

Abstract
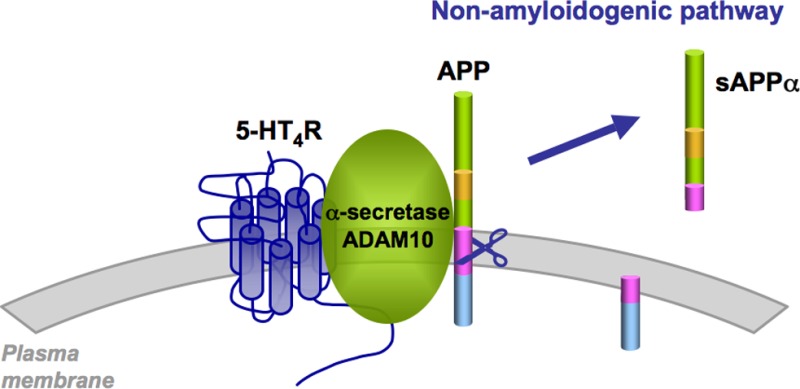
In addition to the amyloidogenic pathway, amyloid precursor protein (APP) can be cleaved by α-secretases, producing soluble and neuroprotective APP alpha (sAPPα) (nonamyloidogenic pathway) and thus preventing the generation of pathogenic amyloid-β. However, the mechanisms regulating APP cleavage by α-secretases remain poorly understood. Here, we showed that expression of serotonin type 4 receptors (5-HT4Rs) constitutively (without agonist stimulation) induced APP cleavage by the α-secretase ADAM10 and the release of neuroprotective sAPPα in HEK-293 cells and cortical neurons. This effect was independent of cAMP production. Interestingly, we demonstrated that 5-HT4 receptors physically interacted with the mature form of ADAM10. Stimulation of 5-HT4 receptors by an agonist further increased sAPPα secretion, and this effect was mediated by cAMP/Epac signaling. These findings describe a new mechanism whereby a GPCR constitutively stimulates the cleavage of APP by α-secretase and promotes the nonamyloidogenic pathway of APP processing.
Keywords: Alpha-secretase, Alzheimer’s disease, sAPP alpha, serotonin
According to the amyloid hypothesis, alteration in synaptic transmission and neuronal loss observed in Alzheimer’s disease (AD) mainly result from the formation of toxic amyloid-β (Aβ) oligomers followed by the extracellular accumulation of Aβ aggregates. Aβ is produced by the successive cleavage of a transmembrane amyloid precursor protein (APP) by β-secretase and γ-secretase.1 In addition to this amyloidogenic pathway, APP can be cleaved by α-secretases, a set of membrane-bound proteases of the ADAM (A disintegrin and metalloprotease) family, generating the soluble APP ectodomain (sAPPα) and a membrane-bound carboxy-terminal fragment (nonamyloidogenic pathway). As α-secretases cleave APP within the Aβ sequence, APP shedding by α-secretases prevents the generation of the pathogenic Aβ peptide.2,3 Therefore, enhancing α-secretase expression or activity has been considered as a valuable strategy for inhibiting Aβ formation. For instance, it has recently been shown that activation of the transcription of the gene encoding the α-secretase ADAM10, by retinoic acid and sirtuin 1 (SIRT1), reduces Aβ production.4−6
Previous reports have shown that G protein-coupled receptors (GPCRs) can differentially affect Aβ peptide production by either modulating the cellular trafficking of APP or by influencing the activity and trafficking of α-, β- and γ-secretases. Moreover, both the expression and the stimulation of GPCRs can affect APP metabolism.7 GPCRs that enhance sAPPα production by stimulating α-secretase activities include the muscarinic M1-M3 receptors, mGlu2 metabotropic glutamate receptor, serotonin 2A (5-HT2A) and 5-HT2C receptors, corticotropin-releasing factor (CRF) receptor 1, purinergic receptor P2X7 and pituitary adenylate cyclase-activating polypeptide (PACAP) type 1 (PAC1) receptor.7 These GPCRs are supposed to have a beneficial effect, because sAPPα exerts neuroprotective and neurotrophic effects and enhances long-term potentiation.8,9 In addition, promoting sAPPα formation generally decreases the production of toxic Aβ peptides. Several signaling cascades, such as the PKC, cAMP/PKA, MAP-Kinase, and PI3K pathways, have been involved in the stimulation of sAPPα release mediated by these GPCRs.10 However, the mechanisms by which they activate α-secretases are not always well understood.
The 5-HT4 receptor (5-HT4R) is another GPCR raising particular interest in view of its possible influence at different levels in the pathogenesis of AD. Activation of 5-HT4Rs stimulates acetylcholine release in prefrontal cortex and hippocampus and improves learning and memory in various preclinical paradigms of memory acquisition and retention.11,12 These findings suggest that 5-HT4R agonists might be used for improving cholinergic function and cognition, which are both compromised in AD. Stimulation of 5-HT4Rs also enhances sAPPα release and reduces Aβ formation in neuronal cultures.13,14 Whether the effect of 5-HT4Rs on APP metabolism reflects an increase in α-secretase activity or is the result of a modification of cellular trafficking of α-secretases and APP remains to be elucidated. Like many GPCRs, 5-HT4R interacts with a large set of intracellular proteins. Some of them, such as p11 and SNX27, are known to affect 5-HT4R transport and might also influence the trafficking of proteins physically and functionally connected with the receptor.15,16 Further supporting the implication of 5-HT4R in AD, the number of receptor binding sites is reduced in the hippocampus of patients with AD.17 However, the specific effects of 5-HT4R expression and activation on APP metabolism remain to be elucidated.
In this study, we show that 5-HT4R expression at physiological levels (around 120–550 fmol/mg of protein)17−21 enhances sAPPα release in transfected HEK-293 cells and cortical neurons in primary culture via two different processes, a “constitutive” release which has never been reported and a previously described agonist-regulated one.13,14 We also show that 5-HT4 receptors physically associate with ADAM10, a mechanism that may be involved in the constitutive release of sAPPα.
