

Abstract
Aim
To characterize how nicotinic acetylcholine receptors (nAChRs) influence epidermal barrier function and recovery following prolonged stress or direct nAChR activation or antagonism.
Main Methods
Mice were subjected to psychological stress or treated topically with nAChR agonist or antagonist for 3 days. We assessed barrier permeability and recovery by measuring transepidermal water loss (TEWL) before and after barrier disruption. In parallel, we analyzed the production and localization of several epidermal cornified envelope proteins in mouse skin and in human EpiDerm™ organotypic constructs stimulated with a nAChR agonist (nicotine) and/or a nAChR selective antagonist (α-bungarotoxin).
Key Findings
We determined that psychological stress in mice impairs barrier permeability function and recovery, an effect that is reversed by application of the α7 selective nAChR antagonist, α-bungarotoxin (Bung). In the absence of stress, both topical nicotine or Bung treatment alone impaired barrier permeability. We further observed that stress, topical nicotine, or topical Bung treatment in mice influenced the abundance and/or localization of filaggrin, loricrin, and involucrin. Similar alterations in these three major cornified envelope proteins were observed in human EpiDerm™ cultures.
Significance
Perceived psychological stress and nicotine usage can both initiate or exacerbate several dermatoses by altering the cutaneous permeability barrier. Modulation of nAChRs by topical agonists or antagonists may be used to improve epidermal barrier function in skin diseases associated with defects in epidermal barrier permeability.
Keywords: Nicotinic Receptors, Barrier Permeability, Epidermal Proteins, Acetylcholine
INTRODUCTION
As a barrier between the external and internal environment, the skin functions to prevent the loss of vital fluids, to provide a physical impediment to external stressors, and to defend against pathogen challenge. To maintain this barrier, rapidly proliferating keratinocytes located in the basal layer migrate through the upper layers of the epidermis. During this process, keratinocytes will undergo keratinization and several morphological modifications to generate the outermost protective layer of the epidermis, the stratum corneum. During the granular transition, keratinocytes release the contents of keratohyalin granules containing key structural proteins (i.e. involucrin, loricrin, and profilaggrin) into the cytoplasm (Candi et al., 2005, Kalinin et al., 2001, Steven and Steinert, 1994). Keratinocytes further limit barrier permeability by maintaining cell-cell interactions via desmosomes, adherens junctions, and tight junctions, while packing intercellular spaces with glycoproteins and lipoproteins (Candi, Schmidt, 2005). Desmosomes and adherens junctions provide mechanical intercellular connections by bridging keratin-filament cytoskeletons of adjacent cells (Green and Simpson, 2007), whereas tight junctions fuse plasma membranes and allow selective passage of ions and uncharged small molecules via a paracellular mechanism (O’Neill and Garrod, 2011). During the final stages of terminal differentiation into anucleated corneocytes, keratinocytes located within the granular layer undergo lamellar body secretion to deposit lipids and antimicrobial peptides onto the surface of the skin to provide a physical and chemical shield against water loss, chemical stressors, and pathogen invasion (Aberg et al., 2008, Elias et al., 2000). In parallel, keratin and filaggrin create the primary structural scaffold for the cornified lipid envelope, while involucrin and loricrin provide the filamentous framework for the binding of additional structural proteins and ceramides (Elias, Fartasch, 2000). Ultimately, an effective barrier is dependent upon the tight regulation and organization of these molecules within the stratum corneum. Thus, inadequate barrier function results in excess water loss and penetration of allergens, irritants, and microbes, which likely contribute to or exacerbate pathologic skin diseases.
Acetylcholine (ACh) serves a critical role in both the development of the stratified epidermis and maintenance of the epidermal barrier. Keratinocytes have the capacity to synthesize, process, store, and release ACh to activate nicotinic (nAChR) or muscarinic (mAChR) acetylcholine receptors expressed on the cell surface as a part of the non-neuronal cholinergic system. In the absence of neuronal-derived ACh, keratinocytes employ their inherent cholinergic system to regulate migration, differentiation, proliferation, and apoptosis as a mechanism to help sustain the epidermal barrier (Denda et al., 2000, Grando, 1993). Various combinations of nAChR subunits form ligand-gated ion channels that, upon activation, result in an influx of calcium which acts as a second messenger, while stimulation of mAChRs results in G-protein coupled signaling cascades (Grando, 2012). During keratinocyte differentiation, the diverse expression of nAChR or mAChR subunits and ACh regulatory enzymes facilitates essential functions required for the specific microenvironment of each layer (Kurzen et al., 2004). Furthermore, ACh resides in the epidermis as a chemical gradient, where the highest concentrations of ACh exist in the outermost epidermal compartment to facilitate differentiation (Nguyen et al., 2001). Studies have demonstrated that altered expression or pharmacologic manipulation of nAChRs and mAChRs correlates with impaired differentiation and barrier defects, in part, due to the loss of intercellular connections (acantholysis) in a model of human organotypic epithelial raft cultures (Kurzen et al., 2006). However, few studies have examined the role of these receptors in regulating barrier permeability using in vivo models of nAChR receptor activation or inhibition.
