

Abstract
Chromosome 16 contains multiple copy number variations (CNVs) that predispose to genomic disorders. Here, we differentiate pathogenic duplications of 16p11.2–p12.2 from microscopically similar euchromatic variants of 16p11.2. Patient 1 was a girl of 18 with autism, moderate intellectual disability, behavioural difficulties, dysmorphic features and a 7.71-Mb (megabase pair) duplication (16:21 521 005–29 233 146). Patient 2 had a 7.81-Mb duplication (16:21 382 561–29 191 527), speech delay and obsessional behaviour as a boy and, as an adult, short stature, macrocephaly and mild dysmorphism. The duplications contain 65 coding genes of which Polo-like kinase 1 (PLK1) has the highest likelihood of being haploinsufficient and, by implication, a triplosensitive gene. An additional 1.11-Mb CNV of 10q11.21 in Patient 1 was a possible modifier containing the G-protein-regulated inducer of neurite growth 2 (GPRIN2) gene. In contrast, the euchromatic variants in Patients 3 and 4 were amplifications from a 945-kb region containing non-functional immunoglobulin heavy chain (IGHV), hect domain pseudogene (HERC2P4) and TP53-inducible target gene 3 (TP53TG3) loci in proximal 16p11.2 (16:31 953 353–32 898 635). Paralogous pyrosequencing gave a total copy number of 3–8 in controls and 8 to >10 in Patients 3 and 4. The 16p11.2–p12.2 duplication syndrome is a recurrent genomic disorder with a variable phenotype including developmental delay, dysmorphic features, mild to severe intellectual disability, autism, obsessive or stereotyped behaviour, short stature and anomalies of the hands and fingers. It is important to differentiate pathogenic 16p11.2–p12.2 duplications from harmless, microscopically similar euchromatic variants of proximal 16p11.2, especially at prenatal diagnosis.
Keywords: chromosomes, human, pair 16; chromosome duplication; 16p11.2–p12.2; DNA array; euchromatic variant; copy number variation
Introduction
The proximal short arm of chromosome 16 contains a number of copy number variation (CNV) hotspots1 that predispose to at least four reciprocal deletions and duplications within bands 16p11.2–16p12.2 (Figure 1). From centromere to telomere, these include first, the common microdeletions/duplications of ∼600 kb in 16p11.2 associated with neurocognitive difficulties and obesity;2, 3, 4 second, the microscopically visible duplications of 8–9 Mb from 16p11.2 to 16p12.1/2 associated with developmental delay and autism5, 6, 7, 8, 9, 10, 11 and the reciprocal microdeletions associated with developmental delay, intellectual disability and subtle dysmorphic features;12, 13 within these are, third, the distal (formerly atypical) microdeletions of ∼220 kb in 16p11.2 associated with a phenotype that includes developmental delay or obesity but extends into the normal range14, 15, 16 and, fourth, the microdeletions/duplications of ∼520 kb in 16p12.1 associated with developmental delay.1, 17 Here, we report two new patients with cytogenetically visible duplications of 16p11.2–16p12.2 analysed using oligonucleotide array comparative genomic hybridisation (oaCGH) and compared with 10 previous postnatal patients.5, 6, 7, 8, 9, 10, 11
Figure 1.

UCSC browser screenshot annotated with extent of the duplications, triplication, deletions and euchromatic variants of proximal 16p in the context of the UCSC browser bands and data on genomic variants and segmental duplications (NCBI 36/hg18): included are the 16p11.2–p21.2 duplications in Patients 1 and 2 (light grey bars), the 16p11.2–p21.2 duplications of Finelli et al8 (light grey bar), the 16p11.2–16p12.1 triplication (intermediate grey bar) and duplication (light grey bar) of Ballif et al, 10 subject 5, the duplications of Tabet et al,11 the reciprocal 16p11.2–p21.2 ‘micro'deletions (dark grey bar with narrow bars at either end indicating variable extent), the 16p12.1 microdeletions and duplications (diagonally hatched light and dark grey bars), the distal (formerly atypical) 16p11.2 microdeletions (dark grey bar), the common 16p11.2 microdeletions/duplications (diagonally hatched light and dark grey bars) and the euchromatic amplification variants in Patients 3 and 4 (dotted bars on a light grey background). The approximate start and end points of bands, imbalances, variants and candidate genes (CDR2, PLK1 and SH2B1) are given in Mb.
CNV is common with over 66 000 examples recorded in the Database of Genomic Variants (DGV) and an estimated 1% of the human population having a CNV >1 Mb.1 When copy number is high enough, rare CNVs of 8p23.1, 9p12, 9qh/q12, 9q13, 15q11.2 and 16p11.2 become visible in the light microscope and have been described as euchromatic variants.18 The euchromatic variants of 16p11.2 were originally defined using semi-quantitative cosmid fluorescence in situ hybridisation (FISH) and believed to result from constitutional cytogenetic amplification of paralogous sequences including non-functional IGVH, creatine transporter (SLC6A10P) and TP53TG3 genes.18, 19, 20 Euchromatic variants of 16p11.2 can be mistaken for pathogenic duplications resulting in a termination of pregnancy in at least one documented case.20 Here, we report two new cases in which bacterial artificial chromosome (BAC) FISH was used to assign the amplicons to an ∼1-Mb region of proximal 16p11.2. Pyrosequencing21 was used to quantify amplicon copy number and corroborate estimates made using semi-quantitative FISH in euchromatic variant carriers and controls.
