

Abstract
The incidence of type 2 diabetes mellitus (T2DM) is increasing worldwide and diverse environmental and genetic risk factors are well recognized. Single nucleotide polymorphisms (SNPs) in the calpain-10 gene (CAPN-10), which encodes a protein involved in the secretion and action of insulin, and chronic exposure to inorganic arsenic (iAs) through drinking water have been independently associated with an increase in the risk for T2DM. In the present work we evaluated if CAPN-10 SNPs and iAs exposure jointly contribute to the outcome of T2DM. Insulin secretion (beta-cell function) and insulin sensitivity were evaluated indirectly through validated indexes (HOMA2) in subjects with and without T2DM who have been exposed to a gradient of iAs in their drinking water in northern Mexico. The results were analyzed taking into account the presence of the risk factor SNPs SNP-43 and -44 in CAPN-10. Subjects with T2DM had significantly lower beta-cell function and insulin sensitivity. An inverse association was found between beta-cell function and iAs exposure, the association being more pronounced in subjects with T2DM. Subjects without T2DM who were carriers of the at-risk genotype SNP-43 or -44, also had significantly lower beta-cell function. The association of SNP-43 with beta-cell function was dependent on iAs exposure, age, gender and BMI, whereas the association with SNP-44 was independent of all of these factors. Chronic exposure to iAs seems to be a risk factor for T2DM in humans through the reduction of beta-cell function, with an enhanced effect seen in the presence of the at-risk genotype of SNP-43 in CAPN-10. Carriers of CAPN-10 SNP-44 have also shown reduced beta-cell function.
Introduction
Established risk factors for type 2 diabetes mellitus (T2DM), such as genetics, diet and lifestyle, do not fully explain the increase in the number of cases of this disease and environmental diabetogenic pollutants are increasingly to blame [1]. Among these environmental pollutants, inorganic arsenic (iAs) exposure is a major public health concern. An increase in the incidence and prevalence of T2DM has been observed consistently among residents exposed chronically to high iAs concentrations mainly through naturally contaminated drinking water (>100 µg/l) in countries such as Bangladesh, Taiwan and Mexico [2], [3], [4], [5], [6], [7], as well as in communities exposed to low iAs concentrations (<10 µg/l) in the United States [8]. Furthermore, in pregnant women iAs exposure has been associated with increased risk of impaired glucose tolerance at 24–28 weeks gestation and may therefore be associated with gestational diabetes [9].
Experimental evidence using a wide variety of rodent study models supports the epidemiologic data. In pancreatic beta-cells, low iAs levels (arsenite 0.5–5 µM) decrease insulin transcription and down-regulate its secretion [10] by decreasing the calcium-dependent calpain-10 proteolysis of SNAP-25 [11], a process necessary for glucose-stimulated insulin secretion [12]. Moreover, oxidative stress induced by low levels of iAs has also been shown to disturb beta-cell function, including through the induction of apoptosis [13], [14]. In vitro studies in adipocytes also show that iAs and its metabolites decrease insulin sensitivity [15], [16].
Rodents exposed chronically to high doses of iAs develop hyperglycemia or impaired glucose tolerance, hyperinsulinemia and/or low insulin sensitivity, as well as pancreatic stress and oxidative damage [17], [18], [19]. Additionally, a high-fat diet has been shown to act synergistically with iAs to induce glucose intolerance in mice [20]. An expert committee recently concluded that iAs exposure is associated with an increased risk for T2DM in humans; however, the evidence is still limited and there is need for additional research [21].
In the Mexican population, as well as in other populations, an increased risk of T2DM has been linked to polymorphisms of the calpain-10 gene (CAPN-10) [22], [23], [24]. The calcium-dependent, non-lysosomal cysteine protease calpain-10 is involved in the reorganization of the actin cytoskeleton required for both insulin exocytosis in pancreatic beta-cells [25] and insulin-stimulated GLUT4 translocation to the plasma membrane in adipocytes [26], among other functions. Interestingly, we have shown that iAs can reduce insulin secretion in beta-cells through a decrease in the proteolysis of SNAP-25 by calpain-10 [11]. Calpain-10 as part of the fusion machinery of beta-cells works as a calcium sensor regulating insulin exocytosis [12], [27], a mechanism that can be disturbed by iAs [11].
To understand the mechanism of iAs in T2DM pathogenesis more thoroughly, we performed a pilot study to evaluate insulin secretion and insulin sensitivity indirectly through validated indexes (HOMA2: a computational structural model of the feedback system of glucose and insulin in a steady state) in subjects with and without T2DM exposed to a gradient of iAs in their drinking water in northern Mexico. Because genetics and environment, as well as their interaction, play a crucial roles in T2DM onset [28], the data were analyzed taking into account risk polymorphisms in CAPN-10 conferring risk for T2DM in Mexican-American and Mestizo-Mexican populations: SNP-43 (rs3792267), Indel-19 (rs3842570), SNP-63 (rs5030952) and SNP-44 (rs2975760) [22], [23].
Materials and Methods
Ethics Statement
All individuals were informed about the procedure, signed an informed consent form and answered a questionnaire. The ethics committee from the Instituto de Investigaciones Biomédicas, UNAM and the IRB from the University of Arizona approved the research protocol, and the investigation was conducted according to the Declaration of Helsinki principles.
Study Subjects
Non-diabetic subjects and unrelated individuals with previously diagnosed T2DM were selected from the database of a study previously performed in the region [7]. Criteria for T2DM diagnosis were in accordance with those of the American Diabetes Association [29]. Subjects selected for the current study were recruited from the counties of Granada, Hidalgo, Porvenir, Purísima and San Salvador in the municipalities of Matamoros and Francisco I. Madero, Coahuila, México. The new selection criteria included age between 35 and 65 years with 30 or more years of residence in the study area, and subjects were matched by age, gender, community residence and previously reported total iAs concentration in the urine [7]. Subjects undergoing insulin treatment or diagnosed with any other illness (for example cancer and/or hepatitis) were excluded. The study was conducted with 72 subjects (40 with T2DM and 32 without T2DM). Ten of the individuals with T2DM were not receiving any hypoglycemic drug, one was receiving tolbutamide, two metformin, fifteen glibenclamide, and the remaining twelve subjects were receiving a combined hypoglycemic treatment of metformin and glibenclamide.
Biochemical and anthropometric analysis
Age, sex, height, weight, waist circumference, and blood pressure were recorded for each donor. Blood samples were taken following an 8-hr fast. Glucose, total cholesterol, LDL-cholesterol, HDL-cholesterol, and triglyceride levels were measured in serum with enzymatic methods using commercially available colorimetric kits (Randox Laboratories Ltd., County Antrim, UK). Serum insulin concentration was determined through ELISA (Monobind, Inc., Lake Forest, CA, USA). Body mass index (BMI) was calculated using the formula weight/height2 (kg/m2).
