

Abstract
We have identified a CD8+CXCR5+ T cell that prevents the development of oviduct dilation following C. muridarum genital infection. Phenotypic studies show that CD8+CXCR5+ cells express markers of T regulatory cells (FoxP3, CD25, and GITR) but do not express a necessary component of cytotoxic cells (perforin). Cxcr5−/− mice have significantly lower numbers of CD8+ cells and lack the CD8+CXCR5+ population while the total number of CD4+ cells is equivalent between mouse strains. The transfer of CD8+ splenocytes from WT mice reduces the oviduct dilation seen in Cxcr5−/− mice following C. muridarum infection. Future studies will investigate the mechanism by which this cell type regulates genital tract pathology.
1. Introduction
Pelvic inflammatory disease (PID) is defined as inflammation of the uterus and/or fallopian tubes and is induced by a number of organisms following sexual transmission. Chlamydia trachomatis is the most common reportable sexually transmitted infection (STI) and is responsible for >1 million cases in the US and approximately 92 million cases worldwide each year [1, 2]. Genital infection can lead to immune-mediated damage of the female reproductive organs and serious reproductive disability, including PID that can result in chronic pelvic pain, ectopic pregnancy, and infertility [3]. The risk of developing infertility increases by 40%–70% following reinfection [4]. The reinfection rate is approximately 13% and occurs within 6 months [5]. Delivery of treatments designed to reduce the local inflammation and prevent fibrotic disease to infected individuals may be a viable approach for further reducing PID and the costs associated with its treatment.
Regulatory T cells (Tregs) are comprised of multiple subsets of T cells that suppress other T cells from engaging in detrimental immune responses [6]. Tregs are broadly categorized as natural or inducible. Natural Tregs induce tolerance, delete autoreactive T cells, and dampen inflammation during an autoimmune reaction [7–13]. Inducible Tregs arise during infections in response to the infectious process to restore the homeostatic environment. In some cases, Tregs can be actively induced by the pathogen and promote pathogen survival by preventing elimination [14]. Tregs have also been shown to protect mucosal surfaces of the intestine from inflammation [15]. The linage can be phenotypically identified by the Foxp3 transcription factor [16]. The most widely studied subset is phenotypically defined as CD4+CD25+FoxP3+. This subset has been shown to indirectly prolong microbial growth by interfering with the priming of naive or unstimulated T cells [17].
CD8 cells also have suppressive activity and have been identified with and without FoxP expression to include the following: CD8+CD25+FoxP3+, CD8+CD45RClowFoxP3+, CD8+CD28−FoxP3−, CD8+CD122+FoxP3−, and CD8αα +FoxP3− as in [18, 19]. Recently CD8+ Tregs have emerged as preventing the pathogenic cascade leading to the onset of fibrotic disease in primary biliary cirrhosis and systemic sclerosis [20]. In this study, we identified a CD8+ cell that coexpresses CXCR5 and regulates genital tract pathology in vivo following infection with C. muridarum.
2. Materials and Methods
2.1. Animals, Chlamydia, and Infection of Mice
A breeding colony was established with Cxcr5−/− mice (8 generations in C57BL/6) obtained from Martin Lipp, Delbrück-Center for Molecular Medicine, Berlin, Germany. Female C57BL/6 mice, 5-6 weeks old (Harlan Sprague-Dawley, Indianapolis, IN, USA) were housed according to American Association of Accreditation of Laboratory Animal Care guidelines. Animal experimental procedures were approved by the UCLA Institutional Animal Care and Use Committee. Chlamydia muridarum was grown on confluent McCoy cell monolayers, purified on Renografin gradients and stored at −80°C in sucrose-phosphate-glutamine buffer (SPG) as previously described [21]. Mice were hormonally synchronized by subcutaneous injection with 2.5 mg of medroxyprogesterone acetate (Depo Provera, Upjohn, Kalamazoo, MI, USA) in 100 μL saline 7 days prior to a vaginal challenge with 1.5 × 105 infection forming units (IFUs) of C. muridarum under anesthetization. Depo Provera drives mice into a state of anestrous and eliminates the variability in the rate and severity of infection due to the estrus cycle. Infection was monitored by measuring IFUs from cervical-vaginal swabs (Dacroswab Type 1, Spectrum Laboratories, Rancho Dominguez, CA, USA) as described [21].
