

Abstract
Streptococcus pneumoniae is a diverse species causing invasive as well as localized infections that result in massive global morbidity and mortality. Strains vary markedly in pathogenic potential, but the molecular basis is obscured by the diversity and plasticity of the pneumococcal genome. In the present study, S. pneumoniae serotype 3 blood (n = 12) or ear (n = 13) isolates were multilocus sequence typed (MLST) and assessed for biofilm formation and virulence phenotype. Blood and ear isolates exhibited similar MLST distributions but differed markedly in phenotype. Blood isolates formed robust biofilms only at pH 7.4, which were enhanced in Fe(III)-supplemented medium. Conversely, ear isolates formed biofilms only at pH 6.8, and Fe(III) was inhibitory. Biofilm formation paralleled luxS expression and genetic competence. In a mouse intranasal challenge model, blood isolates did not stably colonize the nasopharynx but spread to the blood; none spread to the ear. Ear isolates colonized the nasopharynx at higher levels and also spread to the ear compartment in a significant proportion of animals; none caused bacteremia. Thus, pneumococci of the same serotype and MLST exhibit distinct phenotypes in accordance with clinical site of isolation, indicative of stable niche adaptation within a clonal lineage.
INTRODUCTION
Streptococcus pneumoniae (the pneumococcus) is responsible for massive global morbidity and mortality. It is a major cause of pneumonia, meningitis, and sepsis, especially in young children and the elderly. S. pneumoniae also causes less serious but highly prevalent infections such as otitis media (OM) and sinusitis (1–5). The World Health Organization estimates that 1.6 million people, of whom 0.7 to 1 million are under the age of 5, die of pneumococcal diseases each year, with the highest incidence in developing countries. Indeed, S. pneumoniae accounts for more deaths worldwide than any other single pathogen (6, 7). In spite of this mortality, S. pneumoniae is part of the commensal nasopharyngeal flora of humans. Most colonized individuals are asymptomatic, and carriers are the principal reservoirs for transmission of S. pneumoniae in the community. In a small proportion of carriers, which nevertheless translates into globally significant total case numbers, S. pneumoniae invades from its nasopharyngeal beachhead to cause disease. This may occur, for example, by aspiration into the lungs to cause pneumonia, by direct invasion of the blood, or by ascension of the Eustachian tube to access the middle ear and cause OM (1, 8, 9). However, the mechanisms whereby pneumococci transition from commensal to pathogen are poorly understood. OM is one of the most common pediatric diagnoses (10), and although not usually life-threatening, it has a massive socioeconomic impact (11).
The capacity to form biofilms is increasingly being recognized as a critical event in the pathogenesis of OM and other pneumococcal diseases, and several studies have shown a strong correlation between biofilm formation in vitro and colonization and lung infection in mice (12–15). Pneumococcal OM is also frequently associated with previous viral respiratory infections that lead to stasis, congestion, and blockade of the normal mucosal ciliary function, as well as Eustachian tube obstruction in very young children, thereby predisposing to secondary bacterial infection (10, 16–18).
It has recently been demonstrated that pneumococcal S-ribosylhomocysteine lyase, encoded by luxS, plays an important role in biofilm formation (19, 20). This enzyme is involved in the S-adenosylhomocysteine pathway that synthesizes autoinducer 2 (AI-2), a molecule involved in bacterial quorum sensing. In this case, extracellular AI-2 recognized by the pneumococcus has an effect on the transcription of genes involved in biofilm formation. Most recently, we have characterized the impact of exogenous Fe(III), as well as the LuxS-mediated AI-2 quorum sensing system, on biofilm formation by S. pneumoniae D39 (19). Fe(III) strongly enhanced biofilm formation, while Fe(III) chelation with deferoxamine was inhibitory. Importantly, Fe(III) also upregulated expression of luxS and piuA (encoding the major iron transporter) in wild-type D39. Similarly, genetic competence, as measured by transformation frequency, as well as expression of competence genes comD, comX, comW, cglA, and dltA, and the murein hydrolase cbpD gene associated with fratricide-dependent DNA release, were all directly related to luxS expression levels and further upregulated by Fe(III).
S. pneumoniae is a genetically plastic and diverse species, comprising 93 capsular serotypes superimposed on over 5,000 clonal groups recognizable by multilocus sequence typing (MLST) (21). Capsule switching experiments also show that both serotype and genetic background influence virulence (22, 23). This strain complexity has complicated attempts to examine whether there is any association between a given clonal lineage or serotype and propensity to cause invasive rather than localized infections, and whether biofilm formation capacity can account for differences in virulence phenotype. In the current study, we have simplified the analysis by confining our examination to S. pneumoniae serotype 3 clinical isolates from cases of sepsis or OM. We present evidence for a strong association between clinical site of isolation, pH-dependent biofilm formation capacity, luxS expression, and virulence phenotype in a murine model.
MATERIALS AND METHODS
Bacterial strains and growth conditions.
S. pneumoniae strains used in this study are listed in Table 1. Cells were grown in a casein-based semisynthetic liquid medium (C+Y) (24) and on Columbia agar supplemented with 5% (vol/vol) horse blood (blood agar [BA]) at 37°C in a CO2-enriched atmosphere. Bacterial stocks were prepared from mid-log-phase cultures and stored at −80°C in 10% glycerol.
