.0001).
We and others observed a difference in Fusobacterium abundance between the colorectal tumor and adjacent non-neoplastic tissue [10], [11], however, it would also be beneficial in future studies to assess the actual adenomas specifically, compared to normal mucosa. Our findings raise several important questions. With regard to colorectal adenomas and cancer, is Fusobacterium causative agent or an opportunistic colonizer? Does Fusobacterium act alone or in concert with other bacteria, viruses or fungi to promote CRC? Are there specific changes in the colonic environment that contribute to increased abundance of Fusobacterium in carcinogenesis? What are the mechanisms involved in this process? These questions will need to be addressed in future studies, particularly in animal models of adenomas and CRC to uncover whether Fusobacterium is a causative agent or opportunistic colonizer of colorectal tissue. Several potential factors such as host inflammation and altered gut environment (pH, bile acids, presence of adenoma or cancer) may contribute to the relationship between Fusobacterium and adenomas. These are addressed below.
Inflammation is a known risk factor for CRC [21], [22]. Interestingly, intestinal inflammation has been repeatedly linked to the gut microbiota [23], [24]. Commensal gut bacteria interact with the host in a symbiotic way to facilitate the operation of the intestinal immune system. However, as reported by several studies, bacterial dysbiosis may contribute to dysregulation of the immune system and mucus production in the gut, ultimately disrupting the delicate homeostatic relationship between commensal bacteria and the human host [3], [25], [26]. Uronis et al. [26] successfully demonstrated a link between the microbiota, intestinal inflammation and increased risk of colitis-associated colorectal cancer (CAC) in a mouse model. More specifically, Fusobacterium has been implicated as a pro-inflammatory pathogen [25], [27], [28] and has been found in a higher abundance of IBD patients [18].
Fusobacterium sp. have been found to flourish primarily in the oral cavity where they have been observed to behighly invasive [18], [29] and adherent [30]–[32]. The ability of Fusobacterium to attach to mucosal surfaces [28] makes it an ideal candidate to study in relation to host immunity and adenomas. Given the reports of previous studies that link Fusobacterium with inflammatory bowel diseases (IBD), we evaluated its relationship with several inflammatory markers. Specifically, we assessed mRNA expression of mucosal inflammatory cytokines IL-6, IL-10, IL-12, IL-17 and TNF-α in normal rectal biopsies and correlated their expression levels with abundance of Fusobacterium species in adenoma and non-adenoma subjects. We observed a positive correlation between the gene expression of several local cytokines and Fusobacterium species in adenoma cases, but not in controls. Similar to previously published findings [25], we saw a significant association between increased abundance of Fusobacterium and TNF-α. Dharmani et al. observed that the presence of invasive F. nucleatum strains correlated with increased TNF-α expression in IBD patients as well as experimental models [25]. Therefore, taking into consideration the observations that Fusobacterium is invasive and adherent, its link with IBD, its increased abundance in adenoma cases compared to controls, as well as a positive correlation with local inflammation, one could suggest that Fusobacterium may contribute to increased mucosal inflammation in adenoma subjects. However, we also detected a significant positive correlation between Fusobacterium and IL-10 expression in adenoma cases. While this observation with IL-10 is interesting, it highlights the complex and multi-factorial relationship between the host and its enteric intestinal bacteria.
