

Abstract
Objective
To examine the apoptotic effect of ent-11α-hydroxy-15-oxo-kaur-16-en-19-oic-acid (5F), a compound isolated from Pteris semipinnata L (PsL), in human lung cancer A549 cells.
Methods
A549 cells were treated with 5F (0–80 μg/ml) for different time periods. Cytotoxicity was examined using a MTT method. Cell cycle was examined using propidium iodide staining. Apoptosis was examined using Hoechst 33258 staining, enzyme-linked immunosorbent assay (ELISA) and caspase-3 activity analysis. Expression of representative apoptosis-related proteins was evaluated by Western blot analysis. Reactive oxygen species (ROS) level was measured using standard protocols. Potential interaction of 5F with cisplatin was also examined.
Results
5F inhibited the proliferation of A549 cells in a concentration- and time-dependent manner. 5F increased the accumulation of cells in sub-G1 phase and arrested the cells in the G2 phase. Exposure to 5F induced morphological changes and DNA fragmentation that are characteristic of apoptosis. The expression of p21 was increased. 5F exposure also increased Bax expression, release of cytochrome c and apoptosis inducing factor (AIF), and activation of caspase-3. 5F significantly sensitized the cells to cisplatin toxicity. Interestingly, treatment with 5F did not increase ROS, but reduced ROS production induced by cisplatin.
Conclusion
5F could inhibit the proliferation of A549 cells by arresting the cells in G2 phase and by inducing mitochondrial-mediated apoptosis.
Key words: Pteris semipinnata L, Lung cancer, G2 cell cycle arrest, Apoptosis, Reactive oxygen species
INTRODUCTION
Lung cancer is one of the most common malignancies. Approximately 85% of lung cancer is non-small cell lung cancer (NSCLC). NSCLC is relatively insensitive to radio- and chemo-therapy. Also, in 75% of the cases, the cancer has already metastasized upon diagnosis[1,2]. As a result, the 5-year overall survival rate is <15%[3]. More effective preventive and therapeutic approaches are urgently needed.
Ent-11-hydroxy-15-oxo-kaur-16-en-19-oic acid (5F) is an active compound in Pteris semipinnata L (PsL)[4-6]. Earlier studies from our laboratory demonstrated 5F could inhibit the in vitro growth of a number of tumors (e.g., colorectal cancer, gastric cancer, anaplastic thyroid carcinoma, liver cancer and laryngeal cancer) via a mitochondrial- mediated apoptotic mechanism[7-11]. We have also compared 5F with 7-hydroxystaurosporine (UCN01), which is currently being evaluated in clinical trials as an antitumor drug in the United State and Japan[12,13]. UCN01 is an inhibitor of protein kinase C, but has higher specificity than its parent compound staurosporine. Our previous study demonstrated equal cytotoxicity of 30 nmol/L 5F to 100 nmol/L UCN01 in colorectal cancer cells. Both UCN01 and 5F increased p38 mitogen-activated protein kinase (MAPK) and nitric oxide (NO) levels. Overexpression of either Bcl-2 or Bcl-xL prevented UCN01-induced cell death. Over- expression of Bcl-2, but not Bcl-xL protected the cells from 5F[7]. These results indicated overlapping yet different antitumor mechanisms for 5F and UCN01.
A recent study in mice from this laboratory showed that 5F is also effective against lung cancer induced by 4-methylnitrosamino-1-3-pyridyl- butanone (NNK) with minimal side effects[14]. The present study examined the potential antitumor action of 5F in a human NSCLC cell line, A549. Possible interaction between 5F and cisplatin was also examined.
MATERIALS AND METHODS
Antibodies and Reagents
Mouse-anti-human monoclonal primary antibodies for β-actin, Bax, cytochrome c, apoptosis inducing factor (AIF), p21 and horseradish peroxidase-conjugated goat-anti- mouse IgG antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). RPMI 1640, fetal bovine serum (FBS), 3-(4,5-dimethyl-thiazol- 2-yl)-2,5-diphenyltetrazoliumbromide (MTT), propidium iodide (PI) and Hoechst 33258 were purchased from Sigma-Aldrich (St. Louis, MO, USA). 5F was isolated from PsL as described previously[15]. All other reagents were purchased from Sigma-Aldrich unless otherwise mentioned.
