

SUMMARY
Genetic mosaic approach is commonly employed in the Drosophila eye by completely abolishing or misexpressing a gene within a subset of cells to unravel its role during development. Classical genetic mosaic approach involves random clone generation in all developing fields. Consequently, a large sample size needs to be screened to generate and analyze clones in specific domains of the developing eye. To address domain specific functions of genes during axial patterning, we have developed a system for generating mosaic clones by combining Gal4/UAS and FLP/FRT system which will allow generation of loss-of-function as well as gain-of-function clones on the dorsal and ventral eye margins. We used the bifid-Gal4 driver to drive expression of UAS-flippase (FLP). This reagent can have multiple applications in (i) studying spatio-temporal function of a gene during dorso-ventral axis specification in the eye, (ii) analyzing genetic epistasis of genes involved in DV patterning and (iii) conducting genome wide screens in a domain specific manner.
Keywords: Drosophila eye, Dorso-ventral (DV) axis, patterning, genetic mosaic, Gal4/UAS technique, compartments
Historically, researchers have exploited the ability to create genetic mosaics to circumvent the problems of lethality associated with studies to analyze function of essential genes. In Drosophila, the genetic mosaic techniques, where animals containing a distinct population of somatic cells lacking the function of a gene, have been used to address many basic biological questions including restriction of cell fates during patterning and growth (Blair, 2003; Duffy et al., 1998; Golic, 1991; Lee and Luo, 1999, 2001; Newsome et al., 2000; Stowers and Schwarz, 1999; Xu and Rubin, 1993). Several other approaches were developed that allowed the gain-of-function as well as the loss-of- function of two different genes in the same set of cells, for example MARCM system (Lee and Luo, 1999, 2001). The development of genetic tools and reagents over the last three decades has vastly facilitated the analysis of gene function along the spatial and temporal axis. Majority of the genetic components of known pathways have been identified but the crosstalk among the various established pathways is yet to be understood. To test and validate the crosstalk among the pathways in a specific context of time, space or domain, it is important to develop simple but effective strategies to study the genetic hierarchy using genetic epistatic approaches. Thus, there is need to develop new tools which can be employed to test both loss-of-function as well as gain-of-function of two different genes in a subset of cells within a developing field.
Genetic mosaic approach has been used in the Drosophila eye to address many biological questions including study of cell fate, patterning, growth, cell death, and disease. There are several genetic mosaic approaches available in Drosophila (Blair, 2003; Golic, 1991; Golic and Lindquist, 1989; Lee and Luo, 1999, 2001; Newsome et al., 2000; Stowers and Schwarz, 1999; Xu and Rubin, 1993). Drosophila eye has also been used for genome wide screens which exploit the FLP recombinase from the yeast to generate FRT mediated recombination to screen for the genes affecting patterning, growth, death and disease (Blair, 2003; Call et al., 2007; Duffy et al., 1998; Janody et al., 2004; Morante et al., 2011; Newsome et al., 2000). The flippase (FLP) enzyme catalyzes recombination at sequence motifs termed FRTs. The heat shock mediated FLP expression to generate random clones may not be suitable in several instances as it may cause lethality due to the fact that extensive numbers of clones are produced (Duffy et al., 1998). The approach of generating random clones to discern the gene function has proved to be extremely useful however, in case of domain specific analysis using this approach requires sampling of a large number of clones. Most of these strategies provide excellent tools for generation of clones in the entire developing eye field. During organogenesis, a developing field gets further sub-divided into smaller regions called compartments (Blair, 2001; Curtiss et al., 2002; Dahmann et al., 2011; Singh et al., 2012). The properties of the cells of a compartment are unique and the cells within a compartment behave differently based on their response to the morphogen gradients. Therefore, to investigate the fine tuning of gene functions in the developing fields, it is important to design tools that will facilitate knocking down a gene in a subset of cells in a developing field or compartment.