Analysis of iAs exposure and metabolism
Urine and drinking water samples were supplied by each donor, collected in polypropylene bottles and stored at −80°C until they were analyzed as previously reported [30], [31]. Briefly, arsenic species in urine [AsV, AsIII, monomethylarsonic acid (MMAV), dimethylarsinic acid (DMAV) and arsenobetaine] were separated by HPLC. Arsenic concentrations in water samples and urine were analyzed by inductively coupled plasma mass spectrometry (ICPMS) utilizing for quality controls the Standard Reference Water, SMR 1640 (NIST, Gaithersburg, MD, USA) and the freeze-dried Urine Reference Material for trace elements (Clinchek-control; RECIPE Chemicals+INSTRUMENTS GmbH, Munich, Germany) for urine [30], [31].
Water consumption habits were ascertained through a standardized questionnaire and iAs consumption per day was determined by multiplying daily water consumption by the concentration of iAs in drinking water for each donor.
Arsenic metabolism efficiency was calculated using the following formulas [32]: first methylation = MMAV/(AsV+AsIII), second methylation = DMAV/MMAV and total methylation = (MMAV+DMAV)/(AsV+AsIII).
Beta-cell function and insulin sensitivity (%)
Beta-cell function and insulin sensitivity values (%) were determined from fasting serum glucose and insulin concentration values using the software HOMA2 (homeostatic model assessment) Calculator V2.2 developed at the University of Oxford [33]. The HOMA2 model is a computational structural model of the feedback system of glucose and insulin in the steady state (overnight fasting). The model consists of a number of nonlinear empirical equations that describe the functions of the organs and tissues involved in glucose regulation in such a way that beta-cell function (insulin secretion) and insulin sensitivity (glucose uptake) percentages can be calculated. The accuracy and precision of HOMA2 correlates highly with the “gold standard” technique of the hyperinsulinemic-euglycemic clamp and with the glucose tolerance test curve, even in subjects taking insulin secretagogues [34], [35], [36]. In six samples with out-of-range insulin values utilized by the HOMA Calculator V2.2 (2.9 to 57.6 mmol/l), it was not possible to calculate beta-cell function or insulin sensitivity percentage.
CAPN-10 genotyping
Genomic DNA was extracted from 1 ml of whole blood using the QIAamp DNA MINI kit (Qiagen, Valencia, CA, USA). The single nucleotide polymorphisms (SNPs) 43 (g.4852, G>A; rs3792267) and 44 (g.4841, T>C; rs2975760) were determined by site-specific Taqman assay by Design (Applied Biosystems, Foster City, CA, USA) using the following primers and probes: For SNP-43 the primers 5′-GCGCTCACGCTTGCT-3′ and 5′-CCTCACCAAGTCAAGGCTTAGC -3′ and the probes 5′-CCTTCAAAcGCCTTAC -3′ labeled with the reporter dye FAM and 5′-CACCTTCAAAtGCCTTAC-3′ labeled with the reporter dye VIC, were used. SNP-44 was amplified with the primers 5′-GCAGGGCGCTCACG-3′ and 5′-CCTCACCAAGTCAAGGCTTAGC-3′ and with the probes 5′-CCTTACTTCaCAGCAAG-3′ labeled with FAM and 5′-CCTTACTTCgCAGCAAG-3′ labeled with VIC. The cycling conditions were 95°C for 15 min followed by 40 cycles of 92°C for 15 sec and 1 min at 60°C. Indel-19 (g.7920 in/del32pb; rs3842570) was analyzed by PCR with the primers 5′-GTTTGGTTCTCTTCAGCGTGGAG-3′ and 5′-CATGAACCCTGGCAGGGTCTAAG-3′ followed by agarose gel electrophoresis to determine the size of the generated products (87 and/or 155 bp) as described [37]. Finally, SNP-63 (g.16378, C>T; rs5030952) was analyzed by direct sequencing with the primers 5′-TCGGGACACTGCTGTTAGGT-3′ and 5′CTGGCTGGAGTTTGGAGAAG-3′. The cycling conditions were 94°C for 2 min, 40 cycles of 94°C for 30 sec, 59°C for 30 sec and 72°C for 80 sec, followed by a final extension of 72°C for 5 min. The direct PCR sequencing reactions were performed using Big Dye V3.0 (Applied Biosystems, Foster City, CA, USA) as previously described [30].
Haplotype for SNPs -43, -19 and -63 was reconstructed using the PHASE program, version 2.1.1 [38], [39] and named according to the nomenclature proposed by Horikawa [22].
Statistical Analyses
The normality of all variables was verified by skewness and kurtosis probes (STATA 9.2), followed by the Kolmogorov-Smirnov test (GraphPad Prism 5.0). All variables that did not fall within the normal distribution were Log-transformed for analysis and are indicated in each case. The statistical significance between both study groups was determined for each variable using an unpaired t-test. Variability by sex was determined using the χ2 test (GraphPad Prism 5.0). Deviation from Hardy-Weinberg equilibrium with Pearson's goodness-of-fit-χ2 test was evaluated using Finetti Generator 3.0.5 [40].
Linear regressions of Log-transformed data were performed to associate arsenic concentration excreted in urine with beta-cell function and with insulin sensitivity values (%) (GraphPad Prism 5.0).
For each group of subjects, regression analyses of Log-transformed data were carried out to determine the association between beta-cell function and SNPs in CAPN-10 as dependent and independent variables, respectively. Associations were adjusted by iAs exposure and known risk factors for T2DM such as age, sex and BMI in a multivariate analysis. Genotypes were coded by dummy variables, three for SNP-43 corresponding to G/G, G/A and A/A alleles and two for SNP-44 analogous to T/T and T/C alleles (STATA 9.2).
In all cases, p values<0.05 were considered statistically significant.
Results
Study subjects and iAs exposure
The only clinical and biochemical characteristic significantly different between the two subject groups (type 2 diabetic and non-diabetic) was higher serum glucose in the diabetic subjects (Table 1). Both groups of subjects had been exposed to similar iAs concentrations through drinking water. The arsenic exposure gradient in the drinking water of subjects ranged from 4.0 to 145.3 µg/l (or ppb) for the non-diabetic group and 2.8 to 131.5 µg/l in the diabetic group.
