

Abstract
The complexities of the involvement of the serotonin transmitter system in numerous biological processes and psychiatric disorders is, to a substantial degree, attributable to the large number of serotonin receptor families and subtypes that have been identified and characterized for over four decades. Of these, the 5-HT1A receptor subtype, which was the first to be cloned and characterized, has received considerable attention based on its purported role in the etiology and treatment of mood and anxiety disorders. 5-HT1A receptors function both at presynaptic (autoreceptor) and postsynaptic (heteroreceptor) sites. Recent research has implicated distinct roles for these two populations of receptors in mediating emotion-related behavior. New concepts as to how 5-HT1A receptors function to control serotonergic tone throughout life were highlights of the proceedings of the 2012 Serotonin Club Meeting in Montpellier, France. Here, we review recent findings and current perspectives on functional aspects of 5-HT1A auto- and heteroreceptors with particular regard to their involvement in altered anxiety and mood states.
Keywords: Serotonin, autoreceptor, heteroreceptor, development, behavior, anxiety, antidepressant
Historical Perspective
Serotonin (5-hydroxytryptamine; 5-HT) is a monoamine neurotransmitter and neurohormone formed by the hydroxylation and subsequent decarboxylation of the essential dietary amino acid l-tryptophan. Serotonin is found primarily in the gastrointestinal tract, platelets, blood vessels, thyroid, pancreas, mammary glands,1−5 and central nervous system (CNS).6,7 In the brain, serotonin is thought to be a key modulatory neurotransmitter involved in the regulation of numerous physiological and behavioral processes including mood- and anxiety-related behavior, cognitive function, food intake, sexual behavior, sleep, cardiovascular function, blood pressure, pain, body temperature, and others.8−11
Serotonin was first reported in 1937 by Vialli and Erspamer and named enteramine.12,13 In 1948, it was identified as a vasoconstrictor in blood serum where it was referred to as “serotonin”.14 Afterward, scientists realized that enteramine and serotonin were one in the same.15,16 Serotonin was recognized as a neurotransmitter when it was discovered in extracts from mammalian brain.17,18 In 1986, the pharmacology of serotonin was reviewed,19 and for the first time the existence of three receptor families (5-HT1–3) was described; additional families were suspected. It is now known that the effects of serotonin are mediated by at least 14 different receptors, which are grouped into subfamilies based on pharmacological responses to specific ligands, sequence similarities at the gene and amino acid levels, gene organization, and second messenger coupling pathways.20−23 Serotonin receptors are assigned to one of seven families, 5-HT1–7, with individual subtypes further designated by letters.
Among serotonin receptors, much attention has been focused on the 1A subtype (5-HT1A). The human 5-HT1A receptor was cloned in 1987 as a single intronless gene24 located on chromosome 5 (5q11.2-q13). In mice, the Htr1a gene resides on the distal part of chromosome 13. The 5-HT1A receptor protein consists of 422 amino acids. Evidence from human and rodent studies suggests that 5-HT1A receptors are implicated in a variety of physiological and pathological processes, such as learning, memory, schizophrenia, Parkinson’s disease, and notably in the etiology and treatment of mood and anxiety disorders.25−32
Adult Expression and Function of 5-HT1A Receptors
Direct Autonomous Inhibition
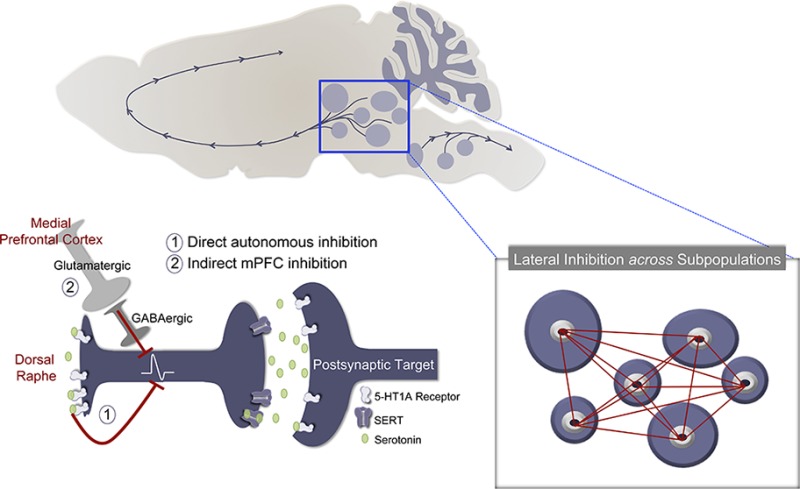
In the mammalian brain, 5-HT1A receptors are divided into two distinct classes based on localization. The cell bodies of central serotonergic neurons are found in the raphe nuclei in the brain stem (Figure 1A),33,34 where 5-HT1A receptors are located on soma and dendrites.35−37 These 5-HT1A autoreceptors exert inhibitory feedback in response to local release of serotonin in the raphe nuclei from axonal collaterals (Figure 1B).38−43 Additionally, sources of local serotonin release have been proposed to arise from somatic and dendritic sites on serotonergic neurons.44−46 Serotonin release in the cell body region results in reductions in serotonergic pacemaker activity and suppression of serotonin synthesis, turnover, and release in projection areas.47−51 In support of this, constitutive 5-HT1A knockout mice show increased rates of serotonin neuronal discharge52 and elevated basal dialysate serotonin levels in frontal cortex and hippocampus.53
Figure 1.

Inhibitory mechanisms of 5-HT1A autoreceptors. (A) Serotonergic cell bodies expressing 5-HT1A autoreceptors are located deep within the brainstem (blue box). Serotonin neurons projecting to the forebrain are organized into two main clusters designated as dorsal and median raphe nuclei with distinct subpopulations within these primary nuclei. 5-HT1A heteroreceptors are localized postsynaptically in various brain regions including the hippocampus (HIP), prefrontal cortex (PFC), thalamus (TH), lateral septum (SEP), amygdala (AMYG), and hypothalamic nuclei (HYP). Many of these regions have been associated with the pathophysiology of mood and anxiety disorders. (B) The top panel depicts a “conventional” serotonin synapse. In the bottom panel, activation of 5-HT1A autoreceptors controls serotonergic tone via one-to-one autoinhibitory feedback to reduce firing rates of serotonin neurons. 5-HT1A heteroreceptors also regulate serotonergic activity through descending glutamatergic projections originating in the medial prefrontal cortex (mPFC). This pathway makes connections with serotonin neurons via brainstem GABAergic inhibitory interneurons. (C) Current hypotheses regarding inhibitory mechanisms of 5-HT1A autoreceptor activation paint a more complex picture. For instance, emerging evidence suggests that serotonin neurons within a subpopulation or even across subpopulations affect each other via “lateral inhibition”. Taken together, new perspectives on the functional aspects of 5-HT1A receptors associated with regulation of serotonergic activity are important avenues for future investigation, particularly regarding increased understanding of the roles of 5-HT1A receptors in the etiology and treatment of psychiatric disorders.
Indirect Inhibition from Medial Prefrontal Cortex
5-HT1A receptors are also expressed by nonserotonergic pyramidal, GABAergic, and cholinergic neurons54−58 in limbic regions such as prefrontal cortex, hippocampus, lateral septum, and amygdala, as well as in several hypothalamic and thalamic nuclei (Figure 1A).37,59,60 Activation of 5-HT1A heteroreceptors mediates hyperpolarizing responses to released serotonin, which typically reduces postsynaptic neuronal excitability and firing rates.37,40,59 As such, 5-HT1A heteroreceptors are involved in the modulation of other neurotransmission systems. For example, 5-HT1A receptors in the medial prefrontal cortex (mPFC) modulate dopamine cell firing and release.61−63
In addition to direct/local 5-HT1A receptor-mediated autoregulation, evidence exists for an indirect negative feedback mechanism that involves 5-HT1A heteroreceptors in the mPFC (Figure 1B).64−66 The mPFC-dorsal raphe nucleus (DRN) pathway comprises glutamatergic descending projections that are hypothesized to decrease serotonin cell firing by activating DRN GABAergic interneurons to inhibit serotonin release.65−72 This pathway has been suggested as one of the neuroanatomical substrates controlling the effects of stress by preventing overactivation of DRN serotonergic neurons.73 A minor glutamatergic pathway may also exist from the mPFC that excites serotonin cell firing directly in the DRN.68,69 It is still not well understood how inhibitory 5-HT1A heteroreceptors excite cortical glutamatergic neurons. The most plausible explanation is the disinhibition of glutamatergic neurons via mPFC GABA interneurons expressing 5-HT1A heteroreceptors.58,74 However, further studies are required to determine the specific mechanisms of 5-HT1A receptors in the mPFC.
Intracellular Signaling
5-HT1A receptors are inhibitory G-protein coupled receptors (GPCRs). Early studies identified that 5-HT1A receptors function by coupling to Gi/Go proteins in most cells.75 Extracellular receptor binding of serotonin or 5-HT1A agonists leads to intracellular exchange of GDP for GTP on Gi/Go alpha subunits. This, in turn, inhibits adenylyl cyclase, which reduces cAMP levels and protein kinase A activity.76 Furthermore, agonist-induced activation of 5-HT1A receptors results in potassium channel activation and calcium channel inhibition.25,59,77−80 5-HT1A receptors are coupled to different GPCR pathways based on localization.81 5-HT1A autoreceptors couple exclusively to Gαi3 leading to partial inhibition of adenylyl cyclase,82−85 an effect that may depend on specific agonists.83,86,87 By contrast, 5-HT1A heteroreceptors couple mainly to Gαo in the hippocampus, and equally to Gαo and Gαi3 in cerebral cortex.88 Differences in 5-HT1A Gα subunit coupling might explain regional differences in activation versus inhibition of intracellular signaling pathways. 5-HT1A autoreceptor desensitization is more pronounced compared to 5-HT1A heteroreceptor desensitization, which also could be related to differential Gα coupling.88,89 Stimulation of 5-HT1A receptors also leads to activation of G-protein-coupled inward rectifying potassium channels (GIRKs)90 in raphe neurons86,91−95 and hippocampus.96−98 Whether 5-HT1A receptors fully couple to inhibit adenylyl cyclase remains controversial, as it has been suggested that the hyperpolarizing response mediated by 5-HT1A autoreceptors is due to the activation of GIRK channels via G-protein βγ subunits.99
In addition to their canonical function, 5-HT1A receptors activate growth factor-regulated signaling pathways, such as mitogen-activated protein kinases (MAPK) and Akt signaling pathways.25 In MAPK signaling, ERK is preferentially affected by 5-HT1A receptors. For example, in RN46A cells, a model of serotonergic raphe neurons that express endogenous 5-HT1A receptors, adenylyl cyclase and ERK1/2 phosphorylation were inhibited by 5-HT1A-receptor activation.100 However, in hippocampal-derived differentiated HN2-5 cells, 5-HT1A agonists increased ERK phosphorylation and activity.101 These and other studies suggest that the modulation of ERK may depend on neuronal origin, as well as maturation states.102−106
The Akt signaling pathway is also activated in 5-HT1A receptor-expressing cells and primary hippocampal neurons.104,107,108 Activation of Akt by 5-HT1A receptors led to inactivation of GSK3β in hippocampal cultures,104,109,110 an effect also observed in raphe cultures.105 Recently, it has been suggested that the GSK3-regulating effects of 5-HT1A receptors are mediated by the PI3K/Akt signaling pathway.111 Taken together, 5-HT1A autoreceptors and heteroreceptors have diverse intracellular signaling capabilities contributing to the complex regulation of the serotonin system, as well as neuronal networks modulated by serotonin.