Table 1.
S. pneumoniae serotype 3 isolates used in this studya
| Strain | MLST | Source |
|---|---|---|
| ST232/1 | ST232 | Blood |
| ST233/3 | ST233 | Blood |
| ST180/4 | ST180 | Blood |
| ST458/5 | ST458 | Blood |
| ST233/6 | ST233 | Blood |
| ST180/7 | ST180 | Blood |
| ST180/8 | ST180 | Blood |
| ST180/15 | ST180 | Blood |
| ST458/20 | ST458 | Blood |
| ST232/23 | ST232 | Blood |
| ST180/24 | ST180 | Blood |
| ST232/25 | ST232 | Blood |
| ST180/2 | ST180 | Ear |
| ST180/9 | ST180 | Ear |
| ST180/10 | ST180 | Ear |
| ST232/11 | ST232 | Ear |
| ST180/12 | ST180 | Ear |
| ST233/13 | ST233 | Ear |
| ST180/14 | ST180 | Ear |
| ST180/16 | ST180 | Ear |
| ST232/17 | ST232 | Ear |
| ST232/18 | ST232 | Ear |
| ST232/19 | ST232 | Ear |
| ST180/21 | ST180 | Ear |
| ST180/22 | ST180 | Ear |
Strains were isolated between 1988 and 1996 from patients at either the Women's and Children's Hospital, North Adelaide, South Australia, or the Alice Springs Hospital, Northern Territory, Australia. Similar proportions of OM and blood isolates were derived from each site.
MLST.
For MLST analysis, the aroE, gdh, gki, recP, spi, xpt, and ddl genes of the S. pneumoniae strains were PCR amplified and sequenced, as described by Spratt (25). The sequence types (ST) of strains were determined from the MLST database (http://www.mlst.net) based on the resulting allelic profiles.
Biofilm assays.
Static biofilm assays were performed by growing bacteria in 24- or 96-well flat-bottom polystyrene plates as described previously (19). Briefly, cells were grown in C+Y medium, adjusted to either pH 7.4 or 6.8, and supplemented with 100 μM Fe(III) nitrate where indicated for 24 h at 37°C in a CO2-enriched atmosphere. Microscopic examination of biofilms was performed by washing wells three times with PBS, desiccation at 50°C, staining with 1% crystal violet for 30 min, and photomicrography under transmitted light. Quantification of bacteria attached to the plastic substratum was performed by washing unstained plates three times, filling wells with 0.1 ml of fresh C+Y medium, detaching cells using a sonicating water bath, and plating serial dilutions onto blood agar. Statistical analysis was performed using 2-tailed Student's t test.
RNA extraction and quantitative real-time RT-PCR.
Selected clinical isolates were grown in C+Y with or without additional Fe(III) (100 μM) for 24 h in a static biofilm model and total RNA was extracted as previously described (26). The level of gene expression relative to that of 16S rRNA was determined using a one-step reverse transcriptase PCR (RT-PCR) kit and a LC480 real-time cycler (Roche) as described previously (19). Statistical analysis was performed using 2-tailed Student's t test.
Mutagenesis of luxS.
The luxS gene was deleted from various type 3 strains using overlap extension PCR and transformation as described previously (19).
Infection of mice.
Animal experiments were approved by the University of Adelaide Animal Ethics Committee. Female outbred 5- to 6-week-old CD-1 (Swiss) mice were inoculated intranasally with 1 × 107 CFU of S. pneumoniae (confirmed retrospectively by viable count) in a volume of 10 μl. Groups of 15 mice were inoculated for each strain, and 5 randomly selected mice from each group were euthanized by CO2 asphyxiation at 24, 48, or 72 h postinfection. Nasal wash, nasopharyngeal tissue, ear tissue, lung, brain, and blood samples were collected and processed as previously described (27, 28). Samples were serially diluted and plated onto BA plates for enumeration of viable pneumococci. Statistical analyses were done using the Mann-Whitney U test.
RESULTS
Serotype 3 biofilm formation is pH dependent.
It has been reported that certain serotypes and STs of S. pneumoniae have a greater potential to cause invasive disease in humans than others; likewise, strains differ in their propensities to cause OM (29, 30). This suggests that strains may differ in their capacity to adapt to and survive or proliferate within distinct host microenvironments. This further implies that clinical isolates from cases of otitis media may exhibit in vitro and in vivo phenotypes distinct from those of blood isolates.