Results and Discussion
Expression of 5-HT4 Receptors Enhances sAPP Release from HEK-293 Cells and Cortical Neurons
To investigate the impact of 5-HT4R expression on APP metabolism, we first cotransfected HEK-293 cells with the SEAP-tagged APP plasmid (Figure 1A) and increasing amounts of the Myc-tagged 5-HT4R construct. 5-HT4R density at the cell surface was assessed by ELISA (Figure 1B) and [3H]GR 113808 saturation binding (from 120 to 750 fmol/mg protein corresponding to 25 to 500 ng cDNA/107 cells, respectively). Expression of increasing amounts of 5-HT4R did not significantly affect total APP (APP band intensity/actin band intensity = 97 ± 4% of control, 96 ± 7%, 108 ± 13%, or 89 ± 18% for 25, 100, 250, or 500 ng of 5-HT4R cDNA respectively, n ≥ 3 per each group) or ADAM10 expression (intensity of ADAM10 bands/actin band intensity = 104 ± 5% of control, 96 ± 11%, 113 ± 18%, or 87 ± 21% for 25, 100, 250, or 500 ng of 5-HT4R cDNA, respectively, n ≥ 3 per each group) or ADAM10 (Figure 1B), while it potently enhanced the release of soluble APP (sAPP) already at low receptor expression (Figure 1B). This effect was independent of the 5-HT4R variant used: the mouse (a, b, e, and f) and human (a, g, and i) C-terminal variants as well as further truncations of the 5-HT4R C-terminal domain (Δ358 and Δ329) could all promote sAPP release (Supporting Information Figure 1A–C). Overexpression of 5-HT4R in primary cultures of cortical neurons produced a comparable increase in sAPP release (Figure 1C) without affecting APP and ADAM10 levels (APP band intensity/actin band intensity = 88 ± 19% of control, intensity of ADAM10 bands/actin band intensity = 111 ± 14% for 2.500 ng of 5-HT4R cDNA n ≥ 3 per each group). Expression of similar amounts of PAC1 or muscarinic M3 receptor, which both increase sAPPα release upon activation by their respective agonists,22,23 failed to enhance sAPP release in HEK-293 cells in the absence of agonist (Figure 1D). Moreover, comparative levels of overexpressed 5-HT6 receptor, another serotonin receptor positively coupled to the Gs/cAMP pathway, did not induce sAPP production (Figure 1D). Similarly, only overexpression of 5-HT4R, but not of PAC1, M3, or 5-HT6 receptors, in cortical neurons enhanced the release of sAPP from neurons (Figure 1E).
Figure 1.

5-HT4R expression constitutively enhances sAPP release. (A) Schematic representation of APP. The position of the SEAP-tag is depicted. The epitopes of the anti-sAPP (22C11) and -sAPPα antibodies (7A6) and the β-, β′-, α-, and γ-secretase cleavage sites are also indicated. Mb, membrane. (B, C) HEK-293 cells (B) or embryonic cortical neurons (C) were transiently transfected with plasmids encoding Myc-tagged 5-HT4R and SEAP-tagged APP (500 ng/107 cells or 2500 ng/6 × 106 cells, respectively). Twenty-four h (B) or 4 days (C) post-transfection, cell surface expression of 5-HT4R was assessed by ELISA in nonpermeabilized cells using the anti-Myc antibody (B and C, white bars). In parallel, sAPP release during a 2 h period (B) or 12 h period (C) was evaluated by measuring the alkaline phosphatase activity (colorimetric assay) of cell supernatants (B and C, gray bars). (B, C) Total APP expression was detected in cell lysates in the corresponding transfection conditions, using the 22C11 antibody. Expression levels of ADAM10, 5-HT4R and actin were provided as controls. (D) HEK-293 cells were transiently transfected with plasmids encoding epitope-tagged 5-HT4R (300 ng/107 cells), PAC1R (700 ng/107 cells), M3R (300 ng/107 cells), or 5-HT6R (75 ng/107 cells) to ensure equivalent cell-surface expression of the different receptors, and SEAP-tagged APP (500 ng/107 cells). Cell surface expression of the receptors (white bars) and quantification of sAPP release (gray bars) were measured as described in (B) using appropriate primary antibodies. (E) Embryonic cortical neurons were transiently cotransfected with a plasmid encoding epitope-tagged 5-HT4R (800 ng/6 × 106 cells), PAC1R (1500 ng/6 × 106 cells), M3R (1000 ng/6 × 106 cells), or 5-HT6R (800 ng/107 cells) and SEAP-tagged APP (2500 ng/6 × 106 cells). Cell surface expression of the receptors (white bars) and quantification of sAPP release during a 4 day period was performed as described in (C). CTL, Condition transfected with only SEAP-tagged APP. Data are the means ± SD of at least three independent experiments. *p < 0.05, **p < 0.01 vs CTL corresponding value.
Expression of 5-HT4 Receptors Specifically Enhances sAPPα Release
To identify the nature of the sAPP form (sAPPα or sAPPβ) secreted upon 5-HT4R expression, we used a sandwich ELISA kit based on two antibodies, with one directed against the N-terminal sequence of APP and the other recognizing the neo-sAPPα epitope. Using this kit, we showed that overexpression of 5-HT4R in both HEK-239 cells and in cortical neurons led to an increase in the release of sAPPα (Figure 2A). Furthermore, SEAP-tagged APP in supernatants from transfected HEK-293 cells was immunoprecipitated with the 22C11 antibody and immunodetected using a site-specific antibody (7A6) that specifically recognizes sAPPα, but not shorter sAPP species, such as sAPPβ′ and sAPPβ (Figures 1A and 2B).24 The immunoreactive signals detected by 7A6 antibody increased concomitantly with 5-HT4R expression, while no significant change of APP and ADAM10 expression was observed (Figure 2B). Mutation of the residues (R609D and K612E) surrounding the α-secretase cleavage site (α-site APP mutant) can impair about 50% of the α-cleavage of APP.25 Coexpression of this mutant with 5-HT4R in HEK-293 cells strongly decreased 5-HT4R-induced sAPP release (Figure 2C), indicating that 5-HT4R expression stimulates the nonamyloidogenic α-cleavage of APP.
Figure 2.

5-HT4R specifically enhances sAPPα release. (A) HEK-293 cells were transiently transfected with plasmids encoding Myc-tagged 5-HT4R (250 ng/107 cells), SEAP-tagged APP (500 ng/107 cells). 2 h-sAPPα release in the medium was detected using an ELISA kit designed to quantify mouse/rat sAPPα. Cortical neurons were transfected with empty plasmid (CTL) or plasmid encoding Flag-tagged 5-HT4R (250 ng/6 × 106 cells). Four days after transfection, sAPPα released by neurons during 12 h was quantified using the same kit. **p < 0.01 vs corresponding CTL value. (B) Proteins in supernatants from HEK-293 cells transiently transfected with plasmids encoding Myc-tagged 5-HT4R and SEAP-tagged APP (500 ng/107 cells) were immunoprecipitated with the 22C11 antibody. Immunoprecipitated complexes were analyzed by Western blotting with the 7A6 (anti-sAPPα) antibody (upper panel). Expression levels of APP, ADAM10, 5-HT4R, and actin in cell lysates were provided as controls (lower panel). (C) HEK-293 cells were transiently cotransfected with plasmids encoding Myc-tagged 5-HT4R (250 ng/107 cells) and either wild type SEAP-tagged APP or the α site APP mutant (R609D, K612E) (500 ng/107 cells). sAPP release during 2 h was evaluated by measuring the alkaline phosphatase activity in the cell medium, **p < 0.01 vs corresponding value in cells expressing the wild type SEAP-tagged APP. (D) HEK-293 cells were transiently transfected with plasmids encoding Myc-tagged 5-HT4R (250 ng/107 cells) and SEAP-tagged APP (500 ng/107 cells). 24 h post-transfection, cells were treated with vehicle alone or 50 μM TAPI-1 and SEAP activity during 30, 60, and 120 min was measured in the spent medium. *p < 0.05, **p < 0.01 vs corresponding value in absence of TAPI-1. (E, F) HEK-293 cells were transiently transfected with plasmids encoding Myc-tagged 5-HT4R (250 ng/107 cells), SEAP-tagged APP (500 ng/107 cells), and either control (CTL) or ADAM10 siRNA. ADAM10 expression was assessed by Western blotting (E) and sAPPα release (F) as described in (A), **p < 0.01 vs 5-HT4R-mediated release in cells transfected with control siRNA. CTL, Condition transfected with only SEAP-tagged APP. Data are the means ± SD of at least three independent experiments.
