

Abstract
The role of savannas during the course of early human evolution has been debated for nearly a century, in part because of difficulties in characterizing local ecosystems from fossil and sediment records. Here, we present high-resolution lipid biomarker and isotopic signatures for organic matter preserved in lake sediments at Olduvai Gorge during a key juncture in human evolution about 2.0 Ma—the emergence and dispersal of Homo erectus (sensu lato). Using published data for modern plants and soils, we construct a framework for ecological interpretations of stable carbon-isotope compositions (expressed as δ13C values) of lipid biomarkers from ancient plants. Within this framework, δ13C values for sedimentary leaf lipids and total organic carbon from Olduvai Gorge indicate recurrent ecosystem variations, where open C4 grasslands abruptly transitioned to closed C3 forests within several hundreds to thousands of years. Carbon-isotopic signatures correlate most strongly with Earth’s orbital geometry (precession), and tropical sea-surface temperatures are significant secondary predictors in partial regression analyses. The scale and pace of repeated ecosystem variations at Olduvai Gorge contrast with long-held views of directional or stepwise aridification and grassland expansion in eastern Africa during the early Pleistocene and provide a local perspective on environmental hypotheses of human evolution.
Keywords: climate, plant waxes, hominins, paleovegetation, paleoclimate
Climate-dependent ecosystem characteristics, such as habitat and water availability, likely influenced natural selection during human evolution (1). For example, woody plants may have influenced thermoregulatory and dietary adaptations in hominins and other terrestrial mammals since the Pleistocene (2–4) about 2.6 Ma. Unfortunately, in many cases, reconstructions of ecosystem characteristics and climate at hominin archaeological sites are limited by poor preservation and coarse temporal resolution. Moreover, discontinuities are common in terrestrial sediment sequences. As a result, much of the environmental context of human evolution has been interpreted based on regional and global conditions reconstructed from marine records (5, 6).
The role of savannas in human evolution remains a subject of debate (5–8). This debate stems, in part, from the historically imprecise definition of savanna for modern and ancient ecosystems and the difficulties of estimating plant community compositions—particularly woody cover—from sediments. Recently, the work by Cerling et al. (7) estimated plant community compositions based on present day relationships between woody cover and carbon-isotope compositions for soil carbonates and soil organic matter (SOM). This approach offers insights into ecosystem structures at hominin archaeological sites (e.g., Omo-Turkana Basin), but it is limited to environments supporting ancient soils (paleosols).
Here, we extend the approach in the work by Cerling et al. (7) to include lipid biomarkers archived in lake sediments deposited between about 2.0 and 1.8 Ma at an important hominin archaeological site—Olduvai Gorge. In addition to including key junctures in human evolution (8), this time interval is associated with important changes in tropical climate, including strengthening of east–west (Walker) atmospheric circulation across the Indian and Pacific Oceans (9). Weakened Walker circulation before about 2.0 Ma was similar to conditions projected to accompany the continued rise in greenhouse gas concentrations during the coming century (10). To examine connections among ocean and atmospheric circulation, regional climate, and plant community composition, we also compare our organic carbon signatures to reconstructions of polar ice volume and sea-surface temperatures (SSTs) in the Atlantic and Indian Oceans.
Background
Site Descriptions.
Olduvai Gorge is just south of the equator in northern Tanzania (2° 48′S, 35° 06′E), where it cuts across a 50-km rift platform basin to expose a 2.0 million y sequence of lake and river sediments (Fig. 1). The basin formed on the western margin of the East African Rift System in response to extension tectonics and the growth of a large volcanic complex (11, 12). During the early Pleistocene, the basin area covered an estimated 3,500 km2 and included a saline–alkaline lake near its center (12). Sediments from this central lake are composed primarily of reworked volcanic material and air-fall tuffs (11). For our study, we use samples recovered from outcrop exposures near the center of the paleolake that preserve a stratigraphic record of continuous deposition (11).
Fig. 1.

Present day precipitation patterns during long rains (A; March to May) and short rains (B; October to December) (75). Bold horizontal lines mark the ITCZ. The IOC (B; dashed line) is prominent during short rains, and its eastward displacement correlates with higher seasonal precipitation (66). Targets show Olduvai Gorge’s location (●, ODP site 662; ○, ODP site 722). (C) Depositional environments at Olduvai Gorge during the early Pleistocene about 2.0–1.8 Ma. Expanded and contracted lake levels are reconstructed based on detailed stratigraphic correlations (11, 12, 43).