ACh is also a key molecule involved in the stress response that is released from neuronal junctions following activation of the parasympathetic nervous system. Stress simultaneously activates both the sympathetic nervous system to increase ACh and catecholamines (e.g. epinephrine and norepinephrine) and the hypothalamic-pituitary-adrenal axis to increase glucocorticoid release. For decades, stress has been implicated as a risk factor for the development or progression of pathologic conditions, including inflammatory skin diseases, delayed wound healing, and an increased risk of infection. Since a defective epidermal barrier is frequently observed in several dermatoses, initial studies were aimed at investigating the influence of psychological stress on factors important for barrier maintenance, such as epidermal lipid and antimicrobial peptide (AMP) production. Stressed mice exhibited delayed barrier recovery following disruption via tape stripping and also decreased epidermal proliferation and differentiation (Choi et al., 2005, Denda, Tsuchiya, 2000). Furthermore, stress reduced the production and secretion of lamellar bodies (Choi, Brown, 2005), epidermal lipids, and AMPs, which was reversed by pretreatment with a glucocorticoid receptor antagonist (Aberg, Man, 2008). These findings established a direct effect of stress-induced neuroendocrine mediators on the biochemical pathways required for normal epidermal barrier homeostasis.
ACh signaling through α7 nAChR ligand-gated ion channels is known to directly regulate the late stages of epidermal differentiation by influencing cell cycle progression, apoptosis, and terminal differentiation (Arredondo et al., 2002). We recently reported that both psychological stress and nAChR activation via topical nicotine treatment reduced epidermal AMP production and activity in mice and human keratinocytes, which could be reversed by the presence of the α7 nAChR selective antagonist, α-bungarotoxin (Bung) (Radek et al., 2010). Given that topical application of Bung to mouse skin was able to reverse the effects of stress on epidermal AMPs, we concluded that non-neuronal epidermal nAChR activation was responsible for this perturbation. Since the permeability barrier and the chemical AMP barrier are co-dependent (Aberg, Man, 2008), we investigated whether manipulation of nAChR stimulation also resulted in functional and structural barrier defects. We explored whether psychological insomnia stress or topical nicotine treatment on mouse skin altered cornified envelope protein localization and production, and we measured barrier integrity by assessing transepidermal water loss (TEWL). We further investigated the specific role of the α7 nAChR by determining if topical application of Bung could restore barrier defects. Finally, we confirmed the effects of nAChR activation on cornified envelope proteins in human EpiDerm™ organotypic constructs, as a means of translating our findings in both in vitro cell culture and animal models into a more clinically relevant system.
MATERIALS AND METHODS
Animals
8–10 week old male C57BL/6 mice (Jackson Laboratories) were divided into 8 treatment groups. Mice were housed under normal conditions or subjected to 3 days of psychological insomnia stress conditions (i.e. crowding, radio static, constant light) in the presence or absence of topical Bung (MP Biomedicals) as described (Aberg et al., 2007). Alternatively, mice were treated topically with vehicle or 1nM nicotine (Sigma) in the presence or absence of 100nM Bung as described (Radek, Elias, 2010). All animal protocols were approved by Loyola University Chicago, Health Sciences Campus IACUC.
Stimulation of Human Epidermal Constructs
EpiDerm™ (EPI-200) was purchased from MatTek Corporation. Constructs were acclimated in modified phenol-red-free MatTek media supplemented with 11ng/ml hydrocortisone and penicillin/streptomycin for 24 hours at 37 C, 5% CO2. Constructs were stimulated for 24 hours with MatTek media containing 0.01nM nicotine, 100nM Bung, or vehicle control.
Immunohistochemistry (IHC) and Lucifer Yellow Permeability
Standard IHC was performed on mouse skin and MatTek organotypic constructs, using primary antibodies for Filaggrin (Covance), Loricrin (Abcam), or Involucrin (Abcam), and appropriate secondary antibodies conjugated to either Cy3 or AlexaFluor 568. For Lucifer Yellow staining, experiments were adapted from (Mildner et al., 2010). In brief, ~20μl of 1mM Lucifer Yellow solution (Sigma) was added to the epidermal surface of skin biopsy punches excised from mice treated as described. Samples were protected from light and incubated at 37 C for 1 hour. Skin was mounted in optimal cutting temperature medium, sectioned (8μM), fixed in acetone, and nuclei were stained using Prolong Antifade Gold with DAPI. Photos were taken on an Evos Digital Inverted Microscope using a 20X objective and analyzed in a blinded manner.