Several metalloproteinases, including ADAM9, ADAM10, and ADAM17 (also called TACE, tumour necrosis factor α-converting enzyme) have been proposed as potential α-secretases that promote the shedding of APP ectodomain. TAPI-1, a metalloproteinase inhibitor, almost completely abolished 5-HT4R-induced sAPPα release from HEK-293 cells (Figure 2D), suggesting the involvement of a member of the ADAM family of metalloproteinases. Moreover, reduction of ADAM10 expression by transfection of ADAM10-specific siRNA (Figure 2E) diminished basal processing of APP in absence of 5-HT4 receptor and also markedly decreased sAPPα release following 5-HT4R expression (Figure 2F). This indicates that ADAM10 contributes significantly to 5-HT4R-induced sAPP release. In contrast, specific inhibitors of β-secretase (KTEEISEVN-statine-VAEF) or γ-secretase (L-685,458) did not affect the release of sAPPα induced by 5-HT4R (Supporting Information Figure 2).
5-HT4R Stimulation Further Enhances sAPPα Release
Previous studies have shown that 5-HT4R stimulation by agonists increases sAPPα production.26 Consistent with these findings, treating HEK-293 cells that transiently express 5-HT4R and SEAP-tagged APP with prucalopride, a 5-HT4R agonist, for 2 h increased by ∼50% the release of sAPPα induced by 5-HT4R expression (Figure 3A and B). This effect was blocked by GR 113808 and RO 116-0086, a neutral antagonist and an inverse agonist of 5-HT4R at Gs signaling, respectively. Neither GR 113808 nor RO 116-0086 alone altered the secretion of sAPP in the absence of the agonist (Figure 3B).
Figure 3.

5-HT4R stimulation further enhances sAPPα release induced by 5-HT4R expression. (A, B) HEK-293 cells were transiently transfected with plasmids encoding Myc-tagged 5-HT4R (250 ng/107 cells) and SEAP-tagged APP (500 ng/107 cells). Twenty-four h post-transfection, cells were treated with vehicle alone (CTL) or 1 μM prucalopride with or without 5 μM GR 113803 or 0.1 μM RO 116-0086 for 2 h, and then sAPP release was evaluated by measuring the alkaline phosphatase activity in the medium. (C, D) Embryonic cortical neurons were transiently transfected with plasmids encoding Flag-tagged 5-HT4R (800 ng/6 × 106 cells) with (C) or without (D) SEAP-tagged APP (2500 ng/6 × 106 cells). Four days after transfection, neurons were treated with increasing concentrations of prucalopride and sAPP release during 12 h was evaluated by measuring the alkaline phosphate activity (C) or by using the sAPPα ELISA kit (D). (E, F) Cortical neurons cotransfected with plasmids encoding Flag-tagged 5-HT4R (250 ng/6 × 106 cells) and SEAP-tagged APP (2500 ng/6 × 106 cells) were exposed to the indicated 5-HT4R agonists/antagonists for 12 h and sAPP release was evaluated by measuring the alkaline phosphatase activity. CTL, Condition transfected with only SEAP-tagged APP. Data are the means ± SD of at least three independent experiments. In (B), (E), and (F), *p < 0.05 vs the corresponding value in cells not treated with agonist.
5-HT4R stimulation by prucalopride increased sAPPα release by cultured cortical neurons that transiently expressed 5-HT4R and SEAP-tagged APP in a concentration-dependent manner (pEC50 = 0.85 ± 0.22 μM, n = 3), as assessed by monitoring the SEAP activity (Figure 3C). Similarly, prucalopride stimulated sAPPα release from endogenous APP, as assessed using a sAPPα specific ELISA kit (Figure 3D). Like in HEK-293 cells, the prucalopride effect was blocked by GR 113808 and RO 116-0086 (Figure 3E). Treatment of neurons with maximally effective concentrations of various synthetic 5-HT4R agonists (10 μM RS 67333, 3 μM prucalopride, 10 μM BIMU8, 1 μM cisapride)27 or with 10 μM 5-HT stimulated sAPP secretion to a similar extent as prucalopride. In contrast, 5-HT4R ligands exhibiting either antagonist (GR 113808, ML 10375) or inverse agonist (RO 116-1148, SB 207266) properties on 5-HT4 receptors coupled to Gs27 did not significantly affect sAPP release from neurons (Figure 3F). However, RO 116-0086 and RO 116-2617, two inverse agonists at Gs signaling decreased basal sAPP release. GPCRs proteins are adapting many conformations. The constitutive 5-HT4R conformation which induces cAMP production (via G protein) is unlikely to be similar to the constitutive conformation of the same 5-HT4 receptor which is producing sAPPα (without a need of G protein). The pharmacology of these two different constitutive conformations is likely to be different. Thus, it is not surprising that some “classical” inverse agonists on the 5-HT4 receptor inducing constitutive cAMP production are not inverse agonists on the constitutive conformation required for sAPPα production.
The Constitutive sAPPα Release Induced by 5-HT4R Expression Is Independent of the cAMP/Epac Pathway
As cAMP and Epac (cAMP-responsive Rap1 guanine nucleotide exchange factor) regulate sAPPα release elicited by 5-HT4R stimulation,13 we investigated whether receptor coupling to Gs and recruitment of the cAMP/Epac pathway were also necessary for enhancing constitutive sAPPα secretion upon 5-HT4R expression. Expression of a dominant-negative form of Gs strongly reduced agonist-independent cAMP accumulation in HEK-293 cells expressing 5-HT4Rs, but did not affect sAPP release (Figure 4A). Moreover, expression of 5-HT4R mutants with high level of constitutive Gs signaling activity (Y302F, N308D, or A258L)28,29 increased basal cAMP production but were less active to increase sAPP release in comparison to wild type 5-HT4R (Figure 4B). Similarly, overexpression of a dominant negative form of Epac1 did not modify the constitutive sAPP secretion induced by 5-HT4R expression (Figure 4C), whereas the agonist (prucalopride)-dependent sAPP release was reduced, as previously reported13 (Figure 4C). As expected, the Epac1 dominant negative form did not affect cAMP production elicited by 5-HT4R activation (Figure 4D). Collectively, these results indicate that only agonist-elicited sAPP secretion is dependent on 5-HT4R coupling to Gs protein and engagement of the cAMP/Epac pathway.
Figure 4.

Role of the Gs/cAMP/Epac pathway in 5-HT4R-induced sAPPα release. HEK-293 cells were transiently transfected with plasmids encoding Myc-tagged 5-HT4R (250 ng/107 cells), SEAP-tagged APP (500 ng/107 cells), and a dominant negative Gs protein (A) or a dominant-negative form of Epac1 (500 ng/107 cells) (C, D). (B) HEK-293 cells were transiently transfected with plasmids encoding Myc-tagged wild type 5-HT4R or mutants (Y302F, N308D, or A258L) with high level of constitutive Gs signaling activity and SEAP-tagged APP (500 ng/107 cells each). Twenty-four h after transfection, cAMP accumulation and sAPP release during 2 h were measured. CTL, Condition transfected with only SEAP-tagged APP. Data are the means ± SD of at least three independent experiments. In (D) and (F), *p < 0.05 vs the corresponding value in cells not treated with agonist.