Table 1. Clinical and biochemical characteristics and exposure to iAs in non-diabetic and diabetic subjects.
| Variables | non-diabetic subjects (n = 32) | Type 2 diabetic subjects (n = 40) | ||||
| Mean ± SD | Median (25th & 75th) | Mean ± SD | Median (25th & 75th) | p | ||
| Sex, Male | (N, %) | 11, 34.4 | 14, 35 | 0.9559 | ||
| Age | (years) | 52.56±2.56 | 53.0 (44.25, 61.5) | 51.85±8.77 | 50.0 (45.0, 59.75) | 0.7406 |
| Weight | (kg) | 76.87±13.47 | 75.80 (68.90, 88.25) | 75.33±12.65 | 74.80 (66.0, 85.5) | 0.6202 |
| Height £ | (m) | 1.59±0.08 | 1.58 (1.54, 1.65) | 1.59±0.10 | 1.55 (1.52, 1.67) | 0.8832 |
| BMI | (kg/m2) | 30.58±5.89 | 29.65 (26.05, 34.18) | 29.91±4.26 | 29.0 (27.3, 32.5) | 0.5810 |
| Waist circumference | (cm) | 102.6±12.26 | 105.0 (93.5, 112.0) | 103.6±10.43 | 103.0 (96.75, 112.0) | 0.7102 |
| Systolic pressure £ | (mmHg) | 128.5±16.67 | 130.0 (113.8, 140.0) | 128.3±16.33) | 130.0 (120.0, 140.0) | 0.9662 |
| Diastolic pressure £ | (mm Hg) | 81±10.46 | 80 (70.0, 90.0) | 79.87±8.89 | 80.0 (70.0, 81.25) | 0.6913 |
| Glucose £ | (mmol/l) | 5.25±0.66 | 5.22 (4.77, 5.79) | 10.66±4.11 | 10.57 (6.93, 13.95) | <0.0001** |
| Total cholesterol £ | (mmol/l) | 5.18±1.47 | 5.0 (3.99, 6.0) | 5.14±1.05 | 5.14 (4.33, 5.63) | 0.8933 |
| LDL-cholesterol £ | (mmol/l) | 1.66±1.12 | 1.52 (0.85, 2.22) | 1.60±0.69 | 1.61 (1.22, 1.80) | 0.6416 |
| HDL-cholesterol £ | (mmol/l) | 1.45±0.64 | 1.30 (1.04, 1.59) | 1.25±0.39 | 1.20 (0.96, 1.51) | 0.1565 |
| Triglycerides £ | (mmol/l) | 23.84±26.83 | 15.65 (10.79, 24.55) | 22.57±16.49 | 18.87 (13.54, 22.13) | 0.5604 |
| Insulin £ | (mU/l) | 14.87±10.81 | 11.80 (6.55, 29.95) | 15.31±8.0 | 14.45 (8.9, 19.08) | 0.3900 |
| Time of diagnosis of T2DM | (years) | 6.77±4.44 | ||||
| Arsenic in drinking water £ | (µg/l) | 78.38±29.35 | 72.21 (60.85, 95.11) | 66.68 36.12 | 71.89 (48.78, 93.39) | 0.0841 |
| Arsenic consumption £ | (µg/day) | 108.60±65.24 | 90.57 (70.16, 148.8) | 125.90 99.04 | 132.70 (49.55, 160.1) | 0.5470 |
| Total arsenic in urine | (µg/l) | 133.40±67.0 | 135.20 (81.04, 168.5) | 100.90 65.21 | 86.92 (52.99, 133.1) | 0.0443* |
Data, except for sex that is expressed by the number and % of male, are represented by mean ± SD and median with 25th and 75th percentiles. Symbol denotes Log-transformation for analysis (£). BMI: body mass index. Symbols denotes statistical significance, p<0.05 (*) and p<0.0001 (**).
Despite the similarity in the concentration of iAs ingested between the two groups, total arsenic excreted in urine, a biomarker of arsenic exposure, was significantly higher in the non-diabetic subjects than in diabetic individuals (Table 1). Analysis of iAs metabolism through species in urine revealed that non-diabetic subjects have a more efficient metabolism of the metalloid, particularly in the first methylation (MMAV/iAs) (Table S1). This finding may explain why iAs excreted in urine was significantly higher in non-diabetic subjects than in diabetic individuals.
Genetic characteristics of the subjects
The allele frequency distribution of at-risk polymorphisms for T2DM in CAPN-10 (SNPs -44, -43, -63 and Indel-19) in non-diabetic and diabetic subjects is shown in Table S2. All of the SNPs were in Hardy-Weinberg equilibrium and none of the donors were homozygous for the C allele of SNP-44. Based on the reconstitution of the haplotype, only two samples from the diabetic group had the at-risk haplotype for T2DM (112/121) for SNPs -43, -19 and -63 (Table S3) previously described for the Mexican-American population [22]. Due to the low frequency of the at-risk haplotype 112/121 found in the study sample, forward genotype stratification was analyzed only with SNPs -43 and -44 because these are the SNPs most associated with T2DM phenotype [23], [41], [42], [43], [44]. The allele frequency for SNP-43 and -44 was in accordance to that previously reported for Mexicans [23].
Association of beta-cell function with iAs exposure
The percentage of functional beta-cells was statistically lower in subjects with T2DM, as expected (Figure 1A). The relationship between the concentration of arsenic in urine and the function of beta-cells in the non-diabetic individuals shows an inversely proportional trend with a slope value of −0.24 but was not statistically significant (r2 = 0.08, p = 0.1441) (Figure 1B). In subjects with T2DM, an inverse association between the concentration of arsenic in urine and the level of beta-cell function was clearly demonstrated, with a slope value of −0.65 (r2 = 0.1067, p = 0.0485) (Figure 1C).
Figure 1. Beta-cell function (%) and association with the concentration of total arsenic excreted in urine.