Cell Culture
A549 cells were obtained from the American Type Culture Collection and cultured in RPMI 1640 medium containing 10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin. Cells were cultured at 37°C in a humidified 5% CO2 incubator. 5F was dissolved in propylene glycol (PG) and diluted with the culture medium immediately before use (final PG concentration ≤1.2%). In all experiments, the cells in RPMI 1640 medium plus PG only were used as the control.
Cell Viability Assay
Cells were seeded in a 96-well plate at a density of 5×103 cells/well. The total volume was adjusted to 200 μl with growth medium. At 24 h after the seeding, the cells were exposed to 5F (0–80 μg/ml), or a combination of 5F (10 μg/ml) and cisplatin (10 μg/ml). Cell viability was examined after 24, 48 or 72 h using a standard MTT method. Drug effect was expressed as percentage relative to the controls. Morphology of the cells was examined after 24 h exposure to 5F under an inverted phase contrast microscope.
Hoechst 33258/PI Staining Assay
Cells (1×105) were seeded onto cover slides and treated with 5F (0–80 μg/ml) for 24 h, then washed with ice-cold PBS and stained with Hoechst 33258 (10 μg/ml) at 37°C for 15 min. After removing free Hoechst 33258 with PBS, cells were stained with PI (20 μg/ml) at 4°C for 15 min, and washed again with PBS prior to observation under a fluorescence microscope (DMLB/ MPS-30/Q500IW, LEICA, Germany).
Cell Cycle Distribution
A549 cells were seeded at a density of 1×105 cells/well in a six-well plate. At 24 h after the seeding, the cells were treated with 5F (0–80 μg/ml) for 24 h at 37°C. Cells were fixed overnight with 70% ethanol at −20°C and stained with PI solution (100 μg/ml). Cell cycle distribution analysis was performed using a flow cytometer (EPICS XL, COULTER, USA).
Apoptosis Enzyme-linked Immunosorbent Assay (ELISA)
Cytoplasmic histone-associated-DNA-fragments (mono- and oligonucleosomes) were determined by Apoptosis Detection ELISAPLUS (Roche, Mannheim, Germany). Briefly, 1×104 cells/well were seeded in a 96-well plate and treated with various concentrations (0–80 μg/ml) of 5F for 24 h. After 24 h incubation, cells were lysed and centrifuged at 1,500 r/min. The cytoplasmic fraction (the supernatant) containing fragmented DNA was transferred to a streptavidin- coated microplate. A mixture of anti-histone-biotin and anti-DNA-peroxidase was added and incubated for 2 h at room temperature. After removal of unbound antibodies by washing, the amount of fragmented DNA of nucleosomes bound to anti-histone-biotin was evaluated by anti-DNA- peroxidase using peroxidase substrate at 405 nm.
Measurement of Caspase-3 Activity
Caspase-3 activity was measured using a spectrometric method as specified in the assay kit instruction (Caspase-3 Colorimetric Assay Kit, KeyGen Biotech, Nanjing, China). Cells were treated with 5F (0–80 μg/ml) for 24 h prior to harvest. Fifty microliter lysis buffer [containing 1% dithiothreitol (DTT)] was added to (3–5)×106 cells. The cells were incubated for 60 min on ice, and vortexed 3–4 times during the incubation. The lysate was centrifuged at 10,400 r/min for 1 min. Protein concentration in the supernatant was measured. Fifty microliter lysate supernantant (containing 100 μg protein) was mixed with 50 μl 2× reaction buffer (containing 1% DTT) and 50 μl caspase-3 substrate, and incubated for 4 h at 37°C in darkness. Absorbance was measured at 405 nm. Caspase-3 activity was expressed as: OD5F/ODcontrol.
Measurement of Reactive Oxygen Species (ROS)
Intracellular ROS was measured using a spectrometric method as specified by manufacturer’s instruction (ROS Assay Kit, GENMED Scientific, Shanghai, China). Cells (1×104 cells/well; 96-well plate) were treated with the test agent for 3 h prior to addition of Reagent A (20 μl/well) and subsequent incubation for 60 min at 37°C. The supernatant was aspirated prior to addition of Reagent B (100 μl/well). After aspiration of the supernatant, Reagent C (100 μl/well) was added. The culture plate was gently stirred to dissolve Reagent C. The absorbance was measured at 650 nm.