5-HT1A Receptors in Development
Serotonin is a morphogenic factor,112 and alteration of serotonin levels during early developmental windows has been shown to influence mood- and anxiety-related behavior in adult animals.113−115 Thus, mapping and understanding factors that regulate the developmental trajectory of 5-HT1A receptor expression are important given that autoreceptors modulate serotonin release and heteroreceptors are widely expressed in emotion-related circuits, where they mediate the effects of released serotonin. In rodents, 5-HT1A receptors appear in early embryonic development. Using in situ hybridization and immunocytochemistry, 5-HT1A mRNA has been detected as soon as embryonic day (E) 12 and 5-HT1A receptor protein by E14 in neuronal cultures prepared from brain stem.116 A surge in 5-HT1A mRNA levels in the brain stem occurs beginning on E13, with peak levels occurring at E15–16.117 Developmentally related changes in 5-HT1A receptor expression also occur at postsynaptic sites. Transient expression is evident in the septum and preoptic region during embryonic development.118 5-HT1A expression occurs at high levels in the cerebellum during the first two postnatal weeks tapering off to near undetectable levels by postnatal day (P) 21.119,120 In brain regions associated with the regulation of mood and anxiety, 5-HT1A-receptor expression likewise exhibits complex patterns during development. In situ hybridization and [3H]-8-OH-DPAT binding both indicate low 5-HT1A expression in the dentate gyrus granule cell layer of the hippocampus at E14.5, with levels gradually increasing during the first few postnatal weeks and reaching near adult levels by P13.118−121
Reports on the ontogeny of 5-HT1A receptors in the developing human brain, albeit fewer, are in agreement with those on rodents. In one study, [3H]-8-OH-DPAT binding was used to examine 5-HT1A receptor expression from tissue acquired at different stages of gestation.122 Although expression levels varied with respect to gestational age (16–22 weeks), the relative regional distribution of 5-HT1A receptors was similar across time points. The highest receptor densities occurred in subregions of the hippocampus and frontal cortex, where a surge in 5-HT1A expression was observed at 18–22 weeks of gestation resulting in levels 3–4 times higher than those reported in adults.122 Others have used in situ hybridization to compare 5-HT1A receptor mRNA levels in human tissue collected at ∼28 weeks of gestation compared to 6–7 years of age.123 Human fetal brain contained the highest 5-HT1A mRNA levels in raphe nuclei, cerebellum, and the CA1 and dentate gyrus regions of the hippocampus. When compared to later ages, only expression in the cerebellum showed dramatic changes, with mRNA levels being lower in children and undetectable in adults.123,124 The latter is similar to the cerebellar expression pattern reported in rodents. Thus, 5-HT1A-receptor expression undergoes tight temporal regulation and in some regions, brief emergence and disappearance, suggesting morphogenic influences, especially in areas of the brain involved in modulating anxiety levels in adults.
A few studies have investigated functional aspects of 5-HT1A receptors during development by means of electrophysiology or pharmacologic challenge. Electrophysiological recordings in the prefrontal cortex of rats indicated that 5-HT1A receptor-mediated hyperpolarization appears late in postnatal development between P16 and P19 corresponding temporally with increases in receptor protein expression.118−120,125 In mice, 5-HT1A-receptor mediated outward currents were not evident at P4 but were observed by P12.126 5-HT1A responses developed after the appearance of physiology characteristic of serotonergic neurons, which appeared to be fully developed by P12. 5-HT1A receptor-signaling pathways important for mediating developmental processes are complex. For example, 8-OH-DPAT-induced activation of the MAPK pathway, and specifically ERK1/2 kinases, requires different PKC isozymes to regulate distinct developmental processes (e.g., cell division versus synaptic activity/strengthening) at P6 versus P15.127 More work will be needed to understand functional changes in 5-HT1A mediated pre- and postsynaptic responses during postnatal development, particularly in terms of critical developmental windows important for shaping and determining anxiety-related behavior later in life.
5-HT1A Receptors in Mood and Anxiety
Mechanisms of Antidepressants
5-HT1A autoreceptors play an important role in regulating serotonergic activity through feedback inhibition pathways. As such, the involvement of 5-HT1A receptors in the mechanism of action of antidepressants has been widely investigated, and literature supporting a role for this receptor population in the delayed efficacy of antidepressants has evolved. 5-HT1A autoreceptors (and possibly heteroreceptors) limit increases in extracellular serotonin levels induced by serotonin selective reuptake inhibitors (SSRIs). Even after chronic SSRI treatment, 5-HT1A autoreceptors maintain some control over serotonin release.128−132
The delayed efficacy of SSRIs has been partially attributed to the need for desensitization of 5-HT1A autoreceptors in the raphe nuclei, enabling firing rates of serotonergic neurons to overcome inhibition.128,133−137 However, only ∼50% of studies in rodents show increased extracellular serotonin after chronic administration of serotonin reuptake inhibiting antidepressants.138 In this context, concomitant administration of 5-HT1A receptor antagonists with SSRIs has been hypothesized to hasten or potentiate changes in serotonin levels to improve clinical efficacy.54,139 The ability of the partial 5-HT1A antagonist, pindolol, to accelerate antidepressant clinical efficacy has been investigated in patients with major depressive disorder. Meta-analyses suggest that pindolol administered in combination with SSRIs augments and accelerates symptomatic improvement after 2 weeks of treatment specifically in patients without prior history of treatment.140−142
However, not all findings support a role for 5-HT1A autoreceptor desensitization and its effects to accelerate SSRI treatment. For example, administration of pindolol in combination with SSRIs to patients with treatment resistant depression or extensive treatment histories does not hasten or produce symptom amelioration.143,144 Human studies examining the role of 5-HT1A receptors in the treatment and etiology of major depressive disorder indicate that sensitivity to receptor signaling after administration of agonists are complicated based on reports of reduced receptor number and binding, as well as genetic influences.145 Additionally, mechanisms of autoreceptor desensitization are not clear and may differ depending on the antidepressant administered. For example, chronic fluoxetine results in desensitization of autoreceptors, which is associated with a reduction in receptor stimulated [35S]GTPγS binding.146−149 However, G-protein coupling is not altered in association with administration of the SSRI sertraline, indicating that antidepressants differ with regard to mechanisms of 5-HT1A desensitization.146
In addition to conflicting findings regarding 5-HT1A autoreceptor desensitization in clinical studies, desensitization occurs in animal models of depression generated via chronic stress paradigms.150−154 5-HT1A autoreceptors and heteroreceptors are also desensitized in serotonin transporter knockout mice in association with a phenotype characterized by enhanced anxiety.155−158 Together, these studies highlight the complex regulation of 5-HT1A pathways, such that desensitization may be common to both the treatment and precipitating etiological factors associated with major depressive disorder.
Animal Models of 5-HT1A Disruption
In light of data suggesting a role for 5-HT1A receptors in mediating antidepressant responses in humans, it is logical to suspect that these receptors also function in the neurocircuitry and pathophysiology of emotion-related behavior. Genetic manipulation of the murine Htr1A gene has been carried out to study the behavioral effects of receptor under- or overexpression. In both cases, the majority of data point to a role for 5-HT1A receptors in anxiety-related behavior. Table 1 in the Supporting Information provides a comprehensive summary of the literature to date focused on anxiety- and depression-like behaviors, and learning and memory in animal models of altered 5-HT1A expression. Taken together, these studies provide convincing evidence that global reductions in 5-HT1A receptor expression lead to increased anxiety-related behavior in adult animals across different strains of mice and behavior test paradigms.
The first reports on mice with reduced 5-HT1A expression appeared simultaneously in 1998. Three separate laboratories produced constitutive 5-HT1A knockout mice, each in a different background strain (Supporting Information Table 1).159−161 Consistent changes in behavior were observed in tests for anxiety-related behavior associated with conflict in novel environments, including exploration in the center of the open field or the open arms of the elevated plus maze. Changes in behavior were more consistently replicated across tests in male versus female mice. In terms of depression-like behaviors, mice with reduced 5-HT1A expression displayed reduced immobility in the forced swim160,161 and tail suspension tests.159 However, interpretations of these behavioral changes were not in agreement. Some authors perceived decreased immobility as an increased stress response,160 while others took a more traditional view of reduced immobility162 as indicative of decreased learned helplessness/behavioral despair or an increased antidepressant-like response.159,160
Following these initial reports, others confirmed increased anxiety-like behavior in 5-HT1A knockout mice when presented with conflict situations (see Supporting Information Table 1 for a full list of references). Findings also included modestly enhanced fear responses to stressful stimuli such as foot shock.163−166 In terms of learning and memory, mice with reduced 5-HT1A expression displayed deficits in hippocampal-dependent learning indicated by increased latencies and path lengths to find the hidden platform in the Morris water maze.167,168 As opposed to mice with constitutive deletion of 5-HT1A receptors, mice with developmentally limited overexpression of 5-HT1A heteroreceptors in dentate gyrus and cortex also displayed reduced anxiety in novel, conflict environments.169 By contrast, mice with permanent overexpression failed to show changes in anxiety-like behaviors.170 Similar to 5-HT1A knockout mice, transient 5-HT1A overexpression resulted in deficits in hippocampal-dependent learning and memory in the Morris water maze.171 Deficits were less pronounced or absent in permanent overexpressing mice.172 Information pertaining to changes in learning and memory after 5-HT1A receptor manipulation is reviewed in detail elsewhere.172,173
Recently, studies have been carried out to delete specific subpopulations of 5-HT1A receptors enabling the differentiation of pre- versus postsynaptic behavioral effects.174 When 5-HT1A autoreceptors were selectively inactivated, mice exhibited increased anxiety-like behavior similar to constitutive knockout mice. These findings highlight the importance of raphe 5-HT1A receptors in mediating anxiety responses. By contrast, when 5-HT1A heteroreceptors were specifically targeted, normal anxiety-like behavior was observed; however, increased depression-like behavior was observed in the forced swim test.174 Increased anhedonic behavior related to reduced consumption of palatable food has also been observed in mice with 5-HT1A heteroreceptor knockout. The latter findings on depression-related behavior illustrate an interesting dichotomy since mice with constitutive deletion of 5-HT1A auto- and heteroreceptors show decreases in depression-like behavior (see Supporting Information Table 1). Thus, the synergistic effects of genetically inactivating auto- and hetero 5-HT1A receptors cannot be predicted by studying knockout of either receptor population in isolation.