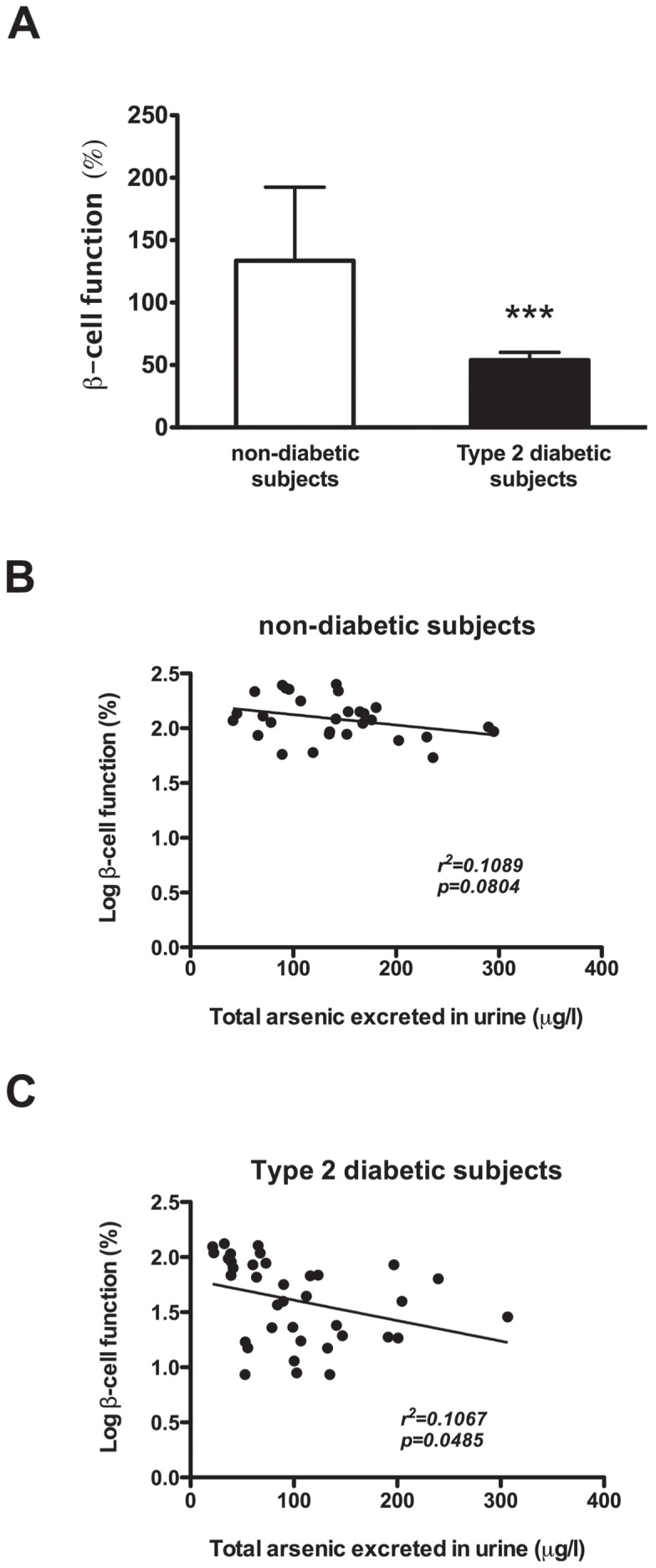
A) Beta-cell function (%) in subjects with and without T2DM; values represent mean ± SD. Symbol (***) denotes statistical significance p<0.0001 (data Log-transformed for analysis). B) Linear regression of the concentration of total arsenic excreted in urine and the % of Log-beta-cell function in non-diabetic donors, and C) Linear regression of the concentration of total arsenic excreted in urine and the % of Log-beta-cell function in diabetic donors.
Stratification of beta-cell function by CAPN-10 genotype with and without consideration of iAs exposure
Because calpains, including calpain-10, are involved in the secretion of insulin and can be altered by arsenic treatment in vitro and because certain SNPs in this gene have been associated with T2DM, the values of the function of beta-cells were stratified by allele for SNP-43 and SNP-44.
When beta-cell function was analyzed based on the SNP-43 genotype, the subjects without T2DM with the at-risk genotype G/G had a percent of beta-cell function statistically lower than those with the genotypes G/A or A/A (Figure 2A). Similarly, when data were analyzed based on the SNP-44 genotype, the subjects without T2DM with the at-risk T/C genotype had statistically lower beta-cell function than those with the T/T genotype (individuals with the C/C genotype were not found in this study) (Figure 2B). In the group of individuals with T2DM, differences were not found between any of those with these two SNPs, as expected (Figure 2C and 2D).
Figure 2.

Beta-cell function and insulin sensitivity (%) stratified by CAPN-10 genotype (SNP-43 and -44) in A) non-diabetic subjects, SNP-43, B) non-diabetic subjects, SNP-44, C) Type 2 diabetic subjects, SNP-43 and D) Type 2 diabetic subjects, SNP-44. Values represent mean ± SD. Symbol (*) denotes statistical significance p<0.05 between alleles (G/G vs. G/A+A/A for SNP-43 and T/T vs. T/C for SNP-44). Beta-cell function values were Log-transformed for statistical analyses.
The association of CAPN-10 genotypes with beta-cell function, taking into account the exposure to iAs and established risk factors for T2DM such as age, sex, and BMI is shown in Table 2. Non-diabetic individuals with the not at-risk genotype A/A for SNP-43 had higher beta-cell function than those with the genotype G/G (p = 0.037); nevertheless, when the association was adjusted by arsenic excreted in urine or other known risk factors for T2DM, the association lost its significance.
Table 2. Univariate and multivariate association between alleles for SNPs-43 and -44 in CAPN-10 and Log-beta cell function (%) in non-diabetic and diabetic subjects.
| Adjusted by | |||||||
| arsenic in urine | arsenic in urine, age, sex and BMI | ||||||
| ß coefficient | p | ß coefficient | p | ß coefficient | p | ||
| Non-diabetic subjects | SNP-43 | ||||||
| G/A vs A/A | −0.34 | 0.226 | −0.28 | 0.302 | −0.09 | 0.748 | |
| G/G vs A/A | −0.57 | 0.037* | −0.49 | 0.073 | −0.32 | 0.278 | |
| P for trend | 0.024* | 0.050 | 0.128 | ||||
| SNP-44 | |||||||
| C/T vs T/T | −0.43 | 0.022* | −0.44 | 0.014* | −0.38 | 0.028* | |
| Type 2 diabetic subjects | SNP-43 | ||||||
| G/A vs A/A | −0.02 | 0.964 | −0.05 | 0.912 | 0.14 | 0.763 | |
| G/G vs A/A | 0.05 | 0.914 | −0.01 | 0.989 | 0.12 | 0.814 | |
| P for trend | 0.871 | 0.978 | 0.874 | ||||
| SNP-44 | |||||||
| C/T vs T/T | 0.47 | 0.211 | 0.56 | 0.156 | 0.57 | 0.160 | |
ß coefficients refer to the estimated change in Log-transformed beta-cell function (%) values between each allele. Symbol denotes statistical significance p<0.05 (*).
The association of beta-cell function with SNP-44 in non-diabetic individuals also showed that subjects with the not at-risk genotype T/T for SNP-44 had higher beta-cell function in comparison to individuals with the T/C genotype (p = 0.024) independent of arsenic exposure (p = 0.014) or age, gender and BMI (p = 0.028). As expected, in subjects with T2DM, neither of the two SNPs showed a statistically significant association with beta-cell function (Table 2).