We have combined the Gal4/UAS system used for targeted misexpression (Brand and Perrimon, 1993) with the FLP/FRT system (Evans et al., 2009) to generate genetic mosaics. In our study, we have employed an optomotor blind (omb) or bifid (hereafter bi) Gal4 driver (Calleja et al., 1996; Lecuit et al., 1996) to selectively target FLP on the Dorso-Ventral (DV) margins of the eye. The FLP will act on the DV margins to enable generation of genetic mosaic clones in a domain specific manner. Domain specific clonal analysis is essentially a useful strategy to maximize the yield of the clones of desired genotype and where sampling hundreds of clones are not required. There are several other eye specific enhancers which are being used like Glass Multiple Repeat (GMR)-FLP (Lee and Luo, 2001) or eyeless (ey)-FLP (Newsome et al., 2000). But these enhancers will not be able to address the requirement for the dorso-ventral axis determination in the eye as GMR will drive expression in all the differentiating neurons and eyeless will drive expression in the entire early eye field.
We first investigated the expression of bi-Gal4 driver in the developing imaginal disc using a UAS-GFP reporter gene. In the early first instar eye imaginal disc bi-Gal4 drives the expression of GFP (bi>GFP) reporter in a small subset of cells in the posterior margin in the glial precursor cells in the optic nerve (Fig. 1a). In the late first instar eye disc bi-Gal4 expression refines and starts expressing in a couple of cells on the margin of the eye imaginal disc (Fig. 1B; arrow). In the early second instar bi-Gal4 driven GFP reporter expression becomes robust in the cells on both the dorsal and ventral margins of the developing eye imaginal disc (Fig. 1C). During late second instar stage of development, the bi-Gal4 expression domain remain restricted to the dorsal and ventral eye margins and the number of cells expressing the GFP reporter is increased to 100–150 cells (Fig. 1D). In the third instar eye imaginal disc, bi-Gal4 expression is observed in the dorsal and the ventral margins as well as in some glial cells within the eye field (Fig. 1E). The third instar eye imaginal discs exhibit clearly demarcated eye field as evident from the expression of the pan neural marker ELAV that marks the photoreceptor neurons (Fig. 1E). Thus, bi-Gal4 domain is restricted to the dorsal and ventral margins of the developing eye field right from early larval development. The adult eye pigmentation pattern which is dependent on expression of mini-white reporter exhibits DV specific distribution in bi-Gal4 flies. The mini-white expression is enriched on the dorsal and ventral margins of the adult eye and is absent near the equator (Fig. 1F).
FIG. 1. Domains of bi-Gal4 expression in Drosophila imaginal discs.

bifid-Gal4 (Calleja et al., 1996; Lecuit et al., 1996) drives Green Fluorescent Protein (GFP) reporter expression on the dorsal and ventral margins of the developing eye-antennal imaginal disc of (A, B) first-, (C, D) second- and (E) third- instar larval eye disc. (B) Arrow in the late first instar disc marks the cells where bi-Gal4 drives expression on the margin. Note that imaginal discs are stained with Disc large (Dlg: red) a membrane specific reporter, a pan neural marker ELAV (blue). (F) The mini-white reporter in bi-Gal4 shows a domain specific expression on the dorsal and ventral margins of the adult eye. (G–I) bi-Gal4 drive GFP expression in (G) leg, and (H, I) wing imaginal disc. The magnification of images is as follows: panel A–E, G, H is 20X, panel F is 10X and panel I is 40X.
The expression of bi-Gal4 driver is not only restricted to the developing eye imaginal disc but is also present in the other developing fields including wing and leg imaginal discs (Lecuit et al., 1996). In the leg imaginal disc bi-Gal4 can drive expression in a proximal subset of cells (Fig. 1G). In the wing imaginal disc the expression of bi-Gal4 driven GFP reporter was observed in a domain which extends both in the anterior and posterior compartments as well as the dorsal and ventral margin, in the area which gives rise to the wing blade (Fig. 1H, I).
Using suitable genetic crosses to combine the FLP/FRT and Gal4/UAS system, we generated stocks where bi-Gal4 can drive expression of FLP within its expression domain, and trigger homologous recombination at the site of the FRT cassettes to generate homozygous loss-of-function clones (Fig. 2). We established fly stocks which allow generation of mutant clones for genetic mutations on particular arms (2R, 3R) of the second and third chromosomes. In the first set of flies, the genetic mosaic clones can be detected by loss of GFP (ubi-GFP) expression. Secondly, we tested the approach where the homozygous wild-type twin clones generated by recombination in somatic cells can be eliminated by using the “cell-lethal” mutations (Newsome et al., 2000; Stowers and Schwarz, 1999). The “cell-lethal” approach can prove advantageous to study the effects of loss-of-function of slow growing cells of mutant clones that are otherwise competed out by their wild-type counterparts or neighbors. This approach can be useful in addressing the issues of cell-competition (Newsome et al., 2000; Stowers and Schwarz, 1999). Furthermore, these tools allow misexpression of transgenes (UAS-X) of interest in the dorsal and ventral margins of the developing eye. Since misexpression of genes will be restricted to a smaller domain within the eye, it will facilitate the analysis of gene function which may otherwise be compromised by the death of the disc or organism.