In addition to mice with complete loss of 5-HT1A autoreceptors, mice with 30% reductions in 5-HT1A autoreceptor expression have been generated.175 These mice display potentiated responses to the administration of fluoxetine as indicated by increased dialysate serotonin levels. These findings are important in the context of human HTR1A gene polymorphisms, including the C(−1019)G HTR1A single nucleotide polymorphism,176−178 which is hypothesized to affect the expression and function of 5-HT1A receptors and to contribute to the high numbers of antidepressant nonresponders.179
New Perspectives on 5-HT1A Autoreceptors
Networks and 5-HT1A Inhibitory Function
It is widely accepted that 5-HT1A receptors located on serotonergic neurons participate in autoinhibition. However, a more complex picture of inhibitory function than simple one-to-one regulation of serotonin cell firing is emerging. Serotonin neurons are anatomically heterogeneous and their projections to the forebrain are topographically organized.180−183 Furthermore, distinct, reproducible patterns of serotonin neuron activation within the raphe nuclei can be seen in response to specific external stimuli, suggesting that anatomical topography underlies functional topography.182,184 Additionally, microdialysis studies examining extracellular serotonin levels after the application of various stressors have supported the idea of defined network patterns of serotonin neurons such that stress-induced changes in serotonin levels are stressor- and region specific.185 These observations suggest that feedback inhibitory pathways might work to control serotonin-network activity. In one model, raphe serotonin neurons exhibit autonomous feedback such that groups of functionally similar serotonergic neurons regulate themselves in a homeostatic manner (Figure 1C).180
Nonautonomous feedback has also been suggested in which crosstalk between distinct groups of serotonin neurons provides “lateral inhibition” influencing patterns of activation (Figure 1C).180 Along these lines, recent work by Commons and Sperling has demonstrated complementary patterns of activation of serotonergic neurons after exposure to and withdrawal from nicotine, which are different and reciprocally switched after 5-HT1A receptor blockade.181 These findings suggest that endogenous feedback inhibition provided by 5-HT1A receptors might be regionally organized and depend on behavioral states. While these observations do not exclude an autoregulatory function, they raise the possibility that 5-HT1A receptors might also operate in a nonautonomous fashion to mediate communication between different groups of serotonin neurons, thus suggesting that 5-HT1A receptors are involved in a regional form of autoregulation between different raphe subfields. These studies are consistent with anatomical studies showing that there are many interconnections between the different raphe nuclei180 and axon collaterals of serotonin neurons travel for some distance within the dorsal raphe nuclei.186
There are also a number of distinct serotonin subsystems with unique genetic programming and functions.187 Recent studies have shown that, in raphe nuclei, there are at least three different serotonin cell-types grouped by anatomical, physiological, and molecular characteristics, and their distribution transcends the traditional anatomical classification of raphe subfields. Furthermore, forebrain regions receiving serotonergic projections are innervated by serotonin neurons with distinct characteristics, forming a highly organized circuit.188 Therefore, identifying and understanding functionally specific axon collaterals within the raphe that are involved in controlling emotional behaviors is likely to be an important future direction of research.
Future Prospects
Targeting Specific Receptor Populations
The evolving distinctions between the specific and diverse roles for 5-HT1A auto- versus heteroreceptors and the manner in which different populations of neurons expressing 5-HT1A receptors elicit control over serotonergic tone is now shifting away from simplified views centered on autonomous autoreceptor-mediated feedback inhibition. Classical definitions of autoreceptors, which have long been associated with controlling serotonergic firing rates via feedback inhibition, remain an important functional property of these receptors. Yet, rather than one-to-one associations, emerging findings point to the idea that inhibitory regulation is also more complex and might operate at several levels that involve multiple pathways and/or networks that influence neighboring circuits.
As the rich complexity of 5-HT1A receptor organization and function continues to be uncovered, optimizing treatment strategies for mood and anxiety disorders by preferentially targeting different 5-HT1A receptor populations or networks to produce therapeutic effects should become possible. For example, the novel 5-HT1A agonist, F15599, has been shown to reduce immobility in the forced swim test189 and to promote these effects primarily through activation of 5-HT1A heteroreceptors localized in the frontal cortex.106 These findings support the idea that in addition to 5-HT1A autoreceptor desensitization following antidepressant treatment, increased 5-HT1A heteroreceptor activity occurs to promote antidepressant efficacy.190−193 Furthermore, in support of the roles of 5-HT1A autoreceptors in mood and anxiety-related behavior, recent advances using siRNA to silence 5-HT1A autoreceptor expression have been shown to ameliorate immobility associated with learned helplessness and to augment fluoxetine-induced increases in serotonin levels in postsynaptic regions.194,195
5-HT1A Receptors: A View of the Future
The serotonin system, and 5-HT1A receptors in particular, exhibit properties that continue to challenge and enlarge our understanding of receptor function. In addition to direct autonomous inhibition, evidence now exists for indirect inhibition of raphe serotonergic neurons arising from a mPFC-DRN pathway. It will be informative to sort out whether direct and indirect pathways work independently or synergistically to limit serotonin release in the mPFC. Moreover, the actions of serotonin might be coordinated across different brain regions as part of a global circuitry via lateral inhibition within and across subpopulations of raphe neurons, some of which have different developmental origins and genetic lineages. Thus, a picture of 5-HT1A receptor-mediated control of emotion-related behavior via coordination of brain-wide networks is coming to light. Still, additional work will be needed to uncover precise interactions between different parts of this network, hierarchical principles of 5-HT1A receptor function, and early life sequencing with regard to lasting consequences for network development.
Many animal models of altered 5-HT1A receptor expression have been created. Taken together, studies on these models highlight two important aspects of 5-HT1A receptors in regard to anxiety-related behavior. First, most studies point to 5-HT1A receptors, particularly during postnatal development, as key regulators of anxiety (see Supporting Information Table 1). By contrast, pharmacologic inhibition or genetic manipulation of 5-HT1A receptors in adult animals fails to produce anxiogenic behavior.196,197 Second, unraveling the significance of 5-HT1A auto- versus heteroreceptors in regard to establishing baseline anxiety behavior is important. Null 5-HT1A mutant mice in which 5-HT1A heteroreceptors were ectopically overexpressed during development169 or where 5-HT1A receptor expression was restored in forebrain regions, for example, hippocampus, cortex, and striatum,196 exhibit reversal or rescue of increased anxiety behavior, respectively. However, the importance of heteroreceptors in gain-of-function experiments has been questioned by recent tissue-specific conditional knockout strategies where 5-HT1A autoreceptors, but not heteroreceptors, have been shown to be critical components for establishing normal levels of anxiety.174 As additional information on the specific roles of different 5-HT1A receptor subpopulations and their influence over emotion-related behaviors during various timeframes become clear, avenues for novel treatment strategies for mood and anxiety disorder should become evident.
Further, as we sort out which 5-HT1A receptor inhibitory circuits play key roles in shaping specific types of behavior, these are also expected to become targets for the development of more selective and, hopefully, more effective therapeutics. Recent findings have shown that different populations of 5-HT1A receptors are coupled to different/multiple intracellular signaling pathways. This suggests the possibility of developing allosteric modulators or other types of small-molecule drugs that modify specific intracellular signaling pathways in cases where a particular type of receptor is coupled to more than one pathway in the same cell type or to different Gα subunits in different cells types.
Finally, the serotonin system remains unique among known neurotransmitter systems in that it is the only system identified as having two molecularly distinct autoreceptors. In addition to 5-HT1A receptors, serotonergic function is regulated by 5-HT1B autoreceptors, which are expressed presynaptically on serotonin axon terminals, in addition to postsynaptic axonal localization associated with dopaminergic, GABAergic, and glutamatergic systems.198 Thus, both 5-HT1A and 5-HT1B receptors function as autoreceptors and heteroreceptors. It is tempting to think that 5-HT1B receptors might exhibit complexity similar to 5-HT1A receptors in terms of multiple inhibitory circuits or hierarchical organization. Perhaps, continued investigation of 5-HT1A (and 5-HT1B) receptors, as well as other receptor subtypes that function both as auto- and heteroreceptors, for example, dopamine D2 and noradrenergic alpha2 receptors, will reveal additional modes of regulatory feedback and network organization. Nonetheless, it appears that recent advances in understanding 5-HT1A receptor function lead the way in terms of extending and expanding our thinking as to how behavioral circuits are organized and controlled by a specific receptor subtype. They also provide exciting new opportunities for drug discovery and development, particularly for the treatment of mood and anxiety disorders.
Supporting Information Available
Table detailing behavior phenotypes in models of 5-HT1A receptor disruption or overexpression. This material is available free of charge via the Internet at http://pubs.acs.org.
The authors declare no competing financial interest.
Author Contributions
⊥ These authors contributed equally.
