

Abstract
Pathogenesis studies of SIV infection have not been performed to date in wild monkeys due to difficulty in collecting and storing samples on site and the lack of analytical reagents covering the extensive SIV diversity. We performed a large scale study of molecular epidemiology and natural history of SIVagm infection in 225 free-ranging AGMs from multiple locations in South Africa. SIV prevalence (established by sequencing pol, env, and gag) varied dramatically between infant/juvenile (7%) and adult animals (68%) (p<0.0001), and between adult females (78%) and males (57%). Phylogenetic analyses revealed an extensive genetic diversity, including frequent recombination events. Some AGMs harbored epidemiologically linked viruses. Viruses infecting AGMs in the Free State, which are separated from those on the coastal side by the Drakensberg Mountains, formed a separate cluster in the phylogenetic trees; this observation supports a long standing presence of SIV in AGMs, at least from the time of their speciation to their Plio-Pleistocene migration. Specific primers/probes were synthesized based on the pol sequence data and viral loads (VLs) were quantified. VLs were of 104–106 RNA copies/ml, in the range of those observed in experimentally-infected monkeys, validating the experimental approaches in natural hosts. VLs were significantly higher (107–108 RNA copies/ml) in 10 AGMs diagnosed as acutely infected based on SIV seronegativity (Fiebig II), which suggests a very active transmission of SIVagm in the wild. Neither cytokine levels (as biomarkers of immune activation) nor sCD14 levels (a biomarker of microbial translocation) were different between SIV-infected and SIV-uninfected monkeys. This complex algorithm combining sequencing and phylogeny, VL quantification, serology, and testing of surrogate markers of microbial translocation and immune activation permits a systematic investigation of the epidemiology, viral diversity and natural history of SIV infection in wild African natural hosts.
Author Summary
We simultaneously assessed, for the first time in a natural host, the epidemiology, diversity and natural history of SIVagmVer infection in wild vervet populations from South Africa. We report that African green monkeys (AGMs) have likely been infected with SIVagm for a long period, ranging from the time of their speciation to Plio-Pleistocene migrations, refuting previous molecular clock calculations suggesting SIVagm to be of recent occurrence. As a result of virus-host coadaptation, SIVagmVer infection is characterized by a lack of disease progression in spite of robust viral replication. We show that very active SIVagm transmission in adult AGMs contrasts with a very limited transmission to their offspring, in spite of massive exposure to SIVagm both in utero and through breastfeeding. The observation that some AGMs remain uninfected in spite of life-long exposure to SIVagm identifies wild vervets as an acceptable animal model for the exposed uninfected individuals, which can be used to identify correlates of resistance to HIV/SIV infection.
Introduction
Over 40 African nonhuman primate (NHP) species are naturally infected with simian immunodeficiency viruses (SIVs) [1]–[3]. Among these, African green monkeys (AGMs) (Chlorocebus genus) are the most numerous, most widely geographically spread and most commonly infected with SIV in the wild [1].
According to Groves [4], [5], AGMs are divided into species that are phenotypically and geographically distinct: vervets (C. pygerythrus) are the most widely spread in East and Southern Africa, ranging from the eastern Rift Valley in Ethiopia, Somalia and extreme southern Sudan to the Cape region in South Africa; grivets (C. aethiops) inhabit the area east of the White Nile in Ethiopia, Somalia from Khartoum to Mongalla, Eritrea and Ethiopia south to the Omo; tantalus monkeys (C. tantalus) are the prevalent species in north Central Africa from Ghana to Sudan and Kenya; green monkeys (C. sabaeus) reside in West Africa from Senegal to the Volta River, and have been introduced to Cape Verde Island as well as to the Caribbean [4]. A fifth AGM species, the malbrouck (C. cynosuros), is located in Central and South-Central Africa ranging from the Albertine Rift in the Democratic Republic of Congo to the Atlantic coast and Northern Namibia and Zambia. Finally, a sixth species, the Bale Mountain vervet (C. djamdjamensis) has a very limited distribution in the bamboo forests of highland Ethiopia. This taxonomic classification, however, is not universally accepted [6].
With the exception of the malbrouck and the Bale mountain vervet, each of the AGM species have been shown to be infected in the wild with species-specific SIVagm (defined as SIVagmVer, SIVagmGri, SIVagmTan and SIVagmSab) [7]–[11], with the reported prevalence rates ranging from30 to 50% [8], [12]–[19]. Phylogenetic analyses of SIVagm strains have shown that the spectrum of genetic diversity of SIVagm surpasses that of other primate lentiviruses [7], [9], [19]–[21], presumed to be due to codivergence/host-dependent evolution (i.e., the evolution of SIVagm from grivet, vervet, tantalus and green monkey species occurred in concert with host speciation) [8], [10], [11]. Together, these findings indicate that AGMs have coexisted with SIVagm for a long period of time, possibly even before their radiation and divergence from their common ancestor, approximately 1.5–3 million years ago [22].
Pathogenesis studies support an ancient coevolutionary relationship by showing that SIVagm are highly adapted to their hosts and that SIVagm-infected AGMs generally do not progress to AIDS [23], [24]. Key features of SIVagm infection in AGMs, as established by experimental studies performed on captive AGMs, include: (i) active viral replication, with set-point viral loads (VLs) similar to or higher than those found in HIV-infected patients [25]–[29]; (ii) significant depletion of CD4+ T cells during acute infection [25], [26], [30], [31], followed by rapid restoration to near preinfection levels in the peripheral blood [17], [25], [26], [30], [31] and delayed and incomplete restoration at mucosal sites [30]; (iii) maintenance of the balance between Th17 and T regulatory cells, due to preservation of the Th17 cell subset [32]; (iv) vigorous but transient inflammatory responses to the virus during acute infection, which are resolved with the transition from acute-to-chronic infection [30], [33], [34]; (v) productive infection of short-lived cells [31]; (vi) partial control of virus replication by the adaptive immune responses to SIV [35]–[41];(vii) no significant increase in CD4+ T cell apoptosis during either acute or chronic infection [30], [42], [43]. These normal levels of CD4+ T cell apoptosis together with preservation of Th17 cells probably allow AGMs to avoid enteropathy, breaches in the mucosal barrier and subsequent microbial translocation (MT) [30], as well as chronic immune activation and disease progression, while allowing CD4+ T cell recovery in the presence of high VLs [30], [44]. Furthermore, similar to other natural hosts of SIVs [45], AGMs have a number of adaptations that spare CD4+ T cells from virus-mediated killing in vivo. These features include a lower fraction of CD4+ T cells expressing the CCR5 coreceptor [46], [47] and down-regulation of the CD4 molecule by T helper cells as they enter the memory cell pool [48]–[50]. Altogether, these adaptations support the concept that the benign course of SIV infection in natural hosts is the result of coevolution over millennia [1], [51].
The ancestry of SIV infections and the codivergence of SIV types with their host species, however, could be accounted for by a model of preferential host-switching [52]. In the case of AGMs it has recently suggested that the phylogenetic profiles of SIVagm can be explained by a pattern of west-to-east transmission of the virus across existing AGM geographic ranges and that the viral divergence may have occurred more recently than previously suggested [53]. This study included only a small number of SIV strains covering a limited geographical distribution, which may have biased a full appreciation of the phylogenetic relationships and ancestry of SIVagm from different AGM species and likely altered the timing calculations for SIVagm emergence. A recent study in which calibration of molecular clocks was based on biogeographical features, reported that SIVs are actually older than estimated based on molecular clock calculations only [2].
The major focus of studies carried out thus far in wild African NHP species has been on identification and characterization of species-specific SIVs [54], [55]. A common denominator of these studies has been the difficulty of obtaining samples from wild animals and the lack of research infrastructure for pathogenesis studies in the field. Several approaches have been used to circumvent these limitations: (i) estimation of the SIV prevalence in captive monkeys, which significantly underestimates the prevalence in the wild, as most monkeys are captured at a young age and maternal-to-infant transmission rates are low in African NHP species [56]; (ii) use of bush meat samples. These are easy to obtain and thus this strategy has proven instrumental in identifying numerous SIVs and generating meaningful prevalence data [2], [57]–[62]. However, bush meat samples tend to be from adult populations and do not permit viral isolation or assessment of critical parameters of infection, such as VLs, immunophenotyping or assessment of the levels of immune activation; and (iii) use of noninvasive sampling (i.e., collection of fecal or urine samples), which has also been instrumental in assessing the prevalence of SIV infection in natural hosts, most notably in identifying and characterizing the animal reservoir of pandemic HIV strains [63]–[67]. However, similar to the use of bush meat samples, noninvasive samples cannot be used to assess key parameters of SIV infection. Thus, although SIVs have been identified and characterized for over two decades, studies of the natural history of SIV infection in the wild have only been performed at a cursory level [1], [54], [68].
Here, we performed for the first time in an African NHP host, a comprehensive large-scale characterization of the epidemiology and natural history of SIV infection in vervet monkeys living freely in different parks, nature reserves and farms from three provinces of the Republic of South Africa, to validate the virological and immunological features of experimental studies in natural hosts.
Results
Prevalence of retroviral infections in vervets from South Africa
Blood samples were collected from two hundred and twenty-five wild-trapped vervets over a 4.5 month period. Details on monkey capture and sample collection are presented as Supplementary information. The samples included represent 11 populations from 9 parks, game reserves and farms located in 3 provinces of South Africa–the Free State, KwaZulu-Natal, and Eastern Cape–covering majority significant proportion of vervet habitat in this region (Figure 1). Both sexes were equally represented. Samples from infants (<1 year old) and juvenile AGMs were included (Figure 1).
Figure 1. Satellite map of the Republic of South Africa indicating the geographical origin of the samples included in this study.

Vervet samples from areas from three provinces (Free State, KwaZulu Natal and Eastern Cape) were included, as indicated. Sex and age-group coverage is illustrated.
Diagnosis of SIVagmVer infection was performed by PCR, which allows for the evaluation of SIVagm diversity in addition to prevalence estimates. Also, in infants, PCR permits the diagnosis of infected offspring (as opposed to serological assays which are not specific, i.e., may also detect passively transmitted maternal antibodies). RNA was extracted from all plasma samples and subjected to RT-PCR analysis using consensus primers designed to amplify a 600-bp fragment in pol integrase and a 900-bp fragment in the gp120 gene encompassing the V3–V5 regions and the 5′ end of the gp41 gene. In addition, gag PCR (846 bp fragment in the p24 gene) was performed on selected samples from each location. This analysis identified 103 AGMs that were virion RNA (vRNA) positive using one, two or three primer sets (Table 1), giving an overall prevalence of SIVagmVer infection of 59% (73/123) in females and 29% (30/102) in males, in the range of those found in previous reports in AGMs [8], [15], [17]. Prevalence levels were relatively similar between different locations (Table S1) and higher than in a cohort of semifree animals (Table S2). SIVagm prevalence was very uneven in different age groups: 7% (3/44) in infants (males: 4%, 1/26; females: 11%, 2/18), 16% (9/58) in juveniles (males: 15%, 5/34; females: 21%, 4/21) and 71% (91/128) in adults (males: 57%, 24/42; females: 78%, 67/86). Thus, we confirmed on a very large number of samples previous results reporting that a dramatic increase in the SIVagm prevalence and incidence occurs with the passage to sexual maturity in AGMs [16]. Interestingly, and different from previous reports, we identified cases of SIVagmVer infection in very young AGMs, which may be suggestive of vertical transmission of the virus in the wild.
Table 1. Age- and sex-related prevalence of SIVagmVer in wild vervets from South Africa.
| Females | Males | |||||
| Total | SIV+ | Prevalence (%) | Total | SIV+ | Prevalence (%) | |
| Infants | 18 | 2 | 11 | 26 | 1 | 4 |
| Juveniles | 19 | 4 | 21 | 34 | 5 | 15 |
| Adults | 86 | 67 | 78 | 42 | 24 | 57 |
Genetic diversity of SIVagm in vervets from South Africa
To determine the relationships of the newly identified SIVagm strains to each other and to previously characterized SIVagm strains, we constructed phylogenetic trees from amplified pol and env nucleotide sequences using a maximum likelihood approach (Figure 2 and Figure S1). While newly identified SIVagmVer strains naturally infecting vervets from South Africa exhibited a high genetic diversity, with average genetic distances in the pol gene of 16.2±4.8%, phylogenetic analyses also identified SIVagm strains that differed in less than 2% of their pol and env nucleotide sequences, indicating epidemiologically linked infections (Figure 2 and Figure S1). In general, strains originating from vervets from the same area clustered together (Figure 2) with a few exceptions that are probably due to male migration between groups when they reach sexual maturity.