Today, annual precipitation patterns at Olduvai Gorge and surrounding regions of eastern Africa are defined by monsoon circulation by two major convergence zones (13)—the Intertropical Convergence Zone (ITCZ) and the Interoceanic Confluence (IOC). The ITCZ marks convergence of regional trade winds, whereas the IOC marks zonal confluence of water vapor derived from the Atlantic and Indian Oceans (Fig. 1). Seasonal migrations of the ITCZ and the IOC result in an annual cycle consisting of two rainy seasons separated by arid conditions that last from May to September. Long rains (March to May) produce the largest proportion of total annual precipitation; short rains (October to December) are more variable but also correlate with total annual precipitation (14). Today, Olduvai Gorge experiences mean annual precipitation (MAP) of about 550 mm; several independent proxy archives suggest that MAP ranged between about 400 and 900 mm during the early Pleistocene (15–17).
We compare our data with coeval records for alkenone-derived SSTs from the eastern Atlantic and western Indian Oceans (18–20) (Fig. 1). Ocean Drilling Program (ODP) site 662 (1° 23′S, 11° 44′W, 3,824 m water depth) is in the eastern Atlantic Ocean in the Gulf of Guinea. ODP site 722 (16° 37′N, 59° 48′E, 2,028 m water depth) is in the northwestern Indian Ocean. Today, SSTs at both sites are sensitive to monsoon-driven seasonal upwelling (21–23), but surface sediment calibrations indicate that alkenone signals reflect mean annual SSTs (22, 23).
Sedimentary Organic Matter.
Organic matter in lake sediments derives from bacteria, algae, and plants (24), and it integrates contributions from the surrounding watershed (25–28). In contrast, organic matter in soils depends more strongly on local sources and preservation (29, 30), and it can vary spatially over scales of 10 m2 or less (25).
Biomarkers are molecular fossils that have structures with biological specificity (31). Those biomarkers from plants and algae carry isotopic signals derived from terrestrial and aquatic sources, respectively. For instance, long straight-chained hydrocarbons, such as nonacosane (nC29) and nC31, occur abundantly in leaves (32) and indicate sedimentary inputs from terrestrial plants (33). Although somewhat less specific, certain shorter-chained hydrocarbons (e.g., nC17 and nC19) indicate contributions from algae and cyanobacteria (31).
Carbon Isotopes in Leaves, Biomarkers, and Soil Organic Matter.
Photosynthetic carbon-isotopic fractionation is defined between atmospheric carbon dioxide (δ13CCO2) and leaf tissues (δ13Cleaf), and its magnitude varies with plant functional type and water availability (34, 35):
We note that ɛ-values are expressed in permil, which is a unit of parts per thousand. Variability in ɛCO2/leaf, which is approximately equivalent to negative Δleaf values (widely used in ecological literature) (34), is well-constrained for different photosynthetic pathways (35), but these modern relationships are not easily applied to ancient plants because of limited preservation of leaf tissues through time. Relatively recalcitrant leaf lipids, such as nC31, afford an opportunity to circumvent this challenge provided that fractionation between δ13Cleaf and nC31 (δ13C31) during biosynthesis can be documented for subtropical and tropical plants (36).
We evaluated carbon-isotopic relationships between soil organic matter (δ13CSOM), leaf tissues, and leaf lipids using published data for 64 plants species and 288 soils from nearly 300 tropical and subtropical localities (Fig. 2). For C3 plant soil systems, we find that nC31 is 13C-depleted by about 7‰ (n = 45) with respect to leaf tissue, whereas SOM is 13C-enriched by about 2‰ (n = 184) relative to leaves. Thus, in C3 ecosystems, there is an isotopic difference (ɛSOM/31) between nC31 and SOM equal to about 9‰:
Fig. 2.

Histograms and box-and-whisker plots for ɛSOM/31 from published δ13CSOM and δ13C31 values from subtropical and tropical regions (SI Appendix). Published data are binned at 0.5‰ intervals (counts) based on site averages. Values for δ13CSOM (dark gray) and δ13C31 (light gray) are plotted relative to δ13Cleaf. Values of ɛSOM/31 equal about 9‰ in both C3 and C4 ecosystems. Individual boxes contain interquartile ranges (IQRs). Bold vertical lines within boxes mark median values. ○, mean values. Horizontal whiskers mark minimum and maximum values, except values outside of 1.5 IQR (●). Notch half-width values indicate confidence in differentiating median values.
In C4 plant soil systems, both nC31 and SOM are 13C-depleted with respect to leaf tissue by about 10‰ (n = 19) and 1‰ (n = 104), respectively. Accordingly, ɛSOM/31 in C4 ecosystems also equals about 9‰. Therefore, we apply ɛSOM/31 of 9‰ to estimate δ13CSOM from δ13C31 values, thus extending to plant waxes the predictive capabilities of δ13CSOM for estimating woody cover (7) (Fig. 3). We caution that the value of 9‰ for ɛSOM/31 derived here is based on data from xeric woodlands and scrublands, tropical deciduous forests, and C4 grasslands, and it may not be representative for other ecosystems (36).
Fig. 3.