Transepidermal Water Loss (TEWL)
TEWL measurements were taken on the dorsum of live, anesthetized mice prior to and following barrier disruption via tape stripping (20 strips) after 0, 3, 6, and 24 hours using a Courage and Khazaka TM-300 TEWAmeter. Measurements were taken continuously until a steady value was reached. Higher raw values typically indicate an abnormal and/or disrupted barrier. Percent barrier recovery 3 hours post tape-stripping was calculated as follows:
where 100% recovery indicates return to the baseline (pre-tape-stripping) value.
Statistical Analysis
TEWL was calculated as the mean ± standard error of the mean (SEM). Data were analyzed using GraphPad Prism v5 by One-way ANOVA and Dunnet’s multiple comparison post test, where p<0.05 values were considered statistically significant.
RESULTS
We previously reported that nAChR activation via topical nicotine (1nM) diminished extractable antimicrobial activity against Staphylococcus aureus in mouse skin, and further determined that topical application (100 nM) of the α7-nAChR selective antagonist, α-bungarotoxin (Bung) (100nM= 10-fold > than Kd of Bung) could reverse the susceptibility of mice to cutaneous infection induced by prolonged psychological stress (Radek, Elias, 2010). Furthermore, psychological stress delays barrier recovery in both mouse and human skin (Choi et al., 2006, Garg et al., 2001). Taken together, these findings suggested that the cholinergic branch of the stress response, and specifically the α7 nAChR, negatively regulated AMP production and activity in the epidermis.
TEWL measurements are a useful biomarker for determining stratum corneum integrity and permeability and elevated TEWL has been observed following physical disruption and is associated with several skin dermatoses (Elias, Fartasch, 2000). Our first goal was to determine if Bung could reverse the increase in TEWL observed during stress by blocking α7-nAChR activation. We found that vehicle-treated stressed mice had significantly higher initial TEWL values following three days of psychological stress compared to unstressed vehicle-treated mice. However, this effect was abolished in the presence of Bung, as stressed mice treated with topical Bung were not significantly different from unstressed vehicle-treated mice (Figure 1A). To determine if direct, topical nAChR stimulation or antagonism alters barrier permeability, TEWL was assessed in unstressed mice treated topically with nicotine or Bung. Compared to vehicle, both treatments resulted in a significant increase in TEWL values (Figure 1B), indicating a defect in barrier permeability homeostasis following non-neuronal nAChR activation or antagonism. Thus, our results from Figure 1A and B suggest that the presence of a nAChR antagonist (e.g. Bung) prior to excess agonist induced by stress (e.g. ACh) can successfully block the effects of the agonist by acting as a competitive inhibitor. However, the presence of either agonist or antagonist alone promotes similar alterations in TEWL to suggest a more dynamic role for nAChR antagonists in maintaining barrier permeability To next evaluate whether nAChR activation changes normal barrier recovery, we measured TEWL 3, 6, and 24 hours following barrier disruption via tape-stripping, a superficial wounding that extracts cells and lipids within the stratum corneum. Unstressed vehicle-treated mice exhibited a 64% barrier recovery 3 hours post-tape stripping (Figure 1C), whereas unstressed mice treated with Bung exhibited only a 49% recovery. Vehicle-treated stressed mice merely demonstrated a 41% barrier recovery, which was significantly less than vehicle-treated unstressed mice. Interestingly, stressed mice treated with Bung exhibited a 55% recovery, restoring barrier recovery similar to that of vehicle-treated unstressed mice. Similar recoveries were observed at 6 and 24 hours, although vehicle-treated stressed mice demonstrated a statistical difference from unstressed-vehicle treated mice only at the 24 hour time point. We further observed a trend towards delayed barrier recovery following nAChR activation by nicotine, as mice treated with nicotine alone exhibited a 49% recovery compared to 64% in vehicle-treated mice (Figure 1D). Barrier recovery improved to 61% with the addition of Bung, although differences were not statistically significant (Figure 1D). Similar results were observed at 6 and 24 hours post-barrier disruption (Figure 1D). However, topical application of muscarine (150μM, 10-fold >than the Kd), a selective mAChR agonist, did not alter initial TEWL or barrier recovery (data not shown).
Figure 1. Stress and topical treatment with nicotine or α-bungarotoxin (Bung) promotes epidermal barrier disruption and reduces barrier recovery following barrier disruption.