2.2. Histology
The genital tracts (GTs) were removed and, fixed in 10% formalin overnight, followed by 70% ethanol. Tissues were embedded en bloc in paraffin, sectioned (5 mm), and stained with hematoxylin and eosin. Tissue blocks were cut transversally from the ovary, and sections were collected at the beginning of the transitional region between ovary and oviduct. A veterinarian pathologist scored 2 sections from the right and left oviducts of each mouse for luminal dilation; 0 = luminal oviduct size of naïve mice, 1+ = mildly increased luminal oviduct size, 2+ = moderately increased luminal oviduct size, 3+ = severely increased luminal oviduct size, and 4+ = severely increased luminal oviduct size in greater than 75% of oviducts.
2.3. Lympholyte Isolation and FACS Identification
Spleen (Spl) and mesenteric lymph nodes (MLN) were harvested from individual mice. Single cell suspensions were attained by dissociating cells within the organs. Lymphocytes were incubated in RPMI 1640 in the presence of PMA and ionomycin. Brefeldin A (Sigma-Aldrich, St. Louis, MO, USA) was added 4 hr before the end of the culture period. The cells were then stained with fluorochrome-labeled antibodies against CD3 (clone 145-2C11), CD4 (clone GK1.5), CD8β (clone eBioH35-17.2), CXCR5 (clone 2G8), CD25 (clone PC61.5), GITR (DTA-1), CD122 (clone TM-beta 1), CD127 (clone A7R34), TCRβ (clone H57-597), TCRγδ (clone eBioGL3), α-GalCer-CD1d-tetramer a gift from Mitchell Kronenberg [22], and irrelevant control abs (eBioscience, San Diego, CA or BD Biosciences, San Jose, CA, USA) for 20 min on ice. After being washed, the cells were incubated with Cytofix/Cytoperm (eBioscience) for 1 hr and stained with fluorochrome-conjugated FoxP3 (clone FJK-16s) for 20 min on ice, washed again, and resuspended in Cell Fix solution (eBioscience). Flow cytometry was performed on a fluorescence activated cell sorting analyzer equipped with a 488 nm argon laser and CellQuest software (FACScan; BD Biosciences). The instrument was calibrated with beads (CaliBRITE; Becton Dickinson), using AutoCOMP software. Dead cells were excluded on the basis of forward angle and 90° light scatter, and 10,000 gated cells were analyzed for each sample.
2.4. Cell Sorting and Adoptive Transfer
Lymphocytes were isolated from the spleens of infected and uninfected mice as described above. Cells were stained for CD3 and CD8 and sorted for the CD3+CD8+ population using FACSAria cell sorter (BD Bioscience) in the UCLA FACS Core Labaratory. The purity of the sorted population was 99%. Cells were resuspended in saline and 1 × 106 cells were intravenously injected into the recipient through the tail vein. At the time of transfer, the mice were infected with C. muridarum as described above. These mice were also synchronized with medroxyprogesterone acetate 7 days prior to infection as described above.
2.5. Statistics
The percentage of CD4 and CD8 cells, oviduct luminal dilation scores, and IFU counts from WT and Cxcr5−/− mice were compared using Student's t-test, Kruskal-Wallis, and ANOVA tests, respectfully, with software from GraphPad Software, Inc. (La Jolla, CA, USA). Groups were considered statistically different at P values of <0.05.
3. Results and Discussion
3.1. Phenotypic Characterization of CD8+CXCR5+ Cells
We identified a population of CD8+ cells that expressed CXCR5 and comprised 2%-3% of CD8 T cells in the MLN (Figure 1(a)) and spleen (data not shown). The percentage of CD8+CXCR5+ cells in the MLN and spleen did not change in a naive mice or following C. muridarum genital infection (data not shown). Phenotypic analysis found that these cells were memory cells by high expression of CD44. Analysis of cytotoxic markers, granzyme B and perforin, showed that these cells did not express perforin. Although granzyme B was expressed, these cells are not considered cytotoxic since perforin is required for entry of granzymes into the cell cytoplasm as shown in mice deficient in perforin [23–25]. In addition, cytotoxic memory cells express high levels of both granzyme B and perforin (Figure 1(b)) [26]. Examination of markers of Tregs showed a subpopulation of CD8+CXCR5+ cells that expressed CD25, Foxp3 and GITR (glucocorticoid-induced tumor necrosis factor receptor family related gene) upon stimulation from mice during a genital infection with C. muridarum (Figure 1(c)). Analysis of CD122 or CD127 was negative (data not shown). Interestingly, induction of this population could not be found on stimulated cells from naive mice (Figure 1(d)). This finding indicates that a subpopulation of CD8+CXCR5+ cells express phenotypic markers associated with Tregs and not cytotoxic cells.