Association of insulin sensitivity with arsenic exposure and CAPN-10 genotype
The level of insulin sensitivity (reciprocal to insulin resistance) was statistically higher in subjects without T2DM in comparison to those with T2DM (Figure 3A). However, unlike beta-cell function, insulin sensitivity, and therefore insulin resistance, was not associated with the excretion of total arsenic in urine in either of the two analyzed groups (Figure 3B and 3C). Furthermore, insulin sensitivity was also stratified by alleles for SNP-43 and SNP-44 because calpain-10 also participates in insulin-dependent glucose uptake [26], [45], [46]. Insulin sensitivity measured through HOMA2 was not significantly influenced by these SNPs (Figure 2).
Figure 3. Insulin sensitivity (%) and its association with the concentration of total arsenic excreted in urine.

A) Insulin sensitivity (%) of subjects with and without T2DM; values represent mean ± SD. Symbol (*) denotes statistical significance p<0.05 (data Log-transformed for analysis). B) Linear regression of Log-insulin sensitivity and arsenic in urine in non-diabetic subjects C) Linear regression of Log-insulin sensitivity and arsenic in urine in type 2 diabetic subjects.
Discussion
Epidemiological studies in humans [2], [3], [4], [5], [6], [7], [8], [9] as well as in vitro and in vivo studies in rodents [10], [11], [13], [14], [16], [17], [18], [19], [20], have been conducted to determine if iAs exposure is a risk factor for T2DM. Analyses of human samples from subjects exposed to iAs are lacking, especially studies of the effects of the metalloid in tissues such as pancreas and liver. Therefore, pilot studies such as this one are very valuable, even though the number of subjects involved was small. Although evidence shows that high iAs exposure and T2DM are significantly associated, additional research is needed to determine whether the relationship is causal as proposed by The National Institute of Environmental Health Sciences (NIEHS), Division of the National Toxicology Program (NTP) this past year [21]. Moreover, evidence showing that iAs exposure is associated with an increased risk for T2DM in humans is still limited [21], and until now, studies that try to explain the mechanism of how arsenic acts as a risk factor for T2DM have been carried out in rodents or in in vitro studies but never in humans; therefore, this pilot study is relevant. The data obtained from this work support the previous findings from studies carried out in non-human subjects, especially those focusing on beta-cell dysfunction.
In this pilot study, we measured the interaction of iAs on glucose metabolism in humans using non-invasive methods. We also analyzed the association between this environmental metalloid and genetic factors for the disease. Our results show that iAs indeed does affect the pathogenesis of T2DM, particularly in disturbing beta-cell function, and this in turn is also associated with SNPs found in CAPN-10, a susceptibility gene for T2DM in certain populations, including Mexicans [22], [23], [24].
It was expected that daily arsenic consumption and levels of arsenic excreted in the urine would be correlated. Nevertheless, non-diabetic subjects consumed less iAs but had greater concentrations of arsenic in the urine than subjects with T2DM. This phenomenon could be due to differences in arsenic metabolism or, to a less extent, to an inaccuracy in self-reporting of water consumption habits.
The relationship between the concentration of arsenic in urine and the function of beta-cells in diabetic and non-diabetic subjects show the same trend, but the association was statistically significant only in the diabetic subjects. These data suggest that iAs exposure is a risk factor for T2DM rather than a direct inducer of the disease. Failure of beta-cell function is necessary for progression from insulin resistance to T2DM; nevertheless, it is not the only event that is required because many other risk factors are also needed for the development of the diabetic state [47]. In this work, we noticed that, in subjects without T2DM, iAs exposure was associated with a decrease in the function of beta-cells, but this was not sufficient for the development of T2DM. Other risk factors would need to exert their actions, indicating a multifactorial metabolic disruption in the organism. Another possibility may be that the beta-cells of diabetic subjects are extremely sensitive to iAs exposure; however, in this study it was not possible to determine this as a cause (i.e., beta-cells are impaired directly by iAs) or a consequence (i.e., the function of beta-cells from diabetic subjects is deficient and thus cells are more susceptible to the effects of iAs). However, in vitro studies have shown that iAs impairs insulin secretion, leading to beta-cell insensitive to glucose [10], [11]. It could also be possible that by increasing the sample size, a clearer association could be determined between iAs exposure and beta-cell function in non-diabetic subjects.
T2DM is a complex disease, and an interaction between environmental factors and genetic components occurs in the pathogenesis of the disease. In this work we evaluated the influence of SNPs in CAPN-10 on beta-cell dysfunction. Other genes have been associated with T2DM such as TCF7L2, SLC30A8, HHEX, CDKAL1, IGF2BP2, CDKN2A/B, PPARG, KCNJ11, KCNQ1 and MTNR1B, as determined through genome-wide association studies (GWAS) [48], [49]. However, this study analyzed SNPs in CAPN-10 because these have been associated with T2DM in the Mexican population [23] and calpain-10 participates in both glucose-stimulated insulin secretion in beta-cells [25] and in insulin-stimulated GLUT4 translocation to the plasma membrane in adipocytes [26]. Moreover, we previously showed that iAs can reduce insulin secretion in beta-cells through a decrease in the proteolysis of SNAP-25 by calpain-10 [11].
SNP-43 in CAPN-10 is involved in many of the phenotypes associated with T2DM, such as insulin resistance, lipogenesis, insulin secretion and microvascular function [41], [42], [43], [44], while SNP-44 has been associated with increase risk for T2DM in Mexican population [23] and with other populations as evaluated through meta-analysis [24]. SNP-44 has also been associated with alterations in the pattern of the oral glucose tolerance curve [50] and is in linkage disequilibrium with the missense variant T504A (SNP-110), which unlike the other SNPs, is located in the open reading frame of CAPN-10 [22], [23].
Beta-cell function was lower in subjects without T2DM that carried the at-risk genotypes G/G at SNP-43 and T/C at SNP-44. In subjects with T2DM, the influence of the genotype on beta-cell function was not evident because the level of function of these pancreatic cells is already altered due to the disease per se and to all the factors that contributed to the onset of it; therefore, the influence of the genotype has been lost. Once the disease has been established, which not only involves the dysfunction of beta-cells but also other events [47], it is likely that at-risk variants in CAPN-10 are not more associated with the level of beta-cell function, even when iAs exposure occurs. Instead, other mechanisms, such epigenetic modifications, may be taking place, as has been revealed in a genome wide site-specific DNA methylation study conducted in peripheral blood lymphocyte DNA of 16 individuals, half with established elevated levels of iAs exposure and showing signs of arsenicosis (skin lesions). The investigators in this study found that epigenetic changes in genes were associated with arsenic-induced diseases, including T2DM [51].