Figure 2. SIVagmVer diversity in wild AGMs from different parks and game reserves in the Republic of South Africa.

Maximum likelihood tree for pol gene. Sequences are colored depending on the region in which the sequences were sampled, with red indicating sequences from Free State, green indicating sequences from KwaZulu-Natal, and blue indicating sequences from Eastern Cape territories. Reference SIVagm strains from vervet monkeys in Kenya are shown in black. The tree is based on a 592 bp pol sequence alignment (after gap-containing sites were removed). Maximum likelihood estimates were performed using 1000 replicate bootstrap analysis. Internal nodes indicate level of support values for internal branching. Bar represents number of amino acid replacements (0.09) per site. The strain nomenclature includes the identification number, monkey status (M-male, F-female; I-infant; J-juvenile and A-adult),and the site and state of origin (FS-Free State; KZN-KwaZulu Natal; EC-Eastern Cape). Putative heterosexual transmission cases documented to be of recent occurrence based on VL and serology testing are underlined. Putative maternal-to-infant transmission cases are boxed.
As expected, phylogenetic analyses showed that SIVagmVer strains from South Africa clustered with the SIVagmVer reference strains (Figure 2 and Figure S1) and were only distantly related to the SIVagm strains from other AGM species (Figure 2). Furthermore, the newly characterized SIVagmVer strains from South Africa formed a subcluster within the SIVagmVer group, in agreement with the fact that all the reference strains available thus far have a distinct geographical origin, being collected from vervets in Kenya, Ethiopia and Tanzania [9], [19], [20]. Average genetic distances between the reference SIVagmVer strains (collected from Ethiopia, Kenya and Tanzania) and the newly characterized SIVagmVer strains were 25.4±2.1%, significantly higher than the overall genetic distances between South African strains (p<0.0001) (Table S3).
Similar to previous reports [66], comparison between the pol and env tree topologies showed numerous differences (Figure 2 and Figure S1), indicating extensive recombination within the SIVagm radiation (Figure S2). Mosaic branches occurred both near the tips of the trees and deeper in the trees, indicating that extensive recombination events have occurred throughout the evolutionary history of SIVagmVer [66]. This high frequency of recombinations between divergent SIVs indicates that SIVagm coinfection and/or superinfection occur frequently in wild vervets.
Remarkably, a geographically-associated clustering pattern can be defined for the South African SIVagmVer strains, with all but one of the SIVagmVer sequences from AGMs inhabiting different areas in the Free State forming a monophyletic cluster in the phylogenetic trees (Figure 2 and Figure S1).
Bootstrap analysis strongly supports a single origin of the Free State cluster, however parental nodes were associated with lower bootstrap support in the pol tree, suggesting ambiguity as to where the root of the Free State cluster should be placed. Lower bootstrap support may be indicative of recombination; a SplitsTree analysis shows that some KwaZulu-Natal sequences are likely to be recombinants of East Coast sequences, indicating a mixing between these two populations (Figure S2).
Interestingly, the SplitsTree analysis also identified one Free State sequence (VMT17083-IM-Gariep_FS) as potentially stemming from a distant recombination event with outgroup SIVagmVer sequences, this may suggest that this sequence is in fact a recombinant with an unknown parent (Figure S2).
The most likely explanation for these phylogenetic relationships is that the Drakensberg Mountains acted as a barrier to separate the vervet populations. We estimate that this separation could have occurred in a time frame ranging from 3 million years ago (during the AGM spread throughout sub-Saharan Africa) to 100,000 years ago (during the mass migrations that occurred in the Plio-Pleistocene glacial periods). We tested this hypothesis by calibrating a relaxed molecular clock to specify the time to the most recent ancestor (TMRCA) of the SIVagm samples surrounding the Drakensberg Mountains range to a time-frame spanning 100,000 to 3,000,000 years since the present for the env tree, and in the pol tree we additionally included SIV strains from several species and further calibrated the tree based on isolates from Bioko Island [2]. According to this analysis, divergence of the Free State lineage is likely to be closer to the 100,000 year mark, with mean most recent common ancestor (MRCA) for the South African SIVagm strains estimated at 329 thousands of years (kYa) (with 95% highest posterior density of 100–1,077 kYa) and 104 kYa (100–112 kYa), in the env tree (Table S4) and pol tree (Figure 3 and Table S4), respectively. This analysis places the root of the SIV tree older than previous studies [2], with the env tree giving a time estimate of SIV endemicity in simian species for 790 kYa (range: 151–2,595 kYa), and the pol tree indicating 248 kYa (range: 189–317 kYa) (Table S4).
Figure 3. Molecular clock trees for pol gene based on the newly derived SIVagmVer sequences from vervets in South Africa.

Relaxed molecular clock tree show the ultrametric divergence pattern between lineages. Calibration was based on internal nodes indicated with a red circle, by setting priors that span the presumed divergence MRCA date ranges, 100,000–3,000,000 years for SIVagm sequences surrounding the Drakensberg Mountains in South Africa, and 10,000±1,000 for SIVdrl sequences sampled from Bioko Island [2]. Internal nodes show divergence date estimates. Sequences are colored based on the territory from which they were sampled, with South African sequences colored red, green and blue, for Free State, KwaZulu Natal and Eastern Cape, respectively, and Bioko sequences colored in yellow.
Analyses of the env gene
SIVagmVer envelope sequences (909 bp) encompassing the V3–V5 regions of the surface (SU) envelope glycoprotein and the 5′ end of the transmembrane (TM) domain were amplified by nested PCR from 48 plasma samples. Alignment of their deduced amino acid sequences with those of published reference SIVagm strains from all four species of AGMs identified hypervariable (V3–V5) as well as conserved envelope domains (Figure 4), as reported previously [10].
Figure 4. Alignment of partial envelope protein sequences from multiple SIVagmVer isolates.

PCR-derived env nucleotide sequences were translated, aligned with previously reported SIVAGM env sequences [133] and compared with a consensus sequence generated by BioEdit. Dots denote sequence identity with the consensus sequence, while dashes represent gaps introduced to optimize the alignment. Triangles (∧) denote N-linked glycosylation sites. Bullets (•) mark CD4 binding sites. V3, V4 and V5 designate hypervariable SIVagm Env domains as previously described [10]. The envelope precursor cleavage site is indicated by an arrow above the consensus sequence. Strain nomenclature includes the identification number, monkey status (M-male, F-female; I-infant; J-juvenile and A-adult), and the site and the state of origin (FS-Free State; KZN-KwaZulu Natal; EC-Eastern Cape).
Regions corresponding to HIV-1 envelope domains of known function, including the envelope glycoprotein precursor cleavage site, the CD4 binding domain and the viral fusion peptide (N-terminus of gp41) were all highly conserved (Figure 4). Also conserved were the majority of cysteine residues (11 of 12) and, as reported previously [10], [18], there was very little sequence variability in the region that corresponds to the hypervariable V3 loop of HIV-1. Pairwise amino acid sequence comparisons were performed to examine the extent of SIVagmVer genetic diversity in the PCR amplified envelope fragment. As for the pol sequence analysis, env analysis revealed that SIVagmVer sequences collected from monkeys in the Free State form a distinct cluster from the monkeys living on coastal areas from both KwaZulu Natal and Eastern Cape provinces (Figure S1). This separate cluster was supported by both specific sequence signatures in the Free State-originating strains and by smaller genetic distances between these strains (Figure 4 and Figure S1).
Thus, sequence analysis identified: (i) a high diversity of SIVagm in South Africa, with the viruses collected from AGMs in the Free State forming an independent cluster, clearly separated from the strains collected from AGMs on the Indian Ocean coast; (ii) high frequency of recombination, suggestive of extensive transmission of the virus; and (iii) occurrence of highly related strains, suggestive of identifiable transmission clusters.
Assessment of the natural history of SIVagmVer infection in wild AGMs
It is generally assumed that SIV infection is nonpathogenic in natural hosts [23], [51], [56]. However, this paradigm was recently challenged by studies demonstrating that SIVcpz has a substantial negative impact on the health, reproduction and lifespan of chimpanzees in the wild [68]–[70]. We therefore took advantage of this unique sample set to assess for the first time the natural history of SIV infection in a natural host monkey species in the wild.
Clinical data
During sample collection, all included AGMs had a thorough clinical assessment. None of the vervets presented with any of the clinical signs associated with SIV infection, including fever, weight loss, lymphadenopathy or opportunistic infections. Although the cross-sectional nature of this study precludes assessment of changes in monkey weight and thus a direct assessment of weight loss, we performed comparative analyses of the body mass index (BMI) between SIV-positive and SIV-negative monkeys and showed that SIV status does not have an impact on normal weight of wild vervets (Figure S3).
Genotypic assessment of the coreceptor usage of SIVagmVer strains from South Africa
Coreceptor usage by the different SIV strains may have a significant impact on the natural history of SIV infection in the wild. Previous studies reported that SIVagmSab strains have the ability to use the CXCR4 coreceptor for viral entry, in addition to CCR5 [26]. Since the amounts of plasma available from the AGMs in South Africa precluded in vitro phenotypic testing of coreceptor usage by these new strains, we estimated coreceptor use in silico by analyzing the V3 region. V3 sequence impacts SIVmac and SIVagm tropism [71]–[73].The algorithms for genotypic assessment of coreceptor usage based on the HIV-1 V3 sequences cannot be implemented for SIV [74], [75]. We therefore assessed the overall net charges of the V3 loop for these sequences and found that all the viruses exhibited the same V3 loop net charge (+7), similar to that of other SIV strains that are documented to use CCR5 [1]. No significant differences in the potential N-glycosylation sites located in the V3–V5 region could be documented for the newly derived SIVagmVer strains from South Africa (Figure 4). We therefore inferred that SIVagmVer strains identified here likely use CCR5 as the major coreceptor for entry, similar to the majority of SIV isolates [26], [40], [76].
Staging of SIVagmVer infection in the wild: VL and serological testing
The levels of chronic viral replication are highly predictive of the outcome of HIV-1 infection [77], [78] and as we previously reported, may also predict the outcome of SIV infection in natural hosts, as the handful of African NHPs reported to progress to AIDS exhibited higher chronic VLs than nonprogressors [79]. To date, VLs in natural hosts of SIVs such as AGMs, sooty mangabeys and mandrills have only been assessed in either experimentally intravenously-infected monkeys [25], [80]–[84] or captive naturally infected monkeys [17], [28], [29], [79], [85], [86]. Only one study reported VLs in wild African monkeys on a very limited number of samples [85].
We used the generated SIVagmVer pol sequences to design specific primers and probes for viral quantification. The integrase region amplified by the pol primers used in our study is relatively well conserved; thus, the primers and probe had an excellent coverage of SIVagmVer diversity. We then quantified the VLs in 87 available samples from the 103 SIVagmVer-infected vervets. Results are shown in Figure 5. Plasma SIVagmVer VLs ranged from 104 to 107 copies/ml and were higher in the juvenile than in adult AGMs, albeit this difference was not statistically significant.
Figure 5. Cross-sectional analysis of SIVagm RNA levels in plasma of naturally infected AGM using a real-time RT-PCR assay.
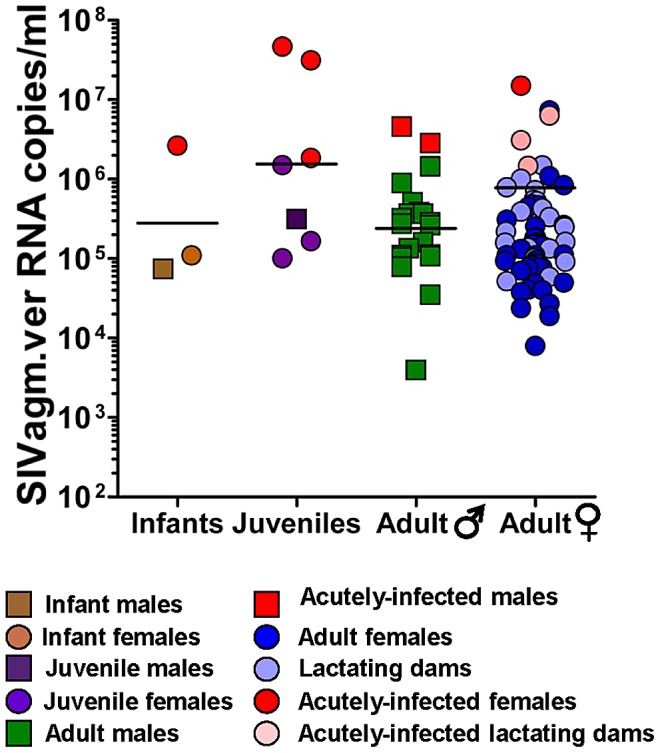
No significant difference could be observed between infant, juvenile and adult AGM or between female and male monkeys. Samples with high VLs assessed as acutely infected are shown. VLs of lactating dams are depicted in lighter colors. Detection limit: 100 copies/ml.