Figure 1.
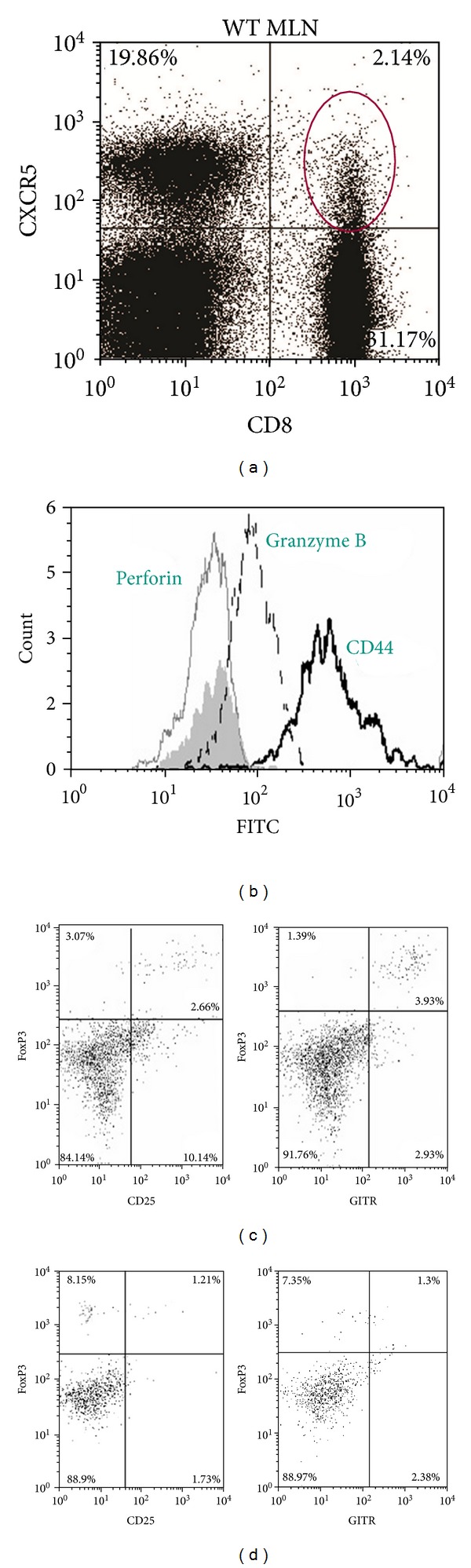
CD8+CXCR5+ cells express the phenotype of Tregs. MLNs were isolated from C57BL/6 WT mice 7 days after genital infection with C. muridarum and stained with CD3, CD8, CXCR5, and various markers of cytotoxic and regulatory T cells. (a) The dotplot shows the percentage of CD8 and CXCR5 on CD3+ cells. (b) Histogram of CD3+CD8+CXCR5+ cells showing the individual expression of perforin (gray line), granzyme B (dash line), and CD44 (black line). Irrelevant control antibody stained cells appear as a shaded histogram. (c) Dotplot showing the percent of CD3+CD8+CXCR5+ cells expressing of FoxP3 and CD25 or FoxP3 and GITR following stimulation of spleen cells from mice during genital infection with C. muridarum. (d) Dotplot showing the percent of CD3+CD8+CXCR5+ cells expressing of FoxP3 and CD25 or FoxP3 and GITR following stimulation of spleen cells from naive mice.
3.2. CD8+CXCR5+ Population Is Absent in Cxcr5−/− Mice
To further characterize the phenotype and examine the function of CD8+CXCR5+ cells, we evaluated the CD8 population in Cxcr5−/− mice. As expected, we did not find any CD8+CXCR5+ cells in mice lacking CXCR5 (Figure 2(a)). Cxcr5−/− mice had slightly less CD3+ T cells (data not shown) but had similar numbers of CD4 cells. In addition, the mice had significantly less CD8 cells compared to WT mice (Figure 2(b)). Further analysis showed that these cells were CD8 T cells by expression of the TCR and CD8 β chain and not a γδ T cell or NKT cell that could express CD8 (Figure 2(c)).