Supplementary Material
References
- Cote F.; Fligny C.; Bayard E.; Launay J. M.; Gershon M. D.; Mallet J.; Vodjdani G. (2007) Maternal serotonin is crucial for murine embryonic development. Proc. Nat.l Acad. Sci. U.S.A. 104, 329–334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gershon M. D. (2000) 5-HT (serotonin) physiology and related drugs. Curr. Opin. Gastroenterol. 16, 113–120. [DOI] [PubMed] [Google Scholar]
- Paulmann N.; Grohmann M.; Voigt J. P.; Bert B.; Vowinckel J.; Bader M.; Skelin M.; Jevsek M.; Fink H.; Rupnik M.; Walther D. J. (2009) Intracellular serotonin modulates insulin secretion from pancreatic beta-cells by protein serotonylation. PLoS Biol. 7, e1000229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walther D. J.; Peter J. U.; Bashammakh S.; Hortnagl H.; Voits M.; Fink H.; Bader M. (2003) Synthesis of serotonin by a second tryptophan hydroxylase isoform. Science 299, 76. [DOI] [PubMed] [Google Scholar]
- Berger M.; Gray J. A.; Roth B. L. (2009) The expanded biology of serotonin. Annu. Rev. Med. 60, 355–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deneris E. S.; Wyler S. C. (2012) Serotonergic transcriptional networks and potential importance to mental health. Nat. Neurosci. 15, 519–527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hensler J. G. (2006) Serotonergic modulation of the limbic system. Neurosci. Biobehav. Rev. 30, 203–214. [DOI] [PubMed] [Google Scholar]
- Jonnakuty C.; Gragnoli C. (2008) What do we know about serotonin?. J. Cell. Physiol. 217, 301–306. [DOI] [PubMed] [Google Scholar]
- Lam D. D.; Garfield A. S.; Marston O. J.; Shaw J.; Heisler L. K. (2010) Brain serotonin system in the coordination of food intake and body weight. Pharmacol., Biochem. Behav. 97, 84–91. [DOI] [PubMed] [Google Scholar]
- Watts S. W.; Morrison S. F.; Davis R. P.; Barman S. M. (2012) Serotonin and blood pressure regulation. Pharmacol. Rev. 64, 359–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veasey S. C.; Fornal C. A.; Metzler C. W.; Jacobs B. L. (1995) Response of serotonergic caudal raphe neurons in relation to specific motor activities in freely moving cats. J. Neurosci. 15, 5346–5359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vialli M. E., V. (1937) Richerche sul secreto delle cellule enterocromaffini IX Intorno alla natura chimica della sostanza specifica. Boll. Soc. Med.-Chir. Pavia 51, 1111–1116. [Google Scholar]
- Vialli M. E., V. (1940) Richerche di caratterizzazione chimica delle sostanze fenoliche presenti negli estratti acetonici di ghiandole salivari posteriori di Octopus vulgaris. Arch. Fisiol. 40, 293–302. [Google Scholar]
- Rapport M. M.; Green A. A.; Page I. H. (1948) Crystalline Serotonin. Science 108, 329–330. [DOI] [PubMed] [Google Scholar]
- Erspamer V.; Asero B. (1952) Identification of enteramine, the specific hormone of the enterochromaffin cell system, as 5-hydroxytryptamine. Nature 169, 800–801. [DOI] [PubMed] [Google Scholar]
- Erspamer V.; Asero B. (1953) Isolation of enteramine from extracts of posterior salivary glands of Octopus vulgaris and of Discoglossus pictus skin. J. Biol. Chem. 200, 311–318. [PubMed] [Google Scholar]
- Twarog B. M.; Page I. H. (1953) Serotonin content of some mammalian tissues and urine and a method for its determination. Am. J. Physiol. 175, 157–161. [DOI] [PubMed] [Google Scholar]
- Amin A. H.; Crawford T. B.; Gaddum J. H. (1954) The distribution of substance P and 5-hydroxytryptamine in the central nervous system of the dog. J. Physiol. 126, 596–618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradley P. B.; Engel G.; Feniuk W.; Fozard J. R.; Humphrey P. P.; Middlemiss D. N.; Mylecharane E. J.; Richardson B. P.; Saxena P. R. (1986) Proposals for the classification and nomenclature of functional receptors for 5-hydroxytryptamine. Neuropharmacology 25, 563–576. [DOI] [PubMed] [Google Scholar]
- Pytliak M.; Vargova V.; Mechirova V.; Felsoci M. (2011) Serotonin receptors - from molecular biology to clinical applications. Physiol. Res. 60, 15–25. [DOI] [PubMed] [Google Scholar]
- Filip M.; Bader M. (2009) Overview on 5-HT receptors and their role in physiology and pathology of the central nervous system. Pharmacol. Rep. 61, 761–777. [DOI] [PubMed] [Google Scholar]
- Hannon J.; Hoyer D. (2008) Molecular biology of 5-HT receptors. Behav. Brain Res. 195, 198–213. [DOI] [PubMed] [Google Scholar]
- Nichols D. E.; Nichols C. D. (2008) Serotonin receptors. Chem. Rev. 108, 1614–1641. [DOI] [PubMed] [Google Scholar]
- Kobilka B. K.; Frielle T.; Collins S.; Yang-Feng T.; Kobilka T. S.; Francke U.; Lefkowitz R. J.; Caron M. G. (1987) An intronless gene encoding a potential member of the family of receptors coupled to guanine nucleotide regulatory proteins. Nature 329, 75–79. [DOI] [PubMed] [Google Scholar]
- Polter A. M.; Li X. (2010) 5-HT1A receptor-regulated signal transduction pathways in brain. Cell. Signalling 22, 1406–1412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akimova E.; Lanzenberger R.; Kasper S. (2009) The serotonin-1A receptor in anxiety disorders. Biol. Psychiatry 66, 627–635. [DOI] [PubMed] [Google Scholar]
- Gordon J. A.; Hen R. (2004) The serotonergic system and anxiety. NeuroMol. Med. 5, 27–40. [DOI] [PubMed] [Google Scholar]
- Hirvonen J.; Karlsson H.; Kajander J.; Lepola A.; Markkula J.; Rasi-Hakala H.; Nagren K.; Salminen J. K.; Hietala J. (2008) Decreased brain serotonin 5-HT1A receptor availability in medication-naive patients with major depressive disorder: An in-vivo imaging study using PET and [carbonyl-11C]WAY-100635. Int. J. Neuropsychopharmacol. 11, 465–476. [DOI] [PubMed] [Google Scholar]
- Le Francois B.; Czesak M.; Steubl D.; Albert P. R. (2008) Transcriptional regulation at a HTR1A polymorphism associated with mental illness. Neuropharmacology 55, 977–985. [DOI] [PubMed] [Google Scholar]
- Lesch K. P.; Gutknecht L. (2004) Focus on The 5-HT1A receptor: Emerging role of a gene regulatory variant in psychopathology and pharmacogenetics. Int. J. Neuropsychopharmacol. 7, 381–385. [DOI] [PubMed] [Google Scholar]
- Ohno Y. (2011) Therapeutic role of 5-HT1A receptors in the treatment of schizophrenia and Parkinson’s disease. CNS Neurosci. Ther. 17, 58–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strobel A.; Gutknecht L.; Rothe C.; Reif A.; Mossner R.; Zeng Y.; Brocke B.; Lesch K. P. (2003) Allelic variation in 5-HT1A receptor expression is associated with anxiety- and depression-related personality traits. J. Neural Transm. 110, 1445–1453. [DOI] [PubMed] [Google Scholar]
- Sotelo C.; Cholley B.; El Mestikawy S.; Gozlan H.; Hamon M. (1990) Direct Immunohistochemical Evidence of the Existence of 5-HT1A Autoreceptors on Serotoninergic Neurons in the Midbrain Raphe Nuclei. Eur. J. Neurosci. 2, 1144–1154. [DOI] [PubMed] [Google Scholar]
- Jans L. A.; Riedel W. J.; Markus C. R.; Blokland A. (2007) Serotonergic vulnerability and depression: assumptions, experimental evidence and implications. Mol. Psychiatry 12, 522–543. [DOI] [PubMed] [Google Scholar]
- Kia H. K.; Miquel M. C.; Brisorgueil M. J.; Daval G.; Riad M.; El Mestikawy S.; Hamon M.; Verge D. (1996) Immunocytochemical localization of serotonin1A receptors in the rat central nervous system. J. Comp. Neurol. 365, 289–305. [DOI] [PubMed] [Google Scholar]
- Pazos A.; Palacios J. M. (1985) Quantitative autoradiographic mapping of serotonin receptors in the rat brain. I. Serotonin-1 receptors. Brain Res. 346, 205–230. [DOI] [PubMed] [Google Scholar]
- Riad M.; Garcia S.; Watkins K. C.; Jodoin N.; Doucet E.; Langlois X.; el Mestikawy S.; Hamon M.; Descarries L. (2000) Somatodendritic localization of 5-HT1A and preterminal axonal localization of 5-HT1B serotonin receptors in adult rat brain. J. Comp. Neurol. 417, 181–194. [PubMed] [Google Scholar]
- Hjorth S.; Sharp T. (1991) Effect of the 5-HT1A receptor agonist 8-OH-DPAT on the release of 5-HT in dorsal and median raphe-innervated rat brain regions as measured by in vivo microdialysis. Life Sci. 48, 1779–1786. [DOI] [PubMed] [Google Scholar]
- Meller E.; Goldstein M.; Bohmaker K. (1990) Receptor reserve for 5-hydroxytryptamine1A-mediated inhibition of serotonin synthesis: Possible relationship to anxiolytic properties of 5-hydroxytryptamine1A agonists. Mol. Pharmacol. 37, 231–237. [PubMed] [Google Scholar]
- Sprouse J. S.; Aghajanian G. K. (1986) (-)-Propranolol blocks the inhibition of serotonergic dorsal raphe cell firing by 5-HT1A selective agonists. Eur. J. Pharmacol. 128, 295–298. [DOI] [PubMed] [Google Scholar]
- Verge D.; Daval G.; Patey A.; Gozlan H.; el Mestikawy S.; Hamon M. (1985) Presynaptic 5-HT autoreceptors on serotonergic cell bodies and/or dendrites but not terminals are of the 5-HT1A subtype. Eur. J. Pharmacol. 113, 463–464. [DOI] [PubMed] [Google Scholar]
- Wang R. Y.; Aghajanian G. K. (1977) Inhibiton of neurons in the amygdala by dorsal raphe stimulation: Mediation through a direct serotonergic pathway. Brain Res. 120, 85–102. [DOI] [PubMed] [Google Scholar]
- Pineyro G.; Blier P. (1999) Autoregulation of serotonin neurons: role in antidepressant drug action. Pharmacol. Rev. 51, 533–591. [PubMed] [Google Scholar]