At least 23 of the 49 SIVagm-infected dams for which the VLs were available (47%) were documented as lactating at the time of sampling (Figure 5). The VLs of lactating dams are detailed in Figure 5 and demonstrate a large exposure of infant AGMs to SIV through breastfeeding.
While VLs of the majority of SIVagm-infected animals fell within a relatively close range, consistent with the range previously reported in experimentally infected African NHPs that are natural hosts of SIV [27], VLs were higher than expected in approximately 10% of vervets (Figure 5). Since such high VLs were observed in the majority of juvenile AGMs, we reasoned that they may be recent infections [27].
High levels of viral replication observed around the peak of VLs during acute HIV/SIV infection generally occur prior to seroconversion [87] and correspond to the Fiebeg II stage of HIV-1 infection [87]. Hence, to stage the SIVagm infection in AGMs with high VLs, we further designed a gp41 peptide ELISA and assessed the levels of anti-SIVagm antibodies in the available samples. As shown in Figure 6, the majority of SIVagm-infected AGMs harbored detectable levels of anti-gp41 antibodies. Conversely, most of the AGMs presenting with high VLs were seronegative. Altogether, these results strongly suggest that AGMs with high VLs were in the acute stage of infection [87]. Note, however, that definitive proof in support of this conclusion would rely on serial sampling and demonstration of seroconversion and partial control of viral replication [87].
Figure 6. anti-SIVagmVer IgG antibody seroprevalence in vervets from South Africa.
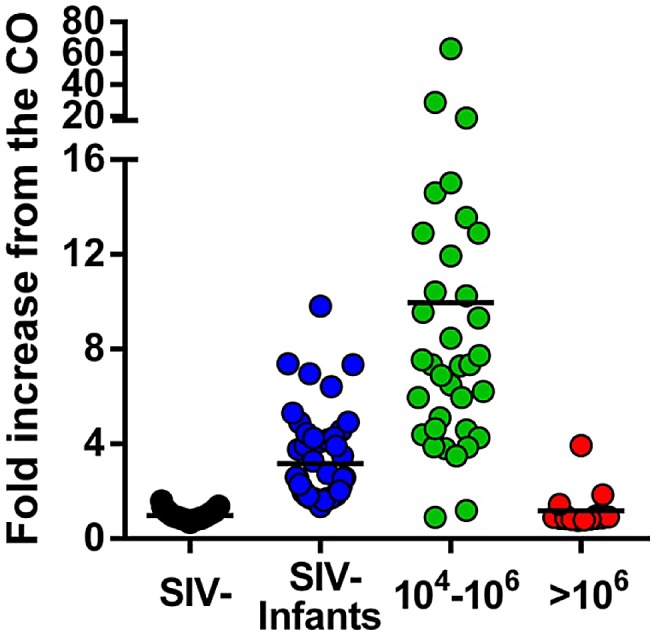
Testing was done by peptide ELISA (mapping the immunodominant region of gp41 of SIVagmVer). Samples were grouped based on PCR and VL quantification results as originating from uninfected vervets, SIV-infected vervets with VLs>106 SIVagmVer RNA copies/ml, SIV-infected vervets with VLs ranging from 104 to 106 SIVagmVer RNA copies/ml and samples from infant vervets. Samples in the latter group are negative by PCR but show different levels of anti-SIVagm IgG, therefore being suggestive of transplacental crossing of maternal antibodies. Values on the Y-axis represent fold increases of the optical density of the samples (measured at 492 nm) over the cut-off value (established at an optical density of 0.2).
Interestingly, serological testing showed that the majority (72%) of the plasma samples collected from infant AGMs were seropositive for SIVagm (Figure 6), while SIVagm could be amplified from only 3 of the infant samples (Figure 2 and Table 1), suggesting passive transfer of maternal antibodies and documenting massive in utero exposure of AGM offspring to SIVagm.
In conclusion, monitoring the viral replication and the anti-gp41 antibodies in SIVagmVer-infected AGMs allowed us to demonstrate that: (i) naturally occurring chronic SIV infection in a natural host is characterized by high VLs, which are in the same range as in experimentally-infected animals [27]; (ii) SIVagm transmission is very active in the wild; and (iii) the relatively low prevalence of SIVagm infection in offspring strongly contrasts with their massive in utero and breastfeeding exposure.
Assessment of the integrity of mucosal barrier
Both SIV-infected AGMs and SMs were reported to maintain the integrity of mucosal barrier and thus to control MT [30], [84], which has been reported to be the main factor behind increased immune activation that drives disease progression in pathogenic HIV/SIV infections [88], [89]. To assess MT in wild vervets, we tested the levels of sCD14 as a surrogate biomarker. As shown in Figure 7, the levels of sCD14 were similar between SIV-infected and SIV-uninfected AGMs and were in the range of those we previously documented in captive uninfected and experimentally-infected AGMs. Even the samples collected from acutely infected monkeys did not show a significant increase in the levels of sCD14, in agreement with our previous results from captive experimentally infected AGMs [30], [90], and suggesting that wild SIVagm-infected AGMs maintain the integrity of the mucosal barrier.
Figure 7. Plasma sCD14 levels in uninfected and SIVagmVer-infected vervets from South Africa.
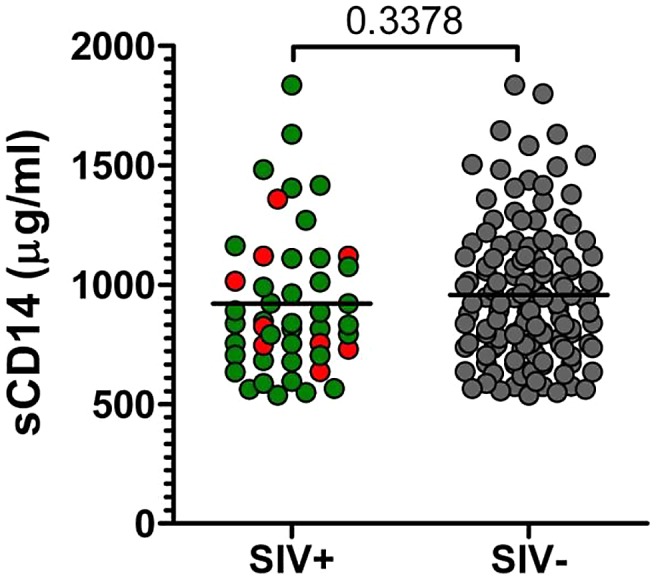
P value was calculated by the Mann-Whitney U-test. Animals diagnosed as acutely infected are denoted by light red circles.
Cytokine and chemokine testing
Finally, to assess the immune activation status between SIVagm-infected and uninfected AGMs in the wild, we measured and compared the levels of a large array of cytokines and chemokines in the available samples collected from the two groups in a Luminex assay. As shown in Figure 8, there was no significant increase in the cytokine/chemokine levels between SIV-infected and uninfected AGMs, with the exception of IL-6. In general, these cytokine/chemokine levels were in the same range of those previously reported by us and others [34] in captive uninfected and experimentally-infected AGMs during the chronic stage of SIVagm infection. This difference was mainly due to elevated levels of IL-6 in four of nine acutely-infected monkeys tested. When these samples were removed from the analysis, there were no significant differences in the levels of IL-6 between infected and uninfected AGMs. The increases in IL-6 probably reflect the transient increase in the levels of immune activation previously reported to occur during acute SIV infection in natural hosts [30], [33], [34], [91].
Figure 8. Cytokine and chemokine levels in uninfected and SIVagmVer-infected vervets from South Africa.

P value was calculated by the Mann-Whitney U-test. Animals diagnosed as acutely infected are denoted by light red circles.
Thus, our analyses demonstrate the maintenance of the mucosal barrier and the lack of increased immune activation in acutely and chronically-infected AGMs in the wild. Immune activation being the major determinant of disease progression in pathogenic HIV/SIV infections, our results further support the clinical assessment of a benign outcome of SIV infection in wild AGMs.
Discussion
AGMs are the largest reservoir of SIV in the wild [23] and one of the main animal models for the study SIV pathogenesis in natural hosts [23], [92]. However, so far no study has systematically investigated the prevalence and incidence of SIVagmVer in vervets and whether the natural history of SIVagm infection in wild free living animals differs from that of captive populations. To address these two key questions, we performed a large-scale analysis of SIVagmVer in wild vervets from the Republic of South Africa. Using only 0.5 ml of plasma and a new complex algorithm combining sequencing and phylogeny with virus quantification, serology, and assessment of surrogate markers of microbial translocation and immune activation, we concomitantly investigated, for the first time in a wild natural host African NHP monkey species, SIV diversity and evolution simultaneously with epidemiology and natural history. The strategy outlined here enables a systematic investigation of the natural history of SIV infection in wild natural hosts.
Epidemiologic considerations
Vervets are abundant throughout East Africa along the Indian Ocean coast. However, most studies carried out thus far focused only on vervets from Northeast Africa (Kenya, Ethiopia and Tanzania) [14], and reported an SIVagmVer prevalence rate of 50% [17]. Conversely, our large-scale survey of SIVagm prevalence and diversity in vervets includes samples collected from all age groups and from multiple locations in South Africa. We confirm that the overall prevalence of SIVagm infection in the wild is high (46%), supporting the notion that SIVagm is uniformly distributed and widespread in AGMs [15], [16], [93]. These prevalence levels are similar to those previously reported for sooty mangabeys [66], [94], [95] and mandrills [96], which are among the most commonly infected NHP species known to date. For the first time, we document that one of the reasons for such extensive spread of SIV in wild vervets is the high steady-state SIVagm replication which probably facilitates virus transmission between AGMs through sexual contact [97].
SIVagm prevalence is higher in AGM females than in males. This difference is not surprising given that SIVagm transmission has lower efficacy through the foreskin compared to vaginal exposure [98] and that most males are expected to have significantly lower exposure, as only a few attain the dominance status necessary for sexual access to breeding females [99]. Note, however, that the robust levels of prevalence in males may be explained by the observation that despite the sexually restrictive nature of male dominance hierarchies in vervets, many subordinate males in multimale, multifemale groups can gain sexual access to females through a number of strategies [100]–[102], and thus become infected [103]. The relatively long breeding season, which lasts up to three months, may also reduce the ability of the alpha male to effectively monopolize fertile females, and increase the chances of transmission through sexual contact in subordinate males [104]. Furthermore, SIVagm transmission between males probably also occurs through aggressive, blood-exposing contacts for dominance, similar to other species [105], as many of the males sampled had open wounds or scars from such contact. Additionally, many females show marked aggression against dispersing males attempting to migrate into their social groups, often resulting in serious injury, providing another route of infection between males and infected females [106].
The very uneven distribution of SIVagm infection among different age groups strongly supports the sexual route as the main transmission route for SIVagm, together with a significantly less effective maternal-to-infant transmission. The low rates of mother-to-infant transmission are paradoxical, as our study documented large SIVagm exposure of the offspring both in utero and through breastfeeding. The rates of maternal-to-infant transmission in wild AGMs are significantly lower than the 35 to 40% mother-to-infant transmission rates reported in HIV infection (http://www.unaids.org) or 40–70% transmission rates in rhesus macaques [107] and are likely the result of virus-host coadaptation [97]. It is also possible that, similar to previous reports in captive sooty mangabeys and AGMs [108], [109], SIV infection of infant AGMs is associated with significantly lower levels of viremia than infection of adult monkeys, which may therefore go undetected by our assay.