Stable carbon-isotope compositions for SOM (δ13CSOM; bottom x axis) and nC31 (δ13C31; top x axis) relative to woody plant cover (A), ecosystem (B), and plant community composition (C). The published relationship between woody plant cover and δ13CSOM is described by the following function, where −13 ≤ δ13CSOM ≥ −31‰ (10): 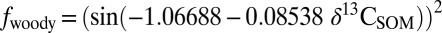 . Here, we relate δ13CSOM to δ13C31 using a value of 9‰ for ɛSOM/31. The relationship between woody plant cover and δ13C31 is, thus, described by the following function, where −22 ≤ δ13CSOM ≥ −40‰):
. Here, we relate δ13CSOM to δ13C31 using a value of 9‰ for ɛSOM/31. The relationship between woody plant cover and δ13C31 is, thus, described by the following function, where −22 ≤ δ13CSOM ≥ −40‰): 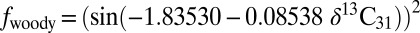 . Ecosystem definitions adhere to African plant communities according to UNESCO terminology (40).
. Ecosystem definitions adhere to African plant communities according to UNESCO terminology (40).
The fractional abundance of C3 plants is inversely related to C4 plants in tropical ecosystems (25, 37). However, the relationship between woody cover (fwoody) and C4 abundance is nonlinear (7), because herbaceous C3 plants can occur in both open and wooded ecosystems (38). As a result, for δ13CSOM values between near-total C3 composition (−30‰) and negligible C3 cover (−14‰), we follow the approach in the work by Cerling et al. (7) to estimate fwoody (where −13 ≤ δ13CSOM ≥ −31‰):
Structural Classification for Ancient Ecosystems.
The broad functional definition for savannas as a continuous herbaceous understory with irregular distributions of trees or bushes does not account for differences in fwoody (39). Therefore, we adopt definitions for African plant communities defined by the United Nations Educational, Scientific and Cultural Organization (UNESCO) (40). According to this classification scheme, (i) forests display continuous tree cover (>10 m) with interlocking crowns and poorly developed understory, (ii) woodlands—including bush/shrublands—display open or closed stands of shrubs or trees (up to 8 m) with at least 40% woody plant cover and understory with grasses and other herbs, (iii) wooded grasslands display 10–40% woody plant cover and well-developed groundcover with grasses and other herbs, (iv) grasslands display less than 10% woody plant cover and well-developed groundcover with grasses and other herbs, and (v) deserts display sparse groundcover and sandy, stony, or rocky substrate. UNESCO does not distinguish between forests and woodlands in terms of woody plant cover, but here, we consider forests to display greater than 80% woody plant cover (7, 39).
The evolutionary implications of orbital forcing on environmental change were recognized over a century ago (41) but remain controversial today (3–6). Marine sediments just off African shores document orbital rhythms in terrestrial inputs during the Pleistocene (5), and a growing number of terrestrial sequences hint at a similar pacing for environmental changes in eastern Africa (42, 43). Marine sequences are often indirectly or too poorly constrained in time to infer relationships between key junctures in human evolution and terrestrial conditions or change (6). Lithostratigraphic patterns in lake margin sediments at Olduvai Gorge reveal five episodes of lake expansion between ca. 1.85 and 1.74 Ma, suggesting that lake level changes may have tracked orbital precession (43). Here, we use sedimentary organic matter signatures in high-resolution and temporally well-constrained lake sediments to evaluate the magnitude and timing of ecosystem changes associated with lake level changes.
Results and Discussion
Ecosystem Change and Woody Cover.
We observe repeated shifts in δ13C31 values between about −36‰ and −20‰ (Fig. 4), which track closely with orbitally paced lake margin lithostratigraphic patterns, suggesting that ecosystem changes at Olduvai Gorge were also influenced by orbital cycles (Fig. 5). Total organic carbon (TOC) δ13C values (δ13CTOC) show a smaller isotopic range of about 9‰ and correlate tightly with δ13C31 values (r2 = 0.86); δ13CTOC values follow terrestrial plant inputs but are attenuated, likely by algal or macrophytic inputs. Taken together, δ13C31 and δ13CTOC values suggest rapid local ecosystem shifts between closed C3 woodlands and open C4 grasslands. These changes were comparable with extreme events, such as the greening of the Sahara about 120,000 y ago, that accompanied the dispersal of modern humans out of Africa (44).
Fig. 4.