- Colgan L. A.; Cavolo S. L.; Commons K. G.; Levitan E. S. (2012) Action potential-independent and pharmacologically unique vesicular serotonin release from dendrites. J. Neurosci. 32, 15737–15746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colgan L. A.; Putzier I.; Levitan E. S. (2009) Activity-dependent vesicular monoamine transporter-mediated depletion of the nucleus supports somatic release by serotonin neurons. J. Neurosci. 29, 15878–15887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Kock C. P.; Cornelisse L. N.; Burnashev N.; Lodder J. C.; Timmerman A. J.; Couey J. J.; Mansvelder H. D.; Brussaard A. B. (2006) NMDA receptors trigger neurosecretion of 5-HT within dorsal raphe nucleus of the rat in the absence of action potential firing. J. Physiol. 577, 891–905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jolas T.; Haj-Dahmane S.; Lanfumey L.; Fattaccini C. M.; Kidd E. J.; Adrien J.; Gozlan H.; Guardiola-Lemaitre B.; Hamon M. (1993) -)Tertatolol is a potent antagonist at pre- and postsynaptic serotonin 5-HT1A receptors in the rat brain. Naunyn-Schmiedeberg's Arch. Pharmacol. 347, 453–463. [DOI] [PubMed] [Google Scholar]
- Bohmaker K.; Eison A. S.; Yocca F. D.; Meller E. (1993) Comparative effects of chronic 8-OH-DPAT, gepirone and ipsapirone treatment on the sensitivity of somatodendritic 5-HT1A autoreceptors. Neuropharmacology 32, 527–534. [DOI] [PubMed] [Google Scholar]
- Kennett G. A.; Marcou M.; Dourish C. T.; Curzon G. (1987) Single administration of 5-HT1A agonists decreases 5-HT1A presynaptic, but not postsynaptic receptor-mediated responses: Relationship to antidepressant-like action. Eur. J. Pharmacol. 138, 53–60. [DOI] [PubMed] [Google Scholar]
- Blier P.; de Montigny C. (1987) Modification of 5-HT neuron properties by sustained administration of the 5-HT1A agonist gepirone: Electrophysiological studies in the rat brain. Synapse 1, 470–480. [DOI] [PubMed] [Google Scholar]
- Sprouse J. S.; Aghajanian G. K. (1988) Responses of hippocampal pyramidal cells to putative serotonin 5-HT1A and 5-HT1B agonists: A comparative study with dorsal raphe neurons. Neuropharmacology 27, 707–715. [DOI] [PubMed] [Google Scholar]
- Richer M.; Hen R.; Blier P. (2002) Modification of serotonin neuron properties in mice lacking 5-HT1A receptors. Eur. J. Pharmacol. 435, 195–203. [DOI] [PubMed] [Google Scholar]
- Parsons L. H.; Kerr T. M.; Tecott L. H. (2001) 5-HT(1A) receptor mutant mice exhibit enhanced tonic, stress-induced and fluoxetine-induced serotonergic neurotransmission. J. Neurochem. 77, 607–617. [DOI] [PubMed] [Google Scholar]
- Artigas F.; Adell A.; Celada P. (2006) Pindolol augmentation of antidepressant response. Curr. Drug Targets 7, 139–147. [DOI] [PubMed] [Google Scholar]
- Azmitia E. C.; Gannon P. J.; Kheck N. M.; Whitaker-Azmitia P. M. (1996) Cellular localization of the 5-HT1A receptor in primate brain neurons and glial cells. Neuropsychopharmacology 14, 35–46. [DOI] [PubMed] [Google Scholar]
- Cassel J. C.; Jeltsch H. (1995) Serotonergic modulation of cholinergic function in the central nervous system: Cognitive implications. Neuroscience 69, 1–41. [DOI] [PubMed] [Google Scholar]
- Palchaudhuri M.; Flugge G. (2005) 5-HT1A receptor expression in pyramidal neurons of cortical and limbic brain regions. Cell Tissue Res. 321, 159–172. [DOI] [PubMed] [Google Scholar]
- Santana N.; Bortolozzi A.; Serrats J.; Mengod G.; Artigas F. (2004) Expression of serotonin1A and serotonin2A receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb. Cortex 14, 1100–1109. [DOI] [PubMed] [Google Scholar]
- Hamon M.; Gozlan H.; el Mestikawy S.; Emerit M. B.; Bolanos F.; Schechter L. (1990) The central 5-HT1A receptors: Pharmacological, biochemical, functional, and regulatory properties. Ann. N.Y. Acad. Sci. 600, 114–129discussion 129–131.. [DOI] [PubMed] [Google Scholar]
- Beck S. G.; Choi K. C.; List T. J. (1992) Comparison of 5-hydroxytryptamine1A-mediated hyperpolarization in CA1 and CA3 hippocampal pyramidal cells. J. Pharmacol. Exp. Ther. 263, 350–359. [PubMed] [Google Scholar]
- Diaz-Mataix L.; Artigas F.; Celada P. (2006) Activation of pyramidal cells in rat medial prefrontal cortex projecting to ventral tegmental area by a 5-HT1A receptor agonist. Eur. Neuropsychopharmacol. 16, 288–296. [DOI] [PubMed] [Google Scholar]
- Diaz-Mataix L.; Scorza M. C.; Bortolozzi A.; Toth M.; Celada P.; Artigas F. (2005) Involvement of 5-HT1A receptors in prefrontal cortex in the modulation of dopaminergic activity: Role in atypical antipsychotic action. J. Neurosci. 25, 10831–10843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakaue M.; Somboonthum P.; Nishihara B.; Koyama Y.; Hashimoto H.; Baba A.; Matsuda T. (2000) Postsynaptic 5-hydroxytryptamine(1A) receptor activation increases in vivo dopamine release in rat prefrontal cortex. Br. J. Pharmacol. 129, 1028–1034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp T.; Boothman L.; Raley J.; Queree P. (2007) Important messages in the ’post’: recent discoveries in 5-HT neurone feedback control. Trends Pharmacol. Sci. 28, 629–636. [DOI] [PubMed] [Google Scholar]
- Celada P.; Puig M. V.; Casanovas J. M.; Guillazo G.; Artigas F. (2001) Control of dorsal raphe serotonergic neurons by the medial prefrontal cortex: Involvement of serotonin-1A, GABA(A), and glutamate receptors. J. Neurosci. 21, 9917–9929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajos M.; Hajos-Korcsok E.; Sharp T. (1999) Role of the medial prefrontal cortex in 5-HT1A receptor-induced inhibition of 5-HT neuronal activity in the rat. Br. J. Pharmacol. 126, 1741–1750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casanovas J. M.; Vilaro M. T.; Mengod G.; Artigas F. (1999) Differential regulation of somatodendritic serotonin 5-HT1A receptors by 2-week treatments with the selective agonists alnespirone (S-20499) and 8-hydroxy-2-(Di-n-propylamino)tetralin: Microdialysis and autoradiographic studies in rat brain. J. Neurochem. 72, 262–272. [DOI] [PubMed] [Google Scholar]
- Hajos M.; Richards C. D.; Szekely A. D.; Sharp T. (1998) An electrophysiological and neuroanatomical study of the medial prefrontal cortical projection to the midbrain raphe nuclei in the rat. Neuroscience 87, 95–108. [DOI] [PubMed] [Google Scholar]
- Jankowski M. P.; Sesack S. R. (2004) Prefrontal cortical projections to the rat dorsal raphe nucleus: Ultrastructural features and associations with serotonin and gamma-aminobutyric acid neurons. J. Comp. Neurol. 468, 518–529. [DOI] [PubMed] [Google Scholar]
- Stratz T.; Farber L.; Varga B.; Baumgartner C.; Haus U.; Muller W. (2001) Fibromyalgia treatment with intravenous tropisetron administration. Drugs Exp. Clin. Res. 27, 113–118. [PubMed] [Google Scholar]
- Varga V.; Szekely A. D.; Csillag A.; Sharp T.; Hajos M. (2001) Evidence for a role of GABA interneurones in the cortical modulation of midbrain 5-hydroxytryptamine neurones. Neuroscience 106, 783–792. [DOI] [PubMed] [Google Scholar]
- Borsini F.; Ceci A.; Bietti G.; Donetti A. (1995) BIMT 17, a 5-HT1A receptor agonist/5-HT2A receptor antagonist, directly activates postsynaptic 5-HT inhibitory responses in the rat cerebral cortex. Naunyn-Schmiedeberg's Arch. Pharmacol. 352, 283–290. [DOI] [PubMed] [Google Scholar]
- Amat J.; Baratta M. V.; Paul E.; Bland S. T.; Watkins L. R.; Maier S. F. (2005) Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat. Neurosci. 8, 365–371. [DOI] [PubMed] [Google Scholar]
- Aznar S.; Qian Z.; Shah R.; Rahbek B.; Knudsen G. M. (2003) The 5-HT1A serotonin receptor is located on calbindin- and parvalbumin-containing neurons in the rat brain. Brain Res. 959, 58–67. [DOI] [PubMed] [Google Scholar]
- De Vivo M.; Maayani S. (1986) Characterization of the 5-hydroxytryptamine1a receptor-mediated inhibition of forskolin-stimulated adenylate cyclase activity in guinea pig and rat hippocampal membranes. J. Pharmacol. Exp. Ther. 238, 248–253. [PubMed] [Google Scholar]
- Cooper D. M.; Londos C. (1982) GTP-dependent stimulation and inhibition of adenylate cyclase. Horiz. Biochem. Biophys. 6, 309–333. [PubMed] [Google Scholar]
- Albert P. R.; Robillard L. (2002) G protein specificity: Traffic direction required. Cell. Signalling 14, 407–418. [DOI] [PubMed] [Google Scholar]
- Barnes N. M.; Sharp T. (1999) A review of central 5-HT receptors and their function. Neuropharmacology 38, 1083–1152. [DOI] [PubMed] [Google Scholar]
- Haj-Dahmane S.; Hamon M.; Lanfumey L. (1991) K+ channel and 5-hydroxytryptamine1A autoreceptor interactions in the rat dorsal raphe nucleus: An in vitro electrophysiological study. Neuroscience 41, 495–505. [DOI] [PubMed] [Google Scholar]
- Lanfumey L.; Hamon M. (2000) Central 5-HT(1A) receptors: Regional distribution and functional characteristics. Nucl. Med. Biol. 27, 429–435. [DOI] [PubMed] [Google Scholar]
- Mannoury la Cour C.; El Mestikawy S.; Hanoun N.; Hamon M.; Lanfumey L. (2006) Regional differences in the coupling of 5-hydroxytryptamine-1A receptors to G proteins in the rat brain. Mol. Pharmacol. 70, 1013–1021. [DOI] [PubMed] [Google Scholar]
- Liu Y. F.; Ghahremani M. H.; Rasenick M. M.; Jakobs K. H.; Albert P. R. (1999) Stimulation of cAMP synthesis by Gi-coupled receptors upon ablation of distinct Galphai protein expression. Gi subtype specificity of the 5-HT1A receptor. J. Biol. Chem. 274, 16444–16450. [DOI] [PubMed] [Google Scholar]
- Marazziti D.; Palego L.; Giromella A.; Mazzoni M. R.; Borsini F.; Mayer N.; Naccarato A. G.; Lucacchini A.; Cassano G. B. (2002) Region-dependent effects of flibanserin and buspirone on adenylyl cyclase activity in the human brain. Int. J. Neuropsychopharmacol. 5, 131–140. [DOI] [PubMed] [Google Scholar]
- Palego L.; Giromella A.; Marazziti D.; Borsini F.; Naccarato A. G.; Giannaccini G.; Lucacchini A.; Cassano G. B.; Mazzoni M. R. (1999) Effects of postmortem delay on serotonin and (+)8-OH-DPAT-mediated inhibition of adenylyl cyclase activity in rat and human brain tissues. Brain Res. 816, 165–174. [DOI] [PubMed] [Google Scholar]