Materials and methods
Conventional G-banded chromosome analysis was carried out in lymphocytes (Patients 1 and 2) and amniotic fluid cells (Patients 3 and 4) using standard techniques. FISH was carried out with non-functional IGVH (cos11, 13, 33, 97 and 98) and creatine transporter (c329B6) cosmids as before.19 Dual colour BAC FISH and oaCGH with a customised 4 × 44K Agilent oligonucleotide array (Santa Clara, CA, USA) were performed as described previously.22 Base pair coordinates correspond to NCBI release 36.1 (hg18). Copy number in euchromatic variant carriers and normal controls was determined by co-amplifying paralogous sequences at 16p11.2 and Xq2823 and quantifying the paralogous sequence mismatches (PSM) between them using pyrosequencing according to the manufacturer's instructions. The resultant PSM allele frequencies were calculated using the Pyrosequencing AQ software (Biotage AB, Uppsala, Sweden) and reflect the relative frequency of the paralogous sequence on each chromosome.
Clinical reports
Patient 1
This 18-year-old girl (Figures 2a–e) was the first child of healthy unrelated Caucasian parents. She had a younger brother aged 15 with normal development. The pregnancy was complicated by hyperemesis. She was born at 36 weeks gestation by spontaneous vaginal delivery weighing 2920 g (90th centile). Discharge was delayed by 5 days because of neonatal jaundice. She was a poor feeder owing to poor suck and also needed waking for feeds. She had poor weight gain and lactose intolerance. Weaning to solid food was slow and achieved by 3 years of age. Developmentally she sat at 8 months and walked at 13 months. By 18 months she had four words with meaning. During infancy, she was asthmatic and prone to respiratory tract infections but had outgrown this by the age of 9. She had otitis media and grommets inserted at the age of 3. In childhood she was a poor sleeper but this improved after the age of 12. She had vacant staring episodes but no seizures and her EEG was normal. She was toilet trained aged 7 years. She was small for her age until the age of 12 but has since caught up. She had onset of menarche aged 14 years. She has had orthodontic treatment since the age of 15 years. A Ruth Griffiths assessment at chronological age 51 months showed mental age results ranging from 28 to 40 months (sub quotient 55–78). From the age of 5 years she attended a special school for moderate learning difficulties. Since the age of 16 years she has been attending a transition college and doing a life skills course. She can dress herself, but needs help with small buttons.
Figure 2.

(a–e) The phenotype of Patient 1 aged 15 years: (a) Facial features showing heavy eyebrows, synophrys, slight ptosis, left convergent squint, broad nasal bridge, bow shaped upper lip and full lower lip; (b) lateral facial features showing overfolded helix, prominent glabella, upturned nose with a slightly bulbous tip and full everted lower lip; (c) oral features showing prominent upper incisors and dental crowding; (d) hands of the proband showing brachydactyly and squared off finger tips; and (e) feet of the proband showing bilateral medial protuberances near her ankles.
From early on her behaviour was described as fixed and repetitive. She did not like plans to be changed. She would prepare well in advance for any event out of her normal routine and become fixated on it. She fixated on certain ideas or phrases and would repeat a phrase over and over to herself. She generally enjoyed meeting new people and talked very quickly to them without accounting for their lack of prior knowledge. She could be inappropriately familiar and affectionate with strangers. At other times she would happily entertain herself and not interact with others.
Clinical evaluation at the age of 15 revealed a weight of 48 kg (<25th centile), height of 158.2 cm (<25th centile) and an OFC of 55 cm (<50th centile). She had a prominent glabella, heavy eyebrows, synophrys, slight ptosis, left convergent squint, broad nasal bridge, an upturned nose with a slightly bulbous tip, bow shaped upper lip, full everted lower lip, prominent upper incisors, dental crowding and overfolded helices (Figures 2a–c). She had tapering, hypermobile fingers with brachydactyly, squared off finger tips (Figure 2d) and prominent foetal finger pads. She has had problems with painful flat feet and had bilateral medial bony protuberances near the ankles for which she has worn orthotics (Figure 2e). Her skin was soft and velvety with wrinkled skin on the palms and soles. Her hands were clammy. She had a postural kyphosis. Neurological examination revealed poor balance and coordination.
She had an autism diagnostic interview-revised (ADI-R) assessment and observations by an occupational therapist, an art therapist and a consultant psychiatrist at the age of 16 years and 9 months and the results confirmed the diagnosis of autism. The assessment showed difficulties with non-verbal regulation of social interaction, difficulty in developing peer relationships and a lack of shared enjoyment and socio-emotional reciprocity. She had difficulty initiating and sustaining conversation in a reciprocal way, some stereotyped and repetitive speech and did not have the social awareness to avoid asking inappropriate things at inappropriate times. The assessment showed that she had more difficulties with minor changes in routine or environment when she was younger compared with when she was assessed. Her parents had made a great deal of effort to avoid any changes of her routine to prevent distress. She has become more aware of her difficulties with time and she gets frustrated at not being able to do things alone. As a result, she has been verbally and physically aggressive towards her brother and peers at times. At college, one to one support is being put in place to try and help her behaviour. She enjoys reading and writing.
Patient 2 (Database of Chromosomal Imbalance and Phenotype in Humans Using Ensembl Resources (DECIPHER) 2131)
This male patient was referred at the age of 45 so that his elderly parents could plan for his future. He had been a full-term normal delivery with a good birth weight and no very early health concerns. He was a little late to walk but his speech was markedly delayed and a diagnosis of glue ear was made. He attended special school from the age of 6 after an extra year at nursery school. He was a generally good natured boy with some obsessional behaviour. He had an abnormal fixation on buses and trucks, which involved large amounts of time engaged in spinning the wheels, but is not regarded as having had autism. He can now read well but cannot write more than a poor rendition of his name and is not good with numbers. He was unable to pass any exams but does have a number of skills such as excellent map-reading ability. He was generally healthy as a child but did have one episode of unexplained hypercalcemia. Since leaving school he has had jobs in sheltered employment. As an adult he has been well other than recurrent rectal prolapse. On examination, he is short at 158 cm (0.4th centile), has an OFC of 59 cm (98th centile) and is of average build. He has a wide mouth, a large furrowed tongue and broad alveolar margins. His hands are broad with a generalised brachydactyly and his digits are untapered. He has one large lipoma on his back and freckling on the palms and soles. A diagnosis of the Bannayan–Riley–Ruvalcaba syndrome was considered but molecular analysis of the phosphatase and tensin homologue (PTEN) gene was negative.