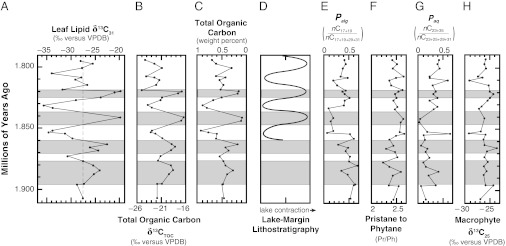
Biomarker and isotopic signatures for organic matter preserved in lake sediments at Olduvai Gorge. (A) Leaf lipid δ13C values for nC31 (δ13C31). (B) TOC δ13C values (δ13CTOC). (C) TOC percentages (%TOC). (D) Diagrammatic depiction of relative lake level changes at Olduvai Gorge based on lake margin lithostratigraphy (43). (E) Ratios of algal lipids (nC17 + nC19) relative to algal and terrestrial plant lipids (nC17 + nC19 + nC29 + nC31). Higher values reflect relatively increased algal inputs (31). (F) Ratios of pristane (Pr) to phytane (Ph). In lake sediments, values above two generally reflect a dominance of terrestrial plant inputs (31). (G) Ratios of macrophytic lipids (nC23 + nC25) relative to macrophytic and terrestrial plant lipids (nC23 + nC25 + nC29 + nC31; that is, Paq). Higher values reflect relatively increased macrophytic inputs (50). (H) Values of δ13C for the macrophytic lipid nC25 (δ13C25). Horizontal gray bands highlight periods of time characterized by δ13C31 values higher than about −28‰ (i.e., open grassland and wooded grassland ecosystems).
Fig. 5.
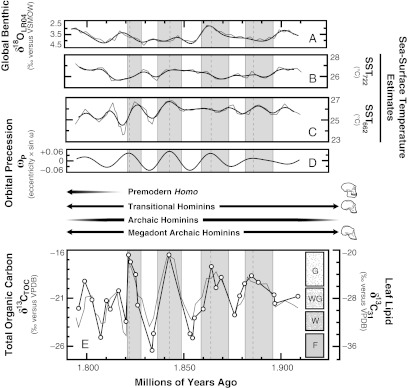
Time series for global, regional, and local proxy indicators during the early Pleistocene. (A) Global benthic oxygen-isotopic composite (19). Gray lines connect original data, and the bold black line shows a five-point smoothing. (B and C) Alkenone-derived SST estimates for the western Indian Ocean (SST722) and eastern Atlantic Ocean (SST662), respectively (18–20). Gray lines connect original data, and bold black lines show five-point smoothing. (D) Calculated orbital precession (ωp). Values for ωp equal the product of calculated eccentricity (e) and the sine function of the longitude of the perihelion (ω). (E) Leaf lipid δ13C values for nC31 (δ13C31; ○ connected by bold black lines) and TOC δ13C values (δ13CTOC; gray lines). F, forest; G, grassland; W, bush/shrubland and woodland; WG, wooded grassland. Ecosystem definitions adhere to African plant communities according to UNESCO terminology (40) based on δ13C31 values (Fig. 4). Vertical gray bands highlight periods of time characterized by δ13C31 values higher than about −28‰ (i.e., open grassland and wooded grassland ecosystems). Hominin taxonomic grades and fossil occurrences are illustrated using information in the work by Wood (76); skulls denote taxonomic grades that have been identified at Olduvai Gorge.
Carbon-isotope evidence for pronounced ecosystem shifts at Olduvai Gorge contrasts with previous reconstructions for eastern Africa that proposed that ecosystems were stable at local to regional scales in the early Pleistocene (15, 45, 46) and generally lacked closed woodlands near hominin archaeological sites since 6 Ma (7). Dramatic and rapid changes in δ13C31 values highlight ecosystem instability in this region; furthermore, δ13C31 values indicate that closed woodlands dominated local landscapes for up to 20% of the time.
Differences in the interpretation of ancient ecosystems, in addition to problems related to savanna heterogeneity, can stem from inherent proxy biases (6). For instance, δ13CSOM values can overrepresent C4 inputs as a result of 13C enrichment during organic matter decomposition (47). Values for δ13C31 can also overrepresent C4 inputs as a result of inorganic carbon (e.g., bicarbonate) assimilation by macrophytes, although δ13C31 values in arid environments are more likely biased to wet conditions (when plants synthesize most leaf lipids) (48). Although specific mechanisms responsible for differences among ecosystem proxies cannot necessarily be reconciled here, we suggest that carbon-isotopic signals for leaf lipids can complement δ13CSOM values, which otherwise can skew to C4—and thus, arid—signals on seasonal and longer timescales.
Biogeochemical Variability at the Ecosystem Scale.
Organic matter in lake sediments incorporates inputs from both aquatic and terrestrial photosynthetic organisms and can vary in proportion with productivity and deposition in a lake and the surrounding watershed. At Olduvai Gorge, δ13CTOC values correlate strongly with TOC (%TOC; r2 = 0.84) but show no clear relationship with algal vs. terrestrial plant inputs (Fig. 4):
The abundance ratio of pristane vs. phytane is constantly near 2.5, suggesting that organic carbon deposition was dominated by terrestrial plant input (31). These observations corroborate strong correlation between δ13CTOC and δ13C31 values and suggest that δ13CTOC values primarily follow terrestrial plant inputs.