Figure 2.
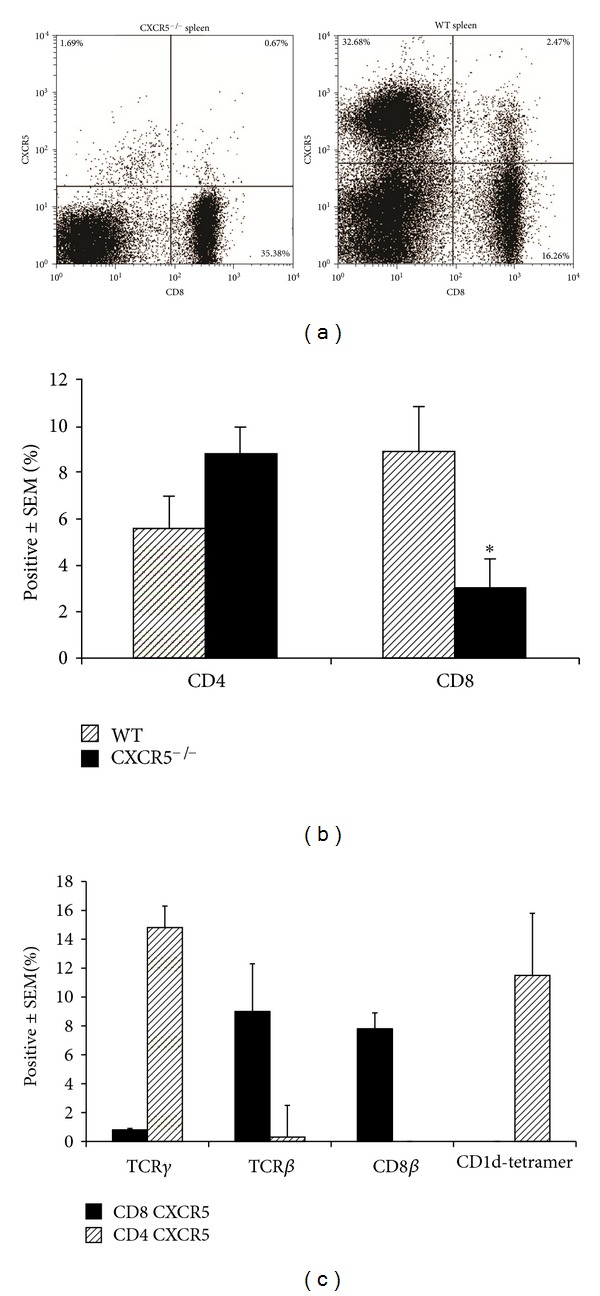
Cx cr5−/− mice lack CD8+CXCR5+ cells. Spleen cells were isolated from C57BL/6 WT mice 7 days after genital infection with C. muridarum and stained with CD3, CD4 or CD8, CXCR5, and markers of T cells; TCRγδ, TCRβ, CD8β or NKT cells; CD1d-tetramer. (a) The dotplot shows the percentage of CD8 and CXCR5 of CD3+ cells from Cxcr5−/− and WT mice. (b) The percent of CD4 or CD8 populations from groups of 6 mice were compared between Cxcr5−/− and WT mice by Student's t-test. The ∗ indicates P < 0.01. (c) The percentage of CD4 or CD8 cells expressing markers of T cells; TCRγ, TCRβ, CD8β or NKT cells; CD1d-tetramer.
3.3. Adoptive Transfer of CD8+ Cells Reversed Oviduct Dilation after C. muridarum Infection in Cxcr5−/− Mice
WT and Cxcr5−/− mice were intravaginally infected with C. muridarum and splenic lymphocytes were purified 7 days after infection. The lymphocytes or purified CD8+ cells were adoptively transferred into WT or Cxcr5−/− recipients. The genital tracts were harvested seven weeks after infection, hematoxylin and eosin stained, and scored by a veterinarian pathologist. We found that the adoptive transfer of WT lymphocytes into WT recipients had significantly reduced oviduct dilation compared to Cxcr5−/− mice given Cxcr5−/− lymphocytes. Further, Cxcr5−/− mice given purified CD8+ cells from WT mice also showed significantly reduced oviduct dilation (Figure 3(a)). It is possible that the transfer of CD8 effector cells could account for a reduction in oviduct dilation by perhaps reducing bacterial burden. However, there was no difference in bacterial burden in the GT on days 9 following infection between Cxcr5−/− mice given WT-CD8 cells or Cxcr5−/− lymphocytes or WT mice given WT lymphocytes (Figure 3(b)). Taken together, these data strongly suggest that CD8+CXCR5+/+ (WT mice) prevent or interfere with the development of oviduct dilation following C. muridarum genital infection.