The human host and the gut bacteria share a complex symbiotic relationship in which they both exert considerable influence on each other. Tumors are known to acidify their microenvironment and cause fluctuations in pH, thus the presence of an adenoma or cancer could lead to stress of the gut environment [33]. The presence of adenoma or cancer could change the luminal environment by altering levels of bile acids, pH, nutrient levels and redox potential, all of which could impact the gut microbial community and favor overgrowth of opportunistic pathogens. Altered colonic pH could affect metabolic activity, absorption of short-chain fatty acids (SCFA), and composition of gut microflora and mucosal cell proliferation [34]. Walker et al. observed that even a single unit change in pH affected the host microbial community composition as well as production of SCFA, especially among butyrate-producing species [35], a group within which Fusobacterium is included [36]–[39]. While most bile acids aid solubilization of lipids and facilitate nutrient absorption, some are transformed by intestinal bacteria into toxic secondary bile acids in the colon [40]. Thus, the transformation of secondary bile acids in the colon could have cytotoxic effects that may contribute to various gastrointestinal diseases including colorectal cancer [41]–[43]. Therefore, it is possible that Fusobacterium is a commensal that gains from the disruption of intestinal homeostasis as a result of the presence of adenoma or cancer. We evaluated the relationship between Fusobacterium species and adenoma size and frequency. However, there were no significant relationships observed between Fusobacterium and adenoma size (small, medium and large) or number of adenomas, suggesting that the abundance Fusobacterium in colonic mucosa is not impacted by adenoma size or frequency. These results are consistent with the findings of Castellarin et al. [10] who observed no association between Fusobacterium abundance and colorectal cancer tumor stage, site, treatment, patient age or survival.
Our goal was to study adenomas, precursors to colorectal cancer, to assess whether an association with Fusobacterium was present. We found an increased abundance of Fusobacterium among adenoma cases compared to controls. Our findings that Fusobacterium is associated with colorectal adenomas implicate its possible involvement early in carcinogenesis. Our work builds on the findings of previous studies reporting an association between Fusobacterium and colorectal cancer. [8], [10], [11]. Our observation of a positive correlation between Fusobacterium and mucosal inflammation in adenoma cases suggests that this relationship may possibly be mediated by inflammation. However, given the complex environment in which Fusobacterium exists with other bacteria [18], we exercise caution in assigning causality as we recognize that more mechanistic studies are needed to elucidate such an association. Recently the alpha-bug and driver-passenger models of CRC proposed by Sears et al. and Tjalsma et al. respectively [44], [45] defined bacterial drivers (or alpha bugs) as gut bacteria with pro-carcinogenic features such as possession of virulence factors, ability to directly modulate mucosal immune responses and ability to alter bacterial community composition to favor proliferation of opportunistic bacteria (passengers). Thus, under these models Fusobacterium could be an important player. Future studies in animal models could help tease apart the precise contribution of Fusobacterium and other bacteria to colorectal carcinogenesis.
Materials and Methods
Ethics Statement
Institutional approval was provided by University of North Carolina, School of Medicine Institutional Review Board.
Study Population and Sampling
Two sets of human samples were analyzed in this study. The first group of subjects was drawn from participants in the Diet and Health Study V who underwent routine colonoscopy screening at UNC Hospitals, Chapel Hill, NC. Eligible subjects 30 years of age or older gave written informed consent to provide colorectal biopsies as well as a phone interview involving questions about diet and lifestyle. At the time of the colonoscopy procedure, the research assistant obtained anthropometric measures to determine body mass index (BMI) and waist–hip ratio (WHR) [13], [20]. Biopsy samples from a total of 115 randomly selected subjects (48 adenoma cases and 67 non-adenoma controls) were used in this study. Subjects with known or suspected colorectal cancer or with inadequate colon prep were excluded from the study. Before the endoscopy procedure was performed, biopsies were taken 8–12 cm from the anal verge of the normal rectal mucosa, and immediately flash frozen in liquid nitrogen. Biopsies were stored at −80°C. Participants with reported adenomas were classified as “cases” and those with no adenomas as “controls” [20].
The second group of samples was made up of de-identified matched tumor and normal tissue biopsies from 10 patients with colorectal cancer. Samples were obtained from UNC Tissue Procurement Facility to confirm previously reported studies.
Fusobacterium Culture
Fusobacterium nucleatum subsp. nucleatum (ATCC® 25586™) was obtained from the American Type Culture Collection (ATCC) and cultured according to their instructions for use as a positive control. The strain was grown on Reinforced Clostridial Medium (Difco, Becton Dickinson, Franklin Lakes, NJ) under anaerobic conditions at 37°C.
DNA Extraction
DNA was extracted from normal rectal mucosal biopsies as well as matched tumor/normal tissue using the Qiagen DNeasy Blood and Tissue Kit (Cat# 69504) and a modified protocol with lysozyme and bead-beating [13], [20]. Pure cultures of F. nucleatum were centrifuged, re-suspended in kit-provided lysis buffer, and DNA extraction was performed using the same extraction method used for biopsies.