Macrophytes contain abundant intermediate-chain n-alkanes (e.g., nC23 and nC25) but limited long-chain homologs compared with most terrestrial plants (49, 50). Thus, relative abundances of nC23 and nC25 vs. nC29 and nC31 (that is, Paq) can provide estimates for macrophytic vs. terrestrial plant inputs (50):
In lake sediments from Olduvai Gorge, δ13C31 values correlate weakly with both Paq (r2 = 0.17) and nC25 δ13C values (r2 = 0.19), suggesting that macrophytes did not significantly contribute to nC31 inputs. Paq and Palg are measures of relative abundance of molecules. They do not directly indicate a proportion of biomass from different organisms; instead, these ratios are useful for identifying organic facies or correlations with other biogeochemical signals.
Mechanisms of Ecosystem Change.
Atmospheric CO2 concentrations (pCO2), temperature, seasonality, and water availability are potential determinants of C3 vs. C4 plant abundance (38). Since the middle Pleistocene, records of pCO2 correlate strongly with polar ice volume changes (51, 52), which were obliquity paced before ∼1 Ma (5). Lake sediment δ13CTOC values for Olduvai Gorge correlate weakly with reconstructed polar ice volumes (r2 = 0.16) based on marine oxygen-isotopic records (9, 19), suggesting that ecosystem changes in this region did not track 41,000-y glacial cycles. If polar ice volume is a representative proxy for pCO2 during the early Pleistocene, then local ecosystem changes were not exclusively tied to pCO2 changes. This conclusion contrasts with suggestions of a dominant role for pCO2 in southern African ecosystems during the early Pleistocene based on speleothem carbonate δ13C values (53), but it is in agreement with marine oxygen-isotopic evidence for eastern African climate sensitivity to polar ice volume only after 1.0 Ma (54). Values for δ13CTOC correlate strongly with precession (ωp) and thus, do not support temperature as primary determinant of ecosystem change, because ωp negligibly influences mean annual temperatures (55). Similarly, paleosol carbonates indicate that mean annual temperatures varied by less than about ±5 °C at and around Olduvai Gorge during the early Pleistocene (56, 57). Strong correlation between δ13CTOC and ωp also suggests that biotic (e.g., herbivory) or abiotic disturbances, such as fire, were not primary determinants of ecosystem change, although they may have served as feedback mechanisms that accelerated changes. In agreement with a variety of other studies (58, 59), we suggest that changing C3 and C4 plant abundances at Olduvai Gorge varied with orbital precession in response to water availability.
Mechanisms of Hydroclimate Change.
Cycles of about 21,000 y are common in a variety of hydroclimate proxy records in eastern Africa since the Pliocene (5), and δ13CTOC values correlate strongly with ωp (r2 = 0.61) in single-factor regression (SI Appendix). Although specific mechanisms responsible for these cycles remain unclear, the timing and magnitude of local and regional hydroclimate changes are consistent with theoretical effects of ωp on monsoon strength (60) (that is, higher summer insolation would enhance land–ocean temperatures contrasts, resulting in stronger monsoons and increased precipitation).
Insolation alone cannot account for the magnitude of hydroclimate change in eastern Africa (61). Previous reconstructions based on pollen and oxygen-isotope compositions of soil carbonates suggest that MAP fluctuated between ∼400 and 800 mm at Olduvai Gorge and surrounding regions during the early Pleistocene (15–17, 43). However, in climate simulations, insolation variability accounts for MAP fluctuations of less than 200 mm and mostly affects long rains (60, 61). Thus, precipitation amounts in the past must have been impacted by multiple factors, the same as they are today (13).
Today, precipitation in eastern Africa responds sensitively to SSTs in the Indian Ocean and Atlantic Ocean (62). In particular, intensifications of short rains (up to 200 mm) accompany coordinated warm and cold SST anomalies in the western Indian Ocean and eastern Atlantic Ocean (63–65), respectively, as a result of transcontinental surface pressure gradients across Africa and monsoon displacement of the IOC from west to east (66). Partial regressions reveal that SSTs for ODP sites 662 (SST662) and 722 (SST722) are significant (P < 0.001) secondary predictors that are statistically independent of covariance with ωp, and the combination of ωp, SST662, and SST722 explains 73% of the variability in δ13CTOC values in a multiple regression model. During the early Pleistocene, both SST662 and SST722 show strong ωp and 41,000-y (obliquity) periodicity (20), but only SST662 shows a consistent relationship with monsoon-driven upwelling (20). Because upwelling in the eastern Atlantic correlates positively with monsoon strength during late boreal summer (20–22), we suggest that obliquity-paced cooling in the eastern Atlantic Ocean and monsoon strengthening (and therefore, stronger westerly winds) resulted in more frequent eastward displacements of the IOC and intensification of short rains in eastern Africa.