From the analysis of beta-cell function stratified by CAPN-10 genotypes through multivariate analysis, it can be postulated that the at-risk heterozygote allele T/C in SNP-44 is associated with decreased beta-cell function independently of arsenic exposure or other risk variables for T2DM, such as age, sex and BMI. On the contrary, the risk for the development of T2DM with the G/G genotype at SNP-43 depends upon intrinsic and environmental factors, including iAs exposure.
As mentioned above, for T2DM to occur, other components, such as insulin resistance, are also needed. In contrast to in vitro findings in the literature in which arsenic induces insulin resistance in adipocytes [15], [16], in this pilot study insulin sensitivity was not associated with iAs exposure or with at-risk alleles in CAPN-10. This lack of correlation may be due to the relationship between glucose and insulin in the basal state (fasting) in the HOMA analysis, which reflects the balance between hepatic glucose output and insulin secretion that is maintained by a feedback loop between the liver and the pancreas [35], and less with muscle or adipose tissue insulin sensitivity, which are the organs where glucose uptake occurs in an insulin-dependent manner. In fact, the studies of in vitro arsenic effects were conducted mostly in adipocytes and not in hepatic cells. Additionally, it cannot be ruled out that the lack of correlation was influenced by metformin treatment in diabetic subjects (50%) or by the small sample size.
In a recent cross-sectional study conducted in Zimapán, México, in addition to the prevalence of diabetes being positively associated with iAs in drinking water and with the concentration of its metabolite dimethylarsinous acid in urine, an inverse association (p<0.01) between HOMA-insulin resistance and iAs exposure was also shown, contrary to our results and to in vitro findings [52]. Nonetheless, more studies in humans are needed to understand more deeply this phenomenon.
Taken together, these results provide evidence in humans that support the idea that iAs is a risk factor for T2DM through the disturbance of beta-cell function. As is known, individuals with beta-cells unable to sustain increased insulin secretion to compensate for insulin resistance will develop T2DM, and thus inadequate beta-cell function is essential to the course of the disease [28]. Here we show that both iAs exposure, as well as presence of the at-risk alleles of SNP-43 and SNP-44 in CAPN-10, a protein that plays an active role in insulin secretion as part of the machinery of the secretion complex [12], have an adverse effect on beta-cell function. It is possible that beta-cell failure may occur earlier and/or in an exacerbated manner in individuals with the at-risk genotypes in SNP-44 (T/C) and/or in subjects with the at-risk genotypes in SNP-43 (G/G) and exposure to iAs.
Epidemiological associations between chemical exposure and disorders of glucose metabolism are increasing every day. According to The Environmental Protection Agency (EPA), an endocrine disrupting chemical (EDC) is defined as an exogenous agent that interferes with the production, release, transport, metabolism, binding, action, or elimination of natural hormones in the body responsible for the maintenance of homeostasis, reproduction, development and/or behavior [1]. iAs as well as other pollutants, such as polychlorinated biphenyls, organochlorine pesticides, dioxins, and bisphenol A, among others [1], [53], [54], need to be seriously taken into account as environmental factors that increase the risk for T2DM.
Finally, we believe this study improves the current understanding of T2DM, particularly by adding evidence of iAs acting as a diabetogen in humans and contributing to the fact that environmental factors other than diet and exercise, together with genetic influences, are involved in the development of the disease. In additional to epidemiological studies, it is very important to conduct biological analyses in humans and to find the way to eliminate efficiently iAs exposure.
Supporting Information
Arsenic metabolism. Data are represented by mean ± SD and median with 25th and 75th percentiles. Symbol denotes Log-transformation for analysis (£). Symbol (*) denotes statistical significance p<0.05.
(DOC)
Allelic frequencies of CAPN-10 polymorphisms (SNP-44, -43, -63 and Indel-19) in non-diabetic and diabetic subjects. p values of the deviation from Hardy-Weinberg equilibrium were 0.419, 0.879, 0.339 and 0.945 for SNP-44, SNP-43, Indel-19 and SNP-63, respectively for non-diabetic subjects. For diabetic subjects the p values were 0.544, 0.610, 0.436 and 0.376 for SNP-44, SNP-43, Indel-19 and SNP-63, respectively.
(DOC)
SNP-43, Indel-19 and SNP-63 haplotype frequency in non-diabetic and diabetic subjects. The nomenclature for haplotype is as follows: SNP-43, allele 1, G, allele 2, A; Indel-19, allele 1, 2 repeats of 32 bp sequence, allele 2, 3 repeats; SNP-63, allele 1, C, allele 2, T. The haplotype associated with T2DM is 112/121 (¥), denoting G/G for SNP43, 2 repeats/3 repeats for Indel-19 and T/C for SNP-63.
(DOC)
Acknowledgments
The authors thank Michael Kopplin for performing the analyses of arsenic quantification in urine and water samples.