Protein Extraction
The total protein, cytosolic and mitochondrial protein fractions were isolated according to previous publication[16]. Briefly, whole cell extract was prepared using a lysis buffer containing: 50 mmol/L Tris, pH 7.4, 10 mmol/L EDTA, 1% Triton X-100, 26% urea and 1:25 protease inhibitor cocktail. For cellular fractionation, cells were resuspended in cytosolic lysis buffer (250 mmol/L sucrose, 20 mmol/L HEPES, 10 mmol/L KCl, 1.5 mmol/L MgCl2, 1 mmol/L EDTA, 1 mmol/L EGTA, 1 mmol/L DTT, 17 mg/ml phenylmethanesulfonyl fluoride (PMSF), 8 mg/ml aprotinin, and 2 mg/ml leupeptin, pH 7.4). After removing unlysed cells and neclei by centrifugation for 10 min at 2,800 r/min, the extract was centrifuged at 12,300 r/min for 15 min to derive the cytosolic (supernatant), and mitochondrial fractions (pellet).
Western Blot Analysis of Bax, Cytochrome c, AIF and P21
Proteins samples (20 μg protein for mitocondrial or cytosolic fractions for Bax, cytochrome c and AIF assays, and 50 μg of total protein from treated cells for P21 and β-actin assays) were subjected to electrophoresis using a 12% sodium dodecyl sulfate-polyacrylamide gel. Separated proteins were transferred to a nitrocellulose membrane, and blocked for 1 h using 10% skim milk in water. The membrane was incubated overnight with one of the following primary antibodies: Bax (1:200), cytochrome c (1:500), AIF (1:100), P21 (1:50) and β-actin (1:200). The membrane was then incubated with secondary antibodies for 3 h incubation at room temperature. Protein bands were visualized using chemiluminescent reagent.
Statistical Analysis
Data are presented as ‾x±s. The differences between the groups were examined with one-way analysis of variance (ANOVA) using the SPSS 13 software (SPSS Inc., Chicago, IL, USA). Statistical significance was defined as P<0.05.
RESULTS
Cell Viability
5F treatment inhibited the proliferation of A549 cells in a time- and dose-dependent manner (Figure 1A). 5F-treated cells exhibited a rounded and granulated morphology, and were detached from culture wall after 24 h exposure (Figure 1B). 5F also significantly sensitized A549 cells to cisplatin-induced cytotoxicity after 24, 48 and 72 h of treatment (Figure 1C).
Figure 1.

Cell viability. A: Effects of 5F on cell viability. *P<0.05 vs. blank control. B: Effect of 5F on cell morphology. ×100, under an inverted microscope. C: Synergistic cytotoxic effect of 5F with cisplatin. *P<0.05 compared with no 5F treatment.
Cell Cycle and Apoptosis
To explain the mechanism of cell growth inhibition by 5F concerning cell cycle change, we analyzed the effects of 5F treatment on cell cycle. 5F treatment increased the accumulation of G2 DNA in a concentration-dependent manner (P<0.05), indicating that 5F could result in G2 cell cycle arrest. More apoptotic cells (sub-G1 peak) were detected with increasing 5F concentration (P<0.05) (Figure 2).
Figure 2.

Effect of 5F on cell cycle and apoptosis, as measured by flow cytometric analysis. Cells were treated with 5F (0–80 μg/ml) for 24 h. A: Representative cell cycle histogram; B: Percentage of the cells in sub-G1 fraction and G2 phase. *P<0.05 vs. blank control.
The apoptosis was confirmed by three additional methods: DNA fragmentation, Hoechst 33258/PI staining and caspase-3 activity. DNA fragmentation was measured by ELISA, and was based on the detection of mono- and oligonucleosomes in the cytoplasmic fractions of cell lysates. As shown in Figure 3, 5F significantly induced DNA fragmentation in A549 cells after 24 h treatment. Hoechst 33258/PI staining revealed nuclear condensation, nuclear fragmentation and granular apoptotic bodies after 24-h 5F treatment (Figure 4). Caspase-3 plays a pivotal role in the terminal execution phase of apoptosis[17]. As shown in Figure 5, 5F significantly increased caspase-3 activity at 10–80 μg/ml, suggesting an apoptotic effect of 5F in A549 cells.
Figure 3.