Patient 3
This healthy 43-year-old woman was in her third pregnancy and delivered a boy with normal measurements at the fortieth week of pregnancy (weight 3160 g, length 54 cm, OFC 34 cm and APGAR scores 10/10/10). The healthy boy is now 6 years old and will start school in the autumn of this year.
Patient 4
This patient was referred for prenatal diagnosis after two miscarriages. No further clinical details were available.
Results
Patient 1
An extra G-dark band was present in the proximal short arm of chromosome 16 (Figure 3a). FISH with non-functional IGVH (cos33 and 98) and creatine transporter cosmids (c329B6) from proximal 16p11.219 was normal but signals with BAC CTB2515A14 from 16p12.1 were duplicated (data not shown) (Table 1). OaCGH confirmed a duplication of 16p12.2–16p11.2 with a minimum size of 7.71 Mb and a maximum of 8.07 Mb (Figure 4e) (Table 1). An additional duplication CNV of band 10q11.21 had a minimum size of 1.11 Mb and a maximum of 2.47 Mb. Parental chromosomes were normal. The karyotype of the proband was: 46,XX,dup (16) (p11.2–p12.2)dn.ish dup (16) (p11.2–12.2)(CTB2515A14++,CTB74E23+RP11-165M2+).arr 16p12.2p11.2(21 507 188 × 2 21 521 005-29 233 146 × 3 29 581 455 × 2), 10q11.21 (45 480 521 × 2 46 371 243-47 485 737 × 3 47 946 216 × 2).
Figure 3.
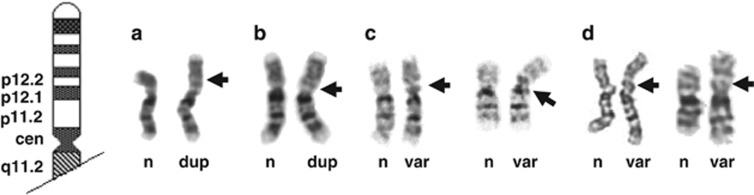
(a–d) G-banded partial karyotypes from Patients 1, 2, 3 and 4 with the normal chromosome (n) on the left and the duplicated (dup) or euchromatic variant (var) chromosome 16 on the right: (a) Patient 1 and (b) Patient 2 with the duplicated region of chromosome 16 indicated by the large black arrows; (c) Patient 3 and (d) Patient 4 with the euchromatic variant region of chromosome 16 indicated by the large black arrows. Note the similarity between the G-banded duplications in the Patients 1 and 2 (a, b) and the euchromatic variants in Patients 3 and 4 (c, d).
Table 1. FISH and oaCGH results.

Figure 4.

(a–f) Dual colour BAC FISH results and oaCGH results: (a) Distinct pairs of signals from RP11-142A12 (red arrows) indicating the duplication in Patient 2; (b) enh (large red arrow) and normal signals (small red arrow) with RP11-408D2 (red) consistent with amplification in a metaphase cell from Patient 3; (c) increased interval (blue arrows) between BAC RP11-67I10 (red) and the 16q11.2 microdissection probe (green) in a pair of chromosomes 16 from Patient 4; (d) enh (large red arrows) and normal signals (small red arrows) with RP11-408D2 (red) consistent with amplification in interphase and metaphase cells from Patient 4 (the centromeres of chromosomes 1, 5 and 19 have also been labelled in green in these images); (e) and (f) oaCGH results from Patients 1 (e) and 2 (f) analysed with the Agilent Analytics software. The vertical coloured bars and background represent the extent of the duplications adjacent to idiograms of chromosome 16 in the left hand panels, magnified in the right hand panels, with black dots representing oligonucleotides with normal copy number and red dots oligonucleotides with increased copy number. The colour reproduction of this figure is available at the European Journal of Human Genetics journal online.
Patient 2 (DECIPHER 2131)
An extra G-dark band was present in the proximal short arm of chromosome 16 (Figure 3b) and duplicated signals were found using FISH with BAC RP11-142A12 from distal 16p12.1 (Figure 4a) (Table 1). OaCGH confirmed a duplication of 16p12.2–16p11.2 (Figure 4a) with a minimum of 7.81 Mb and a maximum of 8.37 Mb (Table 1). An additional duplication CNV of 1.52 Mb from Xp22.31 was found with a minimum size of 1.52 Mb. Parental chromosomes were normal and the karyotype of the proband was: 46,XY,dup (16) (p11.2–p12.2) dn.ish dup (16) (p11.2–12.2) (RP11-142A12,RP11-165M2+).arr 16p12.1p11.2 (21 209 438 × 2 21 382 561-29 191 527 × 3 29 581 455 × 2), Xp22.31(6 467 403-7 992 061) × 3.
Patient 3
An extra GTG-positive band in proximal 16p was found during chromosome analysis of amniotic fluid cultures and lymphocytes from the mother (Figure 3c). The extra band was CBG negative but enhanced (enh) signals were seen using FISH with IGVH cosmids (cos11, 13, 33, 97 and 98) (data not shown)19 Enh signals were also seen with BACs RP11-408D2 (Figure 4b) and RP11-378C4. The results with BAC RP5-1044J9 were less convincing than those in Patient 4 implying an amplicon of 692 kb (Table 1). The duplicated signals with BAC RP11-410P5 are likely to represent independent variation as they are non-contiguous. The karyotype was 46,XX,var (16) (p11.2p11.2) mat.ish var (16) (RP11-410P5++,RP11-196G11+,RP5-1044J9?enh,RP11-408D2enh,RP11-378C4enh,RP11-488I20+,RP11-80F22+,RP11-65B9+). Pyrosequencing gave a total diploid copy number of 8 (Supplementary Figure 1c) and, assuming that the normal chromosome had the median copy number of 2, the euchromatic variant chromosome would have at least six copies of the amplicon spanning 4.2 Mb.