Two conditions that vary significantly between different niches of the human body are metal ion concentrations (31) and pH; the pH of the blood is typically around 7.4, while in the (uninfected) ear cavity it is in the range of 6.5 to 6.8 (32). We first examined whether a relationship exists between the site of isolation and ability to form biofilms under different pH and [Fe(III)] conditions. We tested 12 blood isolates and 13 ear isolates belonging to serotype 3, a type that is frequently associated with both OM and invasive disease. Interestingly, the two groups contained representatives of the same four STs in similar proportions (ST180, ST232, ST233, and ST458, in order of prevalence) (Table 1). All 25 strains were in the opaque phase, as judged by opacity phenotype when grown on THY-catalase plates (28), and there was no significant difference in the level of type 3 capsular polysaccharide production, as determined by uronic acid assay (33) (results not presented). In the first instance, strains were grown in C+Y medium (pH 7.4) with or without supplementation with 100 μM Fe(III) nitrate and were tested in the static biofilm assay. Pneumococcal growth rates were not significantly different in the presence of this Fe(III) concentration compared with rates in standard C+Y medium, which contains 0.8 μM total Fe, as measured by inductively coupled plasma mass spectrometry (ICPMS) (data not shown). After 24 h of incubation, a marked increase in biofilm density in the presence of Fe(III) was observed in strains isolated from the blood compared with strains isolated from the ear, as judged by viable counts of dispersed, unstained biofilms (P < 0.001) (Fig. 1A). The biofilm density was also significantly greater in blood isolates in the presence of Fe(III) than in standard C+Y medium (P < 0.001) (Fig. 1A), consistent with our previous observation for the type 2 laboratory strain D39 (19). However, the presence of Fe(III) had no significant effect on the low level of biofilm formation by the type 3 ear isolates at pH 7.4 (Fig. 1A). In marked contrast, when biofilm formation assays were performed using C+Y medium with the pH adjusted to 6.8 (a level within the range found in the ear cavity), the biofilm density of the ear isolates was significantly greater than that of the blood isolates (P < 0.01). Interestingly, supplementation of C+Y (pH 6.8) medium with Fe(III) abolished the ability of ear isolates to form biofilms (P < 0.001) and did not increase the low level of biofilm formation by blood isolates at the lower pH (Fig. 1A). Microscopic examination of crystal violet-stained biofilm assay plates confirmed that robust biofilms were formed by blood isolates only at pH 7.4 and were boosted further by Fe(III), whereas ear isolates formed robust biofilms only at pH 6.8 and in the absence of Fe(III) (Fig. 2).
Fig 1.

Biofilm formation and gene expression. (A) Biofilm formation by clinical isolates (12 from blood and 13 from ear) after 24 h of growth at either pH 7.4 or 6.8, with or without 100 μM Fe(III), determined by viable count. Data are the means ± standard deviations for three independent experiments (*, P < 0.05; **, P < 0.01; and ***, P < 0.001; 2-tailed Student's t test). (B and C) Expression of luxS (B) and piuA (C) relative to 16S rRNA in the indicated clinical isolates grown either at pH 7.4 or 6.8, with or without addition of 100 μM Fe(III). Data are the means ± standard deviations for three independent experiments (*, P < 0.05; **, P < 0.01; and ***, P < 0.001; 2-tailed Student's t test).
Fig 2.
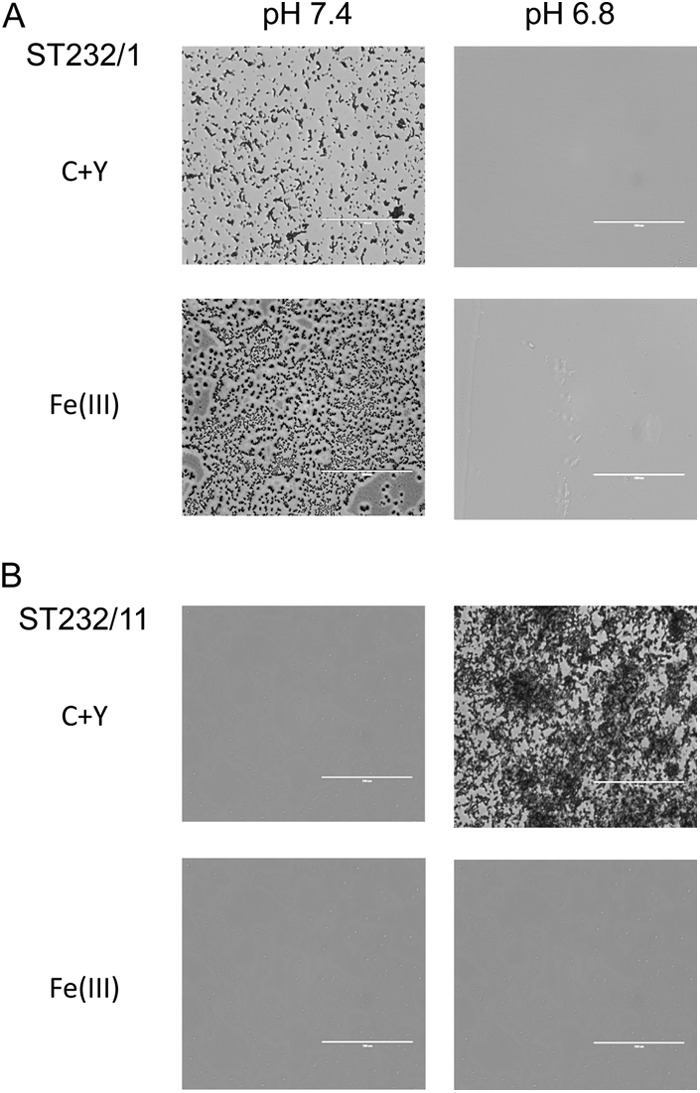
Microscopic analysis of crystal violet-stained 24-h biofilms of the blood isolate ST232/1 (A) and the ear isolate ST232/11 (B) grown at pH 7.4 or 6.8 in the presence or absence of 100 μM Fe(III). Scale bar, 100 μm.