FIG. 2.

Schematics of genetic crosses to generate reagents for the dorso-ventral genetic mosaic system on the second and third chromosome.
Generation of Clones
We tested these stocks using the mutant alleles of Lobe (L), a gene involved in ventral eye development. Loss-of-function of L results in loss of the ventral eye whereas the dorsal clones do not have any effect (Singh and Choi, 2003). Earlier L clones were generated using an ey promoter which drives FLP expression in the entire developing eye field continuously (Reuter et al., 2003; Singh and Choi, 2003). Even though L is required for ventral eye development, it results in generation of L loss-of-function clones in the entire developing eye field during all stages of development. Loss-of-function clones of L using the bi-Gal4 driven FLP mediated recombination of FRT cassette on 2R resulted in selective loss of ventral eye (Fig. 3C, D). These clones were generated using the “cell-lethal” approach (Newsome et al., 2000). It has been shown that L loss-of-function phenotype can be rescued by blocking caspase dependent cell death (Singh et al., 2006) using misexpression of the baculo-virus P35 transgene caspase inhibitor (Hay et al., 1994). We also tested if our domain specific genetic mosaic approach can rescue this loss of ventral eye phenotype of L loss-of-function clones by blocking caspase dependent cell death. Misexpression of P35 on the dorsal and ventral margin using our bi-Gal4 driver can rescue the loss of ventral eye phenotype of L loss-of-function clones (Fig. 3E, F). We also tested the other strategy of making random clones by heat shock FLP or ey promoter linked to FLP to make clones of L both on dorsal and ventral margins of the developing eye and found similar phenotypes of loss of ventral eye (data not shown). This phenotype of loss of ventral in L mutant clones can be rescued by misexpression of P35 (Fig. 3E, F). The third chromosomal stocks were also used to generate clones of homothorax (hth). As reported earlier, loss of function of hth resulted in enlargement of the ventral eye (Pai et al., 1998); we found similar ventral eye enlargements using our system (data not shown).
FIG. 3. Phenotypes of loss-of-function clones on dorsal and ventral eye margins.
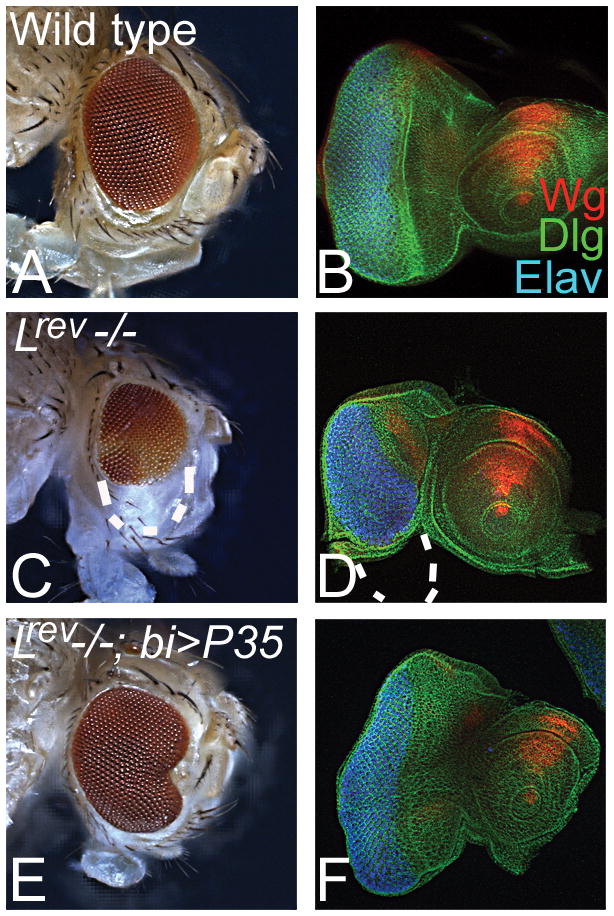
(A, B) Wild-type (A) adult eye and (B) eye imaginal disc. (B) Eye imaginal disc is stained for a membrane specific marker Disc large (Dlg: green channel), a signaling molecule Wingless (Wg: red channel), and a pan neural marker Elav (blue) that marks the photoreceptor neurons of the eye disc. (C, D) Loss-of-function clones of L on the dorsal and ventral margins of the developing eye imaginal disc by using cell-lethal strategy results in preferential loss of ventral eye phenotype as observed in the (C) adult eye and the (D) eye imaginal disc. The outline of ventral eye is marked by white dotted line both in the adult eye and the eye imaginal disc. Note that the dorsal eye margin does not exhibit any effect on the eye development and differentiation. (E, F) Misexpression of P35, to block caspase dependent cell death, in loss-of-function clones of L (using cell-lethal approach) in dorsal and ventral eye margins, result in suppression of loss-of-ventral eye phenotype as seen in the (E) adult eye and (F) eye imaginal disc. These reagents can be used for generating loss-of-function of a gene along with gain-of-function of another gene both in the dorsal and ventral eye margins. Magnification of all adult eye images (panel A, C, E) is 10X and all imaginal discs images (panel B, D, F) are 20X.