Patient 4
An extra GTG-positive band was found during chromosome analysis of amniotic fluid cultures (Figure 3d). No information on the karyotypes of the parents was available. Enh signals were seen using FISH with IGVH cosmids (cos11, 33 and 98; data not shown)19 and an increased interval between the 16p BAC RP11-67I10 and a 16q11.2-microdissection probe (Figure 4c). Enh signals were also found with BACs RP5-1044J9, RP11-408D2 and RP11- 378C4 (Figure 4d) (Table 1) implying an amplicon of 945 kb. Enh signals from non-contiguous BACs RP11-410P5 and RP11-80F22 (Table 1) were thought to be independent variation especially as RP11-80F22 is one of the commonest CNVs found using diagnostic oaCGH.24 The formal karyotype was: 46,XY,var(16) (p11.2p11.2). ish var (16) (RP11-410P5enh,RP11-196G11+,RP5-1044J9enh,RP11-408D2enh,RP11-378C4enh,RP11-488I20+,RP11-80F22enh,RP11-65B9+). Pyrosequencing gave a total diploid copy number of >10 (Supplementary Figure 1d) and, if the normal chromosome had the median copy number of 2, the euchromatic variant chromosome would have at least eight copies of the amplicon spanning 7.6 Mb.
Copy number in controls
Paralogous pyrosequencing gave total copy numbers of between three and eight in 46 controls (20 males and 26 females) with a median value of four (Supplementary Figures 1a and b).
Discussion
Our results indicate how similar large single-copy duplications of 16p11.2–p12.2 and amplification variants of 16p11.2 appear under the light microscope (Figures 3a–d) and how distinct they are at the molecular level (Figure 1). The importance of differentiating duplications from variants has been illustrated by the termination of a pregnancy when a euchromatic variant was misinterpreted as a duplication.20 The presence or absence of the pathogenic duplication can be tested using FISH, qPCR or microarrays (Table 1). The same methods can be used to obtain results consistent with a euchromatic variant (Figures 4a–d), but care is needed in the interpretation of results from this copy number variable region, qPCR can be sensitive to template DNA quality and targeted arrays may lack adequate coverage and require careful choice of controls. Accurate determination of copy numbers higher than 10 remains problematic even with the pyrosequencing assay used here. The less likely possibility of an unbalanced insertion would require a combination of techniques. Both pathogenic duplications of 16p11.2–p12.1 and euchromatic variants of 16p11.2 have been directly transmitted from parents to children but one of the parents has been affected, to some extent, in all the duplications.7
16p11.2–p21.2 duplication phenotype
Table 2 summarises the differing phenotypic consequences in Patients 1 and 2 together with seven individuals with duplications of 16p11–16p13,5, 6, 7, 8, 9 monozygotic twins with a slightly larger duplication11 and a patient with a triplication and duplication (Ballif et al,10 subject 5). A prenatal case was excluded as no sonographic anomalies were found before termination of the pregnancy.25 Only developmental and/or psychomotor delay was present in all patients. In one patient and the twins, normal development was followed by a marked decline after the age of 2. Intellectual disability in nine patients varied from severe in four to mild or moderate in others. Autism or autistic behaviour was recorded in eight patients but only Patient 1, the twins and one other patient had a formal ADI-R or DSM IV diagnosis. Patient 2 was not thought to have autism as a child. Obsessive and/or stereotyped behaviour was found in seven patients, social problems in six, ADHD or hyperactivity in four and echolalia in three. MRI/CT scan anomalies were noted in four patients and epilepsy, seizures and/or EEG anomalies in three.
Table 2. Phenotypic features found in patients with postnatal duplications of proximal 16p.