LuxS is involved in serotype 3 biofilm formation.
To test whether the iron- and pH-dependent biofilm formation of serotype 3 strains is linked to the activity of the LuxS quorum sensing system, a relationship previously identified in strain D39 serotype 2 (19), the level of luxS and piuA expression was measured using real-time RT-PCR. For this analysis, one blood isolate and one ear isolate belonging to each of the three major STs (ST180, ST232, and ST233) were selected. The difference in luxS expression levels (Fig. 1B) between strains and growth conditions closely paralleled the pattern of biofilm formation (Fig. 1A). In the blood isolates ST180/15, ST232/1, and ST233/3, luxS expression was significantly higher when cells were grown at pH 7.4 in the presence of Fe(III) than under other growth conditions (P < 0.001 for ST180/15 and ST232/1; P < 0.01 for ST233/3). In contrast, for the ear isolates ST180/2, ST232/11, and ST233/13, luxS expression was significantly higher when cells were grown at pH 6.8 in the absence of Fe(III) than under any of the other conditions (P < 0.001 for ST180/2 and ST233/13; P < 0.05 for ST232/11). However, for each ST pair, the overall level of expression of luxS of the ear isolate was lower than that of the corresponding blood isolate under their respective optimal conditions (P < 0.001 for ST180 and ST232; P < 0.01 for ST233) (Fig. 1B).
Since iron has been shown to stimulate biofilm formation in the blood isolates, we also quantified the expression level of piuA, which encodes the major pneumococcal iron uptake system (34). A significantly higher level of expression of piuA was observed in blood isolates grown at pH 7.4 in the presence of Fe(III) than under other growth conditions (P < 0.01 for ST180/15 and ST233/3; P < 0.001 for ST232/1) (Fig. 1C). In contrast, piuA expression was very low in all the ear isolates, and there was no evidence for upregulation of piuA in these isolates in the presence of iron, except for ST233/13 at pH 7.4 (Fig. 1C).
luxS mutation reduces the ability of serotype 3 clinical isolates to form biofilms.
To further confirm the role of luxS in biofilm formation by serotype 3 strains, luxS was deleted in all 6 clinical isolates mentioned above and the mutants were then tested for biofilm forming ability. Viable counts of the dispersed biofilms revealed a significant reduction in biofilm formation for the luxS mutants of the three ear isolates compared to the wild type when grown in C+Y (pH 6.8) medium (P < 0.001 for ST180/2 and ST233/13; P < 0.01 for ST232/11) (Fig. 3A). Microscopic examination after crystal violet staining confirmed the results of viable counts (Fig. 4). A similar finding was also observed for the luxS mutants of all three blood isolates grown at pH 7.4 in the presence of Fe(III) (data not shown).
Fig 3.

(A) Effect of luxS mutation on biofilm formation by ear isolates, determined by viable count. Data are the means ± standard deviations for three independent experiments (**, P < 0.01, and ***, P < 0.001; 2-tailed Student's t test). (B) Transformability of the reference invasive type 2 strain D39 and the type 3 clinical isolates (from blood or ear) grown at either pH 7.4 or 6.8. Data are the total numbers of transformants (means ± standard deviations for three independent experiments) (**, P < 0.01, and ***, P < 0.001; 2-tailed Student's t test). Each transformation reaction mixture contained approximately 107 competent pneumococci.
Fig 4.
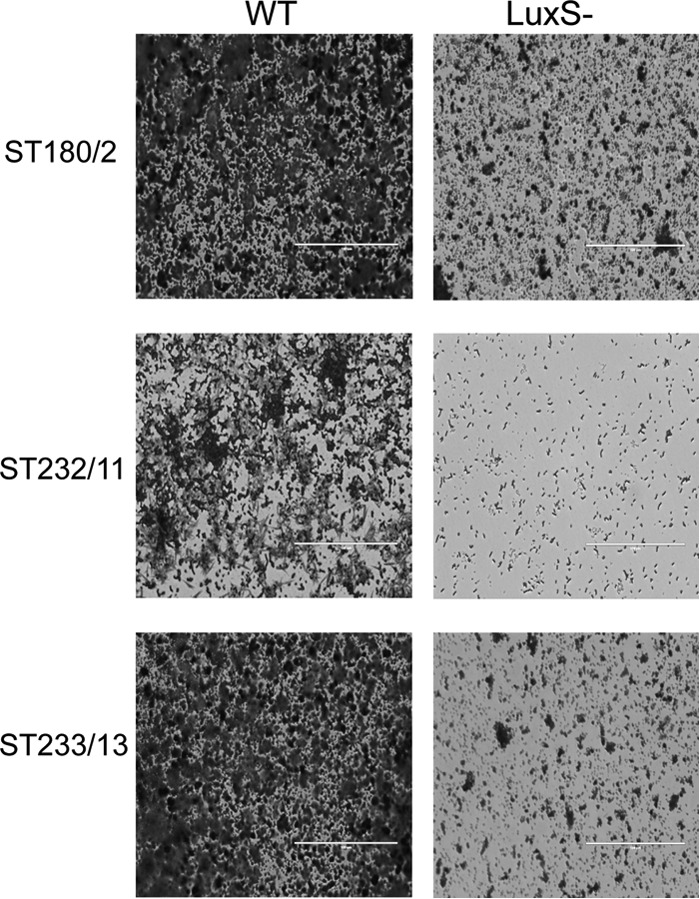
Microscopic analysis of crystal violet-stained biofilms formed at pH 6.8 by ear isolates and their respective luxS mutant derivatives. WT, wild type. Scale bar, 100 μm.