Figure 3.
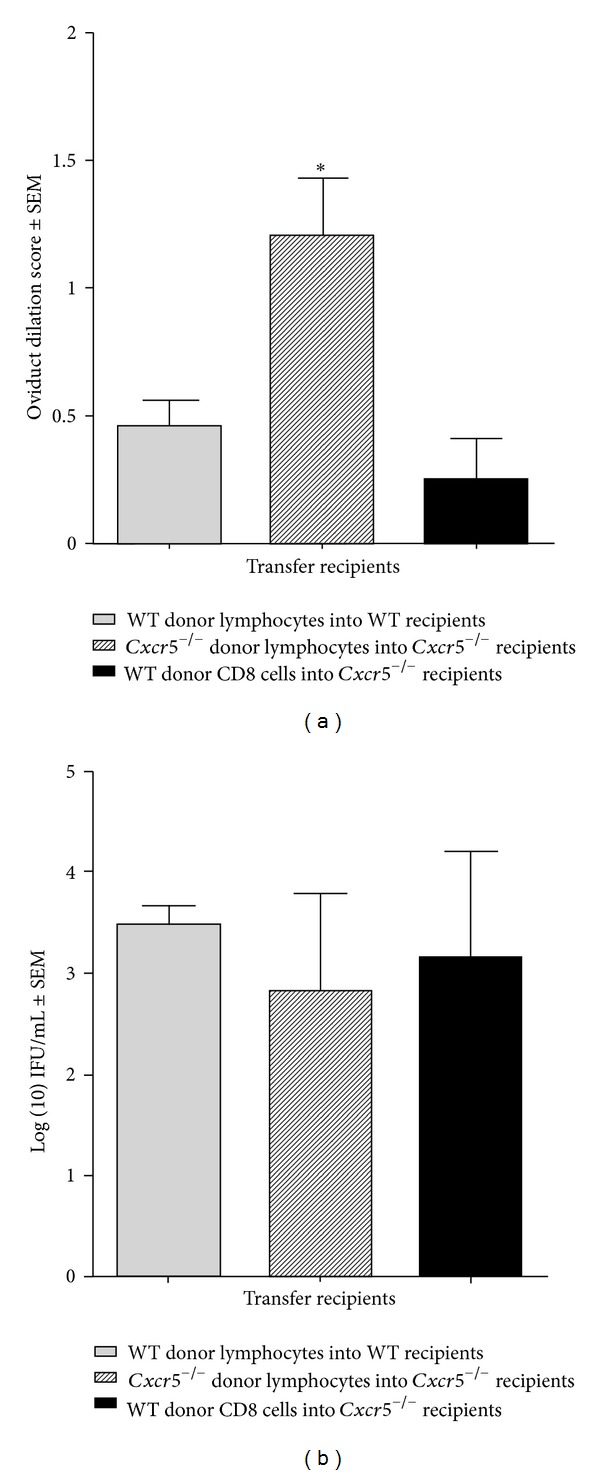
Adoptive transfer of CD8+ cells reversed oviduct dilation late after C. muridarum infection in Cxcr5−/− mice. (a) Donor Cxcr5−/− and WT mice were infected with C. muridarum and the spleens were harvested 7 days later and lymphocytes were obtained by dissociating cells within the organs. Donor C57BL/6 WT mice were infected with C. muridarum and the spleens harvested 7 days after infection. CD8+ cells were purified by FACS sorting for expression of CD8. Cxcr5−/− lymphocytes and WT-CD8+ were adoptively transferred (1 × 106) into Cxcr5−/− mice as indicated. Genital tracts were harvested 7 weeks later and hemetoxylin and eosin staining was performed and scored. Oviducts were scored individually and were compared between recipient groups using Kruskal-Wallis test. Transfer groups are, WT donor into WT recipient; n = 6 mice (12 oviducts), Cxcr5−/− donor into Cxcr5−/− recipient, n = 6 mice (12 oviducts), and WT-CD8 donor into Cxcr5−/− recipient, n = 2 mice (4 oviducts). The bars indicate the mean ± SEM and *P = 0.02. (b) Recipient mice were vaginally swabbed 9 days after infection and all groups were compared by one-way ANOVA. There were no differences between groups. The bars indicate the mean IFU ± SEM.