- Valdizan E. M.; Castro E.; Pazos A. (2010) Agonist-dependent modulation of G-protein coupling and transduction of 5-HT1A receptors in rat dorsal raphe nucleus. Int. J. Neuropsychopharmacol. 13, 835–843. [DOI] [PubMed] [Google Scholar]
- Clarke W. P.; Yocca F. D.; Maayani S. (1996) Lack of 5-hydroxytryptamine1A-mediated inhibition of adenylyl cyclase in dorsal raphe of male and female rats. J. Pharmacol. Exp. Ther. 277, 1259–1266. [PubMed] [Google Scholar]
- Johnson R. G.; Fiorella D.; Winter J. C.; Rabin R. A. (1997) [3H]8-OH-DPAT labels a 5-HT site coupled to inhibition of phosphoinositide hydrolysis in the dorsal raphe. Eur. J. Pharmacol. 329, 99–106. [DOI] [PubMed] [Google Scholar]
- Mannoury la Cour C.; Boni C.; Hanoun N.; Lesch K. P.; Hamon M.; Lanfumey L. (2001) Functional consequences of 5-HT transporter gene disruption on 5-HT(1a) receptor-mediated regulation of dorsal raphe and hippocampal cell activity. J. Neurosci. 21, 2178–2185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaput Y.; Blier P.; de Montigny C. (1986) In vivo electrophysiological evidence for the regulatory role of autoreceptors on serotonergic terminals. J. Neurosci. 6, 2796–2801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luscher C.; Jan L. Y.; Stoffel M.; Malenka R. C.; Nicoll R. A. (1997) G protein-coupled inwardly rectifying K+ channels (GIRKs) mediate postsynaptic but not presynaptic transmitter actions in hippocampal neurons. Neuron 19, 687–695. [DOI] [PubMed] [Google Scholar]
- Bayliss D. A.; Li Y. W.; Talley E. M. (1997) Effects of serotonin on caudal raphe neurons: Activation of an inwardly rectifying potassium conductance. J. Neurophysiol. 77, 1349–1361. [DOI] [PubMed] [Google Scholar]
- Clarke W. P.; De Vivo M.; Beck S. G.; Maayani S.; Goldfarb J. (1987) Serotonin decreases population spike amplitude in hippocampal cells through a pertussis toxin substrate. Brain Res. 410, 357–361. [DOI] [PubMed] [Google Scholar]
- Katayama J.; Yakushiji T.; Akaike N. (1997) Characterization of the K+ current mediated by 5-HT1A receptor in the acutely dissociated rat dorsal raphe neurons. Brain Res. 745, 283–292. [DOI] [PubMed] [Google Scholar]
- Kelly J. S.; Larkman P.; Penington N. J.; Rainnie D. G.; McAllister-Williams H.; Hodgkiss J. (1991) Serotonin receptor heterogeneity and the role of potassium channels in neuronal excitability. Adv. Exp. Med. Biol. 287, 177–191. [DOI] [PubMed] [Google Scholar]
- Loucif A. J.; Bonnavion P.; Macri B.; Golmard J. L.; Boni C.; Melfort M.; Leonard G.; Lesch K. P.; Adrien J.; Jacquin T. D. (2006) Gender-dependent regulation of G-protein-gated inwardly rectifying potassium current in dorsal raphe neurons in knock-out mice devoid of the 5-hydroxytryptamine transporter. J. Neurobiol. 66, 1475–1488. [DOI] [PubMed] [Google Scholar]
- Andrade R.; Nicoll R. A. (1987) Pharmacologically distinct actions of serotonin on single pyramidal neurones of the rat hippocampus recorded in vitro. J. Physiol. 394, 99–124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colino A.; Halliwell J. V. (1987) Differential modulation of three separate K-conductances in hippocampal CA1 neurons by serotonin. Nature 328, 73–77. [DOI] [PubMed] [Google Scholar]
- Oleskevich S. (1995) G alpha o1 decapeptide modulates the hippocampal 5-HT1A potassium current. J. Neurophysiol. 74, 2189–2193. [DOI] [PubMed] [Google Scholar]
- Kovoor A.; Lester H. A. (2002) Gi Irks GIRKs. Neuron 33, 6–8. [DOI] [PubMed] [Google Scholar]
- Kushwaha N.; Albert P. R. (2005) Coupling of 5-HT1A autoreceptors to inhibition of mitogen-activated protein kinase activation via G beta gamma subunit signaling. Eur. J. Neurosci. 21, 721–732. [DOI] [PubMed] [Google Scholar]
- Adayev T.; El-Sherif Y.; Barua M.; Penington N. J.; Banerjee P. (1999) Agonist stimulation of the serotonin1A receptor causes suppression of anoxia-induced apoptosis via mitogen-activated protein kinase in neuronal HN2–5 cells. J. Neurochem. 72, 1489–1496. [DOI] [PubMed] [Google Scholar]
- Albert P. R.; Tiberi M. (2001) Receptor signaling and structure: Insights from serotonin-1 receptors. Trends Endocrinol. Metab. 12, 453–460. [DOI] [PubMed] [Google Scholar]
- Chang C. W.; Poteet E.; Schetz J. A.; Gumus Z. H.; Weinstein H. (2009) Towards a quantitative representation of the cell signaling mechanisms of hallucinogens: Measurement and mathematical modeling of 5-HT1A and 5-HT2A receptor-mediated ERK1/2 activation. Neuropharmacology 56(Suppl 1), 213–225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowen D. S.; Johnson-Farley N. N.; Travkina T. (2005) 5-HT receptors couple to activation of Akt, but not extracellular-regulated kinase (ERK), in cultured hippocampal neurons. J. Neurochem. 93, 910–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Druse M.; Tajuddin N. F.; Gillespie R. A.; Le P. (2005) Signaling pathways involved with serotonin1A agonist-mediated neuroprotection against ethanol-induced apoptosis of fetal rhombencephalic neurons. Brain Res. 159, 18–28. [DOI] [PubMed] [Google Scholar]
- Newman-Tancredi A.; Martel J. C.; Assie M. B.; Buritova J.; Lauressergues E.; Cosi C.; Heusler P.; Bruins Slot L.; Colpaert F. C.; Vacher B.; Cussac D. (2009) Signal transduction and functional selectivity of F15599, a preferential post-synaptic 5-HT1A receptor agonist. Br. J. Pharmacol. 156, 338–353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsiung S. C.; Tamir H.; Franke T. F.; Liu K. P. (2005) Roles of extracellular signal-regulated kinase and Akt signaling in coordinating nuclear transcription factor-kappaB-dependent cell survival after serotonin 1A receptor activation. J. Neurochem. 95, 1653–1666. [DOI] [PubMed] [Google Scholar]
- Hsiung S. C.; Tin A.; Tamir H.; Franke T. F.; Liu K. P. (2008) Inhibition of 5-HT1A receptor-dependent cell survival by cAMP/protein kinase A: Role of protein phosphatase 2A and Bax. J. Neurosci. Res. 86, 2326–2338. [DOI] [PubMed] [Google Scholar]
- Chen S.; Owens G. C.; Crossin K. L.; Edelman D. B. (2007) Serotonin stimulates mitochondrial transport in hippocampal neurons. Mol. Cell Neurosci. 36, 472–483. [DOI] [PubMed] [Google Scholar]
- Talbot J. N.; Jutkiewicz E. M.; Graves S. M.; Clemans C. F.; Nicol M. R.; Mortensen R. M.; Huang X.; Neubig R. R.; Traynor J. R. (2010) RGS inhibition at G(alpha)i2 selectively potentiates 5-HT1A-mediated antidepressant effects. Proc. Natl. Acad. Sci. U.S.A. 107, 11086–11091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polter A. M.; Yang S.; Jope R. S.; Li X. (2012) Functional significance of glycogen synthase kinase-3 regulation by serotonin. Cell. Signalling 24, 265–271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauder J. M.; Wallace J. A.; Krebs H.; Petrusz P.; McCarthy K. (1982) In vivo and in vitro development of serotonergic neurons. Brain Res. Bull. 9, 605–625. [DOI] [PubMed] [Google Scholar]
- Oberlander T. F.; Gingrich J. A.; Ansorge M. S. (2009) Sustained neurobehavioral effects of exposure to SSRI antidepressants during development: Molecular to clinical evidence. Clin. Pharmacol. Ther. 86, 672–677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trowbridge S.; Narboux-Neme N.; Gaspar P. (2011) Genetic models of serotonin (5-HT) depletion: What do they tell us about the developmental role of 5-HT?. Anat. Rec. 294, 1615–1623. [DOI] [PubMed] [Google Scholar]
- van Kleef E. S.; Gaspar P.; Bonnin A. (2012) Insights into the complex influence of 5-HT signaling on thalamocortical axonal system development. Eur. J. Neurosci. 35, 1563–1572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hillion J.; Catelon J.; Raid M.; Hamon M.; De Vitry F. (1994) Neuronal localization of 5-HT1A receptor mRNA and protein in rat embryonic brain stem cultures. Brain. Res. 79, 195–202. [DOI] [PubMed] [Google Scholar]
- Hillion J.; Milne-Edwards J. B.; Catelon J.; de Vitry F.; Gros F.; Hamon M. (1993) Prenatal developmental expression of rat brain 5-HT1A receptor gene followed by PCR. Biochem. Biophys. Res. Commun. 191, 991–997. [DOI] [PubMed] [Google Scholar]
- Bonnin A.; Peng W.; Hewlett W.; Levitt P. (2006) Expression mapping of 5-HT1 serotonin receptor subtypes during fetal and early postnatal mouse forebrain development. Neuroscience 141, 781–794. [DOI] [PubMed] [Google Scholar]
- Daval G.; Verge D.; Becerril A.; Gozlan H.; Spampinato U.; Hamon M. (1987) Transient expression of 5-HT1A receptor binding sites in some areas of the rat CNS during postnatal development. Int. J. Dev. Neurosci. 5, 171–180. [DOI] [PubMed] [Google Scholar]
- Miquel M. C.; Kia H. K.; Boni C.; Doucet E.; Daval G.; Matthiessen L.; Hamon M.; Verge D. (1994) Postnatal development and localization of 5-HT1A receptor mRNA in rat forebrain and cerebellum. Brain Res. 80, 149–157. [DOI] [PubMed] [Google Scholar]
- Patel T. D.; Zhou F. C. (2005) Ontogeny of 5-HT1A receptor expression in the developing hippocampus. Brain Res. 157, 42–57. [DOI] [PubMed] [Google Scholar]
- Bar-Peled O.; Gross-Isseroff R.; Ben-Hur H.; Hoskins I.; Groner Y.; Biegon A. (1991) Fetal human brain exhibits a prenatal peak in the density of serotonin 5-HT1A receptors. Neurosci. Lett. 127, 173–176. [DOI] [PubMed] [Google Scholar]
- del Olmo E.; Lopez-Gimenez J. F.; Vilaro M. T.; Mengod G.; Palacios J. M.; Pazos A. (1998) Early localization of mRNA coding for 5-HT1A receptors in human brain during development. Brain Res. 60, 123–126. [DOI] [PubMed] [Google Scholar]
- del Olmo E.; Diaz A.; Guirao-Pineyro M.; del Arco C.; Pascual J.; Pazos A. (1994) Transient localization of 5-HT1A receptors in human cerebellum during development. Neurosci. Lett. 166, 149–152. [DOI] [PubMed] [Google Scholar]