Transformability is pH dependent in serotype 3 strains.
Induction of the competence state has been shown to parallel biofilm formation in S. pneumoniae (13, 19), and so the transformability of the six serotype 3 clinical isolates, as well as the reference invasive type 2 strain D39, was measured in planktonic cells grown at pH 7.4 and pH 6.8, as described previously (19). The transformability of the strains was found to be strongly influenced by the pH; blood isolates exhibited a significantly greater propensity to take up external DNA at pH 7.4 (P < 0.001 for ST180/15 and ST232/1; P < 0.01 for ST233/3), as did the reference strain D39. In contrast, the ear isolates showed higher rates of transformability at pH 6.8 (P < 0.001 for ST180/2 and ST232/11; P < 0.01 for ST233/13) (Fig. 3B). Indeed, for the ST180 and ST232 strains, the blood isolates were completely untransformable at pH 6.8, while the ear isolates were not transformable at pH 7.4. Strains of ST233 were transformable under both conditions tested, but a significant increase in efficiency was observed for the blood isolate at pH 7.4 and ear isolate at pH 6.8.
Clinical isolation site corresponds with virulence phenotype in mice.
To investigate the virulence profile of the clinical isolates, we used a murine nasopharyngeal inoculation model, which mimics the natural route of infection for S. pneumoniae. At all the time points examined (24, 48, and 72 h postinfection), the majority of mice in each group infected with blood isolates (ST180/15, ST232/1, or ST233/3) showed bacteremia, whereas no bacteremia could be detected in any of the mice in groups challenged with the ear isolates (ST180/2, ST232/11, or ST233/13) (Fig. 5C). Collectively, the degree of bacteremia for blood isolates was significantly greater than that for the combined ear isolate groups at all time points (P < 0.001). When individual groups within an ST were compared, the bacteremia level for the ST180/15 group was significantly greater than that for the ST180/2 group at all time points (P < 0.05), while bacteremia levels in the ST232/1 and ST233/3 groups were significantly greater than those for ST232/11 and ST233/13, respectively, at 48 h (P < 0.05). The situation was reversed in the nasopharyngeal tissue (Fig. 5A), where blood isolates as a whole were inferior to ear isolates in terms of overall colonization levels at 48 and 72 h (P < 0.001 in both cases). Comparing the individual STs, ST232/11 colonized the nasopharyngeal tissue to a significantly greater extent than ST232/1 at both 48 and 72 h (P < 0.05 and P < 0.01, respectively), while ST180/2 and ST233/13 exhibited significantly higher rates of colonization at 72 h than the corresponding ST-matched blood isolate (P < 0.01 and P < 0.05, respectively). Bacteria in ear tissue samples were detected only in groups challenged with the ear isolates, and this difference was statistically significant (P < 0.05) at both 48 and 72 h (Fig. 5B).
Fig 5.

Virulence phenotypes of blood and ear isolates. Groups of 15 mice were infected intranasally with 107 CFU of the indicated strain. At the indicated times, 5 mice from each group were euthanized and numbers of pneumococci in the indicated tissues were quantitated. (A) Nasopharyngeal tissue; (B) ear; (C) blood. Viable counts are shown for each mouse at each site; horizontal bars indicate the median value for each group/time point. Blood isolates are represented by solid symbols; ear isolates are represented by open symbols. Differences between groups were analyzed by 1-tailed Mann-Whitney U test (*, P < 0.05; **, P < 0.01; and ***, P < 0.001).
Interestingly, at 24 h postinfection, pneumococci could not be detected in the lungs of any of the mice challenged with blood isolates, in spite of significant bacteremia in the majority of animals in each ST group (Fig. 6C). Moreover, only 4 of the 15 mice challenged with blood isolates had evidence of pneumococci in the lungs at either 48 or 72 h. In contrast, no bacteria could be detected in the lungs of any of the mice challenged with the ear isolates. Similar findings were observed in brain tissue (Fig. 6B). It is also interesting to note that pneumococci could not be detected in nasopharyngeal washes of any mice 24 h postinfection (Fig. 6A). At 48 h, less than half of the mice had pneumococci in nasal wash fluid, with fewer again at 72 h.
Fig 6.

Virulence phenotypes of blood and ear isolates. Groups of 15 mice were infected intranasally with 107 CFU of the indicated strain. At the indicated times, 5 mice from each group were euthanized and numbers of pneumococci in the indicated tissues were quantitated. (A) Nasal wash; (B) brain; (C) lungs. Viable counts are shown for each mouse at each site; horizontal bars indicate the median value for each group and time point. Blood isolates are represented by solid symbols; ear isolates are represented by open symbols. Differences between groups were analyzed by 1-tailed Mann-Whitney U test (*, P < 0.05).