4. Conclusions
Our study describes the identification of CD8+CXCR5+ cells that possess the ability to regulate oviduct dilation that occurs following the immune response to C. muridarum genital infection. This study highlights the finding that the CD8+ population is comprised of multiple subsets with differing function. Murthy et al., showed that CD8 cells secreting TNFα cause oviduct dilation and hydrosalpinx [27]. Individual differences in PID or infertility may be influenced by the proportions of CD8+ cell subsets. The mechanism whereby regulation of genital tract pathology occurs is not known. However, a small percentage of CD8+CXCR5+ cells express markers of Tregs; FoxP3, CD25, and GITR but not perforin, a marker of cytotoxic cells, and suggest that these cells function as T regulatory cells.
Our data show that adoptive transfer of CD8+CXCR5+ cells prevent oviduct dilation following genital infection with C. muridarum and suggests that this cell type is necessary for preventing oviduct dilation following genital infection. However, CD8+ cells with suppressive activity require IL-10+ APC for expansion [28]. IL-10+ APC have been shown to reduce the number of antichlamydial Th1 cells that develop and this results in a prolonged infection [29]. Others have suggested that there are a number of immune responses operating in different phases of the immune response and this could possibly explain the conflicting role of IL-10+ APC during C. muridarum infection.
A population of CD8 cells that also expresses CXCR5 has been reported but their function has not been identified [30]. There are a number of subsets of CD8+ Tregs that have been identified and include the following: CD8+CD25+FoxP3+, CD8+CD45RClowFoxP3+, CD8+CD28−FoxP3−, CD8+CD122+FoxP3−, and CD8ααFoxP3− [18]. We found that a small percentage of CD8+CXCR5+ cells expressed FoxP3 upon antigen stimulation but did not express CD122 or CD8αα. The mechanism of suppression of the CD8αα subset has been reported. This subset is restricted to nonclassical MHC class Ib molecules, Qa-1, and has been found to suppress autoantibody formation and development of systemic lupus erythematosus (SLE-) like disease through inhibition of TFH [31]. Although CXCR5 expression does not prevent entry into germinal centers, the Cxcr5−/− mice used in our study do not have TFH cells or germinal centers and have reduced levels of immunoglobulins [32, 33]. Likewise we also find a reduction in antichlamydial IgM and IgG levels (data not shown). Additionally, regulation by the CD8αα subset depended on perforin and perforin was not found on activated CD8+CXCR5+ cells in this study confirming that CD8+CXCR5+ identified in this study are not the CD8αα expressing cell described previously.
T regulatory cells primarily effect T cells and/or dendritic cells and have four mechanisms by which they suppress immune responses. T regulatory cells secrete inhibitory cytokines such as IL-10 and TGFβ, induce apoptosis of target cells using the granzyme-perforin pathway, deprive effector T cells of IL-2 which leads to apoptosis of effector T cells, and inhibit dendritic cell function [19]. Our study has found that the granzyme-perforin pathway is unlikely to be used by CD8+CXCR5+ based on a lack of expression of the effector molecule, perforin. It would appear unlikely that suppressive cytokines are secreted since the resolution of infection is similar between Cxcr5−/− and WT mice and IL-10 secretion has been shown to prolong the course of infection [29]. Future studies will determine whether CD8+CXCR5+ function as T regulatory cells and act directly on T cells or on another cell type such as dendritic cells.
Conflict of Interests
The authors do not have a commercial or other association that might pose a conflict of interests.
Acknowledgments
The authors thank the past and present members of the laboratory for their contributions and to Nora Rozengurt for scoring the genital tract sections. This work was supported by the National Institutes of Health, AI026328 (K. A. Kelly) and a sponsored research award from Abraxis Bioscience, Inc. (K. A. Kelly). This work was presented in part at the 6th Meeting of the European Society for Chlamydia Research, Aarhus, Denmark, July 1–4, 2008.