Transepidermal water loss (TEWL) was measured on the mouse dorsum (n=10–18/group) following three days of psychological stress in the presence or absence of Bung (A), or in unstressed mice treated topically with vehicle, nicotine, or Bung (B). TEWL measurements taken 3, 6 and 24 hours following barrier disruption via tape stripping were used to assess barrier recovery following stress (C) or nicotine treatment (D) +/− Bung. All comparisons represent differences from unstressed, vehicle (A, C) or vehicle alone (B, D) within the same time-point by One-way ANOVA with Dunnett’s multiple comparison post-test where *p < 0.05 was considered significant. #p = 0.06
To confirm that increased TEWL following nAChR activation via nicotine treatment corresponds to increased barrier permeability, we utilized an ex vivo approach using the hydrophilic fluorescent dye, Lucifer Yellow. An intact, effective barrier normally limits Lucifer Yellow penetration beyond the stratum corneum, as observed in vehicle-treated mice (Figure 2A). However, Lucifer Yellow penetration into the basal layer and partial dermis was observed in nicotine-treated (Figure 2B) and Bung-treated mice (Figure 2C), confirming a compromised epidermal barrier following agonist or antagonist treatment. In a separate pilot study, we observed that mice treated with Bung prior to treatment with nicotine did not have significantly different initial TEWL values when compared to vehicle-treatment alone (data not shown). Accordingly, we observed Lucifer Yellow was confined to the stratum corneum in mice treated topically with Bung prior to nicotine treatment (Figure 2D), suggesting a unique role for Bung in the presence of an agonist to block the effects on barrier permeability.
Figure 2. Topical nicotine and α-bungarotoxin (Bung) treatment increases barrier permeability beyond the stratum corneum.
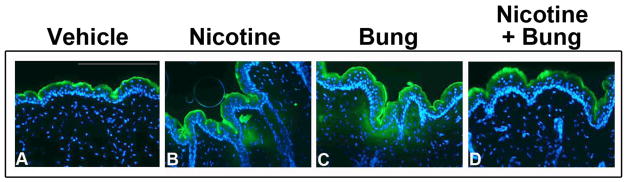
Lucifer Yellow was retained in the stratum corneum in vehicle (A) treated mouse skin. Lucifer Yellow penetrated into the basal layers of the epidermis and dermis in nicotine (B) and Bung (C) treated mouse skin. Lucifer Yellow was retained in the epidermis in combined nicotine + Bung treated mouse skin (D). Scale bar= 200μm
Since nicotine and Bung both independently increased barrier permeability, we next sought to determine if these treatments altered the localization or production of cornified envelope proteins (e.g. filaggrin, loricrin, and involucrin) in mouse skin or human EpiDerm™ constructs by immunohistochemistry. We previously determined that ACh and nicotine elicited similar effects within the same dosing range in keratinocytes (0.01–1nM), with maximal suppression of AMP responses observed at 0.01nM ACh or nicotine compared to controls (Radek, Elias, 2010). In parallel studies, we observed that 100nM Bung, which is ~10-fold greater than its reported Kd, was able to reverse the suppression of AMP responses in keratinocytes. Since AMP production and barrier permeability are co-dependent, we stimulated organotypic rafts with 0.01nM nicotine and 100nM Bung, which would mimic our previous conditions using keratinocyte cultures and mouse models of nAChR activation/blockade. In our current study, treatment with the nAChR agonist (nicotine) and antagonist (Bung) increased filaggrin in both mouse skin and in human constructs (Figure 3A). In a healthy epidermis, involucrin is normally confined to the upper spinous layers and the stratum granulosum, as observed in vehicle-treated mouse skin (Figure 3B). However, nicotine treatment decreased involucrin abundance in the more differentiated layer of the epidermis in mouse skin, whereas Bung increased epidermal involucrin. In human EpiDerm™ constructs, greater involucrin production was observed in the stratum corneum in vehicle-treated constructs, which normally acts as an early component in the assembly of the cornified envelope. In contrast, a more diffuse, premature involucrin production was identified following nicotine or Bung treatment (Figure 3B). Lastly, we noted that the production of loricrin in the granular layer of mouse epidermis decreased with nicotine treatment compared to vehicle (Figure 3C). However, Bung treatment robustly increased loricrin production throughout the entire mouse epidermis (Figure 3C). Although changes in the production of loricrin in human EpiDerm™ constructs was not observed, we did detect a profound change in localization with both nicotine or Bung treatments, where loricrin appeared to be more confined to the less differentiated corneocytes in treated constructs compared to vehicle (Figure 3C).
Figure 3. Nicotine and α-bungarotoxin (Bung) treatment alters the abundance or localization of cornified envelope proteins in mouse skin and human epidermal constructs.