DISCUSSION
S. pneumoniae is a diverse and adaptable pathogen, capable of surviving in a range of niches within its human host. Previous studies from our laboratory have identified changes in in vivo transcriptional profile within a given strain that facilitate survival in distinct host niches (nasopharynx, lungs, blood, and brain) and/or aid progression from one niche to another (26, 27, 35, 36). However, genetic differences between strains also result in pneumococci with inherent tropism for one niche over another. Studies in animal models suggest that both capsular and noncapsular loci contribute to these differences in in vivo phenotype (22, 23, 37–40), but interpretation of molecular epidemiological analyses of human isolates is complicated by the vast genetic diversity of strains.
In the present study, we have examined phenotypic differences between clinical isolates from either sepsis or OM (n = 12 and 13, respectively), focusing on strains belonging to serotype 3, an important cause of both systemic and localized pneumococcal infections. All 25 strains were in the opaque phase and produced similar amounts of type 3 capsular polysaccharide. Moreover, the groups of blood and ear isolates comprised the same four MLSTs in similar proportions. Surprisingly, however, the in vitro and in vivo phenotypes of the two groups were strikingly different. Only the blood isolates were capable of forming in vitro biofilms at pH 7.4, and this property was significantly augmented by supplementation of the medium with Fe(III). On the other hand, only the ear isolates were capable of forming biofilms at pH 6.8, and at this pH, Fe(III) was inhibitory.
We then conducted more detailed phenotypic comparisons of matched pairs of isolates from either the blood or ear belonging to the dominant MLSTs (ST180, ST232, and ST233). We have previously reported that the LuxS quorum sensing system is a central regulator of biofilm formation in the invasive type 2 S. pneumoniae strain D39, and in the present study, there was again a close parallel between expression of the luxS gene and biofilm formation at the permissive pH for the respective groups. Moreover, the stimulatory effect of Fe(III) on biofilm formation by the blood isolates at pH 7.4 was matched by upregulated expression of piuA, which encodes the major pneumococcal iron transporter. In contrast, piuA expression was negligible in blood isolates at the nonpermissive pH of 6.8 or in the absence of Fe(III), and in the ear isolates under any of the conditions tested.
Although the level of luxS expression in each of the ear isolates at pH 6.8 in the absence of Fe(III) was significantly greater than that under the other environmental conditions, luxS expression in each of the ear isolates was significantly lower than that in the blood isolate of the corresponding ST under its optimal conditions. This suggested that luxS might play a less important role in biofilm formation by ear isolates relative to blood isolates. Nevertheless, mutagenesis of luxS significantly reduced biofilm formation, regardless of the source of the isolate. The link between the luxS system and genetic competence previously observed in D39 also held for the type 3 isolates, with maximal transformation efficiencies observed under the same environmental conditions as were permissive for biofilm formation.
The consistent and stark phenotypic distinction between blood and ear isolates in terms of preferred pH and requirement for Fe(III) for optimal biofilm formation suggests that there are fundamental genetic differences between S. pneumoniae strains belonging to the same clonal lineage that enable adaptation to the distinct host niches from whence they were isolated. This adaptation to the human niche was closely mimicked by the behavior of the various strains in a mouse intranasal challenge model. All three blood isolates were poor colonizers of the nasopharynx, yet they were able to readily spread directly to the blood in most animals, largely bypassing the lungs. Moreover, none of the blood isolates ever spread to the ear compartment. On the other hand, the ear isolates were able to stably colonize the nasopharyngeal tissue of the vast majority of animals, but they never spread to the blood, brain, or lungs. Nearly half of the animals challenged with OM isolates had pneumococci in the ear compartment by 72 h.
In this study, we have provided compelling evidence that stable adaptation of pneumococci to distinct host niches occurs within clonal lineages. Thus, molecular epidemiological studies aimed at associating MLST with potential to cause invasive versus noninvasive disease should be interpreted with considerable caution. It is now clear that accessory regions outside the core pneumococcal genome are major determinants of virulence phenotype in both humans and animal models. Identification of multiple ST-matched pairs of S. pneumoniae serotype 3 strains with distinct human tissue tropism and virulence profile provides a unique opportunity to identify accessory regions or polymorphisms within the core genome common to isolates from one niche versus the other by genome sequence analysis. This, in turn, will permit direct testing of the role of identified regions in pathogenesis by targeted mutagenesis.
ACKNOWLEDGMENTS
This work was supported by the National Health and Medical Research Council (NHMRC) of Australia (Program Grant 565526 to J.C.P. and A.W.P. and NHMRC Australia Fellowship to J.C.P.), the Australian Research Council (DORA Fellowship to A.W.P.), and the Garnett Passe and Rodney Williams Memorial Foundation (Training Fellowship to C.T.).