Ecosystems and Hominin Evolution.
Fossil evidence for pronounced aridification and faunal turnover in eastern Africa between about 2.0 and 1.8 Ma has sparked hypotheses linking the emergence and dispersal of the genus Homo to climate-driven ecosystem change (2–6). Fossil evidence for cranial expansion in premodern Homo (e.g., H. erectus sensu lato) has been linked to irregular resource distributions (67), and our carbon-isotopic data are consistent with enhanced ecosystem variability as a context for encephalization (Fig. 5). During the early Pleistocene, strong ecosystem preferences are not apparent between transitional (e.g., H. habilis) and archaic (e.g., Paranthropus boisei) hominins (68); however, isotopic and fossil data suggest that transitional species accessed a broad spectrum of dietary resources compared with archaic species (68–70). Among primates, quality (i.e., energy density) of dietary resources correlates strongly with brain size (67). Assuming that dietary resources were primarily unrelated to technological innovations by transitional species (4, 68), we hypothesize that ecosystem variability favored hominin species with large brains that allowed for versatile foraging strategies and dietary diversity.
Conclusions
This study presents high-resolution biomarker and δ13C records of ecosystem variability from lake sediments at Olduvai Gorge that were deposited during an interval of pronounced shifts in vertebrate community and global climate reorganization about 2.0–1.8 Ma. Values of δ13C31 indicate rapid and repeated ecosystem restructuring between closed C3 woodlands and open C4-dominated grasslands. Our δ13C records reveal coupled fluctuations between ecosystem and precession. Additional variability is explained by differences in SST between the Atlantic and Indian Oceans. These observations suggest aridity-controlled, as opposed to carbon dioxide- or temperature-controlled, woody plant cover in eastern Africa during the early Pleistocene. We conclude that highly variable ecosystems accompanied the emergence and dispersal of the genus Homo. Our study also builds on soil data to construct an interpretive framework for ecosystem reconstruction based on leaf lipids.
Materials and Methods
Reconstructions for Polar Ice Volume.
We use the approach in the work by Bintanja and van de Wal (71), which assumes that a global composite record of benthic δ18O values (LR04 Stack) accurately traces polar ice volume during the early Pleistocene (19).
Multiple Regression Analysis.
We used rank transformation (72) and Fourier cross-correlation (73) to compare nonlinear, time-shifted, and unevenly sampled proxy records (SI Appendix). Because ωp correlates most strongly with δ13CTOC in single-factor analyses (r2 = 0.61), we constrain multiple regression models for δ13CTOC values to always include ωp. This approach is justified, because the influence of ωp on climate in eastern Africa is well-supported by theory and other observations (10, 61). Because ωp is correlated with other factors, care is required in evaluating additional influences on δ13CTOC values. We assessed the influence of other factors on δ13CTOC values using multivariate partial regression models that account for covariance between ωp and other factors. Partial regression models with SST662 (partial r2 = 0.17) and SST722 (partial r2 = 0.11) as secondary predictors are the only models with notable explanatory power for δ13CTOC values. The following equation shows the δ13CTOC relationship shared with ωp, SST662, and SST722 (r2 = 0.73):
Lipid Extraction and Purification.
Freeze-dried and ground lake sediments were Soxhlet-extracted with dichloromethane:methanol (9:1 vol/vol) for 12 h. Total lipid extracts were then separated into apolar and polar fractions over alumina with hexanes and methanol, respectively. Apolar molecules were separated into saturated and unsaturated fractions over 5% (wt/wt) silver nitrate-impregnated alumina with hexanes and dichloromethane, respectively. Finally, unsaturated apolar compounds were separated with a 5-Å molecular sieve to isolate n-alkanes.
Isotopic Analysis.
All δ13CTOC values were measured after decarbonation of ground lake sediments with excess 2N hydrochloric acid (74). Residual materials were combusted in an elemental analyzer, and δ13CTOC values were measured in a ThermoFinnigan Delta+ XP. Standard reference materials of known δ13C values, including polyethylene foil (NIST 8540), were used throughout sample runs to ensure accuracy. SD (1σ) for NIST 8540 equaled 0.1‰.
Compound-specific δ13C values were measured by GC–isotope ratio monitoring–MS (Delta+ XP; ThermoFinnigan) with a combustion interface. Carbon dioxide gas of known δ13C value was used as an external standard; internal standards (i.e., nC41, androstane, and squalane; Schimmelmann Standards) were used throughout sample runs to ensure accuracy. SD (1σ) equaled 0.3‰.