Thus, overall this directed genetic mosaic approach using bi-Gal4 presents an efficient strategy to assay the effect of essential genes and their lethal mutations in specific tissues (domains) and time windows. Our results further suggest that these reagents will serve as an excellent tool for testing the function of genes involved in DV patterning of the eye, and genetic epistasis of genes involved in DV patterning of eye. Furthermore, these reagents can be used for genome-wide screening approaches using either loss-of-function clones or misexpression of a gene of interest only on the dorsal and ventral margins of the eye. The cells outside of the Gal4 expression domain in the eye imaginal disc may serve as the control. Temperature sensitive Gal80 ts alleles are available which can block the Gal4 activity (McGuire et al., 2003; McGuire et al., 2004). In the future we can also introduce the Gal80ts in this bi-Gal4 mediated domain specific clone system to develop a more efficient temporal control. We anticipate this system will provide an efficient, genetic tool in the hands of fly researchers to study dorso-ventral patterning in the Drosophila eye.
METHODS
Stocks used in this study were bi-Gal4 (Calleja et al., 1996; Lecuit et al., 1996); y, w; UAS-NLS-GFPS65T (Ito et al., 1997), UAS-FLP/TM6BTb, UAS-FLP/CyO (Duffy et al., 1998), y,w; FRT42D cl w+/CyO (Newsome et al., 2000), y, w; FRT42D ubi-GFP/CyO, y,w; FRT82B ubi-GFP/TM6B Tb. These stocks are described in flybase (http://flybase.bio.indiana.edu). The flies were maintained on standard fly food at 25°C. An outline of the genetic crosses for generation of flies required for domain specific genetic mosaic approach is shown in Fig. 2. The various genotypes used for this study are:
bi-Gal4 (X); FRT42D ubi-GFP/CyO (II); UAS-FLP/TM6B Tb (III)
bi-Gal4 (X); FRT42D cl w[+]/CyO (II); UAS-FLP/TM6B Tb (III)
bi-Gal4 (X); UAS-FLP/CyO (II); FRT82 ubi-GFP/TM6B Tb (III)
Roman numerals denote the chromosomes. The UAS-FLP and FRT stocks were obtained from the Bloomington Stock center.
Genetic Mosaic analysis
We employed genetic mosaic approach to generate loss-of-function clones in the eye (Xu and Rubin, 1993). For the generation of clones in the eye, we have used bi-GAL4 driven UAS-FLP as source of FLP on the dorsal and ventral margin of the eye field. To generate mosaic clones of L in the eye, bi-GAL4; FRT42D ubi-GFP; UAS-FLP virgins were crossed to males of LrevFRT42D/CyO, (ii) LrevFRT42D/CyO; UAS-P35. To test third chromosomal reagent, we crossed bi-Gal4; UAS-FLP/CyO; FRT82 ubi-GFP/TM6B Tb virgins to y, w; FRT 82B hth100-1/TM6BTb males. Mutant tissue was marked by the absence of GFP reporter in the eye imaginal disc. We also extended the bi-Gal4 mediated generation of loss-of-function clones where wild-type cells were eliminated using the cell lethal approach. All these reagents will be readily available to the research community upon acceptance of the manuscript.