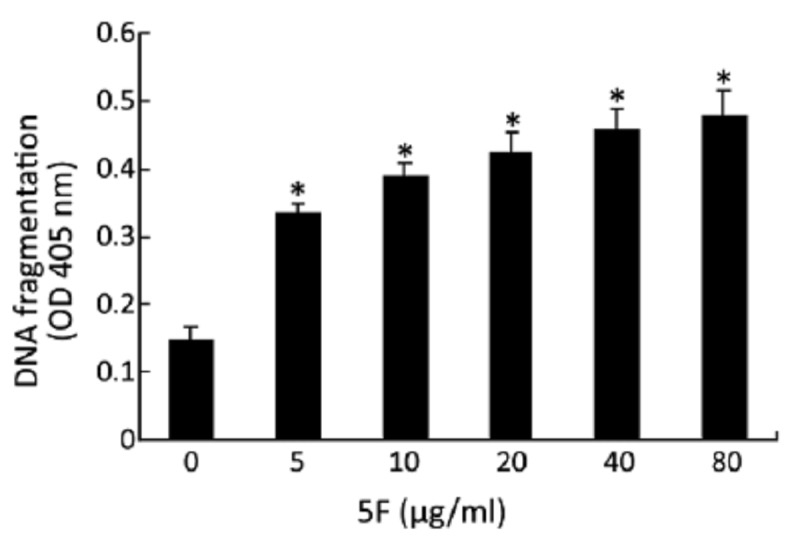
Effect of 5F on DNA fragmentation, as measured by ELISA. *P<0.05 vs. blank control.
Figure 4.

Morphology of apoptotic cells stained with Hoechst 33258/PI. Cells were treated with 5F (0–80 μg/ml) for 24 h prior to staining with Hoechst 33258/PI. ×200, under a fluorescent microscope.
Figure 5.
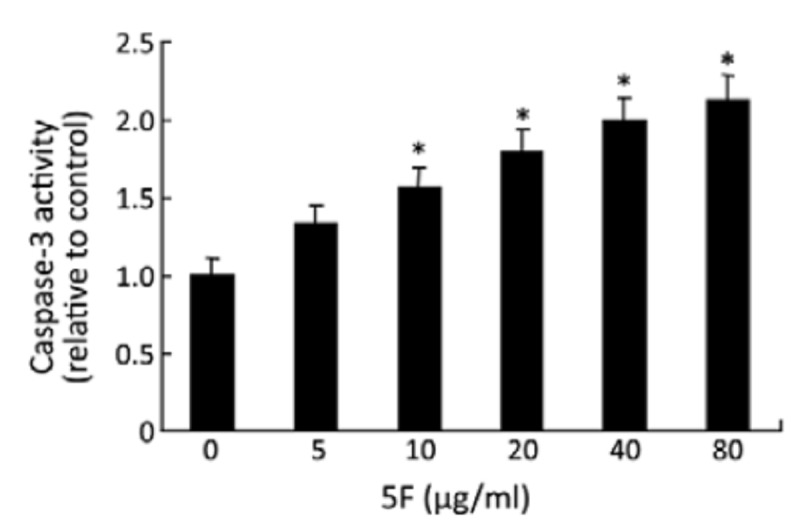
Effect of 5F on caspase-3 activity, as measured by a colorimetric assay. *P<0.05 vs. blank control.
Intracellular ROS Levels
ROS plays an important role in apoptosis in a variety of cells. To investigate the impact of 5F on ROS generation in A549 cells, we measured intracellular ROS level. As shown in Figure 6A, 5F (3-h exposure) significantly decreased ROS generation in A549 cells. It has been reported that cisplatin cytotoxicity is mediated by increased generation of ROS. Recently, several reports have demonstrated that cisplatin-induced cytotoxicity could be ameliorated by using antioxidants or oxygen radical scavengers[18-20]. Hence, we determined whether cisplatin stimulated intracellular ROS generation and whether 5F could affect ROS production. Cisplatin increased ROS production (Figure 6B). 5F treatment reduced ROS production in cisplatin-treated A549 cells.
Figure 6.

Effect on ROS generation. A: 5F, *P<0.05 vs. blank control; B: cisplatin, *P<0.05 compared with each other.
Expression of Proteins Involved in Cell Cycle Progression and Apoptosis
To understand the mechanism by which 5F caused the G2-phase cell cycle arrest, we examined the effects of 5F on the induction of p21, which regulates the G2-phase checkpoint[21-23]. 5F treatment induced p21 expression in a concentration-dependent manner (P<0.05) (Figure 7), suggesting that p21 might regulate the cell cycle arrest by 5F.