Supplementary Material
Acknowledgments
We are grateful to Richard Hay (1926–2006), whose pioneering work at Olduvai Gorge inspired this research. We thank the Winston Churchill Foundation, Carbon Educators and Researchers Together for Humanity (CarbonEARTH) (National Science Foundation Grant Division of Graduate Education 0947962), and the Tanzania Antiquities Department. We also thank the Ngorongoro Conservation Area Authority for field permits to R. Blumenschine and F. Masao. G.M.A. and R. Renaut collected all samples (National Science Foundation Grant Earth Sciences 9903258).
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
See Commentary on page 1144.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1206276110/-/DCSupplemental.
References
- 1.Sabeti PC, et al. Positive natural selection in the human lineage. Science. 2006;312(5780):1614–1620. doi: 10.1126/science.1124309. [DOI] [PubMed] [Google Scholar]
- 2.Teaford MF, Ungar PS. Diet and the evolution of the earliest human ancestors. Proc Natl Acad Sci USA. 2000;97(25):13506–13511. doi: 10.1073/pnas.260368897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pickering TR, Bunn HT. The endurance running hypothesis and hunting and scavenging in savanna-woodlands. J Hum Evol. 2007;53(4):434–438. doi: 10.1016/j.jhevol.2007.01.012. [DOI] [PubMed] [Google Scholar]
- 4.McNabb J. Hominins and the Early-Middle Pleistocene transition: evolution, culture and climate in Africa and Europe. Geological Society Special Publication. 2005;247:287–304. [Google Scholar]
- 5.deMenocal P. African climate change and faunal evolution during the Pliocene-Pleistocene. Earth Planet Sci Lett. 2004;220(1-2):3–24. [Google Scholar]
- 6.Kingston JD. Shifting adaptive landscapes: Progress and challenges in reconstructing early hominid environments. Am J Phys Anthropol. 2007;134(Suppl 45):20–58. doi: 10.1002/ajpa.20733. [DOI] [PubMed] [Google Scholar]
- 7.Cerling TE, et al. Woody cover and hominin environments in the past 6 million years. Nature. 2011;476(7358):51–56. doi: 10.1038/nature10306. [DOI] [PubMed] [Google Scholar]
- 8.Wood B, Harrison T. The evolutionary context of the first hominins. Nature. 2011;470(7334):347–352. doi: 10.1038/nature09709. [DOI] [PubMed] [Google Scholar]
- 9.Ravelo AC, Andreasen DH, Lyle M, Olivarez Lyle A, Wara MW. Regional climate shifts caused by gradual global cooling in the Pliocene epoch. Nature. 2004;429(6989):263–267. doi: 10.1038/nature02567. [DOI] [PubMed] [Google Scholar]
- 10.Haywood AM, et al. Introduction: Pliocene climate, processes and problems. Philos Transact A Math Phys Eng Sci. 2009;367:3–17. doi: 10.1098/rsta.2008.0205. [DOI] [PubMed] [Google Scholar]
- 11.Hay R. Geology of the Olduvai Gorge. Los Angeles: University of California Press; 1976. [Google Scholar]
- 12.Ashley G, Hay R. Sedimentation patterns in a Plio-Pleistocene volcaniclastic rift-platform basin, Olduvai Gorge, Tanzania. In: Renaut R, Ashley G, editors. Sedimentation in Continental Rifts. Tulsa, OK: Society for Sedimentary Geology; 2002. pp. 107–122. [Google Scholar]
- 13.Nicholson S. The nature of rainfall variability over Africa on time scales of decades to millenia. Global Planet Change. 2000;26(1-3):137–158. [Google Scholar]
- 14.Moron V, Robertson A, Ward M, Camberlin P. Spatial coherence of tropical rainfall at the regional scale. J Clim. 2007;20(21):5244–5263. [Google Scholar]
- 15.Bonnefille R, et al. Glacial/interglacial record from intertropical Africa, high resolution pollen and carbon data at Rusaka, Burundi. Quat Sci Rev. 1995;14(9):917–936. [Google Scholar]
- 16.Sikes NE, Ashley GM. Stable isotopes of pedogenic carbonates as indicators of paleoecology in the Plio-Pleistocene (upper Bed I), western margin of the Olduvai Basin, Tanzania. J Hum Evol. 2007;53(5):574–594. doi: 10.1016/j.jhevol.2006.12.008. [DOI] [PubMed] [Google Scholar]
- 17.Bergner A, Trauth M, Bookhagen B. Paleoprecipitation estimates for the Lake Naivasha basin (Kenya) during the last 175 k.y. using a lake-balance model. Global Planet Change. 2003;36(1-2):117–136. [Google Scholar]
- 18.Clemens SC, Murray DW, Prell WL. Nonstationary phase of the Plio-Pleistocene Asian monsoon. Science. 1996;274(5289):943–948. doi: 10.1126/science.274.5289.943. [DOI] [PubMed] [Google Scholar]