Also conceivable is that, in addition to sexual and maternal-to-infant routes, a nonnegligible proportion of SIVagmVer transmissions occur through biting or fighting, as injuries are frequent (in our study sample, 10% of vervets showed signs of recent injuries that could result in exposure, such as deep lacerations). Injuries preponderantly occur during the mating season, due to both male transfer and to contests for dominance between males, and are usually cleaned by licking, which may involve many different individuals and may explain identification of near-identical viruses in vervets which are not direct mating partners. Studies have shown that monkeys are highly susceptible to SIV infection through oral exposure [110], [111], and the high levels of systemic viral replication documented in wild vervets may facilitate oral transmission during wound cleaning.
At least 10% of the cases of SIVagmVer infection in our group were recent infections (defined by high levels of viral replication close to the peak VL and negative serologies). Recent infections tend to occur in a narrow time frame (10–12 days postinoculation in experimentally infected macaques and African NHPs [27] and 3 weeks postinfection in HIV-1-infected patients [87]); therefore our results point to relatively high rates of SIVagm transmission in the wild. Note, however, that most transmissions are probably condensed in the time frame of the mating season, and thus the overall incidence of SIV infection in AGMs is not as high as suggested by our cross-sectional analysis.
Virus diversity and evolutionary considerations
Our study identified a significant degree of divergence between strains collected from relatively homogenous groups of vervets, similar to previous reports for other species of African NHPs hosts, such as mandrills and sooty mangabeys [61], [66], [95], [96]. Also, we have identified closely related strains, suggestive of identifiable chains of transmission. In some instances, such highly related strains were observed in mother-offspring pairs, and probably occurred as a result of direct mother-to-infant transmission of the virus. In other instances, clusters of closely related viruses included a strain collected from a monkey documented as being recently infected. No clear association could be established for several of the closely related strains, and, as previously mentioned, these infections might have occurred during other social interactions, such as grooming or wound licking.
SIVagmVer strains from different locations were generally separated on the phylogenetic trees. Intermixing of strains was also observed, suggestive of SIVagm spread between communities by dispersing males. Indeed, although in vervet populations males range with their social group, they also migrate between groups upon or shortly after reaching sexual maturity (sometimes multiple times during the males lifetime) [106], [112].
Interestingly, the SIVagmVer strains collected from Free State locations (west of the Drakensberg Mountains) clustered differently within pol and env trees (Figure 2 and S1) as well as in the gag tree (data not shown) from those collected from monkeys situated along the Indian Ocean coast (east of the Drakensberg Mountains). Given that the geographical spread between coastal monkeys and those from Free State is similar, the most likely explanation for these phylogenetic relationships is that the Drakensberg Mountains acted as a barrier (through low temperatures in the elevated areas) to separate the vervet populations on the two sides. The Drakensberg Mountains are very old, with an estimated age of 280 million years, which implies that a separation of the vervet populations could have occurred in a time frame ranging from 3 million years ago (during the AGM spread throughout sub-Saharan Africa) to 100,000 years go (during the mass migrations that occurred in the Plio-Pleistocene glacial periods). The Drakensberg Mountains may also have acted as an effective barrier to separate different populations of at least two other species, the common impala (Aepyceros melampus melampus) [113] and the chacma baboon (Papio ursinus) [114]. We therefore performed relaxed molecular clock analyses to specify TMRCA of the SIVagm samples surrounding the Drakensberg Mountains range to a time-frame spanning 100,000 to 3,000,000 years since the present for the env tree, and in the pol tree we additionally included SIV strains of several species and further calibrated the tree based on isolates from Bioko Island [2]. Molecular clock analysis using Bayesian techniques is highly dependent on priors that are specified [115]; however our analysis is in agreement with another study in which the use of biogeography showed that SIVs are very old [2], and refuted timing calculations based exclusively on molecular clocks without regards to biogeographical calibration points [53].
Taken together, these observations strongly suggest that SIVagm infection of vervets is more ancient than the most recent previous estimate [53] and suggests that the concept of host-dependent evolution (supported by the observation that the four SIVagm types cluster together in the SIV trees) cannot be discarded using current molecular clock calibrations, but may possibly be discernible through the integration of further biogeographical divergence estimates.
Natural history considerations
It is generally assumed that SIV infection is nonpathogenic in natural hosts [1], [23], [44], [51], [54]. Data supporting this assumption are derived from the study of captive monkeys and a very limited number of species (AGMs, sooty mangabeys and mandrills) [1], [23], [44], [51]. Sporadic cases of progression to AIDS were reported in each of these species upon infection with their species-specific viruses [116]–[118] or with cross-species transmitted SIVs [119]. Large scale studies in wild animals are needed to draw definitive conclusions about pathogenicity of SIVs in their natural hosts in the wild, as demonstrated by recent observation that SIVcpz infection is pathogenic in naturally-infected chimpanzees [68], [69], [120]. The fact that such a major effect went undetected for decades supports the need for systematic assessment of the natural history of SIV infection in African NHPs in their native habitat, and not in captive environments (where health status is controlled, nutrition is monitored and exposure to adventitious agents that may impact clinical status is limited), to provide significant information regarding the clinical outcome of SIV infection in natural hosts.
The main limitation of our assessment of natural history of SIVagmVer infection in vervets is that only plasma samples were available, and thus we could not perform immunophenotypic characterization or in vitro replication studies. We circumvented this limitation by employing a combination of biomarkers that could be monitored in plasma.
We report that SIVagmVer infection most likely has a benign outcome in vervets. BMI were not different between SIV-infected and SIV-uninfected vervets. Detected levels of viral replication were high, but in the range of those reported during experimental studies [27]. In the documented cases of acute SIVagmVer infection, the VL levels were also in the range of those observed in experimentally infected monkeys during acute infection [27].
We measured levels of sCD14 to assess the levels of MT, which is one of the proposed mechanisms of excessive immune activation that drives HIV/SIV infection to progress to AIDS [88], [89]. sCD14 is a surrogate marker for direct measurement of endotoxin or Gram negative bacteria, and numerous studies have shown a strong association of increasing sCD14 levels and increasing LPS levels in plasma in both pathogenic HIV and SIV infections (presumably due to loss of barrier function in the gut in turn due to HIV-mediated destruction of GALT) [88], [89], and lack of increase in both markers in the setting of nonpathogenic SIV infections in natural hosts [30], [84], [90], [121].
It is critical to assess the integrity of the mucosal barrier in a natural host in the wild, as we and others have previously reported that lack of MT during chronic infection in AGMs and sooty mangabeys may be one mechanism of controlling chronic immune activation and disease progression [30], [84]. These studies have been done using captive NHPs. Wild monkeys are exposed to many pathogens in a significantly more hostile environment than are captive animals, which may impact the ability of SIV-infected monkeys to maintain the barrier. However, we found no significant difference in the levels of sCD14 between infected and uninfected vervets in the wild.
In addition to being a surrogate marker of MT, sCD14 is a marker of macrophage activation, and thus our results also suggest that there are no significant differences in the levels of macrophage activation between SIV-infected and uninfected AGMs in the wild.
Lack of chronic immune activation was also supported by the similar levels of cytokines and chemokines tested in SIVagmVer-infected and uninfected wild vervets. The only exception was represented by an increased level of IL-6 in SIVagmVer-infected animals, which was driven by high levels of this cytokine in acutely infected monkeys. We and others reported that a transient increase in immune activation occurs during acute SIV infection in natural hosts [33], [35], [36], [91], [122], which is rapidly resolved with the transition to chronic infection [30]. Therefore, the similar levels of cytokine in plasma between SIV-infected and uninfected animals are somewhat surprising in the context of a significant number of acute infections in our study sample. This apparent discrepancy with the results of experimental studies may be due to the fact that most of the experimental studies involved intravenous administration of large amounts of virus that may trigger inflammation and immune activation. Alternatively, as we have previously shown, the acute increases in levels of plasma cytokines and chemokines are very transient in natural hosts, being thus likely that our cross-sectional samples missed such increases.
Altogether, sCD14 and cytokine/chemokine testing data support the conclusion that SIVagmVer infection of wild vervets is not associated with significant increases in levels of immune activation. Immune activation being the main driver of HIV disease progression, our results corroborate the clinical assessment and support a nonprogressive outcome of SIVagmVer infection in the wild.
Final considerations: Wild vervet populations from South Africa as an animal model for exposed-seronegative/exposed-uninfected (ESN/EU) patients
Both clinical experience and a growing medical literature indicate that some persons who have been exposed to HIV infection remain uninfected [123]. Although in some instances this may simply represent good fortune, cohorts of uninfected persons have been reported who are considered at high risk for infection. Clarifying the determinants of protection against HIV infection is a high priority that will require careful selection of high-risk uninfected cohorts, which should undergo targeted studies of plausible mediators and broad screening for unexpected determinants of protection. An animal model for ESN/EU may permit such an assessment while circumventing the major ethical boundaries to the study of ESN/EU humans. Such an animal model is not currently available. The SIVmac/RM model is not relevant for ESN/EU studies because this model was derived for pathogenesis and vaccine studies (thus employing strains selected for pathogenicity and not for their transmission characteristics) [124]. On the other hand, de novo development of such a model in RMs would be extremely expensive and face major macaque availability restrictions. In this context, the SIVagmVer-infected wild vervet cohorts that we identified in South Africa may serve as a useful model for ESN/EU studies. In these populations, there is an enormous exposure to SIVagm. However, while up to 80% of sexually-active adult females are infected with SIVagmVer, the remaining 20% are uninfected in spite of the presumably massive potential for exposure to SIV throughout their lives. Thus, field studies to assess the prevalence and incidence of SIV infection in the wild and prospectively confirm resistance to infection in a subset of ESN/EU monkeys, combined with genetic, immunologic and virologic assessment of the monkeys in the cohorts, may permit identification of correlates of ESN/EU, an instrumental leap for designing new therapeutic strategies aimed at preventing HIV infection.
In conclusion, we report that SIVagmVer is highly prevalent in wild vervet populations from South Africa, that the virus has long been present in this population and that coevolution between SIVagmVer and its natural host, the vervet monkey, resulted in an infection which has a benign outcome in the wild. More importantly, our study has identified a potential model for the study of ESN/EU cases, which has a strong potential to identify correlates of protection against HIV/SIV infection.
Materials and Methods
Ethics statement
All the animals sampled in this study were used according to regulations set forth by the Animal Welfare Act. The animal sampling protocols were approved by the University of Wisconsin-Milwaukee Institutional Animal Care and Use Committee (IACUC). Studies at the UCLA and University of Pittsburgh were carried out through an agreement with the UWM. At the University of the Free State UFS, ethical clearance was provided by the Interfaculty Animal Ethics Committee (project no. 13/2010).
Samples
The study was carried out in feral and semifree vervets (n = 225) living in different areas in South Africa, aged between 6 months and 10 years (Figure 1). Animals were individually trapped using established methods [125]. Details on animal capture are provided as Text S1. Blood was collected through venous puncture. Details are provided as Text S1. Each monkey included in the study had a microchip implanted for further identification and to prevent duplicate sampling. Detailed clinical assessment of the monkeys (including signs associated with immunodeficiency, i.e., lymphadenopathy, low weight, fever or rashes) was performed at the time of blood collection. Details are provided as Text S1. Upon collection, plasma was immediately stored at −80°C. A 500 µl aliquot of plasma from each monkey was available for this study.
Plasma processing and testing strategy
Upon arrival, plasma samples were thawed for RNA extraction and separated into three aliquots: the first aliquot was used for testing the prevalence levels of SIV by PCR. Additional SIV ELISA testing was performed using this aliquot to establish the stage of acute SIV infection and infant exposure to SIVagm. The second aliquot was used for quantification of SIVagm VLs by real-time PCR. Primers and probes were designed based on the SIVagm integrase sequences. Correlation between VLs and serology enabled identification of acutely SIV-infected AGMs [87]. Finally, the third aliquot was used to assess the natural history of SIVagm infection in the wild by testing surrogate markers for microbial translocation and immune activation (sCD14 and cytokine/chemokine levels).