Immunohistochemistry
Imaginal discs were dissected from first-, second- and wandering third-instar larvae in 1XPBS and stained following the standard protocol (Singh et al., 2002). Antibodies used were rat anti-Elav (1:100), mouse anti-Wg (1:50) (Developmental Studies Hybridoma Bank), rabbit anti-Dlg (1:200; a gift from K. Cho). Secondary antibodies (Jackson Laboratories) used in this study were goat anti-rat IgG conjugated with Cy5 (1:200), donkey anti-rabbit IgG conjugated to Cy3 (1:250), donkey anti-rabbit IgG conjugated to FITC, and donkey anti-mouse IgG conjugated to Cy3 (1:200). Tissues were mounted on slides in Vectashield (Vector Laboratories). Immunofluorescent images were analyzed using the Olympus Fluoview 1000 Laser Scanning Confocal Microscope.
Bright Field Imaging
Adult eye images were taken on Axioimager.Z1 Zeiss Apotome. Adult flies were mounted on a needle and then individual image stacks were generated using Z-sectioning approach. The final image was generated using extended depth of focus function of Axiovision software version 4.6.3.
Acknowledgments
Authors thank Bloomington Stock Centre, Henry Sun, Justin Kumar, K Cho, and Kwang-Wook Choi for fly reagents and members of Singh and Kango-Singh Lab for the comments on the manuscript. MT and ORP are supported by the graduate program at the University of Dayton. AS is supported by the NIH grant (1R15 HD064557-01), and MKS is supported by start-up support from the University of Dayton.
LITERATURE CITED
- Blair SS. Cell lineage: compartments and Capricious. Curr Biol. 2001;11:R1017–1021. doi: 10.1016/s0960-9822(01)00614-5. [DOI] [PubMed] [Google Scholar]
- Blair SS. Genetic mosaic techniques for studying Drosophila development. Development. 2003;130:5065–5072. doi: 10.1242/dev.00774. [DOI] [PubMed] [Google Scholar]
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. doi: 10.1242/dev.118.2.401. [DOI] [PubMed] [Google Scholar]
- Call GB, Olson JM, Chen J, Villarasa N, Ngo KT, Yabroff AM, Cokus S, Pellegrini M, Bibikova E, Bui C, Cespedes A, Chan C, Chan S, Cheema AK, Chhabra A, Chitsazzadeh V, Do MT, Fang QA, Folick A, Goodstein GL, Huang CR, Hung T, Kim E, Kim W, Kim Y, Kohan E, Kuoy E, Kwak R, Lee E, Lee J, Lin H, Liu HC, Moroz T, Prasad T, Prashad SL, Patananan AN, Rangel A, Rosselli D, Sidhu S, Sitz D, Taber CE, Tan J, Topp K, Tran P, Tran QM, Unkovic M, Wells M, Wickland J, Yackle K, Yavari A, Zaretsky JM, Allen CM, Alli L, An J, Anwar A, Arevalo S, Ayoub D, Badal SS, Baghdanian A, Baghdanian AH, Baumann SA, Becerra VN, Chan HJ, Chang AE, Cheng XA, Chin M, Chong F, Crisostomo C, Datta S, Delosreyes A, Diep F, Ekanayake P, Engeln M, Evers E, Farshidi F, Fischer K, Formanes AJ, Gong J, Gupta R, Haas BE, Hahm V, Hsieh M, Hui JZ, Iao ML, Jin SD, Kim AY, Kim LS, King M, Knudsen-Robbins C, Kohanchi D, Kovshilovskaya B, Ku A, Kung RW, Landig ME, Latterman SS, Lauw SS, Lee DS, Lee JS, Lei KC, Leung LL, Lerner R, Lin JY, Lin K, Lim BC, Lui CP, Liu TQ, Luong V, Makshanoff J, Mei AC, Meza M, Mikhaeil YA, Moarefi M, Nguyen LH, Pai SS, Pandya M, Patel AR, Picard PD, Safaee MM, Salame C, Sanchez C, Sanchez N, Seifert CC, Shah A, Shilgevorkyan OH, Singh I, Soma V, Song JJ, Srivastava N, StaAna JL, Sun C, Tan D, Teruya AS, Tikia R, Tran T, Travis EG, Trinh JD, Vo D, Walsh T, Wong RS, Wu K, Wu YW, Yang NX, Yeranosian M, Yu JS, Zhou JJ, Zhu RX, Abrams A, Abramson A, Amado L, Anderson J, Bashour K, Beyer E, Bookatz A, Brewer S, Buu N, Calvillo