Pyrosequencing of Colorectal Tumors and Matched Normal Tissue
For CRC tumor and matched normal tissue, the pyrosequencing protocol is as described previously [20]. Briefly, the bacterial 16S rRNA gene was amplified from each DNA sample using barcode-tagged universal 16S primers that span the V1–V3 variable regions. PCR amplification and detection were performed using 454 GS FLX Titanium technology in the UNC Microbiome Core Facility. The primers were composed of the Roche Titanium Fusion Primer A (5′-CGTATCGCCTCCCTCGCGCCATCAG-3′), a 10 bp MID barcode (Roche, Indianapolis, IN) unique to each of the samples and the universal bacteria primer 27F (5′-AGAGTTTGATCCTGGCTCAG-3′)The reverse primers were composed of the Roche Titanium Primer B (5′-CTATGCGCCTTGCCAGCCCGCTCAG-3′) the identical 10 bp MID as the forward primer and the reverse bacteria primer 338R (5′-TGCTGCCTCCCGTAGGAGT-3′) [46].
Analysis of Bacterial Pyrosequencing Data from CRC Tissue
Analysis of deep sequencing data from CRC tumor and matched normal tissue was performed using QIIME [47]. Briefly, the combined raw sequencing data as well as metadata describing the samples were de-multiplexed and filtered for quality control, following which the data were denoised by PyroNoise and sequences were grouped into OTUs (Operational Taxonomic Units) at 97% sequence similarity to approximate genus-level phylotypes. After taxonomic assignment of OTUs, sequences were aligned for phylogenetic analysis as well as alpha and beta diversity calculations. In addition to t-tests, Wilcoxon-rank sum test was used to compare bacterial abundance and diversity in CRC tumor versus normal tissue.
Quantitative Real-Time PCR (qPCR)
For mucosal samples from adenoma cases and non-adenoma controls as well as CRC tumors and matching normal tissue, qPCR was performed to quantify the abundance of Fusobacterium species. A standard curve was generated by amplifying a16S rDNA region of F. nucleatum (ATCC® 25586™) using Fusobacterium-specific primers [48]. Concentration of the PCR product was measured by absorbance and the number of fragment copies was calculated using the following formula:
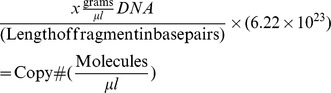 |
Copy number was adjusted to a starting concentration of 1.00×1010 and serial dilutions were performed to create nine standards. 25 µl reactions were prepared containing template DNA, 10µM primer mix, and Fast-SYBR Green Master Mix (Applied Biosystems). The qPCR was performed with an annealing temperature of 60°C for 40 cycles. Finally, the copy number was calculated based on the standard curve, which was adjusted to a starting DNA concentration of 50 ng/µL using the following formula to the unadjusted values:
where A is the concentration of the template DNA and B is the dilution, 1∶10.
Reverse Transcriptase PCR (RT-PCR) for Local Inflammatory Cytokines in Adenoma Cases and Non-adenoma Controls
RT-PCR was performed to assess mRNA expression of inflammatory cytokines IL-6, IL-10, IL-12, IL-17 and TNF-α using ready-to-use optimized primers (SA Biosciences). Expression of each inflammatory cytokine was assessed relative to the housekeeping gene hydroxymethylbilane synthase (HMBS). The qPCR was performed using SYBR Green Master Mix (Applied Biosystems) and each sample was run in duplicate. qPCR results were normalized using the expression of the HMBS gene [49].
Fluorescence in situ Hybridization (FISH)
FISH was performed on paraffin sections of mucosal biopsies fixed with Carnoy’s fixative using a universal bacteria probe (EUB388) and a Fusobacterium-specific probe. These assays used a previously described protocol [13]. Pure cultures of E. Coli and Fusobacterium were used as positive controls.