5-HT4 Receptors Physically Interact with the α-Secretase ADAM10
To further explore the possible mechanism by which 5-HT4R constitutively activates ADAM10-dependent APP cleavage, we examined whether 5-HT4R physically interacts with this α-secretase. ADAM10 coimmunoprecipited with HA-tagged-5-HT4R in HEK-293 cells (Figure 5A). Although two immunoreactive ADAM10 species with apparent molecular masses of 95 (immature pro-enzyme) and 69 kDa (mature, active form) were detected in whole cell lysates by Western blotting, only the 69 kDa mature form, which originates from the immature form by removal of a 194 amino acid pro-domain and is localized at the plasma membrane,30 was coimmunoprecipitated with 5-HT4R. Prucalopride stimulation of 5-HT4R did not modify the ability of the receptor to interact with ADAM10, thus demonstrating the existence of a “constitutive” specific complex composed of 5-HT4R and active ADAM10 (Figure 5A).
Figure 5.

5-HT4R interacts with the α-secretase ADAM10. (A) HEK-293 cells were transiently transfected with plasmids encoding HA-tagged 5-HT4R (250 ng/107 cells) and SEAP-tagged APP (500 ng/107 cells). Twenty-four h after transfection, cells were treated 2 h with vehicle alone or with 1 μM prucalopride (Prucalo), and then protein lysates were immunoprecipitated with the anti-HA antibody. Whole cell extracts and immunoprecipitated material were analyzed by Western blotting using antibodies against ADAM10 (mature (m) and immature (im) forms) and against HA-tag. (B–D) HEK-293 cells were transiently cotransfected with plasmids encoding Myc- (C) or HA-tagged 5-HT4R (B, D), HA-5-HT6R (B), Myc-PAC1R (C), or HA-M3R (D) and SEAP-tagged APP. Twenty-four h after transfection, protein lysates were immunoprecipitated as described in (A). (E) Data correspond to the ratio of ADAM10 coimmunoprecipitation to 5-HT4R immunoprecipitation signals. ND, not detectable. CTL, Condition transfected with only SEAP-tagged APP. The data illustrated are representative of three independent experiments performed with different sets of cultured cells.
Active ADAM10 did not coimmunoprecipitate with overexpressed 5-HT6 (Figure 5B), PAC1 (Figure 5C), or M3 receptors (Figure 5D). Furthermore, different 5-HT4R mutants (A258L, N308D, and Y302F) and truncated forms devoid of the C-terminal domain (Δ327 and Δ329) showed the same ability to interact with ADAM10 as the wild type receptor (Supporting Information Figure 1D–G).
Promotion of Non-Amyloidogenic Pathway by 5-HT4 Receptors
The nature of APP processing is dependent on its cellular localization and on the cellular proteases involved in its cleavage. The present study shows that 5-HT4R expression, at physiological levels (120–550 fmol/mg proteins),17−21 constitutively promotes the nonamyloidogenic processing of APP and that 5-HT4 receptors physically interact with the mature form of the α-secretase ADAM10. Several reports described the overlapping expression of APP and ADAM10 (but not ADAM17) in adult brain neurons31 and suggested that ADAM10 could be the α-secretase involved in the nonamyloidogenic pathway of APP, particularly in neurons.30 This assumption was confirmed by RNAi-mediated silencing of ADAM10 and by using conditional ADAM10 knockout mice.24,32 These findings were extended by the present study, which revealed association of the 5-HT4R with ADAM10. The ability of 5-HT4R to physically interact with mature ADAM10 was not shared by other GPCRs known to stimulate sAPPα secretion upon stimulation by their respective agonists. Indeed, expression of the PAC1 and M3 receptors did not trigger the formation of such a protein complex. Moreover, ADAM10 is retained in the ER by an arginine stretch (RRR).33 Future work should test the possibility that association of ADAM10 with 5-HT4R masks this sequence, allowing the translocation of both proteins to the plasma membrane, a process associated with ADAM10 maturation.
It has been previously reported that stimulation of 5-HT4R promotes sAPPα release in a cAMP/Epac-dependent manner,13 and these findings were confirmed in the present study. Although 5-HT4R displays high level of constitutive activity at the Gs-adenylyl cyclase pathway,28 several lines of evidence argued against a role of the cAMP/Epac pathway in the constitutive effect of 5-HT4R on APP processing: (1) inhibition of cAMP formation by an inverse agonist or by overexpression of the inactive form of Gs had no effect on the level of sAPPα release elicited by 5-HT4R expression; (2) expression of a dominant negative Epac mutant did not affect constitutive induction of sAPPα release by 5-HT4R; and (3) mutants of 5-HT4R that constitutively increase adenylyl cyclase activity did not stimulate sAPPα secretion more than the wild type receptor. Collectively, these findings demonstrate that 5-HT4R promotes the nonamyloidogenic processing of APP via different mechanisms implicating either the canonical Gs protein-dependent signaling (agonist-elicited response) or the formation of a protein complex including the receptor, the ADAM10 α-secretase, and, probably, additional protein partners that remain to be identified (agonist-independent response).
One promising strategy to decrease the accumulation of Aβ peptides in the brain has been to inhibit β- and γ-secretase activities. However, the development of γ-secretase inhibitors has been challenging because this enzyme has many endogenous substrates. Several γ-secretase inhibitors have been synthesized and have shown unequivocal efficiency in reducing Aβ plaque deposition in preclinical models of AD. However, clinical trials have revealed severe adverse side effects associated with the blockade of Notch processing.34 The recent failure of a γ-secretase inhibitor to slow down the progression of the disease, accompanied by worsening of cognitive performances, has halted its development.35 However, the recent discovery that down-regulation of GSAP (γ-secretase activating protein) selectively prevents Aβ peptide production without affecting Notch signaling36 opens new perspectives.
Strategies for increasing the production of the neuroprotective sAPPα are also actively investigated. 5-HT4R expression clearly increased sAPPα release, and it was previously demonstrated that 5-HT4R activation concomitantly decreased Aβ formation while increasing sAPPα release.13,14 However, α-cleavage is not always coupled to a reduction in Aβ level.3 The role of ADAM10, the physiological α-secretase in neurons, in the regulation of late-onset AD has been suggested by the discovery of nonsynonymous mutations in the ADAM10 pro-domain in seven late-onset AD families.37 In line with these findings, activation of ADAM10 expression by vitamin A derivatives4 or the transcription factor SIRT16 decreases Aβ peptide production and amyloid plaque formation in APP/PSEN1 transgenic mice. In addition, SIRT1-induced ADAM10 expression increases Notch processing and the release of Notch/intracellular domain (NICD), which activates genes involved in neuronal repair in adult brain.38 The efficacy of ligands of some GPCRs, such as M1R, 5-HT4R, and PAC1R, in reducing Aβ production and/or improving cognition in preclinical models of AD is now well documented.39 Here, we describe a new mechanism whereby 5-HT4Rs constitutively activate the α-secretase ADAM10 and sAPPα release. Together with the ability of 5-HT4R agonists to increase learning and memory, via acetylcholine release,12 the present findings suggest that this GPCR might represent an interesting target for AD treatment. Several studies reported a significant drop of 5-HT4R binding sites in post-mortem brains of AD patients.17,40,41 In this context, the identification of 5-HT4R-interacting proteins capable of modulating the trafficking of the 5-HT4R/ADAM10 complex will be of high relevance.