Figure 7.
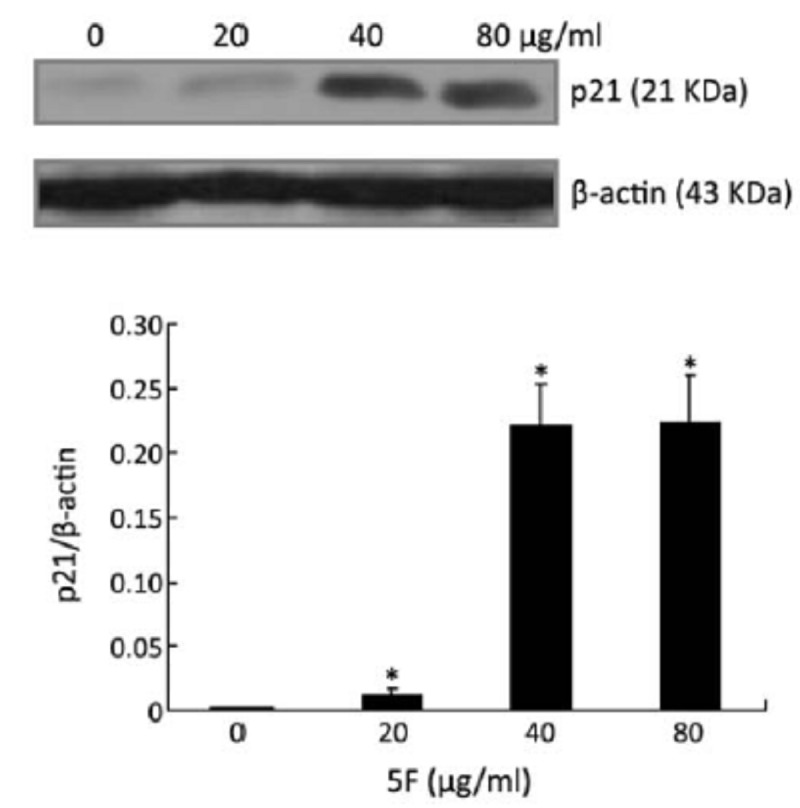
Effect of 5F on p21 expression, as measured by Western blot. *P<0.05 vs. blank control.
The disruption of mitochondrial membrane potential is one critical event in triggering mitochondrial- mediated apoptosis[24,25]. The mitochondrial dysfunction is induced by various cellular stimuli, causing the translocation of Bax from the cytosol to the mitochondria, which subsequently triggers the release of cytochrome c and AIF from the mitochondria into the cytosol, and finally induces apoptosis via either caspase-dependent or caspase- independent pathways. To further clarify the mechanisms of apoptosis in A549 cells triggered by 5F, we targeted apoptotic proteins in the mitochondrial pathway. 5F significantly increased Bax expression in mitochondria (Figure 8). 5F also dose-dependently upregulated the expression of cytochrome c and AIF proteins (P<0.05)
Figure 8.

Effects of 5F on Bax translocation to mitochondria, the release of cytochrome c and AIF from mitochondria into the cytosol. *P<0.05 vs. blank control.
DISCUSSION
Results from the present study demonstrated that 5F could inhibit the growth of A549 cells in a dose- and time-dependent manner. 5F also sensitized A549 cells to cisplatin-induced cytotoxicity. These data suggest that 5F may be a potential anti-lung tumor agent.
Dysregulated cell cycle progression and impaired apoptotic induction are important factors in cancer development[26]. Accordingly, inhibition of cell cycle regulation should be particularly useful in the treatment of cancer. 5F has been demonstrated to arrest cells at G2 in FRO cells (an anaplastic thyroid carcinoma cell line)[9]. Consistent with this report, the present study showed that 5F could induce G2-phase arrest and apoptosis in A549 cells. Regulation of the cell cycle at the G2-M checkpoint is complex and involves multiple molecular processes. p21 appears to be important since introduction of nonfunctional p21 or a p21 antisense oligonucleotide could diminish the G2-M arrest in a variety of cancer cells[27,28]. Apart from directly inhibiting cyclin-dependent kinase activity to promote cell cycle arrest, p21 interacts with proliferating cell nuclear antigens[29]. Our results demonstrated elevated p21 level in A549 cells in response to 5F exposure. Based on previous study, it is possible G2 arrest upon 5F treatment here is caused by P21.