- Beique J. C.; Campbell B.; Perring P.; Hamblin M. W.; Walker P.; Mladenovic L.; Andrade R. (2004) Serotonergic regulation of membrane potential in developing rat prefrontal cortex: Coordinated expression of 5-hydroxytryptamine (5-HT)1A, 5-HT2A, and 5-HT7 receptors. J. Neurosci. 24, 4807–4817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calizo L. H., Piel D. A., and Beck S. G. (2010) Uncovering critical periods in the development of 5-HT neuron physiology and excitatory and inhibitory synaptic activity, Society for Neuroscience, San Diego, CA. [Google Scholar]
- Mehta M.; Ahmed Z.; Fernando S. S.; Cano-Sanchez P.; Adayev T.; Ziemnicka D.; Wieraszko A.; Banerjee P. (2007) Plasticity of 5-HT 1A receptor-mediated signaling during early postnatal brain development. J. Neurochem. 101, 918–928. [DOI] [PubMed] [Google Scholar]
- Gardier A. M.; Malagie I.; Trillat A. C.; Jacquot C.; Artigas F. (1996) Role of 5-HT1A autoreceptors in the mechanism of action of serotoninergic antidepressant drugs: Recent findings from in vivo microdialysis studies. Fundam. Clin. Pharmacol. 10, 16–27. [DOI] [PubMed] [Google Scholar]
- Hervas I.; Queiroz C. M.; Adell A.; Artigas F. (2000) Role of uptake inhibition and autoreceptor activation in the control of 5-HT release in the frontal cortex and dorsal hippocampus of the rat. Br. J. Pharmacol. 130, 160–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hjorth S.; Westlin D.; Bengtsson H. J. (1997) WAY100635-induced augmentation of the 5-HT-elevating action of citalopram: Relative importance of the dose of the 5-HT1A (auto)receptor blocker versus that of the 5-HT reuptake inhibitor. Neuropharmacology 36, 461–465. [DOI] [PubMed] [Google Scholar]
- Popa D.; Cerdan J.; Reperant C.; Guiard B. P.; Guilloux J. P.; David D. J.; Gardier A. M. (2010) A longitudinal study of 5-HT outflow during chronic fluoxetine treatment using a new technique of chronic microdialysis in a highly emotional mouse strain. Eur. J. Pharmacol. 628, 83–90. [DOI] [PubMed] [Google Scholar]
- Trillat A. C.; Malagie I.; Mathe-Allainmat M.; Anmella M. C.; Jacquot C.; Langlois M.; Gardier A. M. (1998) Synergistic neurochemical and behavioral effects of fluoxetine and 5-HT1A receptor antagonists. Eur. J. Pharmacol. 357, 179–184. [DOI] [PubMed] [Google Scholar]
- Artigas F.; Romero L.; de Montigny C.; Blier P. (1996) Acceleration of the effect of selected antidepressant drugs in major depression by 5-HT1A antagonists. Trends Neurosci. 19, 378–383. [DOI] [PubMed] [Google Scholar]
- Blier P.; de Montigny C. (1983) Effects of quipazine on pre- and postsynaptic serotonin receptors: Single cell studies in the rat CNS. Neuropharmacology 22, 495–499. [DOI] [PubMed] [Google Scholar]
- Blier P.; De Montigny C. (1983) Electrophysiological investigations on the effect of repeated zimelidine administration on serotonergic neurotransmission in the rat. J. Neurosci. 3, 1270–1278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawson L. A.; Nguyen H. Q.; Smith D. I.; Schechter L. E. (2000) Effects of chronic fluoxetine treatment in the presence and absence of (±)pindolol: A microdialysis study. Br. J. Pharmacol. 130, 797–804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El Mansari M.; Sanchez C.; Chouvet G.; Renaud B.; Haddjeri N. (2005) Effects of acute and long-term administration of escitalopram and citalopram on serotonin neurotransmission: An in vivo electrophysiological study in rat brain. Neuropsychopharmacology 30, 1269–1277. [DOI] [PubMed] [Google Scholar]
- Luellen B. A., Gilman T. L., and Andrews A. M. (2010) Presynaptic adaptive responses to constitutive versus adult pharmacologic inhibition of serotonin uptake. In Experimental Models in Serotonin Transporter Research (Kalueff A. V. L., J.L., Ed.), Cambridge University Press, Cambridge. [Google Scholar]
- Artigas F.; Perez V.; Alvarez E. (1994) Pindolol induces a rapid improvement of depressed patients treated with serotonin reuptake inhibitors. Arch. Gen. Psychiatry 51, 248–251. [DOI] [PubMed] [Google Scholar]
- Portella M. J.; de Diego-Adelino J.; Ballesteros J.; Puigdemont D.; Oller S.; Santos B.; Alvarez E.; Artigas F.; Perez V. (2011) Can we really accelerate and enhance the selective serotonin reuptake inhibitor antidepressant effect? A randomized clinical trial and a meta-analysis of pindolol in nonresistant depression. J. Clin. Psychiatry 72, 962–969. [DOI] [PubMed] [Google Scholar]
- Ballesteros J.; Callado L. F. (2004) Effectiveness of pindolol plus serotonin uptake inhibitors in depression: a meta-analysis of early and late outcomes from randomised controlled trials. J. Affective Disord. 79, 137–147. [DOI] [PubMed] [Google Scholar]
- Whale R.; Terao T.; Cowen P.; Freemantle N.; Geddes J. (2010) Pindolol augmentation of serotonin reuptake inhibitors for the treatment of depressive disorder: a systematic review. J. Psychopharmacol. 24, 513–520. [DOI] [PubMed] [Google Scholar]
- Portella M. J.; de Diego-Adelino J.; Puigdemont D.; Perez-Egea R.; Alvarez E.; Artigas F.; Perez V. (2009) Pindolol augmentation enhances response outcomes in first depressive episodes. Eur. Neuropsychopharmacol. 19, 516–519. [DOI] [PubMed] [Google Scholar]
- Segrave R.; Nathan P. J. (2005) Pindolol augmentation of selective serotonin reuptake inhibitors: accounting for the variability of results of placebo-controlled double-blind studies in patients with major depression. Hum. Psychopharmacol. 20, 163–174. [DOI] [PubMed] [Google Scholar]
- Savitz J.; Lucki I.; Drevets W. C. (2009) 5-HT(1A) receptor function in major depressive disorder. Prog. Neurobiol. 88, 17–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi D. V.; Burke T. F.; McCasland M.; Hensler J. G. (2008) Serotonin-1A receptor function in the dorsal raphe nucleus following chronic administration of the selective serotonin reuptake inhibitor sertraline. J. Neurochem. 105, 1091–1099. [DOI] [PubMed] [Google Scholar]
- Pejchal T.; Foley M. A.; Kosofsky B. E.; Waeber C. (2002) Chronic fluoxetine treatment selectively uncouples raphe 5-HT(1A) receptors as measured by [(35)S]-GTP gamma S autoradiography. Br. J. Pharmacol. 135, 1115–1122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen C.; Li H.; Meller E. (2002) Repeated treatment with antidepressants differentially alters 5-HT1A agonist-stimulated [35S]GTP gamma S binding in rat brain regions. Neuropharmacology 42, 1031–1038. [DOI] [PubMed] [Google Scholar]
- Castro M.; Diaz A.; del Olmo E.; Pazos A. (2003) Chronic fluoxetine induces opposite changes in G protein coupling at pre and postsynaptic 5-HT1A receptors in rat brain. Neuropharmacology 44, 93–101. [DOI] [PubMed] [Google Scholar]
- Rozeske R. R.; Evans A. K.; Frank M. G.; Watkins L. R.; Lowry C. A.; Maier S. F. (2011) Uncontrollable, but not controllable, stress desensitizes 5-HT1A receptors in the dorsal raphe nucleus. J. Neurosci. 31, 14107–14115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bambico F. R.; Nguyen N. T.; Gobbi G. (2009) Decline in serotonergic firing activity and desensitization of 5-HT1A autoreceptors after chronic unpredictable stress. Eur. Neuropsychopharmacol. 19, 215–228. [DOI] [PubMed] [Google Scholar]
- Lanfumey L.; Pardon M. C.; Laaris N.; Joubert C.; Hanoun N.; Hamon M.; Cohen-Salmon C. (1999) 5-HT1A autoreceptor desensitization by chronic ultramild stress in mice. NeuroReport 10, 3369–3374. [DOI] [PubMed] [Google Scholar]
- Grippo A. J.; Sullivan N. R.; Damjanoska K. J.; Crane J. W.; Carrasco G. A.; Shi J.; Chen Z.; Garcia F.; Muma N. A.; Van de Kar L. D. (2005) Chronic mild stress induces behavioral and physiological changes, and may alter serotonin 1A receptor function, in male and cycling female rats. Psychopharmacology (Berlin, Ger.) 179, 769–780. [DOI] [PubMed] [Google Scholar]
- Evrard A.; Barden N.; Hamon M.; Adrien J. (2006) Glucocorticoid receptor-dependent desensitization of 5-HT1A autoreceptors by sleep deprivation: studies in GR-i transgenic mice. Sleep 29, 31–36. [DOI] [PubMed] [Google Scholar]
- Holmes A.; Lit Q.; Murphy D. L.; Gold E.; Crawley J. N. (2003) Abnormal anxiety-related behavior in serotonin transporter null mutant mice: the influence of genetic background. Genes Brain Behav. 2, 365–380. [DOI] [PubMed] [Google Scholar]
- Kalueff A. V.; Olivier J. D.; Nonkes L. J.; Homberg J. R. (2010) Conserved role for the serotonin transporter gene in rat and mouse neurobehavioral endophenotypes. Neurosci. Biobehav. Rev. 34, 373–386. [DOI] [PubMed] [Google Scholar]
- Li Q.; Wichems C.; Heils A.; Lesch K. P.; Murphy D. L. (2000) Reduction in the density and expression, but not G-protein coupling, of serotonin receptors (5-HT1A) in 5-HT transporter knock-out mice: gender and brain region differences. J. Neurosci. 20, 7888–7895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Q.; Wichems C.; Heils A.; Van De Kar L. D.; Lesch K. P.; Murphy D. L. (1999) Reduction of 5-hydroxytryptamine (5-HT)(1A)-mediated temperature and neuroendocrine responses and 5-HT(1A) binding sites in 5-HT transporter knockout mice. J. Pharmacol. Exp. Ther. 291, 999–1007. [PubMed] [Google Scholar]
- Heisler L. K.; Chu H. M.; Brennan T. J.; Danao J. A.; Bajwa P.; Parsons L. H.; Tecott L. H. (1998) Elevated anxiety and antidepressant-like responses in serotonin 5-HT1A receptor mutant mice. Proc. Natl. Acad. Sci. U.S.A. 95, 15049–15054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parks C. L.; Robinson P. S.; Sibille E.; Shenk T.; Toth M. (1998) Increased anxiety of mice lacking the serotonin1A receptor. Proc. Natl. Acad. Sci. U.S.A. 95, 10734–10739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramboz S.; Oosting R.; Amara D. A.; Kung H. F.; Blier P.; Mendelsohn M.; Mann J. J.; Brunner D.; Hen R. (1998) Serotonin receptor 1A knockout: An animal model of anxiety-related disorder. Proc. Natl. Acad. Sci. U.S.A. 95, 14476–14481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porsolt R. D.; Bertin A.; Blavet N.; Deniel M.; Jalfre M. (1979) Immobility induced by forced swimming in rats: Effects of agents which modify central catecholamine and serotonin activity. Eur. J. Pharmacol. 57, 201–210. [DOI] [PubMed] [Google Scholar]