Methods
Plasmids
The plasmid encoding the inactive form of the Gs protein was kindly given by Dr. Catherine H. Berlot (Weis Center for Research Geisinger Clinic, Danville, Canada). The plasmids encoding c-Myc-tagged 5-HT4R, HA-tagged 5-HT4R, Flag-tagged 5-HT4R, HA-tagged 5-HT6R, and the dominant-negative form of Epac2 were previously described.28,42,43 The SEAP-tagged APP695 cDNA in pRK5 was generated by adding the secreted embryonic alkaline phosphatase (SEAP) sequence, flanked by glycine linkers and the PspX I (5′) or BsrG I (3′) restriction sites, to the N-terminus of mouse APP695, after L398 (Figure 1A). The plasmids encoding the HA-tagged M3 and Myc-tagged PAC1 receptors were kindly provided by Dr. Arnaud Monteil and Dr. Laurent Journot, respectively (Institut de Génomique Fonctionnelle, Montpellier, France).
Antibodies
The mouse anti-sAPP antibody (22C11, directed against the N-terminal part of APP; 1:80 dilution)44 was purchased from Millipore (Billerica, MA). The rat anti-sAPPα antibody (7A6, murine sAPPα-specific, 1:2 dilution) was kindly given by Dr. Stefan F. Lichtenthaler (Adolf Butenandt Institute - Ludwig Maximilians University, Munich, Germany).24 The rabbit anti-ADAM10 antibody (directed against the C-terminal domain, 1:500 dilution), the mouse anti-c-Myc antibody (9E10), and the rabbit anti-Flag antibody were purchased from Sigma-Aldrich (Saint Quentin Fallavier, France). The mouse anti-HA antibody was purchased from Life Technologies (Saint Aubin, France). The rat anti-ADAM10 antibody (directed against the ectodomain domain, 1:500 dilution) was purchased from R&D Systems (Minneapolis, MN). Mouse anti-HA and rabbit anti-GFP antibodies were purchased from Life Technologies-Invitrogen (Carlsbad, CA). The mouse anti-SEAP antibody was purchased from Euromedex (Souffelweyershein, France). Horseradish peroxidase (HRP)-conjugated anti-rabbit and anti-mouse antibodies were from GE Healthcare (Orsay, France).
Materials
All media, sera, and antibiotics used for cell culture were purchased from Lonza (Basel, Switzerland). GR 113808 and GR 125487 were synthesized and generously provided by GlaxoSmithKline (Brentford, U.K.). 5-HT (serotonin creatine sulfate), Phorbol 12-myristate 13-acetate (PMA), the β-secretase inhibitor KTEEISEVN-statine-VAEF, the γ-secretase inhibitor L-685458, (−)-isoproterenol hydrochloride, and metoclopramide ((N-(2-dimethylamino)-ethyl)-4-amino-5-chloro-2-methoxybenzamide) were from Sigma. The TNF-α Protease Inhibitor-1 (TAPI-1) was purchased from Merck (Darmstadt, Germany). Prucalopride was synthesized and kindly provided by Dr. Jan Schuurkes (Shire-Movetis n.v., Turnhout, Belgium). RO 116-0086, RO 116-1148, and RO 116-2617 were generously provided by Roche Bioscience (Palo Alto, CA). RS 67333 was purchased from Tocris Bioscience (Bristol, U.K.). BIMU8 was kindly provided by Boehringer Ingelheim (Rhein, Germany). Cisapride was synthesized and obtained from Janssen Pharmaceutica (Beerse, Belgium). ML 10375 was a generous gift from Dr. Langlois (CNRS-BIOCIS, Châtenay-Malabry, France). SB 207266 was obtained from Fournier-Debat Laboratories (Daix, France).
Cell Culture and Transfections
Human embryonic kidney 293 (HEK-293) cells were grown in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% dialyzed fetal calf serum (FCS). Cells were transfected at 60–70% confluence by electroporation, as previously described.28 Twenty-four hours after transfection, cells were serum starved for 4 h before assays.
Primary cultures of cortical neurons were prepared as previously described.45 Briefly, cells dissociated from the cortex of 15–16 day/old Swiss mouse embryos were plated in serum-free medium in 24-well culture dishes (0.5 × 106 cells/well). Cultures were maintained at 37 °C in a humidified atmosphere in 5% CO2 and 95% H2O, in DMEM/F12 supplemented with 10% hormone mixture for 4 days. Cortical neurons were transfected by electroporation using AMAXA (Lonza, Basel, Switzerland) according to the manufacturer’s instructions. Media were collected for 12 h at DIV4.
Small Interfering RNA (siRNA) Transfection
A mix of double-stranded siRNA sequences 5′-AAUGAAGAGGGACACUUCCCUdTdT-3′ and 5′-AAGUUGCCUCCUCCUAAACCAdTdT-3′ (Eurogentec, Liege, Belgium) was used to selectively target ADAM10. A small RNA duplex, which has no silencing effect, was used as control (negative control siRNA duplex from Eurogentec). All the siRNAs used in this study have been described earlier and validated.46 Early passage HEK-293 cells at 40% confluence in 24-well plates were transiently transfected with 400 ng of plasmid encoding wild type 5-HT4R and 500 ng of plasmid encoding SEAP-tagged-APP. siRNAs were transfected at 150 nM using Lipofectamine 2000, according to the manufacturer’s recommendations (Life Technologies-Invitrogen). Cells were lysed 2 days after siRNA transfection in order to quantify APP expression.
Measurement of sAPP
To determine the total and intracellular concentration of APP and of secreted APP, cells were washed with PBS at the beginning of the assay and preincubated for 2 h in serum-free medium supplemented with 5-HT4R ligands and drugs were added for the indicated times. Cells were then lysed after collection of conditioned supernatants containing secreted proteins according to the two following methods: (i) Collected media were centrifuged at 300g for 10 min and then at 20 000g for 20 min to remove cells and cell debris. Proteins were concentrated by precipitation with 10% ice-cold tricholroacetic acid (TCA) (at 4 °C for 2 h) and centrifuged at 38 000g at 4 °C for 25 min, and then pellets were washed with diethyl ether three times. TCA precipitates were reconstituted in solubilization buffer (20 mM HEPES buffer; 250 mM Tris buffer pH6.8; 40% SDS; 40% glycerol; 40% 2-mercaptoethanol and traces of bromophenol blue). The protein amount of each sample was determined with the bicinchoninic acid assay (Sigma). Following precipitation, samples were separated by electrophoresis on 7.5% SDS-PAGE gels and transferred onto nitrocellulose membranes. (ii) In order to measure sAPP release, HEK-293 cells were transiently transfected with APP-SEAP and plated in 6-well culture dishes. Spent media were incubated overnight at 4 °C with Dynabeads protein G (Life Technologies-Invitrogen, Carlsbad, CA) that had been precoupled to 4 μg of 22C11 polyclonal antibody. After 5 washes in lysis buffer (20 mM HEPES; 150 mM NaCl; 1% NP40; 10% glycerol; 4 mg/mL dodecylmaltoside) supplemented with protease inhibitors and phosphatase inhibitors (10 mM NaF; 2 mM Na+-vanadate and 1 mM Na+-pyrophosphate), immunoprecipitated proteins were eluted in solubilization buffer and separated by electrophoresis (see above). To detect intracellular APP, cells were lysed in 100 μL of lysis buffer (see above), reconstituted in solubilization buffer, and separated by electrophoresis (see above).