Footnotes
Published ahead of print 3 December 2012
REFERENCES
- 1. Gray BM, Converse GM, III, Dillon HC., Jr 1980. Epidemiologic studies of Streptococcus pneumoniae in infants: acquisition, carriage, and infection during the first 24 months of life. J. Infect. Dis. 142:923–933 [DOI] [PubMed] [Google Scholar]
- 2. Ispahani P, Slack RC, Donald FE, Weston VC, Rutter N. 2004. Twenty year surveillance of invasive pneumococcal disease in Nottingham: serogroups responsible and implications for immunisation. Arch. Dis. Child. 89:757–762 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Hussain M, Melegaro A, Pebody RG, George R, Edmunds WJ, Talukdar R, Martin SA, Efstratiou A, Miller E. 2005. A longitudinal household study of Streptococcus pneumoniae nasopharyngeal carriage in a UK setting. Epidemiol. Infect. 133:891–898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Kadioglu A, Weiser JN, Paton JC, Andrew PW. 2008. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Microbiol. 6:288–301 [DOI] [PubMed] [Google Scholar]
- 5. Varon E, Mainardi JL, Gutmann L. 2010. Streptococcus pneumoniae: still a major pathogen. Clin. Microbiol. Infect. 16:401. [DOI] [PubMed] [Google Scholar]
- 6. WHO 2007. Pneumococcal conjugate vaccine for childhood immunization—WHO position paper. Wkly. Epidemiol. Rec. 82:93–104 [PubMed] [Google Scholar]
- 7. Mulholland K. 2003. Global burden of acute respiratory infections in children: implications for interventions. Pediatr. Pulmonol. 36:469–474 [DOI] [PubMed] [Google Scholar]
- 8. Brueggemann AB, Peto TE, Crook DW, Butler JC, Kristinsson KG, Spratt BG. 2004. Temporal and geographic stability of the serogroup-specific invasive disease potential of Streptococcus pneumoniae in children. J. Infect. Dis. 190:1203–1211 [DOI] [PubMed] [Google Scholar]
- 9. Sleeman KL, Griffiths D, Shackley F, Diggle L, Gupta S, Maiden MC, Moxon ER, Crook DW, Peto TE. 2006. Capsular serotype-specific attack rates and duration of carriage of Streptococcus pneumoniae in a population of children. J. Infect. Dis. 194:682–688 [DOI] [PubMed] [Google Scholar]
- 10. Parsons DS, Wald ER. 1996. Otitis media and sinusitis: similar diseases. Otolaryngol. Clin. North Am. 29:11–25 [PubMed] [Google Scholar]
- 11. Teele DW, Klein JO, Rosner B. 1989. Epidemiology of otitis media during the first seven years of life in children in greater Boston: a prospective, cohort study. J. Infect. Dis. 160:83–94 [DOI] [PubMed] [Google Scholar]
- 12. Oggioni MR, Trappetti C, Kadioglu A, Cassone M, Iannelli F, Ricci S, Andrew PW, Pozzi G. 2006. Switch from planktonic to sessile life: a major event in pneumococcal pathogenesis. Mol. Microbiol. 61:1196–1210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Trappetti C, Gualdi L, Di Meola L, Jain P, Korir CC, Edmonds P, Iannelli F, Ricci S, Pozzi G, Oggioni MR. 2011. The impact of the competence quorum sensing system on Streptococcus pneumoniae biofilms varies depending on the experimental model. BMC Microbiol. 11:75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Muñoz-Elías EJ, Marcano J, Camilli A. 2008. Isolation of Streptococcus pneumoniae biofilm mutants and their characterization during nasopharyngeal colonization. Infect. Immun. 76:5049–5061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Trappetti C, Kadioglu A, Carter M, Hayre J, Iannelli F, Pozzi G, Andrew PW, Oggioni MR. 2009. Sialic acid: a preventable signal for pneumococcal biofilm formation, colonization, and invasion of the host. J. Infect. Dis. 199:1497–1505 [DOI] [PubMed] [Google Scholar]
- 16. Brook I. 2011. Microbiology of sinusitis. Proc. Am. Thorac. Soc. 8:90–100 [DOI] [PubMed] [Google Scholar]
- 17. Del Beccaro MA, Mendelman PM, Inglis AF, Richardson MA, Duncan NO, Clausen CR, Stull TL. 1992. Bacteriology of acute otitis media: a new perspective. J. Pediatr. 120:81–84 [DOI] [PubMed] [Google Scholar]
- 18. Faden H, Stanievich J, Brodsky L, Bernstein J, Ogra PL. 1990. Changes in nasopharyngeal flora during otitis media of childhood. Pediatr. Infect. Dis. J. 9:623–626 [PubMed] [Google Scholar]
- 19. Trappetti C, Potter AJ, Paton AW, Oggioni MR, Paton JC. 2011. LuxS mediates iron-dependent biofilm formation, competence, and fratricide in Streptococcus pneumoniae. Infect. Immun. 79:4550–4558 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Vidal JE, Ludewick HP, Kunkel RM, Zahner D, Klugman KP. 2011. The LuxS-dependent quorum-sensing system regulates early biofilm formation by Streptococcus pneumoniae strain D39. Infect. Immun. 79:4050–4060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Enright MC, Spratt BG. 1998. A multilocus sequence typing scheme for Streptococcus pneumoniae: identification of clones associated with serious invasive disease. Microbiology 144:3049–3060 [DOI] [PubMed] [Google Scholar]