S, Cao J, Chan A, Chan J, Chang A, Chang D, Chang Y, Chen Y, Choi J, Chou J, Dang P, Davarifar A, Deravanesian A, Desai P, Fabrikant J, Farnad S, Fu K, Garcia E, Garrone N, Gasparyan S, Gayda P, Go S, Goffstein C, Gonzalez C, Guirguis M, Hassid R, Hermogeno B, Hong J, Hong A, Hovestreydt L, Hu C, Huff D, Jamshidian F, Jen J, Kahen K, Kao L, Kelley M, Kho T, Kim S, Kirkpatrick B, Langenbacher A, Laxamana S, Lee C, Lee SY, Lee TS, Lee T, Lewis G, Lezcano S, Lin P, Luu T, Luu J, Marrs W, Marsh E, Marshall J, Min S, Minasian T, Minye H, Misra A, Morimoto M, Moshfegh Y, Murray J, Nguyen K, Nguyen C, Nodado E, 2nd, O’Donahue A, Onugha N, Orjiakor N, Padhiar B, Paul E, Pavel-Dinu M, Pavlenko A, Paz E, Phaklides S, Pham L, Poulose P, Powell R, Pusic A, Ramola D, Regalia K, Ribbens M, Rifai B, Saakyan M, Saarikoski P, Segura M, Shadpour F, Shemmassian A, Singh R, Singh V, Skinner E, Solomin D, Soneji K, Spivey K, Stageberg E, Stavchanskiy M, Tekchandani L, Thai L, Thiyanaratnam J, Tong M, Toor A, Tovar S, Trangsrud K, Tsang WY, Uemura M, Vollmer E, Weiss E, Wood D, Wu J, Wu S, Wu W, Xu Q, Yamauchi Y, Yarosh W, Yee L, Yen G, Banerjee U. Genomewide clonal analysis of lethal mutations in the Drosophila melanogaster eye: comparison of the X chromosome and autosomes. Genetics. 2007;177:689–697. doi: 10.1534/genetics.107.077735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calleja M, Moreno E, Pelaz S, Morata G. Visualization of gene expression in living adult Drosophila. Science. 1996;274:252–255. doi: 10.1126/science.274.5285.252. [DOI] [PubMed] [Google Scholar]
- Curtiss J, Halder G, Mlodzik M. Selector and signalling molecules cooperate in organ patterning. Nat Cell Biol. 2002;4:E48–51. doi: 10.1038/ncb0302-e48. [DOI] [PubMed] [Google Scholar]
- Dahmann C, Oates AC, Brand M. Boundary formation and maintenance in tissue development. Nat Rev Genet. 2011;12:43–55. doi: 10.1038/nrg2902. [DOI] [PubMed] [Google Scholar]
- Duffy JB, Harrison DA, Perrimon N. Identifying loci required for follicular patterningusing directed mosaics. Development. 1998;125:2263–2271. doi: 10.1242/dev.125.12.2263. [DOI] [PubMed] [Google Scholar]
- Evans CJ, Olson JM, Ngo KT, Kim E, Lee NE, Kuoy E, Patananan AN, Sitz D, Tran P, Do MT, Yackle K, Cespedes A, Hartenstein V, Call GB, Banerjee U. G-TRACE: rapid Gal4-based cell lineage analysis in Drosophila. Nat Methods. 2009;6:603–605. doi: 10.1038/nmeth.1356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golic KG. Site-specific recombination between homologous chromosomes in Drosophila. Science. 1991;252:958–961. doi: 10.1126/science.2035025. [DOI] [PubMed] [Google Scholar]
- Golic KG, Lindquist S. The FLP recombinase of yeast catalyzes site-specific recombination in the Drosophila genome. Cell. 1989;59:499–509. doi: 10.1016/0092-8674(89)90033-0. [DOI] [PubMed] [Google Scholar]
- Hay BA, Wolff T, Rubin GM. Expression of baculovirus P35 prevents cell death in Drosophila. Development. 1994;120:2121–2129. doi: 10.1242/dev.120.8.2121. [DOI] [PubMed] [Google Scholar]
- Ito K, Awano W, Suzuki K, Hiromi Y, Yamamoto D. The Drosophila mushroom body is a quadruple structure of clonal units each of which contains a virtually identical set of neurones and glial cells. Development. 1997;124:761–771. doi: 10.1242/dev.124.4.761. [DOI] [PubMed] [Google Scholar]