Viral RNA extraction and cDNA synthesis
From each plasma specimen, viral RNA was extracted using the QIAamp Viral RNA Mini kit (QIAGEN, Germantown, MD). RNA was eluted and immediately subjected to cDNA synthesis. Reverse transcription of RNA to single-stranded cDNA was performed using SuperScript III reverse transcription according to the manufacturer's recommendations (Invitrogen, Carlsbad, CA). In brief, each cDNA reaction included 1×RT buffer, 0.5 mM of each deoxynucleoside triphosphate, 5 mM dithiothreitol, 2 U/ml RNaseOUT (RNase inhibitor), 10 U/ml of Super-Script III reverse transcriptase, and 0.25 µM antisense primers SIV-POL-OR (5′-ACB ACY GCN CCT TCH CCT TTC-3′), SIV-GAG-B (5′-CCT ACT CCC TGA CAG GCC GTC AGC ATT TCT TC-3′) and SIV-ENV-B (5′-AGA GCT GTG ACG CGG GCA TTG AGG TT-3′). The mixture was incubated at 50°C for 60 min, followed by an increase in temperature to 55°C for an additional 60 min. The reaction was then heat-inactivated at 70°C for 15 min and treated with 2 U of RNase H at 37°C for 20 min. The newly synthesized cDNA was used immediately or frozen at −80°C.
Nested PCR and DNA gel extraction
SIVagm RNA was amplified byPCR using Platinium Taq Polymerase High Fidelity (Invitrogen, Carlsbad, CA). A 600 bp pol integrase fragment was amplified by nested PCR using outer primers POL-IS4 (5′-CCA GCN CAC AAA GGN ATA GGA GG-3′) and POL-OR and inner primers SIV-POL-IS4 and SIV-POL-Unipol2 (5′-CCC CTA TTC CTC CCC TTC TTT TAA AA-3′) [59], [126]. An 846 bp gag fragment was amplified by nested PCR using outer primers GAG-A (5′-AGG TTA CGG CCC GGC GGA AAG AAA A-3′) and GAG-B, and inner primers GAG-C (5′-AGT ACA TGT TAA AAC ATG TAG TAT GGG C-3′) and GAG-F (5′-CCT TAA GCT TTT GTA GAA TCT ATC TAC ATA-3′) [127]. Finally, a 900-bp env fragment encompassing the V3-V5 gp120 region and the 5′ end of gp41 envelope regions was amplified by nested PCR using outer primers ENV-A (5′-GAA GCT TGT GAT AAA ACA TAT TGG AT-3′) and ENV-B, and inner primers ENV-C (5′-GTG CAT TGT ACA GGG TTA ATG AAT ACA ACA G-3′) and ENV-D (5′-TTC TTC TGC TGC AGTA TCC CAG CAA G-3′) [128]. Nested PCR products were subjected to 1% agarose (Invitrogen, Carlsbad, CA) gel electrophoresis and extracted and purified by QIAquick Gel Extraction kit (QIAGEN, Germantown, MD).
DNA sequencing
SIV gene amplicons were directly sequenced by the University of Pittsburgh Genomics and Proteomics Core laboratories (GPCL) using the nested PCR primers. Individual sequence for each amplicon was edited and inspected using BioEdit 7.1.3. Sequences were translated for analyses on the V3–V5 region of the SIVagmVer envelope with BioEdit.
Phylogenetic analysis
The gag, pol and env nucleotide sequence alignments were aligned with reference sequences obtained from the Los Alamos National Laboratory HIV Sequence Database (http://hiv-web.lanl.gov). The alignment for the pol region included nucleotide sequences sampled from the island of Bioko [2]. Newly derived SIV sequences were aligned using MUSCLE sequence alignment software [129] and alignments were edited manually where necessary.
Separate phylogenetic trees for each of the genes were inferred by maximum-likelihood using PhyML 3.0 [130], with a GTR substitution model, gamma-distributed rate heterogeneity at sites and an SPR tree search. Nonparametric bootstrap analysis with 1,000 replicates was performed to assess the reliability of branching order. In addition, calibrated molecular clock trees were calculated using BEAST v1.7.3 [115] to infer likely MRCA dates between different lineages. For molecular clock analysis, we used an HKY substitution model with gamma-distributed rate heterogeneity among sites and an uncorrelated lognormal relaxed molecular clock. A Yule tree prior was used for the pol region, and a constant size coalescent for the env region. Evolutionary rate was calibrated by setting a uniform prior bounded by 10,000 and 3,000,000 years, as TMRCA of SIVagm isolates sampled from areas surrounding the Drakensberg Mountains. For the pol region, a normal distribution with a mean of 10,000 years and a standard deviation of 1,000 years was additionally used to calibrate TMRCA of Bioko and mainland SIVdrl sequences. Chains were run for 107–108 states and ESS values were high (>300). Analysis of the molecular clock trees was performed using Tracer v1.5 (http://tree.bio.ed.ac.uk/software/tracer/), and trees were inferred by maximum clade credibility with mean node heights using TreeAnnotator v1.7.3, and visualized using FigTree v1.3.1 (http://tree.bio.ed.ac.uk/software/figtree/). To test for signals of recombination in the dataset, we ran the Neighbor-Net algorithm implemented in SplitsTree [131] to infer phylogenetic networks for the env and pol regions.
Viral load quantification
We used the generated pol alignment to design specific primers and probe for VL quantification in wild vervets: pol-F (5′-GTA GCC AGT GGG TTC ATA GAA GCA-3′), pol-R (5′-CAT TGC CTT TAC TTC CTG TGA AAT GA-3′) and pol-Probe (5′-/56-FAM/TAG AGA AAC/ZEN/AGG AAA AGA AAC AGC AA/3IABkF/-3′). Primers and probe were synthesized by IDTDNA (Coralville, IA) and were used in a one-step real-time PCR assay with TaqMan One Step PCR Master Mix (Applied Biosystems, Branchburg, NJ). Real-time PCR was performed in MicroAmp Optical 96-well plates (Applied Biosystems, Branchburg, NJ) by mixing the OneStep RT-PCR Enzyme Mix with 5 µl isolated RNA in a 50 µl final reaction volume. Real-time PCR conditions were as follows: 30 min at 50°C, 15 min at 95°C, followed by 45 cycles of 95°C for 15 s and 60°C for 1 min. Dilutions of all components were made using sterile RNase-free water. Data were collected and analyzed using the PE Applied Biosystems software provided. RNA copies/well were adjusted to copies per milliliter of original plasma. Samples were tested in duplicate and the number of RNA copies determined by comparison with a standard curve obtained using known amounts of SIV-pol RNA. The SIVagm RNA standard was produced by cloning one of the SIVagmVerpol gene fragments into Vector pBluescript SK+ and linearizing it by Xba I as a DNA template for RNA in vitro transcription. Standard RNA was produced by using MEGAscript kit (Applied Biosystems, Branchburgh, NJ), according to the manufacturer's instructions. Briefly, the transcription reaction system was assembled at room temperature and mixed thoroughly, followed by incubation at 37°C for 4 hours. One microliter of TURBO DNase was added to the reaction mixture followed by incubation at 37°C for 15 minutes to digest the DNA template. RNA was recovered by Lithium chloride precipitation. The amount of RNA was quantified at A260, aliquoted and immediately stored at −80°C.Detection limit of the SIVagmVer quantification assay was 100 copies/ml.
Serology
Based on the env sequence data, a peptide mapping the immunodominant region in the Gp41 transmembrane glycoprotein was synthesized and used in a peptide enzyme linked immunosorbent assay (ELISA), as described [132]. The inferred peptide sequence was TALEKYLEDQARLNVWGCAWKQVC, and this sequence was very well conserved between different SIVagmVer isolates, including previously reported reference strains from Kenya and Ethiopia [9], [10], [93]. The Gp36 peptide was synthesized to a purity of at least 90% (Fisher Scientific, USA) and the assay was performed as previously reported [132]. The cut-off of the reaction was arbitrarily set at 0.20, as per previous reports [132].
Microbial translocation (MT)
MT was assessed using the levels of soluble CD14 (sCD14) as a surrogate marker [89]. CD14 is a transmembranous protein, which also exists in a soluble form (sCD14; both as a shed membrane form and an alternatively spliced form), as a part of the complex that presents endotoxin (lipopolysaccharide or LPS) to TLR4 on monocytes. When monocytes are activated, ectodomain shedding results in increased sCD14 levels. sCD14 is therefore surrogate for direct measurement of endotoxin or Gram negative bacteria which translocate from the intestinal lumen to general circulation as a result of the immunologic injury inflicted at the mucosal level by the pathogenic HIV/SIV infection [88]–[90], [121]. sCD14 levels were measured using a quantitative sandwich enzyme immunoassay technique (Quantikine Human sCD14 Immunoassay, R&D Systems, Minneapolis, MN). The detection limit of this kit is 200 ng/mL and can range up to 5000 ng/mL, with an interassay coefficient of variability ranging between 7.19% and 10.9%.
Cytokine and chemokine testing
Cytokine testing in plasma was done using a sandwich immunoassay-based protein array system, Cytokine Monkey Magnetic 28-Plex Panel (Invitrogen, Camarillo, CA), as instructed by the manufacturer, and results were read by the Bio-Plex array reader (Bio-Rad Laboratories, Hercules, CA), which uses Luminex fluorescent-bead-based technology (Luminex Corporation, Austin, TX).
Statistical analyses
Statistical analyses were performed to analyze the results of viral load, sCD14 and cytokine testing using Mann-Whitney U-tests and Prism 4.0 software (Prism, Irvine, CA).
Nucleotide sequence accession numbers
The nucleotide sequences of the pol and env sequences from SIVagmVer-infected vervets from South Africa were deposited in the GeneBank (accession numbers: JX462308-JX462444).
Supporting Information
Maximum likelihood trees for env gene of the newly derived SIVagmVer sequences from wild vervets in South Africa. Maximum likelihood estimates were performed using 1000 replicate bootstrap analysis. Internal nodes indicate level of support values for internal branching. Sequences are colored depending on the region in which the sequences were sampled, with red indicating sequences from Free State, green indicating sequences from KwaZulu-Natal, and blue indicating sequences from Eastern Coast territories. In addition, sequences colored in gray are sampled from the Riverside Wildlife Rehabilitation and Environmental Education Centre (RWREC), Letsitele, Limpopo semifree colony that is housing released pets or animals with health problems, and their origin is unknown, and sequences colored in black indicate outgroup sequences. The strain nomenclature includes the identification number, monkey status (M-male, F-female; I-infant; J-juvenile and A-adult), and the site and the state of origin (FS-Free State; KZN-KwaZulu Natal; EC-Eastern Cape).
(TIFF)
SplitsTree showing the pol gene of South African SIVagmVer sequences. Nodes are colored by the region in which they were sampled, with red, blue and green denoting Free State, East Coast and KwaZulu-Natal, respectively. Potentially interesting recombinants have been circled, including VMT17083-IM-Gariep_FS and a cluster indicating possible mixing between East Coast and KwaZulu-Natal sequences.
(TIFF)
Comparative body mass index (BMI) assessment based on age and location in SIV-infected and SIV uninfected wild vervet monkeys ( Chlorocebus pygerithrus ) from South Africa.
(TIFF)
SIV prevalence in vervet monkeys from South Africa based on location of collected samples.
(DOC)
Age- and sex-related prevalence of SIVagmVer in semifree vervets from Riverside Wildlife Rehabilitation and Education Center, Letsitele, Limpopo.
(DOC)
Estimates of evolutionary divergence over sequence pairs between groups. The number of base substitutions per site±standard error estimate(s) from averaging over all sequence pairs between groups are shown± Analyses were conducted using the Maximum Composite Likelihood model [1]. The rate variation among sites was modeled with a gamma distribution (shape parameter = 0.5). env analysis involved 81 nucleotide sequences. All positions containing gaps and missing data were eliminated. There were a total of 822 positions in the final dataset for env. pol analysis involved 98 nucleotide sequences. All positions containing gaps and missing data were eliminated. There were a total of 574 positions in the final pol dataset. Evolutionary analyses were conducted in MEGA5 [2].
(DOC)
Most recent common ancestor (MRCA) estimates using relaxed molecular clocks to incorporate biogeographical assumptions. Calibration for the env gene is based on time to MRCA (TMRCA) of SIVagm sequences sampled around the Drakensberg Mountains, presumed to be 100,000–3,000,000 years of age. Calibration of the pol gene is based on TMRCA of SIVagm sequences as before, as well as SIVdrl sequences sampled from the Bioko Island of Equatorial Guinea, presumed to have been isolated for approximately 10,000 years. These estimates indicate that SIV is potentially considerably older than previous estimates, and refute timing estimates based exclusively on molecular clocks without calibration.