Funding Statement
The work was partially supported by CONACYT, DGAPA-PAPIIT (UNAM) and by the Dean Carter Binational Center for Environmental Health Sciences. The analyses for arsenic determination in water and urine samples were performed by the Analytical Section of the Hazard Identification Core from the NIEHS-supported Superfund Basic Research Program Grant (NIH ES-04940). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Neel BA, Sargis RM (2011) The paradox of progress: environmental disruption of metabolism and the diabetes epidemic. Diabetes 60: 1838–1848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Rahman M, Tondel M, Ahmad SA, Axelson O (1998) Diabetes mellitus associated with arsenic exposure in Bangladesh. Am J Epidemiol 148: 198–203. [DOI] [PubMed] [Google Scholar]
- 3. Rahman M, Tondel M, Chowdhury IA, Axelson O (1999) Relations between exposure to arsenic, skin lesions, and glucosuria. Occup Environ Med 56: 277–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Lai MS, Hsueh YM, Chen CJ, Shyu MP, Chen SY, et al. (1994) Ingested inorganic arsenic and prevalence of diabetes mellitus. Am J Epidemiol 139: 484–492. [DOI] [PubMed] [Google Scholar]
- 5. Tseng CH, Tai TY, Chong CK, Tseng CP, Lai MS, et al. (2000) Long-term arsenic exposure and incidence of non-insulin-dependent diabetes mellitus: a cohort study in arseniasis-hyperendemic villages in Taiwan. Environ Health Perspect 108: 847–851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Wang SL, Chiou JM, Chen CJ, Tseng CH, Chou WL, et al. (2003) Prevalence of non-insulin-dependent diabetes mellitus and related vascular diseases in southwestern arseniasis-endemic and nonendemic areas in Taiwan. Environ Health Perspect 111: 155–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Coronado-González JA, Del Razo LM, García-Vargas G, Sanmiguel-Salazar F, Escobedo-de la Peña J (2007) Inorganic arsenic exposure and type 2 diabetes mellitus in Mexico. Environ Res 104: 383–389. [DOI] [PubMed] [Google Scholar]
- 9. Ettinger AS, Zota AR, Amarasiriwardena CJ, Hopkins MR, Schwartz J, et al. (2009) Maternal arsenic exposure and impaired glucose tolerance during pregnancy. Environ Health Perspect 117: 1059–1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Díaz-Villaseñor A, Sánchez-Soto MC, Cebrián ME, Ostrosky-Wegman P, Hiriart M (2006) Sodium arsenite impairs insulin secretion and transcription in pancreatic beta-cells. Toxicol Appl Pharmacol 214: 30–34. [DOI] [PubMed] [Google Scholar]
- 11. Díaz-Villaseñor A, Burns AL, Salazar AM, Sordo M, Hiriart M, et al. (2008) Arsenite reduces insulin secretion in rat pancreatic beta-cells by decreasing the calcium-dependent calpain-10 proteolysis of SNAP-25. Toxicol Appl Pharmacol [DOI] [PubMed] [Google Scholar]
- 12. Marshall C, Hitman GA, Partridge CJ, Clark A, Ma H, et al. (2005) Evidence that an isoform of calpain-10 is a regulator of exocytosis in pancreatic beta-cells. Mol Endocrinol 19: 213–224. [DOI] [PubMed] [Google Scholar]
- 13. Fu J, Woods CG, Yehuda-Shnaidman E, Zhang Q, Wong V, et al. (2010) Low-level arsenic impairs glucose-stimulated insulin secretion in pancreatic beta cells: involvement of cellular adaptive response to oxidative stress. Environ Health Perspect 118: 864–870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Lu TH, Su CC, Chen YW, Yang CY, Wu CC, et al. (2011) Arsenic induces pancreatic beta-cell apoptosis via the oxidative stress-regulated mitochondria-dependent and endoplasmic reticulum stress-triggered signaling pathways. Toxicol Lett 201: 15–26. [DOI] [PubMed] [Google Scholar]
- 15. Paul DS, Harmon AW, Devesa V, Thomas DJ, Styblo M (2007) Molecular mechanisms of the diabetogenic effects of arsenic: inhibition of insulin signaling by arsenite and methylarsonous acid. Environ Health Perspect 115: 734–742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Xue P, Hou Y, Zhang Q, Woods CG, Yarborough K, et al. (2011) Prolonged inorganic arsenite exposure suppresses insulin-stimulated AKT S473 phosphorylation and glucose uptake in 3T3-L1 adipocytes: involvement of the adaptive antioxidant response. Biochem Biophys Res Commun 407: 360–365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Izquierdo-Vega JA, Soto CA, Sánchez-Peña LC, De Vizcaya-Ruíz A, Del Razo LM (2006) Diabetogenic effects and pancreatic oxidative damage in rats subchronically exposed to arsenite. Toxicol Lett 160: 135–142. [DOI] [PubMed] [Google Scholar]
- 18. Paul DS, Hernandez-Zavala A, Walton FS, Adair BM, Dedina J, et al. (2007) Examination of the effects of arsenic on glucose homeostasis in cell culture and animal studies: development of a mouse model for arsenic-induced diabetes. Toxicol Appl Pharmacol 222: 305–314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Paul DS, Devesa V, Hernandez-Zavala A, Adair BM, Walton FS, et al. (2008) Environmental arsenic as a disruptor of insulin signaling. Met Ions Biol Med 10: 1–7. [PMC free article] [PubMed] [Google Scholar]
- 20. Paul DS, Walton FS, Saunders RJ, Styblo M (2011) Characterization of the impaired glucose homeostasis produced in C57BL/6 mice by chronic exposure to arsenic and high-fat diet. Environ Health Perspect 119: 1104–1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Thayer KA, Heindel JJ, Bucher JR, Gallo MA (2012) Role of environmental chemicals in diabetes and obesity: a National Toxicology Program workshop review. Environ Health Perspect 120: 779–789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Horikawa Y, Oda N, Cox NJ, Li X, Orho-Melander M, et al. (2000) Genetic variation in the gene encoding calpain-10 is associated with type 2 diabetes mellitus. Nat Genet 26: 163–175. [DOI] [PubMed] [Google Scholar]
- 23. Del Bosque-Plata L, Aguilar-Salinas CA, Tusié-Luna MT, Ramírez-Jimenez S, Rodríguez-Torres M, et al. (2004) Association of the calpain-10 gene with type 2 diabetes mellitus in a Mexican population. Mol Genet Metab 81: 122–126. [DOI] [PubMed] [Google Scholar]
- 24. Weedon MN, Schwarz PE, Horikawa Y, Iwasaki N, Illig T, et al. (2003) Meta-analysis and a large association study confirm a role for calpain-10 variation in type 2 diabetes susceptibility. Am J Hum Genet 73: 1208–1212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Turner MD, Fulcher FK, Jones CV, Smith BT, Aganna E, et al. (2007) Calpain facilitates actin reorganization during glucose-stimulated insulin secretion. Biochem Biophys Res Commun 352: 650–655. [DOI] [PubMed] [Google Scholar]
- 26. Paul DS, Harmon AW, Winston CP, Patel YM (2003) Calpain facilitates GLUT4 vesicle translocation during insulin-stimulated glucose uptake in adipocytes. Biochem J 376: 625–632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Aganna E, Burrin JM, Hitman GA, Turner MD (2006) Involvement of calpain and synaptotagmin Ca2+ sensors in hormone secretion from excitable endocrine cells. J Endocrinol 190: R1–7. [DOI] [PubMed] [Google Scholar]
- 28. Kahn SE, Hull RL, Utzschneider KM (2006) Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 444: 840–846. [DOI] [PubMed] [Google Scholar]
- 29. American Diabetes Association (2012) Diagnosis and classification of diabetes mellitus. Diabetes Care 35 Suppl 1: S64–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Meza MM, Yu L, Rodriguez YY, Guild M, Thompson D, et al. (2005) Developmentally restricted genetic determinants of human arsenic metabolism: association between urinary methylated arsenic and CYT19 polymorphisms in children. Environ Health Perspect 113: 775–781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Meza MM, Kopplin MJ, Burgess JL, Gandolfi AJ (2004) Arsenic drinking water exposure and urinary excretion among adults in the Yaqui Valley, Sonora, Mexico. Environ Res 96: 119–126. [DOI] [PubMed] [Google Scholar]
- 32. Del Razo LM, Garcia-Vargas GG, Vargas H, Albores A, Gonsebatt ME, et al. (1997) Altered profile of urinary arsenic metabolites in adults with chronic arsenicism. A pilot study. Arch Toxicol 71: 211–217. [DOI] [PubMed] [Google Scholar]
- 33.The Oxford Centre for Diabetes Endocrinology and Metabolism HOMA Calculator.