The G2-phase arrest is associated with apoptosis in tumor cells treated with chemotherapeutic agents such as cisplatin[30]. In the present study, we demonstrated apoptosis-inducing effects of 5F in A549 cells using PI cell cycle analysis, DNA fragmentation (ELISA), Hoechst 33258/PI staining as well as activated caspase-3. Mitochondrial-mediated apoptotic pathway contributes to apoptosis of cancer cells induced by several anti-tumor agents[31-33]. In our experiment, 5F elevated Bax level in mitochondria, stimulated the release of cytochrome c and AIF from mitochondria into the cytosol, suggesting that 5F-induced apoptosis in A549 cells is also mediated by the mitochondrial pathway.
Excessive production of ROS induces apoptosis[34-36]. Cancer cells have high basal ROS levels, and are therefore more vulnerable to further ROS increase[37,38]. In our experiments, 5F not only decreased the basal level of ROS but also markedly reduced the cisplatin-induced generation of ROS in A549 cells. This finding is in agreement with a recent study that showed decreased ROS generation by 5F in HepG2 cells[39], but appears to be different from another earlier study that showed increased ROS production upon induction of apoptosis by 5F in gastric cells[8]. These inconsistent results may reflect different cancer cell types and ROS detection methods across the studies. In fact, increasing numbers of anticancer drugs have been found to enhance ROS generation.
Cisplatin is one of the most effective chemotherapeutic agents, but has prominent toxicities (e.g., ototoxicity and nephrotoxicity). These toxicities are believed to be related to its ability to induce ROS production[40,41]. Animal studies have indicated that antioxidants or oxygen radical scavengers could attenuate cisplatin toxicity[41,42]. Results from the current study confirmed that 5F possesses antioxidant property in addition to anticancer property. This is a major advantage that could limit its toxicity. The current study was conducted in a single cell line. Potential effects need to be investigated in other lung cancer cells.
In summary, 5F possibly induced G2 phase arrest and apoptotic cell death in A549 cells through the activation of p21 and the mitochondria-mediated apoptotic pathway. 5F sensitized A549 cells to cisplatin- induced cytotoxicity. These findings encourage further development of 5F as a chemotherapeutic agent for lung cancers. The ROS scavenging property of 5F offers a significant advantage over other cytotoxic agents, and forms the theoretical basis of combined use with other chemotherapeutic agents.
REFERENCES
- 1.Yang P, Allen MS, Aubry MC, et al. Clinical features of 5,628 primary lung cancer patients: experience at Mayo Clinic from 1997 to 2003. Chest 2005; 128:452-62 [DOI] [PubMed] [Google Scholar]
- 2.Govindan R, Page N, Morgensztern D, et al. Changing epidemiology of small-cell lung cancer in the United States over the last 30 years: analysis of the surveillance, epidemiologic, and end results database. J Clin Oncol 2006; 24:4539-44 [DOI] [PubMed] [Google Scholar]
- 3.Erridge SC, Møller H, Price A, et al. International comparisons of survival from lung cancer: pitfalls and warnings. Nat Clin Pract Oncol 2007; 4:570-7 [DOI] [PubMed] [Google Scholar]
- 4.Li J, Liang N, Mo L, et al. Comparison of the cytotoxicity of five constituents from Pterissemipinnata L. in vitro and the analysis of their structure-activity relationships. Acta Pharmaceutica Sinica 1998; 33:641-4 [PubMed] [Google Scholar]
- 5.Lu YN, Chen G, Wu KF, et al. Determination of 5F in Pterissemipinnata L injection by HPLC. Xin Yao Cai (in Chinese)2008; 23:2048-58 [Google Scholar]