(DOC)
Details on animal origin, animal capture, sampling methodology and clinical assessment.
(DOC)
Acknowledgments
We thank Drs. Beatrice Hahn, Vanessa Hirsch and Preston Marx for helpful discussion. Samples used in this study were collected as a part of the Systems Biology Sample Repository. We thank Department of Environmental Affairs, South Africa; Department of Economic Development and Environmental Affairs, Eastern Cape; Department of Tourism, Environmental and Economic Affairs, Free State Province; the Ezemvelo KZN Wildlife in KwaZulu Natal Province; and the Department of Economic Development, Environment and Tourism, Limpopo Province. Map of the Republic of South Africa used in Figure 1 was obtained from Wikipedia (http://en.wikipedia.org/wiki/File:South_Africa_sat.jpg).
Funding Statement
This work was supported by NIH grant R01OD010980 from the Office of Research Infrastructure Programs/OD (formerly R01RR016300 from NCRR) to NF and by NIH/NIAID/NCRR grants RO1 AI064066, R01 RR025781, P01 AI088564 to CA and IP. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. VandeWoude S, Apetrei C (2006) Going wild: Lessons from T-lymphotropic naturally occurring lentiviruses. Clin Microbiol Rev 19: 728–762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Worobey M, Telfer P, Souquière S, Hunter M, Coleman CA, et al. (2010) Island biogeography reveals the deep history of SIV. Science 330: 1487. [DOI] [PubMed] [Google Scholar]
- 3. Ahuka-Mundeke S, Ayouba A, Mbala-Kingebeni P, Liegeois F, Esteban A, et al. (2011) Novel multiplexed HIV/simian immunodeficiency virus antibody detection assay. Emerg Infect Dis 17: 2277–2286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Groves C (2001) Primate taxonomy. Washington, DC: Smithsonian Institution Press.
- 5.Groves CP (2005) Order Primates. In: Wilson DE, Reeder DM, editors. Mammal Species of the World. Baltimore, MD: The Johns Hopkins University Press. pp. 111–184.
- 6. Grubb P, Butinsky TM, Oates JF, Bearder SK, Disotell TD, et al. (2003) Assessment of the diversity of African primates. Int J Primatol 24: 1301–1357. [Google Scholar]
- 7. Fomsgaard A, Allan J, Gravell M, London WT, Hirsch VM, et al. (1990) Molecular characterization of simian lentiviruses from east African green monkeys. J Med Primatol 19: 295–303. [PubMed] [Google Scholar]
- 8. Hirsch VM, McGann C, Dapolito G, Goldstein S, Ogen-Odoi A, et al. (1993) Identification of a new subgroup of SIVagm in tantalus monkeys. Virology 197: 426–430. [DOI] [PubMed] [Google Scholar]
- 9. Fukasawa M, Miura T, Hasegawa A, Morikawa S, Tsujimoto H, et al. (1988) Sequence of simian immunodeficiency virus from African green monkey, a new member of the HIV/SIV group. Nature 333: 457–461. [DOI] [PubMed] [Google Scholar]
- 10. Jin MJ, Hui H, Robertson DL, Muller MC, Barre-Sinoussi F, et al. (1994) Mosaic genome structure of simian immunodeficiency virus from west African green monkeys. EMBO J 13: 2935–2947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Allan JS, Short M, Taylor ME, Su S, Hirsch VM, et al. (1991) Species-specific diversity among simian immunodeficiency viruses from African green monkeys. J Virol 65: 2816–2828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Hendry RM, Wells MA, Phelan MA, Schneider AL, Epstein JS, et al. (1986) Antibodies to simian immunodeficiency virus in African green monkeys in Africa in 1957–62. Lancet 2: 455. [DOI] [PubMed] [Google Scholar]
- 13. Lowenstine LJ, Pedersen NC, Higgins J, Pallis KC, Uyeda A, et al. (1986) Seroepidemiologic survey of captive Old-World primates for antibodies to human and simian retroviruses, and isolation of a lentivirus from sooty mangabeys (Cercocebus atys). Int J Cancer 38: 563–574. [DOI] [PubMed] [Google Scholar]
- 14. Ohta Y, Masuda T, Tsujimoto H, Ishikawa K, Kodama T, et al. (1988) Isolation of simian immunodeficiency virus from African green monkeys and seroepidemiologic survey of the virus in various non-human primates. Int J Cancer 41: 115–122. [DOI] [PubMed] [Google Scholar]
- 15. Phillips-Conroy JE, Jolly CJ, Petros B, Allan JS, Desrosiers RC (1994) Sexual transmission of SIVagm in wild grivet monkeys. J Med Primatol 23: 1–7. [DOI] [PubMed] [Google Scholar]
- 16. Jolly C, Phillips-Conroy JE, Turner TR, Broussard S, Allan JS (1996) SIVagm incidence over two decades in a natural population of Ethiopian grivet monkeys (Cercopithecus aethiops aethiops). J Med Primatol 25: 78–83. [DOI] [PubMed] [Google Scholar]
- 17. Goldstein S, Brown CR, Ourmanov I, Pandrea I, Buckler-White A, et al. (2006) Comparison of simian immunodeficiency virus SIVagmVer replication and CD4+ T-cell dynamics in vervet and sabaeus African green monkeys. J Virol 80: 4868–4877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Muller MC, Saksena NK, Nerrienet E, Chappey C, Herve VM, et al. (1993) Simian immunodeficiency viruses from central and western Africa: evidence for a new species-specific lentivirus in tantalus monkeys. J Virol 67: 1227–1235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Johnson PR, Fomsgaard A, Allan J, Gravell M, London WT, et al. (1990) Simian immunodeficiency viruses from African green monkeys display unusual genetic diversity. J Virol 64: 1086–1092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Baier M, Garber C, Muller C, Cichutek K, Kurth R (1990) Complete nucleotide sequence of a simian immunodeficiency virus from African green monkeys: a novel type of intragroup divergence. Virology 176: 216–221. [DOI] [PubMed] [Google Scholar]
- 21. Li Y, Naidu Y, Fultz P, Daniel MD, Desrosiers RC (1989) Genetic diversity of simian immunodeficiency virus. J Med Primatol 18: 261–269. [PubMed] [Google Scholar]
- 22. Xing J, Wang H, Zhang Y, Ray DA, Tosi AJ, et al. (2007) A mobile element-based evolutionary history of guenons (tribe Cercopithecini). BMC Biol 5: 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Pandrea I, Apetrei C (2010) Where the wild things are: Pathogenesis of SIV infection in African nonhuman primate hosts. Curr HIV/AIDS Reports 7: 28–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Pandrea I, Silvestri G, Apetrei C (2009) AIDS in African nonhuman primate hosts of SIVs: A new paradigm of SIV infection. Curr HIV Res 6: 57–72. [DOI] [PubMed] [Google Scholar]
- 25. Pandrea I, Apetrei C, Dufour J, Dillon N, Barbercheck J, et al. (2006) Simian immunodeficiency virus (SIV) SIVagm.sab infection of Caribbean African green monkeys: New model of the study of SIV pathogenesis in natural hosts. J Virol 80: 4858–4867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Pandrea I, Kornfeld C, Ploquin MJ-I, Apetrei C, Faye A, et al. (2005) Impact of viral factors on very early in vivo replication profiles in SIVagm-infected African green monkeys. J Virol 79: 6249–6259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Pandrea I, Silvestri G, Onanga R, Veazey RS, Marx PA, et al. (2006) Simian immunodeficiency viruses replication dynamics in African non-human primate hosts: common patterns and species-specific differences. J Med Primatol 35: 194–201. [DOI] [PubMed] [Google Scholar]
- 28. Broussard SR, Staprans SI, White R, Whitehead EM, Feinberg MB, et al. (2001) Simian immunodeficiency virus replicates to high levels in naturally infected African green monkeys without inducing immunologic or neurologic disease. J Virol 75: 2262–2275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Goldstein S, Ourmanov I, Brown CR, Beer BE, Elkins WR, et al. (2000) Wide range of viral load in healthy african green monkeys naturally infected with simian immunodeficiency virus. J Virol 74: 11744–11753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Pandrea I, Gautam R, Ribeiro R, Brenchley JM, Butler IF, et al. (2007) Acute loss of intestinal CD4+ T cells is not predictive of SIV virulence. J Immunol 179: 3035–3046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Pandrea I, Ribeiro RM, Gautam R, Gaufin T, Pattison M, et al. (2008) Simian immunodeficiency virus SIVagm dynamics in African green monkeys. J Virol 82: 3713–3724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Favre D, Lederer S, Kanwar B, Ma ZM, Proll S, et al. (2009) Critical loss of the balance between Th17 and T regulatory cell populations in pathogenic SIV infection. PLoS Pathogens 5: e1000295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Jacquelin B, Mayau V, Targat B, Liovat AS, Kunkel D, et al. (2009) Nonpathogenic SIV infection of African green monkeys induces a strong but rapidly controlled type I IFN response. J Clin Invest 119: 3544–3555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Kornfeld C, Ploquin MJ, Pandrea I, Faye A, Onanga R, et al. (2005) Antiinflammatory profiles during primary SIV infection in African green monkeys are associated with protection against AIDS. J Clin Invest 115: 1082–1091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Gaufin T, Pattison M, Gautam R, Stoulig C, Dufour J, et al. (2009) Effect of B cell depletion on viral replication and clinical outcome of SIV infection in a natural host. J Virol 83: 10347–10357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Gaufin T, Ribeiro RM, Gautam R, Dufour J, Mandell D, et al. (2010) Experimental depletion of CD8+ cells in acutely SIVagm-infected African green monkeys results in increased viral replication. Retrovirology 7: 42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Zahn RC, Rett MD, Korioth-Schmitz B, Sun Y, Buzby AP, et al. (2008) Simian Immunodeficiency Virus (SIV)-specific CD8+ T cell responses in chronically SIVagm-infected vervet African green monkeys. J Virol 82: 11577–11588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Zahn RC, Rett MD, Li M, Tang H, Korioth-Schmitz B, et al. (2010) Suppression of adaptive immune responses during primary SIV infection of sabaeus African green monkeys delays partial containment of viremia but does not induce disease. Blood 115: 3070–3078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Schmitz JE, Zahn RC, Brown CR, Rett MD, Li M, et al. (2009) Inhibition of adaptive immune responses leads to a fatal clinical outcome in SIV-infected pigtailed macaques but not vervet African green monkeys. PLoS Pathog 5: e1000691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Hirsch VM (2004) What can natural infection of African monkeys with simian immunodeficiency virus tell us about the pathogenesis of AIDS? AIDS Rev 6: 40–53. [PubMed] [Google Scholar]
- 41. Gicheru MM, Otsyula M, Spearman P, Graham BS, Miller CJ, et al. (1999) Neutralizing antibody responses in Africa green monkeys naturally infected with simian immunodeficiency virus (SIVagm). J Med Primatol 28: 97–104. [DOI] [PubMed] [Google Scholar]
- 42. Estaquier J, Idziorek T, de Bels F, Barre-Sinoussi F, Hurtrel B, et al. (1994) Programmed cell death and AIDS: significance of T-cell apoptosis in pathogenic and nonpathogenic primate lentiviral infections. Proc Natl Acad Sci U S A 91: 9431–9435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Cumont MC, Diop O, Vaslin B, Elbim C, Viollet L, et al. (2008) Early divergence in lymphoid tissue apoptosis between pathogenic and nonpathogenic simian immunodeficiency virus infections of nonhuman primates. J Virol 82: 1175–1184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Pandrea I, Sodora DL, Silvestri G, Apetrei C (2008) Into the wild: simian immunodeficiency virus (SIV) infection in natural hosts. Trends Immunol 29: 419–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Paiardini M, Cervasi B, Reyes-Aviles E, Micci L, Ortiz AM, et al. (2011) Reduced CCR5 up-regulation upon activation limits virus replication in central-memory CD4+ T cells of SIV-infected sooty mangabeys. Nature Medicine 17: 830–836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Pandrea I, Apetrei C, Gordon S, Barbercheck J, Dufour J, et al. (2007) Paucity of CD4+CCR5+ T cells is a typical feature of natural SIV hosts. Blood 109: 1069–1076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Pandrea I, Onanga R, Souquiere S, Mouinga-Ondéme A, Bourry O, et al. (2008) Paucity of CD4+CCR5+ T-cells may prevent breastfeeding transmission of SIV in natural non-human primate hosts. J Virol 82: 5501–5509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Beaumier CM, Harris LD, Goldstein S, Klatt NR, Whitted S, et al. (2009) CD4 downregulation by memory CD4+ T cells in vivo renders African green monkeys resistant to progressive SIVagm infection. Nat Med 15: 879–885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Vinton C, Klatt NR, Harris LD, Briant JA, Sanders-Beer BE, et al. (2011) CD4-like immunological function by CD4- T cells in multiple natural hosts of simian immunodeficiency virus. J Virol 85: 8702–8708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Apetrei C, Gaufin T, Gautam R, Vinton C, Hirsch VM, et al. (2010) Pattern of SIVagm infection in patas monkeys suggests that host adaptation to SIV infection may result in resistance to infection and virus extinction. J Infect Dis 202 Suppl 3: S371–376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Silvestri G, Paiardini M, Pandrea I, Lederman MM, Sodora DL (2007) Understanding the benign nature of SIV infection in natural hosts. J Clin Invest 117: 3148–3154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Charleston MA, Robertson DL (2002) Preferential host switching by primate lentiviruses can account for phylogenetic similarity with the primate phylogeny. Syst Biol 51: 528–535. [DOI] [PubMed] [Google Scholar]