| Features | Case 15 | Case 25 | Reference 6 | Mother 7 | Daughtera,7 | P18 | Reference 9 | Subject 510 | P2,3b,11 | Present P1 | Present P2 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Trisomic region | 16p11–p13 | 16p11–p13 | 16p11.2–p12 | 16p11.2–p12.1 | 16p11.2–p12.1 | 16p11.2–p12.2 | 16p11.2–p13.1 | 16p11.2–p12.2 | 16p11.2-p12.2 | 16p11.2-p12.2 | 16p11.2-p12.2 |
| ∼Extent (Mb) | − | − | − | 8 | 8 | 8 | − | 6 trp;1 dup | 8.95 | 7.71-8.07 | 7.81-8.37 |
| Gestation (weeks) | 35 | Full term | Full term | Full term | Full term | Full term | Full term | 36 | Full term | ||
| Birth weight (g) | 2200 | 2630 | 3700 | 4050 (>p97) | 2550 (p3) | 2250 | 2450 (p4) | 2920 (p90) | |||
| Sex (M/F) | F | F | M | F | F | M | F | M | |||
| Age at examination (years) | 2 | 35, 39 and 40 | 5 | 3, 25 | 1.5, 4 | 11 | 17 | 15 | 45 | ||
| Height (cm) | 158 (<p3) | 103 (p3) | 151 (<p3) | NR | −3 to -4 SDS | 173 (mean) | 158 (<p25) | p0.4 | |||
| Weight (kg) | − | 16.3 (p50) | 42 (<p3) | NR | −2 SDS | 49 (−2 SDS) | 48 (< p25) | ||||
| OFC | Micro | 52.5 (<p3) | 49 (p3) | 54 (p25) | Micro | −1SDS | 55 (−1 SDS) | 55 (< p50) | Macro | ||
| Dev/psychomotor delay | + | + | + | LD + | After 24/12 | + | + | ++ | After 26/12 | + | + |
| Autism | Behaviour | DSM-IV | Partial | + | + | − | DSM-IV | ADI-R | − | ||
| Obesessive behaviours | + | + | + | + | + | ||||||
| Stereotyped behaviours | + | − | + | + | + | ||||||
| Echolalia | + | + | + | ||||||||
| Social problems | + | + | + | + | + | ||||||
| ADHD | + | + | H | ||||||||
| Intellectual disability | Mild | Mild | Severe | + | Severe | Severe | Moderate | + | |||
| IQ | 83 | 80 | 25−>39 | 42 | |||||||
| MRI/CT anomalies | − | + | + | + | |||||||
| EEG anomalies | − | + | − | − | + | − | |||||
| Epilepsy/seizures | − | + | + | − | − | + | − | − | |||
| Ataxia | + | + | |||||||||
| Hypotonia | + | − | − | − | − | − | + | − | |||
| Dysmorphism | + | + | + | None | − | + | + | + | + | + | + |
| Depressed/large/broad nasal bridge | + | + | + | − | − | + | + | + | + | + | |
| Palpebral fissures | Upslant | Narrow | − | Upslant | Upslant | Narrow | Upslant | − | − | ||
| Hypertelorism | + | + | − | − | + | + | + | + | |||
| Long philtrum | + | + | + | − | − | − | + | + | − | ||
| Round face | − | + | − | − | − | + | + | − | |||
| Prominent glabella | − | − | − | + | + | + | |||||
| Anteverted nares | + | − | − | − | + | + | + | ||||
| Upper lip anomaly | Thin | − | − | Large | Thin | Thin | − | ||||
| Low set ears | + | + | + | − | − | − | + | − | |||
| Short neck | + | + | − | − | + | − | |||||
| Micro/retrognathia | + | − | − | − | − | − | + | + | − | + | |
| Nystagmus/strabismus | + | + | + | ||||||||
| Finger/hand anomalies | + | + | − | − | − | + | + | + | + | Brachyd | |
| Skin syndactyly of feet | − | + | − | − | − | − | + | + | − | ||
| Recurrent infections | + | Early on | Early on | ||||||||
| Other single features | AD; EF; CD | CF | CPC; HT | HA | EV; MH; NV; VSD | FT; L; MST |
Abbreviations: AD, anxiety disorder; Brachyd, Brachdactyly; CD, cervical dystonia; CF, club feet; CPC, cryptic cleft palate; DSM-IV, diagnostic and statistical manual of mental disorders, fourth edition; EF, emotional flatness; EV, everted lower lip; F, female; FT, furrowed tongue; H, hyperactvity; HA, hyperacusis; HT, hypoplastic thumb; L, lipomata; LD, learning difficulties; M, male; macro, macrocephaly; MH, muscular hypotrophy; micro, microcephaly; MST, macrostomia; NV, non-verbal; P, Patient; p3, third centile etc; SD, sternum deformity; SDS, SD; VSD, ventral septal defect (in twin P2 that spontaneously closed); +, present; −, absent.
Greater than 10: chromosome X=12.8% (two copies), chromosome 16=87.2% (>10 copies).
Data added from Finelli et al ,8 Patient 2.
P2,3=monozygotic twins of Tabet et al,11 Patients 2 and 3.
Dysmorphism was seen in 10 patients but absent in one mother and daughter diad.7 Common dysmorphisms included a depressed, broad or large nasal bridge, upslanting or narrow palpebral features, hypertelorism and a long or tented philtrum. Short stature was found in five patients and microcephaly in four but normal OFC was found in Patient 1 and macrocephaly in Patient 2. Tapering, long, short and/or hypermobile fingers were found in eight patients. Patient 1 also had the prominent fingertip pads noted before (Ballif et al,10 subject 5) as well as brachydactyly and medial bony protuberances on both feet. Skin syndactyly of the feet was seen in three patients. Recurrent infections and nystagmus or strabismus were found in four patients each. Only one of the twins had a congenital heart defect (VSD) that resolved spontaneously.
Candidate genes
A minimum of 65 known coding genes are common to the duplication intervals in Patients 1 and 2 of which 11 are OMIM Morbid genes. Of these 11, many are autosomal recessive and only 3 have an estimated haploinsufficiency likelihood score (HLS), with implied dosage sensitivity, of <10%.26 These include the sarcoplasmic/endoplasmic reticulum calcium ATPase 1 (ATP2A1) gene (8.8%), associated with autosomal recessive Brody myopathy, and the Amiloride-sensitive sodium channel subunits β and γ genes SCNN1B (9.2%) and SCNN1G (8.8%) associated with the autosomal dominant hypertensive disorder Liddle syndrome. Of the other 54 genes, the Polo-like kinase 1 (PLK1) gene has the lowest HLS of 0.4% and no other gene has an HLS of <10%. PLK1 is a protein kinase superfamily member and the human form of Drosophila polo, which is an important cell cycle regulator implicated in mammals as both oncogene and tumour suppressor. Polo mutants can switch development from neurons to additional neuroblasts in Drosophila,27 which suggests that dosage of PLK1 might conceivably have an effect on neuronal development and microcephaly in humans.