Statistical Analysis
Mean and standard errors were computed for continuous variables. Comparison of continuous variables such as age (years), waist hip-ratio, body mass index (Kg/m2), alcohol (grams/day) calories (Kcal/day), fat (grams/day), red meat (ounces/day) and dietary fiber intake (grams/day) between adenoma cases and non-adenoma controls were made using t-tests. Categorical variables for CRC subjects (tumor location, disease stage, adjuvant therapy) were compared by Fisher’s Exact Test. Fusobacterium copy numbers were assessed for normality and log transformed. Mean log Fusobacterium abundance was compared between case and control subjects using t-test. The distribution of Fusobacterium among control subjects were used to generate tertile values for logistic regression analysis. The lowest tertile of Fusobacterium abundance was considered as the reference. Correlation of Fusobacterium and local gene expression of TNF-α, IL-17, IL-12, IL-10 and IL-6 were assessed by Spearman's correlation coefficient.
Supporting Information
Relationship between Fusobacterium abundance and adenoma size.
(DOCX)
Acknowledgments
We wish to thank Dr. Belén Cadenas and the UNC Microbiome Core Facility for expertise and assistance with Fusobacterium culture. We wish to acknowledge the UNC Lineberger Comprehensive Cancer Center Tissue Procurement Core Facility, UNC CGIBD Histology Core (Carolyn Suitt), and the UNC Microscopy Services Laboratory (Dr. Robert Bagnell) and Ward Jarvis for assistance with FISH and microscopy.
Funding Statement
This work was supported by grants from the National Institutes of Health NIH R01 CA04468, R01 CA136887, P50 CA106991 and P30 DK034987. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Chow J, Lee SM, Shen Y, Khosravi A, Mazmanian SK (2010) Advances in Immunology: Elsevier. 243–274 p. [DOI] [PMC free article] [PubMed]
- 2. Savage DC (1977) Microbial ecology of the gastrointestinal tract. Annual review of microbiology 31: 107–133. [DOI] [PubMed] [Google Scholar]
- 3. Arthur JC, Jobin C (2010) The struggle within: Microbial influences on colorectal cancer. Inflammatory Bowel Diseases 17: 396–409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Mutch DM (2004) Impact of commensal microbiota on murine gastrointestinal tract gene ontologies. Physiological Genomics 19: 22–31. [DOI] [PubMed] [Google Scholar]
- 5. Kaur N, Chen CC, Luther J, Kao JY (2011) Intestinal dysbiosis in inflammatory bowel disease. Gut Microbes 2: 211–216. [DOI] [PubMed] [Google Scholar]
- 6. Sasaki M, Klapproth J-MA (2012) The role of bacteria in the pathogenesis of ulcerative colitis. Journal of Signal Transduction 2012: 704953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang F, Zhang P, Jiang H, Cheng S (2012) Gut Bacterial Translocation Contributes to Microinflammation in Experimental Uremia. Digestive Diseases and Sciences. [DOI] [PubMed]
- 8. Marchesi JR, Dutilh BE, Hall N, Peters WHM, Roelofs R, et al. (2011) Towards the Human Colorectal Cancer Microbiome. PloS one 6: e20447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Sobhani I, Tap J, Roudot-Thoraval F, Roperch JP, Letulle S, et al. (2011) Microbial Dysbiosis in Colorectal Cancer (CRC) Patients. PloS one 6: e16393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Castellarin M, Warren RL, Freeman JD, Dreolini L, Krzywinski M, et al. (2012) Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Research 22: 299–306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Kostic AD, Gevers D, Pedamallu CS, Michaud M, Duke F, et al. (2012) Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Research 22: 292–298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Azcarate-Peril MA, Sikes M, Bruno-Barcena JM (2011) The intestinal microbiota, gastrointestinal environment and colorectal cancer: a putative role for probiotics in prevention of colorectal cancer? Am J Physiol Gastrointest Liver Physiol 301: G401–424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Shen XJ, Rawls JF, Randall T, Burcal L, Mpande CN, et al. (2010) Molecular characterization of mucosal adherent bacteria and associations with colorectal adenomas. Gut microbes 1: 138–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Hakansson A, Molin G (2011) Gut Microbiota and Inflammation. Nutrients 3: 637–682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Round JL, Mazmanian SK (2009) The gut microbiota shapes intestinal immune responses during health and disease. Nature Reviews Immunology 9: 313–323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Swidsinski A, Dorffel Y, Loening-Baucke V, Theissig F, Ruckert JC, et al. (2010) Acute appendicitis is characterised by local invasion with Fusobacterium nucleatum/necrophorum. Gut 60: 34–40. [DOI] [PubMed] [Google Scholar]
- 17.Allen-Vercoe E, Strauss J, Chadee K (2011) Fusobacterium nucleatum: An emerging gut pathogen? Gut microbes 2. [DOI] [PubMed]
- 18. Strauss J, Kaplan GG, Beck PL, Rioux K, Panaccione R, et al. (2011) Invasive potential of gut mucosa-derived fusobacterium nucleatum positively correlates with IBD status of the host. Inflammatory Bowel Diseases 17: 1971–1978. [DOI] [PubMed] [Google Scholar]
- 19. Strauss J, White A, Ambrose C, McDonald J, Allen-Vercoe E (2008) Phenotypic and genotypic analyses of clinical Fusobacterium nucleatum and Fusobacterium periodonticum isolates from the human gut. Anaerobe 14: 301–309. [DOI] [PubMed] [Google Scholar]
- 20.Sanapareddy N, Legge RM, Jovov B, McCoy A, Burcal L, et al.. (2012) Increased rectal microbial richness is associated with the presence of colorectal adenomas in humans. The ISME journal: 1–11. [DOI] [PMC free article] [PubMed]
- 21. Coussens LM, Werb Z (2002) Inflammation and cancer. Nature 420: 860–867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Tanaka T (2009) Colorectal carcinogenesis: Review of human and experimental animal studies. J Carcinog 8: 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Rogler G, Haller D, Jobin C (2010) Microbiota in Chronic Mucosal Inflammation. International Journal of Inflammation 2010: 1–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Tlaskalová-Hogenová H, Štěpánková R, Hudcovic T, Tučková L, Cukrowska B, et al. (2004) Commensal bacteria (normal microflora), mucosal immunity and chronic inflammatory and autoimmune diseases. Immunology Letters 93: 97–108. [DOI] [PubMed] [Google Scholar]
- 25. Dharmani P, Strauss J, Ambrose C, Allen-Vercoe E, Chadee K (2011) Fusobacterium nucleatum Infection of Colonic Cells Stimulates MUC2 Mucin and Tumor Necrosis Factor Alpha. Infection and Immunity 79: 2597–2607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Uronis JM, Mühlbauer M, Herfarth HH, Rubinas TC, Jones GS, et al. (2009) Modulation of the Intestinal Microbiota Alters Colitis-Associated Colorectal Cancer Susceptibility. PloS one 4: e6026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Liu H, Redline RW, Han YW (2007) Fusobacterium nucleatum induces fetal death in mice via stimulation of TLR4-mediated placental inflammatory response. J Immunol 179: 2501–2508. [DOI] [PubMed] [Google Scholar]
- 28. Swidsinski A, Dorffel Y, Loening-Baucke V, Theissig F, Ruckert JC, et al. (2011) Acute appendicitis is characterised by local invasion with Fusobacterium nucleatum/necrophorum. Gut 60: 34–40. [DOI] [PubMed] [Google Scholar]
- 29. Han YW, Shi W, Huang GT, Kinder Haake S, Park NH, et al. (2000) Interactions between periodontal bacteria and human oral epithelial cells: Fusobacterium nucleatum adheres to and invades epithelial cells. Infection and Immunity 68: 3140–3146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Edwards AM, Grossman TJ, Rudney JD (2005) Fusobacterium nucleatum Transports Noninvasive Streptococcus cristatus into Human Epithelial Cells. Infection and Immunity 74: 654–662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Han YW, Ikegami A, Rajanna C, Kawsar HI, Zhou Y, et al. (2005) Identification and Characterization of a Novel Adhesin Unique to Oral Fusobacteria. Journal of bacteriology 187: 5330–5340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Weiss EI, Shaniztki B, Dotan M, Ganeshkumar N, Kolenbrander PE, et al. (2000) Attachment of Fusobacterium nucleatum PK1594 to mammalian cells and its coaggregation with periodontopathogenic bacteria are mediated by the same galactose-binding adhesin. Oral microbiology and immunology 15: 371–377. [DOI] [PubMed] [Google Scholar]