sAPP release was also estimated by measuring the extracellular SEAP activity released by HEK-293 cells transiently transfected with the SEAP-APP plasmid. One day post-transfection, drugs were added for the indicated times (see legends). Then cell culture media were collected, and SEAP activity measured by adding the chromogenic substrate p-nitrophenyl phosphate disodium hexahydrate (Sigma) according to the manufacturer’s instructions. The reaction was read at 405 nm using an Infinite 2000 luminescence counter (Tecan, Männedorf, Switzerland). Specific quantification of sAPPα release was performed by using the sAPPα ELISA kit from IBL International (Hamburg, Germany) according to the manufacturer’s instructions. The reaction was read at 620 and 450 nm using an Infinite 2000 luminescence counter (Tecan).
Immunoblotting
Proteins were separated on 7.5% SDS-PAGE gels and transferred by semidry blotting onto nitrocellulose membranes (Hybond-C, Amersham Biosciences, Orsay, France). Membranes were blocked at room temperature in Tris-buffered saline blocking solution containing 5% (w/v) milk powder, 0.25% v/v Tween 20 for 1 h. Membranes were then incubated with the primary antibodies at 4 °C overnight, washed extensively, and incubated with the secondary antibodies at room temperature for 1 h. Immunoreactivity was detected using the Chemiluminescence Reagent Plus kit (Perkin-Elmer, Courtaboeuf, France). Immunoreactive bands were quantified by densitometry using Image J and GraphPad PRISM (GraphPad Software, San Diego, CA).
Co-Immunoprecipitation
Cells were seeded in 150 mm plates (106 cells/plate) 24 h before the experiment. Cross-linking was carried out in PBS completed with 1.25 mM dithiobissuccinimidyl propionate (DSP) (Pierce, Perbio-Brebières, France) (a membrane-permeable, hydrolyzable, covalent cross-linker) for 30 min. The cross-linking reaction was stopped with 50 mM Tris pH 7.4. After one wash in PBS, cells were incubated with lysis buffer (see above) at 4 °C for 1 h. Samples were centrifuged at 20 000g for 15 min. Solubilized proteins were incubated at 4 °C with 20 μL of Dynabeads protein G (Life Technologies-Invitrogen) precoupled with 4 μg of antibody overnight. Immunoprecipitated proteins were eluted in Laemmli sample buffer, resolved by SDS-PAGE gel electrophoresis, and detected by Western blotting.
Determination of cAMP Production
HEK-293 cells were transiently transfected with plasmids encoding the indicated receptors and/or SEAP-APP, and then seeded in 24-well plates (200 000 cells/well). Twenty-four hours after transfection, cells were exposed to the indicated concentrations of 5-HT4R ligands in the presence of 0.1 mM l-ascorbic acid and 0.1 mM of the phosphodiesterase inhibitor RO-20-1724, at 37 °C in 250 μL of HBS (20 mM HEPES; 150 mM NaCl; 4.2 mM KCl; 0.9 mM CaCl2; 0.5 mM MgCl2; 0.1% glucose; 0.1% BSA). Cells were then lysed by addition of the same volume of Triton-X100 (0.1%). Quantification of cAMP production was performed by HTRF by using the cAMP Dynamic kit (Cisbio Bioassays) according to the manufacturer’s instructions.
Cell Surface ELISA (Enzyme-Linked Immunosorbent Assay)
HEK-293 cells transiently transfected with the appropriate cDNAs were seeded in 96-well plates (40 000 cells/well). Twenty-four hours after transfection, cells were treated with drugs, washed, fixed with 4% paraformaldehyde at room temperature for 10 min, and blocked with PBS + 1% FCS. Cells were then incubated with primary antibodies in the same buffer for 1 h followed by HRP conjugated secondary antibodies at 1:8000 for 1 h. After extensive washes, the chromogenic substrate was added (Supersignal ELISA Femto Maximum Sensitivity, Pierce). Chemiluminescence was detected and quantified with an Infinite 2000 luminescence counter (Tecan).
Statistical Analysis
All represented experiments were performed in triplicate. Statistical significance was determined with the Student–Newman–Keuls test using GraphPad Prism (GraphPad Software). P-values < 0.05 were considered as statistically significant. Data corresponded to the mean ± SD of three independent experiments.
Acknowledgments
We are grateful to Gaël Barthet, Federica Bertaso, and Vincent Homburger for constructive discussion. Binding experiments, cAMP measurements, and ELISA were carried out using the ARPEGE Pharmacology Screening Interactome facility at the Institute of Functional Genomics (Montpellier, France).
Glossary
Abbreviations
- APP
amyloid precursor protein
- sAPPα
soluble APP alpha
- sAPPβ
soluble APP beta
- SEAP
secreted embryonic alkaline phosphatase
- 5-HT4R
serotonin type 4 receptor
- AD
Alzheimer’s disease
- ADAM
A disintegrin and metalloprotease
- Aβ
amyloid-β peptide
- GPCR
G protein-coupled receptor
Supporting Information Available
Supplemental Figure 1: 5-HT4R variants (mouse a, b, e, and f; human a, g, and i), truncated forms (Δ358 and Δ329) and mutants (A258L, N308D, and Y302F) promote sAPP release and physically interact with ADAM10. Supplemental Figure 2: sAPP release induced by 5-HT4R expression is not affected by β- or γ-secretase inhibitors. Supplemental figure legends. This material is available free of charge via the Internet at http://pubs.acs.org.
Author Contributions
⊥ These authors contributed equally to this work.
Author Contributions
M.C., J.B., A.D., and S.C. conceived and designed the experiments. M.C., R.D., E.C., and F.G. carried out the experiments. M.C., E.C., F.G., S.F.L., P.M., J.B., A.D., and S.C. analyzed the data. M.C., P.M., J.B., A.D., and S.C. wrote the manuscript.
Supported by grants from CNRS, INSERM, the French Minister of Research (ANR Blanc 2006, to AD and SC, ANR Blanc 2012 “ADAMGUARD” to J.B.), the “Fondation pour la Recherche Médicale” (FRM, to P.M.), the France Alzheimer association (to S.C.), the Soroptimist International (French Union, to S.C.), the Languedoc Roussillon region (“Chercheuse d’Avenir” 2012, to S.C.) and the competence network degenerative dementias (BMBF-KNDD, to S.F.L.). M.C. was a recipient of fellowships from the CNRS, the Languedoc Roussillon region and FRM. E.C. was supported by ANR Blanc 2006.
The authors declare no competing financial interest.