- 53. Wertheim JO, Worobey M (2007) A challenge to the ancient origin of SIVagm based on African green monkey mitochondrial genomes. PLoS Pathog 3: e95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Hahn BH, Shaw GM, De Cock KM, Sharp PM (2000) AIDS as a zoonosis: scientific and public health implications. Science 287: 607–614. [DOI] [PubMed] [Google Scholar]
- 55. Peeters M, Courgnaud V, Abela B, Auzel P, Pourrut X, et al. (2002) Risk to human health from a plethora of simian immunodeficiency viruses in primate bushmeat. Emerg Infect Dis 8: 451–457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Apetrei C, Robertson DL, Marx PA (2004) The history of SIVS and AIDS: epidemiology, phylogeny and biology of isolates from naturally SIV infected non-human primates (NHP) in Africa. Front Biosci 9: 225–254. [DOI] [PubMed] [Google Scholar]
- 57. Courgnaud V, Abela B, Pourrut X, Mpoudi-Ngole E, Loul S, et al. (2003) Identification of a new simian immunodeficiency virus lineage with a vpu gene present among different cercopithecus monkeys (C. mona, C. cephus, and C. nictitans) from Cameroon. J Virol 77: 12523–12534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Courgnaud V, Formenty P, Akoua-Koffi C, Noe R, Boesch C, et al. (2003) Partial molecular characterization of two simian immunodeficiency viruses (SIV) from African colobids: SIVwrc from Western red colobus (Piliocolobus badius) and SIVolc from olive colobus (Procolobus verus). J Virol 77: 744–748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Courgnaud V, Pourrut X, Bibollet-Ruche F, Mpoudi-Ngole E, Bourgeois A, et al. (2001) Characterization of a novel simian immunodeficiency virus from guereza colobus monkeys (Colobus guereza) in Cameroon: a new lineage in the nonhuman primate lentivirus family. J Virol 75: 857–866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Courgnaud V, Salemi M, Pourrut X, Mpoudi-Ngole E, Abela B, et al. (2002) Characterization of a novel simian immunodeficiency virus with a vpu gene from greater spot-nosed monkeys (Cercopithecus nictitans) provides new insights into simian/human immunodeficiency virus phylogeny. J Virol 76: 8298–8309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Apetrei C, Metzger MJ, Robinson D, Ling B, Telfer PT, et al. (2005) Detection and partial characterization of new simian immunodeficiency virus (SIVsm) strains from bush meat samples from rural Sierra Leone. J Virol 79: 2631–2636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Liegeois F, Courgnaud V, Switzer WM, Murphy HW, Loul S, et al. (2006) Molecular characterization of a novel simian immunodeficiency virus lineage (SIVtal) from northern talapoins (Miopithecus ogouensis). Virology 349: 55–65. [DOI] [PubMed] [Google Scholar]
- 63. Keele BF, Van Heuverswyn F, Li Y, Bailes E, Takehisa J, et al. (2006) Chimpanzee reservoirs of pandemic and nonpandemic HIV-1. Science 313: 523–526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Van Heuverswyn F, Li Y, Neel C, Bailes E, Keele BF, et al. (2006) Human immunodeficiency viruses: SIV infection in wild gorillas. Nature 444: 164. [DOI] [PubMed] [Google Scholar]
- 65. Santiago ML, Lukasik M, Kamenya S, Li Y, Bibollet-Ruche F, et al. (2003) Foci of endemic simian immunodeficiency virus infection in wild-living eastern chimpanzees (Pan troglodytes schweinfurthii). J Virol 77: 7545–7562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Santiago ML, Range F, Keele BF, Li Y, Bailes E, et al. (2005) Simian immunodeficiency virus infection in free-ranging sooty mangabeys (Cercocebus atys atys) from the Tai Forest, Cote d'Ivoire: implications for the origin of epidemic human immunodeficiency virus type 2. J Virol 79: 12515–12527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Santiago ML, Rodenburg CM, Kamenya S, Bibollet-Ruche F, Gao F, et al. (2002) SIVcpz in wild chimpanzees. Science 295: 465. [DOI] [PubMed] [Google Scholar]
- 68. Keele BF, Holland Jones J, Terio K, Estes JD, Rudicell RS, et al. (2009) Increased mortality and AIDS-like immunopathology in wild chimpanzees infected with SIVcpz. Nature 460: 515–519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Rudicell RS, Holland Jones J, Wroblewski EE, Learn GH, Li Y, et al. (2010) Impact of simian immunodeficiency virus infection on chimpanzee population dynamics. PLoS Pathog 6: e1001116 doi: 10.1371/journal.ppat.1001116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Terio KA, Kinsel MJ, Raphael J, Mlengeya T, Lipende I, et al. (2011) Pathologic lesions in chimpanzees (Pan trogylodytes schweinfurthii) from Gombe National Park, Tanzania, 2004–2010. J Zoo Wildl Med 42: 597–607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Kirchhoff F, Morrison HG, Desrosiers RC (1995) Identification of V3 mutations that can compensate for inactivating mutations in C4 of simian immunodeficiency virus. Virology 213: 179–189. [DOI] [PubMed] [Google Scholar]
- 72. Konig RR, Flory E, Steidl S, Neumann J, Coulibaly C, et al. (2002) Engineered CD4- and CXCR4-using simian immunodeficiency virus from African green monkeys is neutralization sensitive and replicates in nonstimulated lymphocytes. J Virol 76: 10627–10636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Steidl S, Stitz J, Schmitt I, Konig R, Flory E, et al. (2002) Coreceptor Switch of [MLV(SIVagm)] pseudotype vectors by V3-loop exchange. Virology 300: 205–216. [DOI] [PubMed] [Google Scholar]
- 74. Recordon-Pinson P, Soulie C, Flandre P, Descamps D, Lazrek M, et al. (2010) Evaluation of the genotypic prediction of HIV-1 coreceptor use versus a phenotypic assay and correlation with the virological response to maraviroc: the ANRS GenoTropism study. Antimicrob Agents Chemother 54: 3335–3340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Lengauer T, Sander O, Sierra S, Thielen A, Kaiser R (2007) Bioinformatics prediction of HIV coreceptor usage. Nat Biotechnol 25: 1407–1410. [DOI] [PubMed] [Google Scholar]
- 76. Gautam R, Carter AC, Katz N, Butler IF, Barnes M, et al. (2007) In vitro characterization of primary SIVsmm isolates belonging to different lineages. In vitro growth on rhesus macaque cells is not predictive for in vivo replication in rhesus macaques. Virology 362: 257–270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Mellors JW, Munoz A, Giorgi JV, Margolick JB, Tassoni CJ, et al. (1997) Plasma viral load and CD4+ lymphocytes as prognostic markers of HIV-1 infection. Ann Intern Med 126: 946–954. [DOI] [PubMed] [Google Scholar]
- 78. Mellors JW, Rinaldo CR Jr, Gupta P, White RM, Todd JA, et al. (1996) Prognosis in HIV-1 infection predicted by the quantity of virus in plasma. Science 272: 1167–1170. [DOI] [PubMed] [Google Scholar]
- 79. Apetrei C, Gautam R, Sumpter B, Carter AC, Gaufin T, et al. (2007) Virus-subtype specific features of natural SIVsmm infection in sooty mangabeys. J Virol 81: 7913–7923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Diop OM, Gueye A, Dias-Tavares M, Kornfeld C, Faye A, et al. (2000) High levels of viral replication during primary simian immunodeficiency virus SIVagm infection are rapidly and strongly controlled in African green monkeys. J Virol 74: 7538–7547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Onanga R, Kornfeld C, Pandrea I, Estaquier J, Souquiere S, et al. (2002) High levels of viral replication contrast with only transient changes in CD4+ and CD8+ cell numbers during the early phase of experimental infection with simian immunodeficiency virus SIVmnd-1 in Mandrillus sphinx . J Virol 76: 10256–10263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Onanga R, Souquiere S, Makuwa M, Mouinga-Ondeme A, Simon F, et al. (2006) Primary simian immunodeficiency virus SIVmnd-2 infection in mandrills (Mandrillus sphinx). J Virol 80: 3303–3309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Silvestri G, Fedanov A, Germon S, Kozyr N, Kaiser WJ, et al. (2005) Divergent host responses during primary simian immunodeficiency virus SIVsm infection of natural sooty mangabey and nonnatural rhesus macaque hosts. J Virol 79: 4043–4054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Gordon S, Klatt NR, Milush JM, Engram J, Dunham RM, et al. (2007) Severe depletion of mucosal CD4+ T cells in AIDS-free SIV-infected sooty mangabeys. J Immunol 179: 3026–3034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Pandrea I, Onanga R, Kornfeld C, Rouquet P, Bourry O, et al. (2003) High levels of SIVmnd-1 replication in chronically infected Mandrillus sphinx . Virology 317: 119–127. [DOI] [PubMed] [Google Scholar]
- 86. Silvestri G, Sodora DL, Koup RA, Paiardini M, O'Neil SP, et al. (2003) Nonpathogenic SIV infection of sooty mangabeys is characterized by limited bystander immunopathology despite chronic high-level viremia. Immunity 18: 441–452. [DOI] [PubMed] [Google Scholar]
- 87. Fiebig EW, Wright DJ, Rawal BD, Garrett PE, Schumacher RT, et al. (2003) Dynamics of HIV viremia and antibody seroconversion in plasma donors: implications for diagnosis and staging of primary HIV infection. AIDS 17: 1871–1879. [DOI] [PubMed] [Google Scholar]
- 88. Brenchley JM, Price DA, Douek DC (2006) HIV disease: fallout from a mucosal catastrophe? Nat Immunol 7: 235–239. [DOI] [PubMed] [Google Scholar]
- 89. Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, et al. (2006) Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nature Medicine 12: 1365–1371. [DOI] [PubMed] [Google Scholar]
- 90. Pandrea I, Gaufin T, Brenchley JM, Gautam R, Monjure C, et al. (2008) Experimentally-induced immune activation in natural hosts of SIV induces significant increases in viral replication and CD4+ T cell depletion. J Immunol 181: 6687–6691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Bosinger SE, Li Q, Gordon SN, Klatt NR, Duan L, et al. (2009) Global genomic analysis reveals rapid control of a robust innate response in SIV-infected sooty mangabeys. J Clin Invest 119: 3556–3572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Muller MC, Barre-Sinoussi F (2003) SIVagm: genetic and biological features associated with replication. Front Biosci 8: D1170–1185. [DOI] [PubMed] [Google Scholar]
- 93. Johnson PR, Gravell M, Allan J, Goldstein S, Olmsted RA, et al. (1989) Genetic diversity among simian immunodeficiency virus isolates from African green monkeys. J Med Primatol 18: 271–277. [PubMed] [Google Scholar]
- 94. Apetrei C, Kaur A, Lerche NW, Metzger M, Pandrea I, et al. (2005) Molecular epidemiology of SIVsm in US Primate Centers unravels the origin of SIVmac and SIVstm. J Virol 79: 8991–9005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Chen Z, Telfier P, Gettie A, Reed P, Zhang L, et al. (1996) Genetic characterization of new West African simian immunodeficiency virus SIVsm: geographic clustering of household-derived SIV strains with human immunodeficiency virus type 2 subtypes and genetically diverse viruses from a single feral sooty mangabey troop. J Virol 70: 3617–3627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Souquiere S, Bibollet-Ruche F, Robertson DL, Makuwa M, Apetrei C, et al. (2001) Wild Mandrillus sphinx are carriers of two types of lentivirus. J Virol 75: 7086–7096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Pandrea I, Parrish NF, Raehtz K, Gaufin T, Barbian HJ, et al. (2012) Mucosal SIV transmission in African green monkeys: Susceptibility to infection is proportional to target cell availability at mucosal sites. J Virol 86: 4158–4168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Ma ZM, Keele BF, Qureshi H, Stone M, Desilva V, et al. (2011) SIVmac251 is inefficiently transmitted to rhesus macaques by penile inoculation with a single SIVenv variant found in ramp-up phase plasma. AIDS Res Hum Retroviruses 27: 1259–1269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Andelman SJ (1986) Ecological and social determinants of cercopithecine mating patterns. In: Rubenstein DI, Wrangham RW, editors. Ecological Aspects of Social Evolution. Princeton: Princeton University Press. pp. 201–216.