- 33. Griffiths JR (1991) Are cancer cells acidic? Br J Cancer 64: 425–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Newmark HL, Lupton JR (1990) Determinants and consequences of colonic luminal pH: implications for colon cancer. Nutr Cancer 14: 161–173. [DOI] [PubMed] [Google Scholar]
- 35. Walker AW, Duncan SH, McWilliam Leitch EC, Child MW, Flint HJ (2005) pH and peptide supply can radically alter bacterial populations and short-chain fatty acid ratios within microbial communities from the human colon. Appl Environ Microbiol 71: 3692–3700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Hamer HM, Jonkers D, Venema K, Vanhoutvin S, Troost FJ, et al. (2008) Review article: the role of butyrate on colonic function. Aliment Pharmacol Ther 27: 104–119. [DOI] [PubMed] [Google Scholar]
- 37. Neish AS (2009) Microbes in gastrointestinal health and disease. Gastroenterology 136: 65–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Pryde SE, Duncan SH, Hold GL, Stewart CS, Flint HJ (2002) The microbiology of butyrate formation in the human colon. FEMS Microbiol Lett 217: 133–139. [DOI] [PubMed] [Google Scholar]
- 39. Resmer KL, White RL (2011) Metabolic footprinting of the anaerobic bacterium Fusobacterium varium using 1H NMR spectroscopy. Mol Biosyst 7: 2220–2227. [DOI] [PubMed] [Google Scholar]
- 40. Degirolamo C, Modica S, Palasciano G, Moschetta A (2011) Bile acids and colon cancer: Solving the puzzle with nuclear receptors. Trends Mol Med 17: 564–572. [DOI] [PubMed] [Google Scholar]
- 41. McGarr SE, Ridlon JM, Hylemon PB (2005) Diet, anaerobic bacterial metabolism, and colon cancer: a review of the literature. J Clin Gastroenterol 39: 98–109. [PubMed] [Google Scholar]
- 42. Ridlon JM, Kang DJ, Hylemon PB (2006) Bile salt biotransformations by human intestinal bacteria. J Lipid Res 47: 241–259. [DOI] [PubMed] [Google Scholar]
- 43. Ignacio Barrasa J, Olmo N, Perez-Ramos P, Santiago-Gomez A, Lecona E, et al. (2011) Deoxycholic and chenodeoxycholic bile acids induce apoptosis via oxidative stress in human colon adenocarcinoma cells. Apoptosis 16: 1054–1067. [DOI] [PubMed] [Google Scholar]
- 44. Sears CL, Pardoll DM (2011) Perspective: alpha-bugs, their microbial partners, and the link to colon cancer. J Infect Dis 203: 306–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Tjalsma H, Boleij A, Marchesi JR, Dutilh BE (2012) A bacterial driver-passenger model for colorectal cancer: beyond the usual suspects. Nat Rev Microbiol 10: 575–582. [DOI] [PubMed] [Google Scholar]
- 46. Fierer N, Hamady M, Lauber CL, Knight R (2008) The influence of sex, handedness, and washing on the diversity of hand surface bacteria. Proc Natl Acad Sci U S A 105: 17994–17999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, et al. (2010) QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7: 335–336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Walter J, Margosch D, Hammes WP, Hertel C (2011) Detection of Fusobacterium Species in Human Feces Using Genus-Specific PCR Primers and Denaturing Gradient Gel Electrophoresis Microbial Ecology in Health and Disease 14.
- 49. Jovov B, Araujo-Perez F, Sigel CS, Stratford JK, McCoy AN, et al. (2012) Differential Gene Expression between African American and European American Colorectal Cancer Patients. PloS one 7: e30168. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Relationship between Fusobacterium abundance and adenoma size.
(DOCX)