- 22. Kelly T, Dillard JP, Yother J. 1994. Effect of genetic switching of capsular type on virulence of Streptococcus pneumoniae. Infect. Immun. 62:1813–1819 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. McAllister LJ, Ogunniyi AD, Stroeher UH, Leach AJ, Paton JC. 2011. Contribution of serotype and genetic background to virulence of serotype 3 and serogroup 11 pneumococcal isolates. Infect. Immun. 79:4839–4849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Lacks S, Hotchkiss RD. 1960. A study of the genetic material determining an enzyme in Pneumococcus. Biochim. Biophys. Acta 39:508–518 [DOI] [PubMed] [Google Scholar]
- 25. Spratt BG. 1999. Multilocus sequence typing: molecular typing of bacterial pathogens in an era of rapid DNA sequencing and the internet. Curr. Opin. Microbiol. 2:312–316 [DOI] [PubMed] [Google Scholar]
- 26. Mahdi LK, Ogunniyi AD, LeMessurier KS, Paton JC. 2008. Pneumococcal virulence gene expression and host cytokine profiles during pathogenesis of invasive disease. Infect. Immun. 76:646–657 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. LeMessurier KS, Ogunniyi AD, Paton JC. 2006. Differential expression of key pneumococcal virulence genes in vivo. Microbiology 152:305–311 [DOI] [PubMed] [Google Scholar]
- 28. Trappetti C, Ogunniyi AD, Oggioni MR, Paton JC. 2011. Extracellular matrix formation enhances the ability of Streptococcus pneumoniae to cause invasive disease. PLoS One 6:e19844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Forbes ML, Horsey E, Hiller NL, Buchinsky FJ, Hayes JD, Compliment JM, Hillman T, Ezzo S, Shen K, Keefe R, Barbadora K, Post JC, Hu FZ, Ehrlich GD. 2008. Strain-specific virulence phenotypes of Streptococcus pneumoniae assessed using the Chinchilla laniger model of otitis media. PLoS One 3:e1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Hanage WP, Kaijalainen TH, Syrjanen RK, Auranen K, Leinonen M, Mäkelä PH, Spratt BG. 2005. Invasiveness of serotypes and clones of Streptococcus pneumoniae among children in Finland. Infect. Immun. 73:431–435 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. McDevitt CA, Ogunniyi AD, Valkov E, Lawrence MC, Kobe B, McEwan AG, Paton JC. 2011. A molecular mechanism for bacterial susceptibility to zinc. PLoS Pathog. 7:e1002357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Jahn AF. 2001. Bone physiology of the ear, p. 59–74 In Jahn AF, Santos-Sacchi J. (ed), Physiology of the ear, 2nd ed. Singular, Thomson Learning, San Diego, CA. [Google Scholar]
- 33. Blumenkrantz N, Asboe-Hansen G. 1973. New method for quantitative determination of uronic acids. Anal. Biochem. 54:484–489 [DOI] [PubMed] [Google Scholar]
- 34. Brown JS, Ogunniyi AD, Woodrow MC, Holden DW, Paton JC. 2001. Immunization with components of two iron uptake ABC transporters protects mice against systemic Streptococcus pneumoniae infection. Infect. Immun. 69:6702–6706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Mahdi LK, Wang H, Van der Hoek MB, Paton JC, Ogunniyi AD. 2012. Identification of a novel pneumococcal vaccine antigen preferentially expressed during meningitis in mice. J. Clin. Invest. 122:2208–2220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Ogunniyi AD, Mahdi LK, Trappetti C, et al. Verhoeven N, Mermans D, Van der Hoek MB, Plumptre CD, Paton JC. 2012. Identification of genes that contribute to pathogenesis of invasive pneumococcal disease by in vivo transcriptomic analysis. Infect. Immun. 80:3268–3278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Bättig P, Hathaway LJ, Hofer S, Muhlemann K. 2006. Serotype-specific invasiveness and colonization prevalence in Streptococcus pneumoniae correlate with the lag phase during in vitro growth. Microbes Infect. 8:2612–2617 [DOI] [PubMed] [Google Scholar]
- 38. Sjöström K, Spindler C, Ortqvist A, Kalin M, Sandgren A, Kühlmann-Berenzon S, Henriques-Normark B. 2006. Clonal and capsular types decide whether pneumococci will act as a primary or opportunistic pathogen. Clin. Infect. Dis. 42:451–459 [DOI] [PubMed] [Google Scholar]
- 39. Inverarity D, Lamb K, Diggle M, Robertson C, Greenhalgh D, Mitchell TJ, Smith A, Jefferies JM, Clarke SC, McMenamin J, Edwards GF. 2011. Death or survival from invasive pneumococcal disease in Scotland: associations with serogroups and multilocus sequence types. J. Med. Microbiol. 60:793–802 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Sandgren A, Albiger B, Orihuela CJ, Tuomanen E, Normark S, Henriques-Normark B. 2005. Virulence in mice of pneumococcal clonal types with known invasive disease potential in humans. J. Infect. Dis. 192:791–800 [DOI] [PubMed] [Google Scholar]