- 34. Matthews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher DF, et al. (1985) Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 28: 412–419. [DOI] [PubMed] [Google Scholar]
- 35. Wallace TM, Levy JC, Matthews DR (2004) Use and abuse of HOMA modeling. Diabetes Care 27: 1487–1495. [DOI] [PubMed] [Google Scholar]
- 36. Emoto M, Nishizawa Y, Maekawa K, Hiura Y, Kanda H, et al. (1999) Homeostasis model assessment as a clinical index of insulin resistance in type 2 diabetic patients treated with sulfonylureas. Diabetes Care 22: 818–822. [DOI] [PubMed] [Google Scholar]
- 37. Orho-Melander M, Klannemark M, Svensson MK, Ridderstrale M, Lindgren CM, et al. (2002) Variants in the calpain-10 gene predispose to insulin resistance and elevated free fatty acid levels. Diabetes 51: 2658–2664. [DOI] [PubMed] [Google Scholar]
- 38. Stephens M, Smith NJ, Donnelly P (2001) A new statistical method for haplotype reconstruction from population data. Am J Hum Genet 68: 978–989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Stephens M, Scheet P (2005) Accounting for decay of linkage disequilibrium in haplotype inference and missing-data imputation. Am J Hum Genet 76: 449–462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Insitute of Human Genetics Helmholtz Center Munich (2012) DeFinetti Program.
- 41. Baier LJ, Permana PA, Yang X, Pratley RE, Hanson RL, et al. (2000) A calpain-10 gene polymorphism is associated with reduced muscle mRNA levels and insulin resistance. J Clin Invest 106: R69–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Hoffstedt J, Ryden M, Lofgren P, Orho-Melander M, Groop L, et al. (2002) Polymorphism in the Calpain 10 gene influences glucose metabolism in human fat cells. Diabetologia 45: 276–282. [DOI] [PubMed] [Google Scholar]
- 43. Stumvoll M, Fritsche A, Madaus A, Stefan N, Weisser M, et al. (2001) Functional significance of the UCSNP-43 polymorphism in the CAPN10 gene for proinsulin processing and insulin secretion in nondiabetic Germans. Diabetes 50: 2161–2163. [DOI] [PubMed] [Google Scholar]
- 44. Shore AC, Evans JC, Frayling TM, Clark PM, Lee BC, et al. (2002) Association of calpain-10 gene with microvascular function. Diabetologia 45: 899–904. [DOI] [PubMed] [Google Scholar]
- 45. Logie LJ, Brown AE, Yeaman SJ, Walker M (2005) Calpain inhibition and insulin action in cultured human muscle cells. Mol Genet Metab 85: 54–60. [DOI] [PubMed] [Google Scholar]
- 46. Brown AE, Yeaman SJ, Walker M (2007) Targeted suppression of calpain-10 expression impairs insulin-stimulated glucose uptake in cultured primary human skeletal muscle cells. Mol Genet Metab 91: 318–324. [DOI] [PubMed] [Google Scholar]
- 47. Day C, Bailey CJ (2011) Obesity in the pathogenesis of type 2 diabetes. The British Journal of Diabetes and Vascular Disease 11: 55–61. [Google Scholar]
- 48. Lyssenko V, Groop L (2009) Genome-wide association study for type 2 diabetes: clinical applications. Curr Opin Lipidol 20: 87–91. [DOI] [PubMed] [Google Scholar]
- 49. Imamura M, Maeda S (2011) Genetics of type 2 diabetes: the GWAS era and future perspectives [Review]. Endocr J 58: 723–739. [DOI] [PubMed] [Google Scholar]
- 50. Tschritter O, Fritsche A, Shirkavand F, Machicao F, Haring H, et al. (2003) Assessing the shape of the glucose curve during an oral glucose tolerance test. Diabetes Care 26: 1026–1033. [DOI] [PubMed] [Google Scholar]
- 51. Smeester L, Rager JE, Bailey KA, Guan X, Smith N, et al. (2011) Epigenetic changes in individuals with arsenicosis. Chem Res Toxicol 24: 165–167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Del Razo LM, Garcia-Vargas GG, Valenzuela OL, Castellanos EH, Sanchez-Pena LC, et al. (2011) Exposure to arsenic in drinking water is associated with increased prevalence of diabetes: a cross-sectional study in the Zimapan and Lagunera regions in Mexico. Environ Health 10: 73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Lee DH, Steffes MW, Sjodin A, Jones RS, Needham LL, et al. (2011) Low dose organochlorine pesticides and polychlorinated biphenyls predict obesity, dyslipidemia, and insulin resistance among people free of diabetes. PLoS One 6: e15977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Makaji E, Raha S, Wade MG, Holloway AC (2011) Effect of environmental contaminants on Beta cell function. Int J Toxicol 30: 410–418. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Arsenic metabolism. Data are represented by mean ± SD and median with 25th and 75th percentiles. Symbol denotes Log-transformation for analysis (£). Symbol (*) denotes statistical significance p<0.05.
(DOC)
Allelic frequencies of CAPN-10 polymorphisms (SNP-44, -43, -63 and Indel-19) in non-diabetic and diabetic subjects. p values of the deviation from Hardy-Weinberg equilibrium were 0.419, 0.879, 0.339 and 0.945 for SNP-44, SNP-43, Indel-19 and SNP-63, respectively for non-diabetic subjects. For diabetic subjects the p values were 0.544, 0.610, 0.436 and 0.376 for SNP-44, SNP-43, Indel-19 and SNP-63, respectively.
(DOC)
SNP-43, Indel-19 and SNP-63 haplotype frequency in non-diabetic and diabetic subjects. The nomenclature for haplotype is as follows: SNP-43, allele 1, G, allele 2, A; Indel-19, allele 1, 2 repeats of 32 bp sequence, allele 2, 3 repeats; SNP-63, allele 1, C, allele 2, T. The haplotype associated with T2DM is 112/121 (¥), denoting G/G for SNP43, 2 repeats/3 repeats for Indel-19 and T/C for SNP-63.
(DOC)