Immunohistochemical analyses were performed on mouse skin sections or human EpiDerm™ constructs using primary antibodies for filaggrin (A), involucrin (B), or loricrin (C), and appear as red staining on all images. DAPI nuclear staining appears blue. All panels are oriented with mouse skin in the upper portion and human EpiDerm constructs in the lower portion.
DISCUSSION
Both anecdotal and clinical evidence for the role of perceived stress in skin disease pathophysiology has existed for decades. Psychological stress has been correlated with deterioration of the cutaneous barrier, suggesting that barrier perturbations may trigger or exacerbate the clinical manifestations of skin disease pathogenesis (Garg, Chren, 2001). Furthermore, our recent investigations uncovered a previously unknown pathway for the negative regulation of epidermal AMPs through activation of nAChRs, as AMPs are co-factors which act in parallel with epidermal lipids to maintain epidermal barrier permeability and homeostasis (Radek, Elias, 2010).
The stratum corneum provides the protective interface between the internal and external environment by strategically incorporating lipids, structural proteins, and AMPs into a virtually impermeable barrier. In the present study, we compared the effects of a traditional nAChR agonist and antagonist on barrier permeability and the regulation of the cornified envelope proteins in both mouse and human epidermis. We established that nAChRs contribute to barrier integrity and recovery, which may be due to alterations in the production or localization of cornified envelope proteins. Several studies have implicated defects in cornified envelope proteins in the development or progression of skin dermatoses. Vohwinkel syndrome and progressive symmetric erythrokeratodermia dermatoses are associated with a genetic mutation in loricrin that perturbs cornified envelope formation (Ishida-Yamamoto et al., 1997, Maestrini et al., 1996), and reduced loricrin production has also been observed in skin lesions from atopic dermatitis patients (Jensen et al., 2004). Filaggrin deficiency or mutation is correlated with ichthyosis vulgaris, psoriasis, and atopic dermatitis (Irvine et al., 2011, Kim et al., 2011). Moreover, defective filaggrin processing has been correlated with Netherton’s syndrome, which results in the overproduction of filaggrin monomers and, consequently, compromises barrier permeability and the adhesive capacity of the stratum corneum (Hewett et al., 2005). Since we observed an increase in filaggrin throughout most of the epidermis in response to nAChR stimulation, we speculate that nAChRs may augment epidermal protease activity, resulting in the accumulation of filaggrin monomers within the epidermis.
Earlier studies demonstrated a critical role for nAChR signaling in the development of organotypic epithelial raft cultures, where AChR blockade via mecamylamine and atropine treatment during epidermal development completely inhibited formation of an intact epithelium (Kurzen, Henrich, 2006). Treatment with select cholinergic agonists or antagonists led to variations in the production and localization of structural proteins, including filaggrin and several desmosomal proteins, such as desmoplakin, desmoglein-1, desmoglein-3, desmocollin-1, and the critical tight junction protein, Zona occludens-1 (ZO-1), during different stages of epidermal development (Kurzen, Henrich, 2006). Since the physical foundation of the stratum corneum is catalyzed via transglutaminases by cross-linking structural proteins and covalently attaching lipids to form the cornified envelope, the alternate or parallel mechanism to explain the defects in barrier permeability and/or localization of cornified envelope proteins identified in our study may be associated with alterations in transglutaminase activity (Matsuki et al., 1998). Our findings further highlight the importance of cholinergic activation on regulating epidermal structural proteins in a fully developed epidermis, and emphasize the sensitivity of epidermal barrier function to nAChR agonists and/or antagonists.
Our unpublished results indicate that psychological stress increases the levels of cutaneous ACh in mouse skin. Since ACh serves critical roles in the development of the stratified epidermis and maintenance of the permeability barrier by regulating keratinocyte migration, differentiation, proliferation, and apoptosis, the tight regulation of any of these processes may be disrupted by an increase in cutaneous ACh (Kurzen et al., 2007). This could occur by disturbing the normal epidermal ACh gradient, altering the distribution of specific nAChR subunits, and/or the activity of ACh regulatory enzymes, such as acetylcholinesterase. It was previously demonstrated that stimulation of keratinocyte monolayers with micromolar concentrations of nicotine increased protein levels of filaggrin, involucrin, keratin-10, and transglutaminase type 1 (Grando et al., 1996). Furthermore, during the granular cell-corneocyte transition, the m1-AChR, α7- and α9- nAChRs are required for apoptotic secretion, a process through which keratohyalin contents, including organelles and large amounts of filaggrin, are released in a characteristic membrane blebbing (Nguyen, Ndoye, 2001). Interestingly, we observed alterations in both barrier permeability and the regulation of cornified envelope proteins with both a traditional nAChR agonist (e.g. nicotine) and antagonist (e.g. Bung).