- Janody F, Lee JD, Jahren N, Hazelett DJ, Benlali A, Miura GI, Draskovic I, Treisman JE. A mosaic genetic screen reveals distinct roles for trithorax and polycomb group genes in Drosophila eye development. Genetics. 2004;166:187–200. doi: 10.1534/genetics.166.1.187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lecuit T, Brook WJ, Ng M, Calleja M, Sun H, Cohen SM. Two distinct mechanisms for long-range patterning by Decapentaplegic in the Drosophila wing. Nature. 1996;381:387–393. doi: 10.1038/381387a0. [DOI] [PubMed] [Google Scholar]
- Lee T, Luo L. Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron. 1999;22:451–461. doi: 10.1016/s0896-6273(00)80701-1. [DOI] [PubMed] [Google Scholar]
- Lee T, Luo L. Mosaic analysis with a repressible cell marker (MARCM) for Drosophila neural development. Trends Neurosci. 2001;24:251–254. doi: 10.1016/s0166-2236(00)01791-4. [DOI] [PubMed] [Google Scholar]
- McGuire SE, Le PT, Osborn AJ, Matsumoto K, Davis RL. Spatiotemporal rescue of memory dysfunction in Drosophila. Science. 2003;302:1765–1768. doi: 10.1126/science.1089035. [DOI] [PubMed] [Google Scholar]
- McGuire SE, Roman G, Davis RL. Gene expression systems in Drosophila: a synthesis of time and space. Trends Genet. 2004;20:384–391. doi: 10.1016/j.tig.2004.06.012. [DOI] [PubMed] [Google Scholar]
- Morante J, Erclik T, Desplan C. Cell migration in Drosophila optic lobe neurons is controlled by eyeless/Pax6. Development. 2011;138:687–693. doi: 10.1242/dev.056069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newsome TP, Asling B, Dickson BJ. Analysis of Drosophila photoreceptor axon guidance in eye-specific mosaics. Development. 2000;127:851–860. doi: 10.1242/dev.127.4.851. [DOI] [PubMed] [Google Scholar]
- Pai CY, Kuo TS, Jaw TJ, Kurant E, Chen CT, Bessarab DA, Salzberg A, Sun YH. The Homothorax homeoprotein activates the nuclear localization of another homeoprotein, extradenticle, and suppresses eye development in Drosophila. Genes Dev. 1998;12:435–446. doi: 10.1101/gad.12.3.435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reuter JE, Nardine TM, Penton A, Billuart P, Scott EK, Usui T, Uemura T, Luo L. A mosaic genetic screen for genes necessary for Drosophila mushroom bodyneuronal morphogenesis. Development. 2003;130:1203–1213. doi: 10.1242/dev.00319. [DOI] [PubMed] [Google Scholar]
- Singh A, Choi KW. Initial state of the Drosophila eye before dorsoventral specification is equivalent to ventral. Development. 2003;130:6351–6360. doi: 10.1242/dev.00864. [DOI] [PubMed] [Google Scholar]
- Singh A, Kango-Singh M, Sun YH. Eye suppression, a novel function of teashirt, requires Wingless signaling. Development. 2002;129:4271–4280. doi: 10.1242/dev.129.18.4271. [DOI] [PubMed] [Google Scholar]
- Singh A, Shi X, Choi KW. Lobe and Serrate are required for cell survival during early eye development in Drosophila. Development. 2006;133:4771–4781. doi: 10.1242/dev.02686. [DOI] [PubMed] [Google Scholar]
- Singh A, Tare M, Puli OR, Kango-Singh M. A glimpse into dorso-ventral patterning of the Drosophila eye. Dev Dyn. 2012;241:69–84. doi: 10.1002/dvdy.22764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stowers RS, Schwarz TL. A genetic method for generating Drosophila eyes composed exclusively of mitotic clones of a single genotype. Genetics. 1999;152:1631–1639. doi: 10.1093/genetics/152.4.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu T, Rubin GM. Analysis of genetic mosaics in developing and adult Drosophila tissues. Development. 1993;117:1223–1237. doi: 10.1242/dev.117.4.1223. [DOI] [PubMed] [Google Scholar]