Despite higher HLS scores, the cerebellar degeneration-related protein 2 (CDR2) gene (46.9%) is a candidate for the 16p12.1 microdeletion syndrome28 and might contribute to the developmental or behavioural problems when duplicated. Similarly, duplication of the Ataxin-2-like protein (ATXNL2) gene (31.5%) might be related to the ataxia seen in Patient 1 and Case 1 of Cohen et al.5 The SH2B adapter protein 1 (SH2B1) gene (19.2%) is a candidate for the developmental delay and obesity associated with the 200-kb microdeletions of 16p11.2. This gene lies within the minimum extent of the present and published 16p11.2–p12.2 duplications (Figure 1)15 but not within the triplication that has a more severe but similar phenotype to the duplications reported here (Ballif et al, 10 subject 5).
Modifiers or second hits
The phenotype of patients with 520-kb microdeletions of 16p12.1 was more severe when a second large CNV (gain or loss >500 kb) was present and these microdeletions are thought to predispose to neuropsychiatric phenotypes in the presence of other large deletions or duplications.17, 28 The genes within the remaining 7.2–7.3 Mb minimum interval in Patients 1 and 2 may therefore be sufficient to explain the cognitive phenotype in 16p11.2–p12.2 duplication syndrome but both patients had additional CNVs >500 kb in size. In the female Patient 1 with autism, the additional 1.11-Mb duplication CNV of 10q11.2 overlaps multiple CNVs in the DGV but also contains a minimum of 9 genes. Of these, duplication of the G-protein-regulated inducer of neurite growth 2 (GPRIN2) gene is a candidate modifier of severity in Rett syndrome29 and might conceivably interact with other candidate autism genes in 16p11.2–p12.2 duplications. In the male Patient 2 without autism, an additional duplication CNV of 1.52 Mb in Xp22.31 contains a minimum of 5 genes, including the steroid sulfatase (STS) gene, but there is growing evidence that duplication CNVs containing STS are benign population variants.30, 31 The inheritance pattern of these two CNVs was not determined.
Breakpoints
The breakpoints mapped in the present patients with oaCGH and the previously reported patients with BAC- or M-FISH8, 9, 10, 11 correspond broadly to clusters of segmental duplications in proximal 16p (Figure 1). In both their families, Finelli et al8 mapped the telomeric breakpoints to BAC RP11-98D10 in 16p12.2 (16:21 354 957–21 532 536) and the centromeric breakpoints to BAC RP11-368N21 in 16p11.2 (16:29 408 797-29 609 842). These authors also suggested that the duplications are mediated by non-allelic homologous recombination (NAHR) between a low copy repeat (LCR16v) in 16p11.2 and 16p12.2 that contains a truncated paralogue of the SLCL7A5 gene from 16q24.3.8 Both the centromeric breakpoints in the present the Patients 1 and 2 lie within an interval of 390 kb in 16p11.2 (Table 1) that overlaps with BAC 368N21 and contains the SLCL7A5P1 gene. The telomeric breakpoints lie within different 173 kb and 13.8 kb intervals in 16p12.2 (Table 1) that overlap with BAC 98D10 but do not contain a copy of the SLCL7A5P1 gene or, in the case of Patient 1, correspond to any known segmental duplication. Similar variation has been recorded in the reciprocal deletion syndrome11, 12, 13 (Figure 1) but further high-resolution analysis of the breakpoints will be needed to establish whether all recurrent deletions and duplications of 16p11.2–16p12.2 arise by NAHR between a variety of repeats and/or via the more complex mechanisms being found in other genomic disorders.32
Constitutional euchromatic amplification variants of 16p11.2
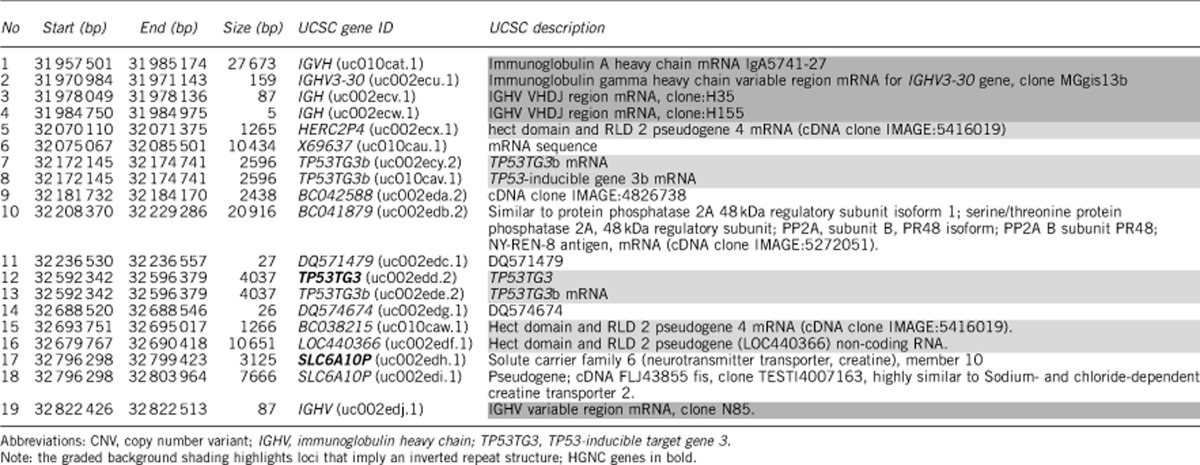
Including the present patients, euchromatic variants of 16p11.2 have been described in 14 families18, 19 and have no known phenotypic consequences. The amplicons in Patients 3 and 4 have been approximately mapped with FISH (Table 1) and come from a 945-kb region of proximal 16p11.2 (16:31 953 353–32 898 635) that contains 19 UCSC (University of California Santa Cruz) sequences of which only two are HUGO Gene Nomenclature Committee (HGNC) genes (Table 2). The region is flanked by non-functional immunoglobulin heavy chain (IGHV) genes, with hect domain pseudogenes (HERC2P4) and TP53-inducible target gene 3 (TP53TG3) protein loci that imply an asymmetrical palindromic structure (Table 3) that might mediate amplification.33
Table 3. UCSC genes within the 945-kb euchromatic CNV region of 16p11.2 (16:31 953 353–32 898 635) (hg18).