Supplementary Material
References
- Selkoe D. J. (2001) Alzheimer’s disease: genes, proteins, and therapy. Physiol. Rev. 81, 741–766. [DOI] [PubMed] [Google Scholar]
- Fahrenholz F. (2007) Alpha-secretase as a therapeutic target. Curr. Alzheimer Res. 4, 412–417. [DOI] [PubMed] [Google Scholar]
- Lichtenthaler S. F. (2010) Alpha-secretase in Alzheimer’s disease: molecular identity, regulation and therapeutic potential. J. Neurochem. 116, 10–21. [DOI] [PubMed] [Google Scholar]
- Tippmann F.; Hundt J.; Schneider A.; Endres K.; Fahrenholz F. (2009) Up-regulation of the alpha-secretase ADAM10 by retinoic acid receptors and acitretin. FASEB J. 23, 1643–1654. [DOI] [PubMed] [Google Scholar]
- Jarvis C. I.; Goncalves M. B.; Clarke E.; Dogruel M.; Kalindjian S. B.; Thomas S. A.; Maden M.; Corcoran J. P. (2010) Retinoic acid receptor-alpha signalling antagonizes both intracellular and extracellular amyloid-beta production and prevents neuronal cell death caused by amyloid-beta. Eur. J. Neurosci. 32, 1246–1255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donmez G.; Wang D.; Cohen D. E.; Guarente L. (2010) SIRT1 suppresses beta-amyloid production by activating the alpha-secretase gene ADAM10. Cell 142, 320–332. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Thathiah A.; De Strooper B. (2011) The role of G protein-coupled receptors in the pathology of Alzheimer’s disease. Nat. Rev. Neurosci. 12, 73–87. [DOI] [PubMed] [Google Scholar]
- Furukawa K.; Sopher B. L.; Rydel R. E.; Begley J. G.; Pham D. G.; Martin G. M.; Fox M.; Mattson M. P. (1996) Increased activity-regulating and neuroprotective efficacy of alpha-secretase-derived secreted amyloid precursor protein conferred by a C-terminal heparin-binding domain. J. Neurochem. 67, 1882–1896. [DOI] [PubMed] [Google Scholar]
- Ishida A.; Furukawa K.; Keller J. N.; Mattson M. P. (1997) Secreted form of beta-amyloid precursor protein shifts the frequency dependency for induction of LTD, and enhances LTP in hippocampal slices. NeuroReport 8, 2133–2137. [DOI] [PubMed] [Google Scholar]
- Thathiah A.; De Strooper B. (2009) G protein-coupled receptors, cholinergic dysfunction, and Abeta toxicity in Alzheimer’s disease. Sci. Signaling 2, re8. [DOI] [PubMed] [Google Scholar]
- Cachard-Chastel M.; Devers S.; Sicsic S.; Langlois M.; Lezoualc’h F.; Gardier A. M.; Belzung C. (2008) Prucalopride and donepezil act synergistically to reverse scopolamine-induced memory deficit in C57Bl/6j mice. Behav. Brain Res. 187, 455–461. [DOI] [PubMed] [Google Scholar]
- Bockaert J.; Claeysen S.; Compan V.; Dumuis A. (2011) 5-HT(4) receptors, a place in the sun: act two. Curr. Opin. Pharmacol. 11, 87–93. [DOI] [PubMed] [Google Scholar]
- Maillet M.; Robert S. J.; Cacquevel M.; Gastineau M.; Vivien D.; Bertoglio J.; Zugaza J. L.; Fischmeister R.; Lezoualc’h F. (2003) Crosstalk between Rap1 and Rac regulates secretion of sAPPalpha. Nat. Cell Biol. 5, 633–639. [DOI] [PubMed] [Google Scholar]
- Cho S.; Hu Y. (2007) Activation of 5-HT4 receptors inhibits secretion of beta-amyloid peptides and increases neuronal survival. Exp. Neurol. 203, 274–278. [DOI] [PubMed] [Google Scholar]
- Warner-Schmidt J. L.; Flajolet M.; Maller A.; Chen E. Y.; Qi H.; Svenningsson P.; Greengard P. (2009) Role of p11 in cellular and behavioral effects of 5-HT4 receptor stimulation. J. Neurosci. 29, 1937–1946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joubert L.; Hanson B.; Barthet G.; Sebben M.; Claeysen S.; Hong W.; Marin P.; Dumuis A.; Bockaert J. (2004) New sorting nexin (SNX27) and NHERF specifically interact with the 5-HT4a receptor splice variant: roles in receptor targeting. J. Cell Sci. 117, 5367–5379. [DOI] [PubMed] [Google Scholar]
- Reynolds G. P.; Mason S. L.; Meldrum A.; De Keczer S.; Parnes H.; Eglen R. M.; Wong E. H. (1995) 5-Hydroxytryptamine (5-HT)4 receptors in post mortem human brain tissue: distribution, pharmacology and effects of neurodegenerative diseases. Br. J. Pharmacol. 114, 993–998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waeber C.; Sebben M.; Nieoullon A.; Bockaert J.; Dumuis A. (1994) Regional distribution and ontogeny of 5-HT4 binding sites in rodent brain. Neuropharmacology 33, 527–541. [DOI] [PubMed] [Google Scholar]
- Domenech T.; Beleta J.; Fernandez A. G.; Gristwood R. W.; Cruz Sanchez F.; Tolosa E.; Palacios J. M. (1994) Identification and characterization of serotonin 5-HT4 receptor binding sites in human brain: comparison with other mammalian species. Mol. Brain Res. 21, 176–180. [DOI] [PubMed] [Google Scholar]
- Bockaert J.; Claeysen S.; Compan V.; Dumuis A. (2004) 5-HT4 receptors. Curr. Drug Targets: CNS Neurol. Disord. 3, 39–51. [DOI] [PubMed] [Google Scholar]
- Manuel-Apolinar L.; Rocha L.; Pascoe D.; Castillo E.; Castillo C.; Meneses A. (2005) Modifications of 5-HT4 receptor expression in rat brain during memory consolidation. Brain Res. 1042, 73–81. [DOI] [PubMed] [Google Scholar]
- Kojro E.; Postina R.; Buro C.; Meiringer C.; Gehrig-Burger K.; Fahrenholz F. (2006) The neuropeptide PACAP promotes the alpha-secretase pathway for processing the Alzheimer amyloid precursor protein. FASEB J. 20, 512–514. [DOI] [PubMed] [Google Scholar]
- Nitsch R. M.; Slack B. E.; Wurtman R. J.; Growdon J. H. (1992) Release of Alzheimer amyloid precursor derivatives stimulated by activation of muscarinic acetylcholine receptors. Science 258, 304–307. [DOI] [PubMed] [Google Scholar]
- Kuhn P. H.; Wang H.; Dislich B.; Colombo A.; Zeitschel U.; Ellwart J. W.; Kremmer E.; Rossner S.; Lichtenthaler S. F. (2010) ADAM10 is the physiologically relevant, constitutive alpha-secretase of the amyloid precursor protein in primary neurons. EMBO J. 29, 3020–3032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Strooper B.; Umans L.; Van Leuven F.; Van Den Berghe H. (1993) Study of the synthesis and secretion of normal and artificial mutants of murine amyloid precursor protein (APP): cleavage of APP occurs in a late compartment of the default secretion pathway. J. Cell Biol. 121, 295–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robert S. J.; Zugaza J. L.; Fischmeister R.; Gardier A. M.; Lezoualc’h F. (2001) The human serotonin 5-HT4 receptor regulates secretion of non-amyloidogenic precursor protein. J. Biol. Chem. 276, 44881–44888. [DOI] [PubMed] [Google Scholar]