- 100.Cords M (2000) The number of males in guenon groups. In: Kappeler P, editor. Primate Males: Causes and Consequences of Variation in Group Composition. Cambridge, UK: Cambridge University Press. pp. 84–96.
- 101. Keddy AC (1986) Female mate choice in vervet monkeys (Cercopithecus aethiops sabaeus). American Journal of Primatology 10: 125–134. [DOI] [PubMed] [Google Scholar]
- 102. Weingrill T, Willems EP, Krutzen M, Noe R (2011) Determinants of Paternity Success in a Group of Captive Vervet Monkeys (Chlorocebus aethiops sabaeus). International Journal of Primatology 32: 415–429. [Google Scholar]
- 103. Cowlishaw G, Dunbar RIM (1991) Dominance rank and mating success in male primates. Animal Behaviour 41: 1045–1056. [Google Scholar]
- 104. Andelman SJ (1987) Evolution of concealed ovulation in vervet monkeys (Cercopithecus aethiops). Am Naturalist 129: 785–799. [Google Scholar]
- 105. Nerrienet E, Amouretti X, Muller-Trutwin MC, Poaty-Mavoungou V, Bedjebaga I, et al. (1998) Phylogenetic analysis of SIV and STLV type I in mandrills (Mandrillus sphinx): indications that intracolony transmissions are predominantly the result of male-to-male aggressive contacts. AIDS Res Hum Retroviruses 14: 785–796. [DOI] [PubMed] [Google Scholar]
- 106. Cheney D, Seyfarth R (1983) Nonrandom dispersal in free-ranging vervet monkeys: Social and genetic consequences. Am Naturalist 122: 392–412. [Google Scholar]
- 107. Amedee AM, Rychert J, Lacour N, Fresh L, Ratterree M (2004) Viral and immunological factors associated with breast milk transmission of SIV in rhesus macaques. Retrovirology 1: 17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Chahroudi A, Meeker T, Lawson B, Ratcliffe S, Else J, et al. (2011) Mother-to-infant transmission of simian immunodeficiency virus is rare in sooty mangabeys and is associated with low viremia. J Virol 85: 5757–5763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Beer B, Denner J, Brown CR, Norley S, zur Megede J, et al. (1998) Simian immunodeficiency virus of African green monkeys is apathogenic in the newborn natural host. J Acquir Immune Defic Syndr Hum Retrovirol 18: 210–220. [DOI] [PubMed] [Google Scholar]
- 110. Ruprecht RM, Baba TW, Liska V, Ray NB, Martin LN, et al. (1999) Oral transmission of primate lentiviruses. J Infect Dis 179 Suppl 3: S408–412. [DOI] [PubMed] [Google Scholar]
- 111. Chenine AL, Siddappa NB, Kramer VG, Sciaranghella G, Rasmussen RA, et al. (2010) Relative transmissibility of an R5 clade C simian-human immunodeficiency virus across different mucosae in macaques parallels the relative risks of sexual HIV-1 transmission in humans via different routes. J Infect Dis 201: 1155–1163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Cheney D, Seyfarth R, Smuts B (1986) Social relationships and social cognition in nonhuman primates. Science 234: 1361–1366. [DOI] [PubMed] [Google Scholar]
- 113. Schwab P, Debes PV, Witt T, Hartl GB, Hmwe SS, et al. (2012) Genetic structure of the common impala (Aepyceros melampus melampus) in South Africa: pylogeography and implications for conservation. J Zool Syst Evol Res 50: 76–84. [Google Scholar]
- 114. Sithaldeen R, Bishop JM, Ackermann RR (2009) Mitochondrial DNA analysis reveals Plio-Pleistocene diversification within the chacma baboon. Mol Phylogenet Evol 53: 1042–1048. [DOI] [PubMed] [Google Scholar]
- 115. Drummond AJ, Suchard MA, Xie D, Rambaut A (2012) Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol Biol Evol 29: 1969–1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Pandrea I, Onanga R, Rouquet P, Bourry O, Ngari P, et al. (2001) Chronic SIV infection ultimately causes immunodeficiency in African non-human primates. AIDS 15: 2461–2462. [DOI] [PubMed] [Google Scholar]
- 117. Traina-Dorge V, Blanchard J, Martin L, Murphey-Corb M (1992) Immunodeficiency and lymphoproliferative disease in an African green monkey dually infected with SIV and STLV-I. AIDS Res Hum Retroviruses 8: 97–100. [DOI] [PubMed] [Google Scholar]
- 118. Ling B, Apetrei C, Pandrea I, Veazey RS, Lackner AA, et al. (2004) Classic AIDS in a sooty mangabey after an 18-year natural infection. J Virol 78: 8902–8908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Apetrei C, Gormus B, Pandrea I, Metzger M, ten Haaft P, et al. (2004) Direct inoculation of simian immunodeficiency virus from sooty mangabeys in black mangabeys (Lophocebus aterrimus): first evidence of AIDS in a heterologous African species and different pathologic outcomes of experimental infection. J Virol 78: 11506–11518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Etienne L, Nerrienet E, LeBreton M, Bibila GT, Foupouapouognigni Y, et al. (2011) Characterization of a new simian immunodeficiency virus strain in a naturally infected Pan troglodytes troglodytes chimpanzee with AIDS related symptoms. Retrovirology 8: 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Pandrea I, Cornell E, Wilson C, Ribeiro RM, Ma D, et al. (2012) Coagulation biomarkers predict disease progression in SIV-infected nonhuman primates. Blood 120: 1357–1366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Lederer S, Favre D, Walters KA, Proll S, Kanwar B, et al. (2009) Transcriptional profiling in pathogenic and non-pathogenic SIV infections reveals significant distinctions in kinetics and tissue compartmentalization. PLoS Pathog 5: e1000296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Lederman MM, Alter G, Daskalakis DC, Rodriguez B, Sieg SF, et al. (2010) Determinants of protection among HIV-exposed seronegative persons: an overview. J Infect Dis 202 Suppl 3: S333–338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Watkins DI, Burton DR, Kallas EG, Moore JP, Koff WC (2008) Nonhuman primate models and the failure of the Merck HIV-1 vaccine in humans. Nat Med 14: 617–621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Grobler P, Turner T (2010) A novel trap design for the capture and sedation of vervet monkeys (Chlorocebus aethiops). South Afr J Wildlife Res 40: 163–168. [Google Scholar]
- 126. Bibollet-Ruche F, Bailes E, Gao F, Pourrut X, Barlow KL, et al. (2004) A new simian immunodeficiency virus lineage (SIVdeb) infecting de Brazza's monkeys (Cercopithecus neglectus): Evidence for a Cercopithecus monkey virus clade. J Virol 78: 7748–7762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Gao F, Yue L, Robertson DL, Hill SC, Hui H, et al. (1994) Genetic diversity of human immunodeficiency virus type 2: evidence for distinct sequence subtypes with differences in virus biology. J Virol 68: 7433–7447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Jin MJ, Rogers J, Phillips-Conroy JE, Allan JS, Desrosiers RC, et al. (1994) Infection of a yellow baboon with simian immunodeficiency virus from African green monkeys: evidence for cross-species transmission in the wild. J Virol 68: 8454–8460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32: 1792–1797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, et al. (2010) New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 59: 307–321. [DOI] [PubMed] [Google Scholar]
- 131. Huson DH, Bryant D (2006) Application of phylogenetic networks in evolutionary studies. Mol Biol Evol 23: 254–267. [DOI] [PubMed] [Google Scholar]
- 132. Simon F, Souquiere S, Damond F, Kfutwah A, Makuwa M, et al. (2001) Synthetic peptide strategy for the detection of and discrimination among highly divergent primate lentiviruses. AIDS Res Hum Retroviruses 17: 937–952. [DOI] [PubMed] [Google Scholar]
- 133.Kuiken C, Foley B, Leitner T, Apetrei C, Hahn B, et al.. (2010) HIV Sequence Compendium. Los Alamos National Laboratory, New Mexico, LA-UR 10-03684: Theoretical Biology and Biophysics Group.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Maximum likelihood trees for env gene of the newly derived SIVagmVer sequences from wild vervets in South Africa. Maximum likelihood estimates were performed using 1000 replicate bootstrap analysis. Internal nodes indicate level of support values for internal branching. Sequences are colored depending on the region in which the sequences were sampled, with red indicating sequences from Free State, green indicating sequences from KwaZulu-Natal, and blue indicating sequences from Eastern Coast territories. In addition, sequences colored in gray are sampled from the Riverside Wildlife Rehabilitation and Environmental Education Centre (RWREC), Letsitele, Limpopo semifree colony that is housing released pets or animals with health problems, and their origin is unknown, and sequences colored in black indicate outgroup sequences. The strain nomenclature includes the identification number, monkey status (M-male, F-female; I-infant; J-juvenile and A-adult), and the site and the state of origin (FS-Free State; KZN-KwaZulu Natal; EC-Eastern Cape).
(TIFF)
SplitsTree showing the pol gene of South African SIVagmVer sequences. Nodes are colored by the region in which they were sampled, with red, blue and green denoting Free State, East Coast and KwaZulu-Natal, respectively. Potentially interesting recombinants have been circled, including VMT17083-IM-Gariep_FS and a cluster indicating possible mixing between East Coast and KwaZulu-Natal sequences.
(TIFF)
Comparative body mass index (BMI) assessment based on age and location in SIV-infected and SIV uninfected wild vervet monkeys ( Chlorocebus pygerithrus ) from South Africa.
(TIFF)
SIV prevalence in vervet monkeys from South Africa based on location of collected samples.
(DOC)
Age- and sex-related prevalence of SIVagmVer in semifree vervets from Riverside Wildlife Rehabilitation and Education Center, Letsitele, Limpopo.
(DOC)
Estimates of evolutionary divergence over sequence pairs between groups. The number of base substitutions per site±standard error estimate(s) from averaging over all sequence pairs between groups are shown± Analyses were conducted using the Maximum Composite Likelihood model [1]. The rate variation among sites was modeled with a gamma distribution (shape parameter = 0.5). env analysis involved 81 nucleotide sequences. All positions containing gaps and missing data were eliminated. There were a total of 822 positions in the final dataset for env. pol analysis involved 98 nucleotide sequences. All positions containing gaps and missing data were eliminated. There were a total of 574 positions in the final pol dataset. Evolutionary analyses were conducted in MEGA5 [2].
(DOC)
Most recent common ancestor (MRCA) estimates using relaxed molecular clocks to incorporate biogeographical assumptions. Calibration for the env gene is based on time to MRCA (TMRCA) of SIVagm sequences sampled around the Drakensberg Mountains, presumed to be 100,000–3,000,000 years of age. Calibration of the pol gene is based on TMRCA of SIVagm sequences as before, as well as SIVdrl sequences sampled from the Bioko Island of Equatorial Guinea, presumed to have been isolated for approximately 10,000 years. These estimates indicate that SIV is potentially considerably older than previous estimates, and refute timing estimates based exclusively on molecular clocks without calibration.
(DOC)
Details on animal origin, animal capture, sampling methodology and clinical assessment.
(DOC)