A diploid copy number of 4 in normal individuals and 10–12 in 16p11.2 euchromatic variant carriers was previously estimated using semi-quantitative cosmid FISH.19 The results of paralogous pyrosequencing (previously reported in abstract34) showed a wider range of copy number from 3 to 8 in controls and 8 to >10 in 16p11.2 euchromatic variant carriers. We could not quantify copy numbers greater than 10 but accurate, cost effective and high-throughput methods are needed to determine the possible effect of large CNVs on quantitative phenotypic traits in large cohorts.35
Conclusions
Our results indicate that the 16p11.2–p12.2 duplication syndrome is a recurrent genomic disorder characterised by developmental delay, autistic and/or repetitive behaviour, dysmorphic features, microcephaly, short stature and tapering fingers. Duplications of 16p11.2–p12.1 can be mistaken for harmless euchromatic variants of proximal 16p11.2 in the light microscope but are distinct at the molecular level and can be distinguished using FISH or aCGH.
Acknowledgments
We thank all four families concerned as well as Dr Christine Garrett and Dr Kay MacDermot for their clinical input on Patient 1. We are grateful to Dr Petra Muschke (Magdeburg) for the information on Patient 3 and for contacting her for the second blood sampling. We thank Dr Ilona Dietze-Armana for sending information on Patient 4. We are also grateful to Professor Hunt Willard for the D16Z2 centromeric probe. VH, SH and HW were supported by the UK Department of Health as part of the National Genetics Reference Laboratory (Wessex).
WEB RESOURCES
DGV: http://projects.tcag.ca/variation/
DECIPHER v5.0: https://decipher.sanger.ac.uk/
Ensembl Genome Browser: http://www.ensembl.org/Homo_sapiens/Info/Index
UCSC Genome Browser: http://genome.ucsc.edu/
The authors declare no conflict of interest.
Footnotes
Supplementary Information accompanies the paper on European Journal of Human Genetics website (http://www.nature.com/ejhg)
Supplementary Material
References
- Itsara A, Cooper GM, Baker C, et al. Population analysis of large copy number variants and hotspots of human genetic disease. Am J Hum Genet. 2009;84:148–161. doi: 10.1016/j.ajhg.2008.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghebranious N, Giampietro PF, Wesbrook FP, et al. A novel microdeletion at 16p11.2 harbors candidate genes for aortic valve development, seizure disorder, and mild mental retardation. Am J Med Genet Part A. 2007;143A:1462–1471. doi: 10.1002/ajmg.a.31837. [DOI] [PubMed] [Google Scholar]
- Weiss LA, Shen Y, Korn JM, et al. Association between microdeletion and microduplication at 16p11.2 and autism. N Engl J Med. 2008;358:667–675. doi: 10.1056/NEJMoa075974. [DOI] [PubMed] [Google Scholar]
- Walters RG, Jacquemont S, Valsesia A, et al. A new highly penetrant form of obesity due to deletions on chromosome 16p11.2. Nature. 2010;463:671–675. doi: 10.1038/nature08727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen MM, Lerner C, Balkin NE. Duplication of 16p from insertion of 16p into 16q with subsequent duplication due to crossing over within the inserted segment. Am J Med Genet. 1983;14:89–96. doi: 10.1002/ajmg.1320140114. [DOI] [PubMed] [Google Scholar]
- Carrasco Juan JL, Cigudosa JC, Otero Gomez A, et al. De novo trisomy 16p. Am J Med Genet. 1997;68 (219-221 doi: 10.1002/(sici)1096-8628(19970120)68:2<219::aid-ajmg19>3.3.co;2-a. [DOI] [PubMed] [Google Scholar]
- Engelen JJ, de Die-Smulders CE, Dirckx R, et al. Duplication of chromosome region (16) (p11.2->p12.1) in a mother and daughter with mild mental retardation. Am J Med Genet. 2002;109:149–153. doi: 10.1002/ajmg.10287. [DOI] [PubMed] [Google Scholar]
- Finelli P, Natacci F, Bonati MT, et al. FISH characterisation of an identical (16) (p11.2p12.2) tandem duplication in two unrelated patients with autistic behaviour. J Med Genet. 2004;41:e90. doi: 10.1136/jmg.2003.016311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behjati F, Shafaghati Y, Firouzabadi SG, et al. M-banding characterization of a 16p11.2p13.1 tandem duplication in a child with autism, neurodevelopmental delay and dysmorphism. Eur J Med Genet. 2008;51:608–614. doi: 10.1016/j.ejmg.2008.06.007. [DOI] [PubMed] [Google Scholar]
- Ballif BC, Hornor SA, Jenkins E, et al. Discovery of a previously unrecognized microdeletion syndrome of 16p11.2-p12.2. Nat Genet. 2007;39:1071–1073. doi: 10.1038/ng2107. [DOI] [PubMed] [Google Scholar]
- Tabet AC, Pilorge M, Delorme R, et al. Autism multiplex family with 16p11.2p12.2 microduplication syndrome in monozygotic twins and distal 16p11.2 deletion in their brother. Eur J Hum Genet. 2012;20:540–546. doi: 10.1038/ejhg.2011.244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Battaglia A, Novelli A, Bernardini L, et al. Further characterization of the new microdeletion syndrome of 16p11.2-p12.2. Am J Med Genet Part A. 2009;149A:1200–1204. doi: 10.1002/ajmg.a.32847. [DOI] [PubMed] [Google Scholar]
- Hempel M, Rivera Brugués N, Wagenstaller J, et al. Microdeletion syndrome 16p11.2-p12.2: clinical and molecular characterization. Am J Med Genet Part A. 2009;149A:2106–2112. doi: 10.1002/ajmg.a.33042. [DOI] [PubMed] [Google Scholar]