From a pharmacological perspective, traditional neuronal nAChR antagonists are a subtype of anti-cholinergic molecules that are expected to inhibit the action elicited by an appropriate agonist, and must not promote any intrinsic activity. nAChR antagonists are usually large molecules that can interact with the agonist binding site and extend beyond the binding site perimeter to block any necessary conformational changes required for ion channel opening and downstream agonist-induced responses. Alternatively, antagonists may interact at unique allosteric sites which do not usually activate the receptor (Albuquerque et al., 2009). However, several intermediate forms of agonists exist, which may explain our divergent observations seen with the nAChR antagonist, Bung. SLURP-1 (secreted mammalian Ly-6/urokinase-type plasminogen activator receptor (uPAR)-related protein-1) is an endogenous, allosteric agonist of nAChRs that is produced and secreted by keratinocytes and acts to finely-tune cholinergic pathways in the presence of the natural agonist, ACh (Chimienti et al., 2003). Interestingly, SLURP-1 and Bung have similar structural characteristics, and both preferentially bind to α7-nAChR due to a higher affinity for the α7 nAChR compared to other receptor subtypes (Arredondo et al., 2005). Hence, it is plausible that in some circumstances, Bung may function similarly to SLURP-1 to modulate cholinergic activation rather than antagonizing α7 nAChR signaling. Furthermore, it is the combined contribution of specific nAChRs and mAChRs that orchestrates the development and maintenance of the stratified epidermis (Grando, 2012). Therefore, antagonism of α7 nAChR by Bung may indirectly augment mAChR or α9 nAChR stimulation, and possibly explain the similar effects we observed with topical nicotine and Bung treatments.
CONCLUSION
Collectively, these investigations demonstrate that non-neuronal nAChRs in keratinocytes play an important role in epidermal barrier permeability maintenance and recovery during stress and following direct nAChR stimulation. This effect may be mediated, in part, by modulating the production or localization of cornified envelope proteins, including filaggrin, involucrin, and loricrin. Our observation that the α7-selective nAChR antagonist, Bung, promotes similar defects in barrier permeability in the absence of nicotine agonist, but is also able to block the detrimental effects in the presence of nicotine suggests that Bung may function similarly to the endogenous allosteric agonist, SLURP-1, to modulate cholinergic activation rather than antagonizing α7 nAChR signaling under certain conditions. Furthermore, the delay in barrier recovery in stressed animals was restored by topical Bung treatment, implicating an important role for the α7 nAChR in barrier recovery during stress. Although we observed changes in several cornified envelope structural proteins, keratinocytes may compensate for the loss of one cornified envelope component by employing or altering the production of other available proteins in an effort to help restore normal barrier function. Normally, nAChR agonists can rapidly promote receptor desensitization to impede their function; however, the data presented here suggest that nAChR antagonists provide an alternative target for the treatment of disease by offering another means to modulate nAChR function. The limited appreciation for nAChR modulators in the skin, as well as the obstacles arising from the existence of multiple nAChR subtypes within the epidermis, emphasize the complexity of the non-neuronal cholinergic system that necessitates further scrutiny. Future investigations may lead to the identification of more potent and selective nAChR targets to treat inflammatory skin diseases associated with barrier defects or dysregulated structural protein expression.
Acknowledgments