References
- 1.World Health Organization DoHA. Global Prevalence and Incidence of Selected Curable Sexually Transmitted Infections. 2001. [PubMed]
- 2.US Department of Health and Human Services DoSP. Sexually Transmitted Disease Surveillance, 2007. Atlanta, Ga, USA: Centers for Disease Control and Prevention; 2007. [Google Scholar]
- 3.US Department of Health and Human Services DoSP, 1999. Sexually Transmitted Disease Surveillance. Atlanta, Ga, USA: (CDC) CfDCaP; 2000. [Google Scholar]
- 4.Westrom L, Joesoef R, Reynolds G, Hagdu A, Thompson SE. Pelvic inflammatory disease and fertility: a cohort study of 1,844 women with laparoscopically verified disease and 657 control women with normal laparoscopic results. Sexually Transmitted Diseases. 1992;19(4):185–192. [PubMed] [Google Scholar]
- 5.Burstein GR, Gaydos CA, Diener-West M, Howell MR, Zenilman JM, Quinn TC. Incident Chlamydia trachomatis infections among inner-city adolescent females. Journal of the American Medical Association. 1998;280(6):521–526. doi: 10.1001/jama.280.6.521. [DOI] [PubMed] [Google Scholar]
- 6.Corthay A. How do regulatory T cells work? Scandinavian Journal of Immunology. 2009;70(4):326–336. doi: 10.1111/j.1365-3083.2009.02308.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Coutinho A, Caramalho I, Seixas E, Demengeot J. Thymic commitment of regulatory T cells is a pathway of TCR-dependent selection that isolates repertoires undergoing positive or negative selection. Current Topics in Microbiology and Immunology. 2005;293:43–71. doi: 10.1007/3-540-27702-1_3. [DOI] [PubMed] [Google Scholar]
- 8.Akdis M. Immune tolerance in allergy. Current Opinion in Immunology. 2009;21(6):700–707. doi: 10.1016/j.coi.2009.07.012. [DOI] [PubMed] [Google Scholar]
- 9.Dittel BN. CD4 T cells: balancing the coming and going of autoimmune-mediated inflammation in the CNS. Brain, Behavior, and Immunity. 2008;22(4):421–430. doi: 10.1016/j.bbi.2007.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. 2008;133(5):775–787. doi: 10.1016/j.cell.2008.05.009. [DOI] [PubMed] [Google Scholar]
- 11.Ozdemir C, Akdis M, Akdis CA. T regulatory cells and their counterparts: masters of immune regulation. Clinical and Experimental Allergy. 2009;39(5):626–639. doi: 10.1111/j.1365-2222.2009.03242.x. [DOI] [PubMed] [Google Scholar]
- 12.Simpson E. Special regulatory T-cell review: regulation of immune responses—examining the role of T cells. Immunology. 2008;123(1):13–16. doi: 10.1111/j.1365-2567.2007.02775.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bluestone JA, Tang Q. How do CD4+CD25+ regulatory T cells control autoimmunity? Current Opinion in Immunology. 2005;17(6):638–642. doi: 10.1016/j.coi.2005.09.002. [DOI] [PubMed] [Google Scholar]
- 14.Belkaid Y. Regulatory T cells and infection: a dangerous necessity. Nature Reviews Immunology. 2007;7(11):875–888. doi: 10.1038/nri2189. [DOI] [PubMed] [Google Scholar]
- 15.Powrie F, Leach MW, Mauze S, Menon S, Caddle LB, Coffman RL. Inhibition of Th1 responses prevents inflammatory bowel disease in scid mice reconstituted with CD45RBhi CD4+ T cells. Immunity. 1994;1(7):553–562. doi: 10.1016/1074-7613(94)90045-0. [DOI] [PubMed] [Google Scholar]
- 16.Josefowicz SZ, Lu LF, Rudensky AY. Regulatory T cells: mechanisms of differentiation and function. Annual Review of Immunology. 2012;30:531–564. doi: 10.1146/annurev.immunol.25.022106.141623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cava AL, Kaer LV, Fu-Dong S. CD4+CD25+ Tregs and NKT cells: regulators regulating regulators. Trends in Immunology. 2006;27(7):322–327. doi: 10.1016/j.it.2006.05.003. [DOI] [PubMed] [Google Scholar]
- 18.Smith TRF, Kumar V. Revival of CD8+ Treg-mediated suppression. Trends in Immunology. 2008;29(7):337–342. doi: 10.1016/j.it.2008.04.002. [DOI] [PubMed] [Google Scholar]