- Bijlsma EK, Gijsbers AC, Schuurs-Hoeijmakers JH, et al. Extending the phenotype of recurrent rearrangements of 16p11.2: deletions in mentally retarded patients without autism and in normal individuals. Eur J Med Genet. 2009;52:77–87. doi: 10.1016/j.ejmg.2009.03.006. [DOI] [PubMed] [Google Scholar]
- Bachmann-Gagescu R, Mefford HC, Cowan C, et al. Recurrent 200-kb deletions of 16p11.2 that include the SH2B1 gene are associated with developmental delay and obesity. Genet Med. 2010;12:641–647. doi: 10.1097/GIM.0b013e3181ef4286. [DOI] [PubMed] [Google Scholar]
- Barge-Schaapveld DQCM, Maas SM, Polstra A, et al. The atypical 16p11.2 deletion: A not so atypical microdeletion syndrome. Am J Med Genet Part A. 2011;155:1066–1072. doi: 10.1002/ajmg.a.33991. [DOI] [PubMed] [Google Scholar]
- Girirajan S, Rosenfeld JA, Cooper GM, et al. A recurrent 16p12.1 microdeletion supports a two-hit model for severe developmental delay. Nat Genet. 2010;42:203–209. doi: 10.1038/ng.534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barber JC. Directly transmitted unbalanced chromosome abnormalities and euchromatic variants. J Med Genet. 2005;42:609–629. doi: 10.1136/jmg.2004.026955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barber JC, Reed CJ, Dahoun SP, et al. Amplification of a pseudogene cassette underlies euchromatic variation of 16p at the cytogenetic level. Hum Genet. 1999;104:211–218. doi: 10.1007/s004390050938. [DOI] [PubMed] [Google Scholar]
- Lopez Pajares I, Villa O, Salido M, et al. Euchromatic variant 16p+ implications in prenatal diagnosis. Prenat Diagn. 2006;26:535–538. doi: 10.1002/pd.1454. [DOI] [PubMed] [Google Scholar]
- Ronaghi M, Uhlén M, Nyrén P. A sequencing method based on real-time pyrophosphate. Science. 1998;281:363–365. doi: 10.1126/science.281.5375.363. [DOI] [PubMed] [Google Scholar]
- Barber JC, Maloney VK, Huang S, et al. 8p23.1 duplication syndrome; a novel genomic condition with unexpected complexity revealed by array CGH. Eur J Hum Genet. 2008;16:18–27. doi: 10.1038/sj.ejhg.5201932. [DOI] [PubMed] [Google Scholar]
- Eichler EE, Lu F, Shen Y, et al. Duplication of a gene rich cluster between 16p11.1 and Xq28: a novel pericentromeric directed mechanism for paralogous genome evolution. Hum Mol Genet. 1996;5:899–913. doi: 10.1093/hmg/5.7.899. [DOI] [PubMed] [Google Scholar]
- Whitby H, Tsalenko A, Aston E, et al. Benign copy number changes in clinical cytogenetic diagnostics by array CGH. Cytogenet Genome Res. 2008;123:94–101. doi: 10.1159/000184696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourthoumieu S, Esclaire F, Terro F, et al. First prenatally diagnosed case of 16p11.2p12.1 duplication. Prenat Diagn. 2008;28:254–256. doi: 10.1002/pd.1949. [DOI] [PubMed] [Google Scholar]
- Huang N, Lee I, Marcotte EM, et al. Characterising and predicting haploinsufficiency in the human genome. PLoS Genet. 2010;6:e1001154. doi: 10.1371/journal.pgen.1001154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H, Ouyang Y, Somers WG, et al. Polo inhibits progenitor self-renewal and regulates Numb asymmetry by phosphorylating Pon. Nature. 2007;449:96–100. doi: 10.1038/nature06056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Girirajan S, Eichler EE. A recurrent 16p12.1 microdeletion suggests a two-hit model for severe developmental delay. Hum Mol Genet. 2010;19:R176–R187. doi: 10.1038/ng.534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Artuso R, Papa FT, Grillo E, et al. Investigation of modifier genes within copy number variations in Rett syndrome. J Hum Genet. 2011;56:508–515. doi: 10.1038/jhg.2011.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li F, Shen Y, Köhler U, et al. Interstitial microduplication of Xp22.31: causative of intellectual disability or benign copy number variant. Eur J Med Genet. 2010;53:93–99. doi: 10.1016/j.ejmg.2010.01.004. [DOI] [PubMed] [Google Scholar]
- Furrow A, Theisen A, Velsher L, et al. Duplication of the STS region in males is a benign copy-number variant. Am J Med Genet Part A. 2011;155A:1972–1975. doi: 10.1002/ajmg.a.33985. [DOI] [PubMed] [Google Scholar]
- Hastings PJ, Lupski JR, Rosenberg SM, et al. Mechanisms of change in gene copy number. Nat Rev Genet. 2009;10:551–564. doi: 10.1038/nrg2593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guenthoer J, Diede SJ, Tanaka H, et al. Assessment of palindromes as platforms for DNA amplification in breast cancer. Genome Res. 2012;22:232–245. doi: 10.1101/gr.117226.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall V, Maloney VK, White H, et al. The use of pyrosequencing to identify copy number variation of 16p11.2 in euchromatic variant carriers and the normal population J Med Genet 200643(Suppl 13.26. [Google Scholar]
- Fode P, Jespersgaard C, Hardwick RJ, et al. Determination of beta-defensin genomic copy number in different populations: a comparison of three methods. PLoS One. 2011;6:e16768. doi: 10.1371/journal.pone.0016768. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.