This research is supported by NIH R01-AR-061497-01A1, NIH P30 1P30-AA-019373-01, NIH T32-AA-013527-09, NIH T32-GM008750-11, and the Dr. Ralph and Marian C. Falk Medical Research Trust. This research was also supported, in part, by resources provided by the Northwestern University Skin Disease Research Center (NU-SDRC) (Chicago, IL) through the NIH NIAMS P30-AR-057216 Award. Any opinions, findings, conclusions, or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the NU-SDRC or the NIH/NIAMS. We thank Paul Hoover and Shuagni Yang at the NU-SDRC for their technical assistance with the organotypic skin raft cultures, and Vanessa Nienhouse at Loyola University Chicago, Health Sciences Campus for critical reading of the manuscript.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Aberg KM, Man MQ, Gallo RL, Ganz T, Crumrine D, Brown BE, et al. Co-regulation and interdependence of the mammalian epidermal permeability and antimicrobial barriers. J Invest Dermatol. 2008;128:917–25. doi: 10.1038/sj.jid.5701099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aberg KM, Radek KA, Choi EH, Kim DK, Demerjian M, Hupe M, et al. Psychological stress downregulates epidermal antimicrobial peptide expression and increases severity of cutaneous infections in mice. The Journal of clinical investigation. 2007;117:3339–49. doi: 10.1172/JCI31726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albuquerque EX, Pereira EF, Alkondon M, Rogers SW. Mammalian nicotinic acetylcholine receptors: from structure to function. Physiological reviews. 2009;89:73–120. doi: 10.1152/physrev.00015.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arredondo J, Chernyavsky AI, Webber RJ, Grando SA. Biological effects of SLURP-1 on human keratinocytes. The Journal of investigative dermatology. 2005;125:1236–41. doi: 10.1111/j.0022-202X.2005.23973.x. [DOI] [PubMed] [Google Scholar]
- Arredondo J, Nguyen VT, Chernyavsky AI, Bercovich D, Orr-Urtreger A, Kummer W, et al. Central role of alpha7 nicotinic receptor in differentiation of the stratified squamous epithelium. J Cell Biol. 2002;159:325–36. doi: 10.1083/jcb.200206096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Candi E, Schmidt R, Melino G. The cornified envelope: a model of cell death in the skin. Nature reviews Molecular cell biology. 2005;6:328–40. doi: 10.1038/nrm1619. [DOI] [PubMed] [Google Scholar]
- Chimienti F, Hogg RC, Plantard L, Lehmann C, Brakch N, Fischer J, et al. Identification of SLURP-1 as an epidermal neuromodulator explains the clinical phenotype of Mal de Meleda. Human molecular genetics. 2003;12:3017–24. doi: 10.1093/hmg/ddg320. [DOI] [PubMed] [Google Scholar]
- Choi EH, Brown BE, Crumrine D, Chang S, Man MQ, Elias PM, et al. Mechanisms by which psychologic stress alters cutaneous permeability barrier homeostasis and stratum corneum integrity. The Journal of investigative dermatology. 2005;124:587–95. doi: 10.1111/j.0022-202X.2005.23589.x. [DOI] [PubMed] [Google Scholar]
- Choi EH, Demerjian M, Crumrine D, Brown BE, Mauro T, Elias PM, et al. Glucocorticoid blockade reverses psychological stress-induced abnormalities in epidermal structure and function. American journal of physiology Regulatory, integrative and comparative physiology. 2006;291:R1657–62. doi: 10.1152/ajpregu.00010.2006. [DOI] [PubMed] [Google Scholar]
- Denda M, Tsuchiya T, Elias PM, Feingold KR. Stress alters cutaneous permeability barrier homeostasis. Am J Physiol Regul Integr Comp Physiol. 2000;278:R367–72. doi: 10.1152/ajpregu.2000.278.2.R367. [DOI] [PubMed] [Google Scholar]
- Elias PM, Fartasch M, Crumrine D, Behne M, Uchida Y, Holleran WM. Origin of the corneocyte lipid envelope (CLE): observations in harlequin ichthyosis and cultured human keratinocytes. J Invest Dermatol. 2000;115:765–9. doi: 10.1046/j.1523-1747.2000.00124-5.x. [DOI] [PubMed] [Google Scholar]
- Garg A, Chren MM, Sands LP, Matsui MS, Marenus KD, Feingold KR, et al. Psychological stress perturbs epidermal permeability barrier homeostasis: implications for the pathogenesis of stress-associated skin disorders. Archives of dermatology. 2001;137:53–9. doi: 10.1001/archderm.137.1.53. [DOI] [PubMed] [Google Scholar]
- Grando SA. Physiology of endocrine skin interrelations. J Am Acad Dermatol. 1993;28:981–92. doi: 10.1016/0190-9622(93)70141-f. [DOI] [PubMed] [Google Scholar]
- Grando SA. Muscarinic receptor agonists and antagonists: effects on keratinocyte functions. Handbook of experimental pharmacology. 2012:429–50. doi: 10.1007/978-3-642-23274-9_18. [DOI] [PubMed] [Google Scholar]
- Grando SA, Horton RM, Mauro TM, Kist DA, Lee TX, Dahl MV. Activation of keratinocyte nicotinic cholinergic receptors stimulates calcium influx and enhances cell differentiation. The Journal of investigative dermatology. 1996;107:412–8. doi: 10.1111/1523-1747.ep12363399. [DOI] [PubMed] [Google Scholar]
- Green KJ, Simpson CL. Desmosomes: new perspectives on a classic. The Journal of investigative dermatology. 2007;127:2499–515. doi: 10.1038/sj.jid.5701015. [DOI] [PubMed] [Google Scholar]