- 19.Peterson RA. Regulatory T-cells: diverse phenotypes integral to immune homeostasis and suppression. Toxicologic Pathology. 2012;40:186–204. doi: 10.1177/0192623311430693. [DOI] [PubMed] [Google Scholar]
- 20.Fenoglio D, Bernuzzi F, Battaglia F, et al. Th17 and regulatory T lymphocytes in primary biliary cirrhosis and systemic sclerosis as models of autoimmune fibrotic diseases. Autoimmunity Reviews. 2012;12(2):300–304. doi: 10.1016/j.autrev.2012.05.004. [DOI] [PubMed] [Google Scholar]
- 21.Maxion HKM, Liu W, Chang MH, Kelly KA. The infecting dose of Chlamydia muridarum modulates the innate immune response and ascending infection. Infection and Immunity. 2004;72(11):6330–6340. doi: 10.1128/IAI.72.11.6330-6340.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Matsuda JL, Naidenko OV, Gapin L, et al. Tracking the response of natural killer T cells to a glycolipid antigen using CD1d tetramers. Journal of Experimental Medicine. 2000;192(5):741–754. doi: 10.1084/jem.192.5.741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kägi D, Ledermann B, Bürki K, et al. Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice. Nature. 1994;369(6475):31–37. doi: 10.1038/369031a0. [DOI] [PubMed] [Google Scholar]
- 24.Lowin B, Hahne M, Mattmann C, Tschopp J. Cytolytic T-cell cytotoxicity is mediated through perforin and Fas lytic pathways. Nature. 1994;370(6491):650–652. doi: 10.1038/370650a0. [DOI] [PubMed] [Google Scholar]
- 25.Kojima H, Shinohara N, Hanaoka S, et al. Two distinct pathways of specific killing revealed by perforin mutant cytotoxic T lymphocytes. Immunity. 1994;1(5):357–364. doi: 10.1016/1074-7613(94)90066-3. [DOI] [PubMed] [Google Scholar]
- 26.Chattopadhyay PK, Betts MR, Price DA, et al. The cytolytic enzymes granyzme A, granzyme B, and perforin: expression patterns, cell distribution, and their relationship to cell maturity and bright CD57 expression. Journal of Leukocyte Biology. 2009;85(1):88–97. doi: 10.1189/jlb.0208107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Murthy AK, Li W, Chaganty BKR, et al. Tumor necrosis factor alpha production from CD8+ T cells mediates oviduct pathological sequelae following primary genital Chlamydia muridarum Infection. Infection and Immunity. 2011;79(7):2928–2935. doi: 10.1128/IAI.05022-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Niederkorn JY. Emerging concepts in CD8+ T regulatory cells. Current Opinion in Immunology. 2008;20(3):327–331. doi: 10.1016/j.coi.2008.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Igietseme JU, Ananaba GA, Bolier J, et al. Suppression of endogenous IL-10 gene expression in dendritic cells enhances antigen presentation for specific Th1 induction: potential for cellular vaccine development. Journal of Immunology. 2000;164(8):4212–4219. doi: 10.4049/jimmunol.164.8.4212. [DOI] [PubMed] [Google Scholar]
- 30.Payne D, Drinkwater S, Baretto R, Duddridge M, Browning MJ. Expression of chemokine receptors CXCR4, CXCR5 and CCR7 on B and T lymphocytes from patients with primary antibody deficiency. Clinical and Experimental Immunology. 2009;156(2):254–262. doi: 10.1111/j.1365-2249.2009.03889.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kim HJ, Verbinnen B, Tang X, Lu L, Cantor H. Inhibition of follicular T-helper cells by CD8+ regulatory T cells is essential for self tolerance. Nature. 2010;467(7313):328–332. doi: 10.1038/nature09370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Arnold CN, Campbell DJ, Lipp M, Butcher EC. The germinal center response is impaired in the absence of T cell-expressed CXCR5. European Journal of Immunology. 2007;37(1):100–109. doi: 10.1002/eji.200636486. [DOI] [PubMed] [Google Scholar]
- 33.Haynes NM, Allen CDC, Lesley R, Ansel KM, Killeen N, Cyster JG. Role of CXCR5 and CCR7 in follicular Th cell positioning and appearance of a programmed cell death gene-1High germinal center-associated subpopulation. Journal of Immunology. 2007;179(8):5099–5108. doi: 10.4049/jimmunol.179.8.5099. [DOI] [PubMed] [Google Scholar]