- 6.Gong XL, Chen ZH, Lu YN, et al. Purification of the antitumor compound 5F from Pterissemipinnata L. Zhong Cheng Yao (in Chinese)2008; 30:1286-9 [Google Scholar]
- 7.Chen GG, Liang NC, Lee JF, et al. Over-expression of Bcl-2 against Pterissemipinnata L-induced apoptosis of human colon cancer cells via a NF-kappa B-related pathway. Apoptosis 2004; 9:619-27 [DOI] [PubMed] [Google Scholar]
- 8.Liu Z, Ng EK, Liang NC, et al. Cell death induced by Pterissemipinnata L. is associated with p53 and oxidant stress in gastric cancer cells. FEBS Lett 2005; 579:1477-87 [DOI] [PubMed] [Google Scholar]
- 9.Liu ZM, Chen GG, Vlantis AC, et al. Cell death induced by ent-11alpha-hydroxy-15-oxo-kaur-16-en-19-oic-acid in anaplastic thyroid carcinoma cells is via a mitochondrial-mediated pathway. Apoptosis 2005; 10:1345-56 [DOI] [PubMed] [Google Scholar]
- 10.Li L, Liu Y, Lu YN, et al. Apoptosis effect and mechanism of 5F from Pterissemipinnata on HepG2 cells. Zhong Yao Cai (in Chinese)2010; 33: 77-80 [PubMed] [Google Scholar]
- 11.Vlantis AC, Lo CS, Chen GG, et al. Induction of laryngeal cancer cell death by Ent-11-hydroxy-15-oxo-kaur-16-en-19-oic acid. Head Neck 2010; 32:1506- 18 [DOI] [PubMed] [Google Scholar]
- 12.Fuse E, Kuwabara T, Sparreboom A, et al. Review of UCN-01 development: a lesson in the importance of clinical pharmacology. J Clin Pharmacol 2005; 45:394-403 [DOI] [PubMed] [Google Scholar]
- 13.Edelman MJ, Bauer KS, Jr, Wu S, et al. Phase I and pharmacokinetic study of 7-hydroxystaurosporine and carboplatin in advanced solid tumors. Clin Cancer Res 2007; 13:2667-74 [DOI] [PubMed] [Google Scholar]
- 14.Li MY, Leung J, Kong AW, et al. Anticancer efficacy of 5F in NNK-induced lung cancer development of A/J mice and human lung cancer cells. J Mol Med 2010; 88:1265-76 [DOI] [PubMed] [Google Scholar]
- 15.Deng YF, Liang NC. Study on extraction and separation of diterpenoids from Pterissemipinnata. ZhongGuo Yao Li XueZaZhi (in Chinese)2004; 39:42-4 [Google Scholar]
- 16.Tian Z, Shen J, Moseman AP, et al. Dulxanthone A induces cell cycle arrest and apoptosis via up-regulation of p53 through mitochondrial pathway in HepG2 cells. Int J Cancer 2008; 122:31-8 [DOI] [PubMed] [Google Scholar]
- 17.Vaux DL, Korsmeyer SJ. Cell death in development. Cell 1999; 96: 245-54 [DOI] [PubMed] [Google Scholar]
- 18.Jiang M, Wei Q, Pabla N, et al. Effects of hydroxyl radical scavenging on cisplatin-induced p53 activation, tubular cell apoptosis and nephrotoxicity. Biochem Pharmacol 2007; 73:1499-510 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kim YH, Kim YW, Oh YJ, et al. Protective effect of the ethanol extract of the roots of Brassica rapa on cisplatin-induced nephrotoxicity in LLC-PK1 cells and rats. Biol Pharm Bull 2006; 29:2436-41 [DOI] [PubMed] [Google Scholar]
- 20.Satoh M, Kashihara N, Fujimoto S, et al. A novel free radical scavenger, edarabone, protects against cisplatin-induced acute renal damage in vitro and in vivo. J Pharmacol Exp Ther 2003; 305:1183-90 [DOI] [PubMed] [Google Scholar]
- 21.Nigg EA. Cyclin-dependent protein kinases: key regulators of the eukaryotic cell cycle. Bioessays 1995; 17:471-80 [DOI] [PubMed] [Google Scholar]
- 22.Gautier J, Solomon MJ, Booher RN, et al. cdc25 is a specific tyrosine phosphatase that directly activates p34cdc2. Cell 1991; 67:197-211 [DOI] [PubMed] [Google Scholar]
- 23.Sherr CJ. Cancer cell cycles. Science 1996; 274:1672-7 [DOI] [PubMed] [Google Scholar]
- 24.Kroemer G.Mitochondrial control of apoptosis: an introduction. Biochem Biophys Res Commun 2003; 304:433-5 [DOI] [PubMed] [Google Scholar]