- Hewett DR, Simons AL, Mangan NE, Jolin HE, Green SM, Fallon PG, et al. Lethal, neonatal ichthyosis with increased proteolytic processing of filaggrin in a mouse model of Netherton syndrome. Hum Mol Genet. 2005;14:335–46. doi: 10.1093/hmg/ddi030. [DOI] [PubMed] [Google Scholar]
- Irvine AD, McLean WH, Leung DY. Filaggrin mutations associated with skin and allergic diseases. The New England journal of medicine. 2011;365:1315–27. doi: 10.1056/NEJMra1011040. [DOI] [PubMed] [Google Scholar]
- Ishida-Yamamoto A, McGrath JA, Lam H, Iizuka H, Friedman RA, Christiano AM. The molecular pathology of progressive symmetric erythrokeratoderma: a frameshift mutation in the loricrin gene and perturbations in the cornified cell envelope. Am J Hum Genet. 1997;61:581–9. doi: 10.1086/515518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen JM, Folster-Holst R, Baranowsky A, Schunck M, Winoto-Morbach S, Neumann C, et al. Impaired sphingomyelinase activity and epidermal differentiation in atopic dermatitis. J Invest Dermatol. 2004;122:1423–31. doi: 10.1111/j.0022-202X.2004.22621.x. [DOI] [PubMed] [Google Scholar]
- Kalinin A, Marekov LN, Steinert PM. Assembly of the epidermal cornified cell envelope. Journal of cell science. 2001;114:3069–70. doi: 10.1242/jcs.114.17.3069. [DOI] [PubMed] [Google Scholar]
- Kim BE, Howell MD, Guttman-Yassky E, Gilleaudeau PM, Cardinale IR, Boguniewicz M, et al. TNF-alpha downregulates filaggrin and loricrin through c-Jun N-terminal kinase: role for TNF-alpha antagonists to improve skin barrier. The Journal of investigative dermatology. 2011;131:1272–9. doi: 10.1038/jid.2011.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurzen H, Berger H, Jager C, Hartschuh W, Naher H, Gratchev A, et al. Phenotypical and molecular profiling of the extraneuronal cholinergic system of the skin. J Invest Dermatol. 2004;123:937–49. doi: 10.1111/j.0022-202X.2004.23425.x. [DOI] [PubMed] [Google Scholar]
- Kurzen H, Henrich C, Booken D, Poenitz N, Gratchev A, Klemke CD, et al. Functional characterization of the epidermal cholinergic system in vitro. The Journal of investigative dermatology. 2006;126:2458–72. doi: 10.1038/sj.jid.5700443. [DOI] [PubMed] [Google Scholar]
- Kurzen H, Wessler I, Kirkpatrick CJ, Kawashima K, Grando SA. The non-neuronal cholinergic system of human skin. Hormone and metabolic research = Hormon- und Stoffwechselforschung = Hormones et metabolisme. 2007;39:125–35. doi: 10.1055/s-2007-961816. [DOI] [PubMed] [Google Scholar]
- Maestrini E, Monaco AP, McGrath JA, Ishida-Yamamoto A, Camisa C, Hovnanian A, et al. A molecular defect in loricrin, the major component of the cornified cell envelope, underlies Vohwinkel’s syndrome. Nat Genet. 1996;13:70–7. doi: 10.1038/ng0596-70. [DOI] [PubMed] [Google Scholar]
- Matsuki M, Yamashita F, Ishida-Yamamoto A, Yamada K, Kinoshita C, Fushiki S, et al. Defective stratum corneum and early neonatal death in mice lacking the gene for transglutaminase 1 (keratinocyte transglutaminase) Proc Natl Acad Sci U S A. 1998;95:1044–9. doi: 10.1073/pnas.95.3.1044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mildner M, Jin J, Eckhart L, Kezic S, Gruber F, Barresi C, et al. Knockdown of filaggrin impairs diffusion barrier function and increases UV sensitivity in a human skin model. J Invest Dermatol. 2010;130:2286–94. doi: 10.1038/jid.2010.115. [DOI] [PubMed] [Google Scholar]
- Nguyen VT, Ndoye A, Hall LL, Zia S, Arredondo J, Chernyavsky AI, et al. Programmed cell death of keratinocytes culminates in apoptotic secretion of a humectant upon secretagogue action of acetylcholine. Journal of cell science. 2001;114:1189–204. doi: 10.1242/jcs.114.6.1189. [DOI] [PubMed] [Google Scholar]
- O’Neill CA, Garrod D. Tight junction proteins and the epidermis. Experimental dermatology. 2011;20:88–91. doi: 10.1111/j.1600-0625.2010.01206.x. [DOI] [PubMed] [Google Scholar]
- Radek KA, Elias PM, Taupenot L, Mahata SK, O’Connor DT, Gallo RL. Neuroendocrine nicotinic receptor activation increases susceptibility to bacterial infections by suppressing antimicrobial peptide production. Cell host & microbe. 2010;7:277–89. doi: 10.1016/j.chom.2010.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steven AC, Steinert PM. Protein composition of cornified cell envelopes of epidermal keratinocytes. Journal of cell science. 1994;107 (Pt 2):693–700. [PubMed] [Google Scholar]