- Bockaert J.; Claeysen S.; Compan V.; Dumuis A. (2008) 5-HT(4) receptors: history, molecular pharmacology and brain functions. Neuropharmacology 55, 922–931. [DOI] [PubMed] [Google Scholar]
- Claeysen S.; Sebben M.; Becamel C.; Bockaert J.; Dumuis A. (1999) Novel brain-specific 5-HT4 receptor splice variants show marked constitutive activity: role of the C-terminal intracellular domain. Mol. Pharmacol. 55, 910–920. [PubMed] [Google Scholar]
- Pellissier L. P.; Sallander J.; Campillo M.; Gaven F.; Queffeulou E.; Pillot M.; Dumuis A.; Claeysen S.; Bockaert J.; Pardo L. (2009) Conformational toggle switches implicated in basal constitutive and agonist-induced activated states of 5-hydroxytryptamine-4 receptors. Mol. Pharmacol. 75, 982–990. [DOI] [PubMed] [Google Scholar]
- Endres K.; Fahrenholz F. (2010) Upregulation of the alpha-secretase ADAM10—risk or reason for hope?. FEBS J. 277, 1585–1596. [DOI] [PubMed] [Google Scholar]
- Marcinkiewicz M.; Seidah N. G. (2000) Coordinated expression of beta-amyloid precursor protein and the putative beta-secretase BACE and alpha-secretase ADAM10 in mouse and human brain. J. Neurochem. 75, 2133–2143. [DOI] [PubMed] [Google Scholar]
- Jorissen E.; Prox J.; Bernreuther C.; Weber S.; Schwanbeck R.; Serneels L.; Snellinx A.; Craessaerts K.; Thathiah A.; Tesseur I.; Bartsch U.; Weskamp G.; Blobel C. P.; Glatzel M.; De Strooper B.; Saftig P. (2010) The disintegrin/metalloproteinase ADAM10 is essential for the establishment of the brain cortex. J. Neurosci. 30, 4833–4844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcello E.; Gardoni F.; Di Luca M.; Perez-Otano I. (2010) An arginine stretch limits ADAM10 exit from the endoplasmic reticulum. J. Biol. Chem. 285, 10376–10384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer A. M. (2011) Neuroprotective therapeutics for Alzheimer’s disease: progress and prospects. Trends Pharmacol. Sci. 32, 141–147. [DOI] [PubMed] [Google Scholar]
- Panza F.; Frisardi V.; Imbimbo B. P.; Capurso C.; Logroscino G.; Sancarlo D.; Seripa D.; Vendemiale G.; Pilotto A.; Solfrizzi V. (2010) REVIEW: gamma-Secretase inhibitors for the treatment of Alzheimer’s disease: The current state. CNS Neurosci. Ther. 16, 272–284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He G.; Luo W.; Li P.; Remmers C.; Netzer W. J.; Hendrick J.; Bettayeb K.; Flajolet M.; Gorelick F.; Wennogle L. P.; Greengard P. (2010) Gamma-secretase activating protein is a therapeutic target for Alzheimer’s disease. Nature 467, 95–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim M.; Suh J.; Romano D.; Truong M. H.; Mullin K.; Hooli B.; Norton D.; Tesco G.; Elliott K.; Wagner S. L.; Moir R. D.; Becker K. D.; Tanzi R. E. (2009) Potential late-onset Alzheimer’s disease-associated mutations in the ADAM10 gene attenuate {alpha}-secretase activity. Hum. Mol. Genet. 18, 3987–3996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costa R. M.; Drew C.; Silva A. J. (2005) Notch to remember. Trends Neurosci 28, 429–435. [DOI] [PubMed] [Google Scholar]
- Jones C. K.; Brady A. E.; Davis A. A.; Xiang Z.; Bubser M.; Tantawy M. N.; Kane A. S.; Bridges T. M.; Kennedy J. P.; Bradley S. R.; Peterson T. E.; Ansari M. S.; Baldwin R. M.; Kessler R. M.; Deutch A. Y.; Lah J. J.; Levey A. I.; Lindsley C. W.; Conn P. J. (2008) Novel selective allosteric activator of the M1 muscarinic acetylcholine receptor regulates amyloid processing and produces antipsychotic-like activity in rats. J. Neurosci. 28, 10422–10433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong E. H.; Reynolds G. P.; Bonhaus D. W.; Hsu S.; Eglen R. M. (1996) Characterization of [3H]GR 113808 binding to 5-HT4 receptors in brain tissues from patients with neurodegenerative disorders. Behav. Brain Res. 73, 249–252. [DOI] [PubMed] [Google Scholar]
- Tsang S. W.; Keene J.; Hope T.; Spence I.; Francis P. T.; Wong P. T.; Chen C. P.; Lai M. K. (2010) A serotoninergic basis for hyperphagic eating changes in Alzheimer’s disease. J. Neurol. Sci. 288, 151–155. [DOI] [PubMed] [Google Scholar]
- Ozaki N.; Shibasaki T.; Kashima Y.; Miki T.; Takahashi K.; Ueno H.; Sunaga Y.; Yano H.; Matsuura Y.; Iwanaga T.; Takai Y.; Seino S. (2000) cAMP-GEFII is a direct target of cAMP in regulated exocytosis. Nat. Cell Biol. 2, 805–811. [DOI] [PubMed] [Google Scholar]
- Meffre J.; Chaumont-Dubel S.; Mannoury la Cour C.; Loiseau F.; Watson D. J.; Dekeyne A.; Seveno M.; Rivet J. M.; Gaven F.; Deleris P.; Herve D.; Fone K. C.; Bockaert J.; Millan M. J.; Marin P. (2012) 5-HT(6) receptor recruitment of mTOR as a mechanism for perturbed cognition in schizophrenia. EMBO Mol. Med. 4, 1043–1056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weidemann A.; Konig G.; Bunke D.; Fischer P.; Salbaum J. M.; Masters C. L.; Beyreuther K. (1989) Identification, biogenesis, and localization of precursors of Alzheimer’s disease A4 amyloid protein. Cell 57, 115–126. [DOI] [PubMed] [Google Scholar]
- Dumuis A.; Bouhelal R.; Sebben M.; Cory R.; Bockaert J. (1988) A nonclassical 5-hydroxytryptamine receptor positively coupled with adenylate cyclase in the central nervous system. Mol. Pharmacol. 34, 880–887. [PubMed] [Google Scholar]
- Janes P. W.; Saha N.; Barton W. A.; Kolev M. V.; Wimmer-Kleikamp S. H.; Nievergall E.; Blobel C. P.; Himanen J. P.; Lackmann M.; Nikolov D. B. (2005) Adam meets Eph: an ADAM substrate recognition module acts as a molecular switch for ephrin cleavage in trans. Cell 123, 291–304. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.