- 25.Green DR, Reed JC. Mitochondria and apoptosis. Science 1998; 281:1309-12 [DOI] [PubMed] [Google Scholar]
- 26.Senderowicz AM. Targeting cell cycle and apoptosis for the treatment of human malignancies. Curr Opin Cell Biol 2004; 16:670-8 [DOI] [PubMed] [Google Scholar]
- 27.De Siervi A, Marinissen M, Diggs J, et al. Transcriptional activation of p21(waf1/cip1) by alkylphospholipids: role of the mitogen-activated protein kinase pathway in the transactivation of the human p21(waf1/cip1) promoter by Sp1. Cancer Res 2004; 64:743-50 [DOI] [PubMed] [Google Scholar]
- 28.Rigberg DA, Blinman TA, Kim FS, et al. Antisense blockade of p21/WAF1 decreases radiation-induced G2 arrest in esophageal squamous cell carcinoma. J Surg Res 1999; 81:6-10 [DOI] [PubMed] [Google Scholar]
- 29.Ando T, Kawabe T, Ohara H, et al. Involvement of the interaction between p21 and proliferating cell nuclear antigen for the maintenance of G2/M arrest after DNA damage. J Biol Chem 2001; 276:42971-7 [DOI] [PubMed] [Google Scholar]
- 30.Chu G.Cellular responses to cisplatin. The roles of DNA-binding proteins and DNA repair. J Biol Chem 1994; 269:787-90 [PubMed] [Google Scholar]
- 31.Malugin A, Kopecková P, Kopecek J.HPMA copolymer-bound doxorubicin induces apoptosis in ovarian carcinoma cells by the disruption of mitochondrial function. Mol Pharm 2006; 3:351-61 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yim EK, Lee KH, Bae JS, et al. Proteomic analysis of antiproliferative effects by treatment of 5-fluorouracil in cervical cancer cells. DNA Cell Biol 2004; 23:769-76 [DOI] [PubMed] [Google Scholar]
- 33.Boulikas T, Vougiouka M.Cisplatin and platinum drugs at the molecular level. Oncol Rep 2003; 10:1663-82 [PubMed] [Google Scholar]
- 34.Sastre J, Pallardó FV, Viña J. Mitochondrial oxidative stress plays a key role in aging and apoptosis. IUBMB Life 2000; 49:427-35 [DOI] [PubMed] [Google Scholar]
- 35.Carmody RJ, Cotter TG. Signalling apoptosis: a radical approach. Redox Rep 2001; 6:77-90 [DOI] [PubMed] [Google Scholar]
- 36.Pelicano H, Feng L, Zhou Y, et al. Inhibition of mitochondrial respiration: a novel strategy to enhance drug-induced apoptosis in human leukemia cells by a reactive oxygen species-mediated mechanism. J Biol Chem 2003; 278:37832-9 [DOI] [PubMed] [Google Scholar]
- 37.Laurent A, Nicco C, Chéreau C, et al. Controlling tumor growth by modulating endogenous production of reactive oxygen species. Cancer Res 2005; 65:948-56 [PubMed] [Google Scholar]
- 38.Gupta A, Rosenberger SF, Bowden GT. Increased ROS levels contribute to elevated transcription factor and MAP kinase activities in malignantly progressed mouse keratinocyte cell lines. Carcinogenesis 1999; 20:2063-73 [DOI] [PubMed] [Google Scholar]
- 39.Li L, Wu KF, Liu Y, et al. Ros is not involved in induction of cell death by induction of cancer cell death by Ent-11-hydroxy-15-oxo-kaur-16-en-19-oic acid in HepG2 cells. Zhong Guo Zhong Yao Za Zhi (in Chinese)2010; 35:1287-91 [DOI] [PubMed] [Google Scholar]
- 40.Chirino YI, Pedraza-Chaverri J. Role of oxidative and nitrosative stress in cisplatin-induced nephrotoxicity. Exp Toxicol Pathol 2009; 61:223-42 [DOI] [PubMed] [Google Scholar]
- 41.Rybak LP, Whitworth CA, Mukherjea D, et al. Mechanisms of cisplatin-induced ototoxicity and prevention. Hear Res 2007; 226:157-67 [DOI] [PubMed] [Google Scholar]
- 42.van den Berg JH, Beijnen JH, Balm AJ, et al. Future opportunities in preventing cisplatin induced ototoxicity. Cancer Treat Rev 2006; 32: 390-7 [DOI] [PubMed] [Google Scholar]