- Groenink L.; van Bogaert M. J.; van der Gugten J.; Oosting R. S.; Olivier B. (2003) 5-HT1A receptor and 5-HT1B receptor knockout mice in stress and anxiety paradigms. Behav. Pharmacol. 14, 369–383. [DOI] [PubMed] [Google Scholar]
- Gross C.; Santarelli L.; Brunner D.; Zhuang X.; Hen R. (2000) Altered fear circuits in 5-HT(1A) receptor KO mice. Biol. Psychiatry 48, 1157–1163. [DOI] [PubMed] [Google Scholar]
- Klemenhagen K. C.; Gordon J. A.; David D. J.; Hen R.; Gross C. T. (2006) Increased fear response to contextual cues in mice lacking the 5-HT1A receptor. Neuropsychopharmacology 31, 101–111. [DOI] [PubMed] [Google Scholar]
- Tsetsenis T.; Ma X. H.; Lo Iacono L.; Beck S. G.; Gross C. (2007) Suppression of conditioning to ambiguous cues by pharmacogenetic inhibition of the dentate gyrus. Nat. Neurosci. 10, 896–902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarnyai Z.; Sibille E. L.; Pavlides C.; Fenster R. J.; McEwen B. S.; Toth M. (2000) Impaired hippocampal-dependent learning and functional abnormalities in the hippocampus in mice lacking serotonin(1A) receptors. Proc. Natl. Acad. Sci. U.S.A. 97, 14731–14736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolff M.; Costet P.; Gross C.; Hen R.; Segu L.; Buhot M. C. (2004) Age-dependent effects of serotonin-1A receptor gene deletion in spatial learning abilities in mice. Brain Res. 130, 39–48. [DOI] [PubMed] [Google Scholar]
- Kusserow H.; Davies B.; Hortnagl H.; Voigt I.; Stroh T.; Bert B.; Deng D. R.; Fink H.; Veh R. W.; Theuring F. (2004) Reduced anxiety-related behaviour in transgenic mice overexpressing serotonin 1A receptors. Brain Res. 129, 104–116. [DOI] [PubMed] [Google Scholar]
- Bert B.; Fink H.; Hortnagl H.; Veh R. W.; Davies B.; Theuring F.; Kusserow H. (2006) Mice over-expressing the 5-HT(1A) receptor in cortex and dentate gyrus display exaggerated locomotor and hypothermic response to 8-OH-DPAT. Behav. Brain Res. 167, 328–341. [DOI] [PubMed] [Google Scholar]
- Bert B.; Dere E.; Wilhelmi N.; Kusserow H.; Theuring F.; Huston J. P.; Fink H. (2005) Transient overexpression of the 5-HT1A receptor impairs water-maze but not hole-board performance. Neurobiol. Learn. Mem. 84, 57–68. [DOI] [PubMed] [Google Scholar]
- Bert B.; Fink H.; Rothe J.; Walstab J.; Bonisch H. (2008) Learning and memory in 5-HT(1A)-receptor mutant mice. Behav. Brain. Res. 195, 78–85. [DOI] [PubMed] [Google Scholar]
- Ogren S. O.; Eriksson T. M.; Elvander-Tottie E.; D’Addario C.; Ekstrom J. C.; Svenningsson P.; Meister B.; Kehr J.; Stiedl O. (2008) The role of 5-HT(1A) receptors in learning and memory. Behav. Brain. Res. 195, 54–77. [DOI] [PubMed] [Google Scholar]
- Richardson-Jones J. W.; Craige C. P.; Nguyen T. H.; Kung H. F.; Gardier A. M.; Dranovsky A.; David D. J.; Guiard B. P.; Beck S. G.; Hen R.; Leonardo E. D. (2011) Serotonin-1A autoreceptors are necessary and sufficient for the normal formation of circuits underlying innate anxiety. J. Neurosci. 31, 6008–6018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson-Jones J. W.; Craige C. P.; Guiard B. P.; Stephen A.; Metzger K. L.; Kung H. F.; Gardier A. M.; Dranovsky A.; David D. J.; Beck S. G.; Hen R.; Leonardo E. D. (2010) 5-HT1A autoreceptor levels determine vulnerability to stress and response to antidepressants. Neuron 65, 40–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blier P. (2010) Altered function of the serotonin 1A autoreceptor and the antidepressant response. Neuron 65, 1–2. [DOI] [PubMed] [Google Scholar]
- Lemonde S.; Du L.; Bakish D.; Hrdina P.; Albert P. R. (2004) Association of the C(−1019)G 5-HT1A functional promoter polymorphism with antidepressant response. Int J Neuropsychopharmacol 7, 501–506. [DOI] [PubMed] [Google Scholar]
- Lemonde S.; Turecki G.; Bakish D.; Du L.; Hrdina P. D.; Bown C. D.; Sequeira A.; Kushwaha N.; Morris S. J.; Basak A.; Ou X. M.; Albert P. R. (2003) Impaired repression at a 5-hydroxytryptamine 1A receptor gene polymorphism associated with major depression and suicide. J. Neurosci. 23, 8788–8799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rush A. J.; Warden D.; Wisniewski S. R.; Fava M.; Trivedi M. H.; Gaynes B. N.; Nierenberg A. A. (2009) STAR*D: Revising conventional wisdom. CNS Drugs 23, 627–647. [DOI] [PubMed] [Google Scholar]
- Bang S. J.; Jensen P.; Dymecki S. M.; Commons K. G. (2012) Projections and interconnections of genetically defined serotonin neurons in mice. Eur. J. Neurosci. 35, 85–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sperling R.; Commons K. G. (2011) Shifting topographic activation and 5-HT1A receptor-mediated inhibition of dorsal raphe serotonin neurons produced by nicotine exposure and withdrawal. Eur. J. Neurosci. 33, 1866–1875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waselus M.; Valentino R. J.; Van Bockstaele E. J. (2011) Collateralized dorsal raphe nucleus projections: a mechanism for the integration of diverse functions during stress. J. Chem. Neuroanat. 41, 266–280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen P.; Farago A. F.; Awatramani R. B.; Scott M. M.; Deneris E. S.; Dymecki S. M. (2008) Redefining the serotonergic system by genetic lineage. Nat. Neurosci. 11, 417–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vasudeva R. K.; Lin R. C.; Simpson K. L.; Waterhouse B. D. (2011) Functional organization of the dorsal raphe efferent system with special consideration of nitrergic cell groups. J. Chem. Neuroanat. 41, 281–293. [DOI] [PubMed] [Google Scholar]
- Adell A.; Casanovas J. M.; Artigas F. (1997) Comparative study in the rat of the actions of different types of stress on the release of 5-HT in raphe nuclei and forebrain areas. Neuropharmacology 36, 735–741. [DOI] [PubMed] [Google Scholar]
- Li Y. Q.; Li H.; Kaneko T.; Mizuno N. (2001) Morphological features and electrophysiological properties of serotonergic and non-serotonergic projection neurons in the dorsal raphe nucleus. An intracellular recording and labeling study in rat brain slices. Brain Res. 900, 110–118. [DOI] [PubMed] [Google Scholar]
- Gaspar P.; Lillesaar C. (2012) Probing the diversity of serotonin neurons. Philos. Trans. R. Soc. London, Ser. B 367, 2382–2394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiyasova V., Bonnavion P., Scotto-Lomassese S., Fabre V., Sahly I., Toronche F., Deneris E., Gaspar P., and Fernandez S. P. (2012) A subpopulation of serotonergic neurons that do not express the 5-HT1A autoreceptor. ACS Chem Neurosci. DOI: 10.1021/cn300157s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Assie M. B.; Bardin L.; Auclair A. L.; Carilla-Durand E.; Depoortere R.; Koek W.; Kleven M. S.; Colpaert F.; Vacher B.; Newman-Tancredi A. (2010) F15599, a highly selective post-synaptic 5-HT(1A) receptor agonist: In-vivo profile in behavioural models of antidepressant and serotonergic activity. Int. J. Neuropsychopharmacol. 13, 1285–1298. [DOI] [PubMed] [Google Scholar]
- Blier P.; Bergeron R.; de Montigny C. (1997) Selective activation of postsynaptic 5-HT1A receptors induces rapid antidepressant response. Neuropsychopharmacology 16, 333–338. [DOI] [PubMed] [Google Scholar]
- Haddjeri N.; Blier P.; de Montigny C. (1998) Long-term antidepressant treatments result in a tonic activation of forebrain 5-HT1A receptors. J. Neurosci. 18, 10150–10156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haddjeri N.; de Montigny C.; Blier P. (1999) Modulation of the firing activity of rat serotonin and noradrenaline neurons by (±)pindolol. Biol. Psychiatry 45, 1163–1169. [DOI] [PubMed] [Google Scholar]
- Haddjeri N.; Ortemann C.; de Montigny C.; Blier P. (1999) Effect of sustained administration of the 5-HT1A receptor agonist flesinoxan on rat 5-HT neurotransmission. Eur. Neuropsychopharmacol. 9, 427–440. [DOI] [PubMed] [Google Scholar]
- Bortolozzi A.; Castane A.; Semakova J.; Santana N.; Alvarado G.; Cortes R.; Ferres-Coy A.; Fernandez G.; Carmona M. C.; Toth M.; Perales J. C.; Montefeltro A.; Artigas F. (2012) Selective siRNA-mediated suppression of 5-HT1A autoreceptors evokes strong anti-depressant-like effects. Mol Psychiatry 17, 612–623. [DOI] [PubMed] [Google Scholar]
- Ferres-Coy A.; Santana N.; Castane A.; Cortes R.; Carmona M. C.; Toth M.; Montefeltro A.; Artigas F.; Bortolozzi A.. Acute 5-HT(1A) autoreceptor knockdown increases antidepressant responses and serotonin release in stressful conditions. Psychopharmacology (Berlin, Ger.) 2012, not supplied. [DOI] [PubMed]
- Gross C.; Zhuang X.; Stark K.; Ramboz S.; Oosting R.; Kirby L.; Santarelli L.; Beck S.; Hen R. (2002) Serotonin1A receptor acts during development to establish normal anxiety-like behaviour in the adult. Nature 416, 396–400. [DOI] [PubMed] [Google Scholar]
- Lo Iacono L.; Gross C. (2008) Alpha-Ca2+/calmodulin-dependent protein kinase II contributes to the developmental programming of anxiety in serotonin receptor 1A knock-out mice. J. Neurosci. 28, 6250–6257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sari Y. (2004) Serotonin1B receptors: from protein to physiological function and behavior. Neurosci. Biobehav. Rev. 28, 565–582. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.