- 19.Lisiecki L, Raymo M. A Plio-Pleistocene stack of 57 globally distributed benthic 18O records. Paleoceanography. 2005 doi: 10.1029/2004PA001071. [DOI] [Google Scholar]
- 20.Cleaveland L, Herbert T. Coherent obliquity band and heterogeneous precession band responses in early Pleistocene tropical sea surface temperatures. Paleoceanography. 2007 doi: 10.1029/2006PA001370. [DOI] [Google Scholar]
- 21.McIntyre A, Ruddiman W, Karlin K, Mix A. Surface water response of the equatorial Atlantic Ocean to orbital forcing. Paleoceanography. 1989 doi: 10.1029/PA004i001p00019. [DOI] [Google Scholar]
- 22.Müller P, et al. Calibration of the alkenone paleotemperature index U37K' based on core-tops from the eastern South Atlantic and the global ocean (60°N–60°S) Geochim Cosmochim Acta. 1998;62(10):1757–1772. [Google Scholar]
- 23.Sonzogni C, et al. Temperature and salinity effects on alkenone ratios measured in surface sediments from the Indian Ocean. Quat Res. 1997;47(3):344–355. [Google Scholar]
- 24.Mead R, et al. Sediment and soil organic matter source assessment as revealed by the molecular distribution and carbon isotopic composition of n-alkanes. Org Geochem. 2005;36(3):363–370. [Google Scholar]
- 25.Wynn J, Bird M. Environmental controls on the stable carbon isotopic composition of soil organic carbon: Implications for modelling the distribution of C3 and C4 plants, Australia. Tellus B Chem Phys Meterol. 2008;60(4):604–621. [Google Scholar]
- 26.Teisserenc R, et al. Integrated transfers of terrigenous organic matter to lakes at their watershed level: A combined biomarker and GIS analysis. Geochim Cosmochim Acta. 2010;74(22):6375–6386. [Google Scholar]
- 27.Oline D, Grant M. Scaling patterns of biomass and soil properties: An empirical analysis. Landsc Ecol. 2002;17(1):13–26. [Google Scholar]
- 28.Aleman J, et al. Reconstructing savanna tree cover from pollen, phytoliths and stable carbon isotopes. J Veg Sci. 2012;23(1):187–197. [Google Scholar]
- 29.Bai E, et al. Spatial patterns of soil δ13C reveal grassland-to-woodland successional processes. Org Geochem. 2012;42(12):1512–1518. [Google Scholar]
- 30.Levin N, et al. Paleosol carbonates from the Omo Group: Isotopic records of local and regional environmental change in East Africa. Palaeogeogr Palaeoclimatol Palaeoecol. 2011;307(1-4):75–89. [Google Scholar]
- 31.Peters K, Walters C, Moldowan J. The Biomarker Guide. Cambridge, New York: Cambridge Univ Press; 2005. [Google Scholar]
- 32.Eglinton G, Hamilton RJ. Leaf epicuticular waxes. Science. 1967;156(3780):1322–1335. doi: 10.1126/science.156.3780.1322. [DOI] [PubMed] [Google Scholar]
- 33.Freeman K, Colarusso L. Molecular and isotopic records of C4 grassland expansion in the late Miocene. Geochim Cosmochim Acta. 2001;65(9):1439–1454. [Google Scholar]
- 34.Diefendorf AF, Mueller KE, Wing SL, Koch PL, Freeman KH. Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proc Natl Acad Sci USA. 2010;107(13):5738–5743. doi: 10.1073/pnas.0910513107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bowling DR, Pataki DE, Randerson JT. Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes. New Phytol. 2008;178(1):24–40. doi: 10.1111/j.1469-8137.2007.02342.x. [DOI] [PubMed] [Google Scholar]
- 36.Diefendorf A, et al. Production of n-alkyl lipids in living plants and implications for the geologic past. Geochim Cosmochim Acta. 2011;75(23):7472–7485. [Google Scholar]
- 37.Lloyd J, et al. Contributions of woody and herbaceous vegetation to tropical savanna ecosystem productivity: A quasi-global estimate. Tree Physiol. 2008;28(3):451–468. doi: 10.1093/treephys/28.3.451. [DOI] [PubMed] [Google Scholar]
- 38.Pearcy R, Ehleringer J. Comparative ecophysiology of C3 and C4 plants. Plant Cell Environ. 1984;7(1):1–13. [Google Scholar]
- 39.Ratnam J, et al. When is a ‘forest’ a savanna, and why does it matter? Glob Ecol Biogeogr. 2011;20(5):653–660. [Google Scholar]
- 40.White F. The Vegetation of Africa. United Nations Scientific and Cultural Organization, Paris; 1983. [Google Scholar]
- 41.Wallace A. Man and natural selection. Nature. 1870;3(53):8–9. [Google Scholar]
- 42.Deino A, et al. Precessional forcing of lacustrine sedimentation in the late Cenozoic Chemeron Basin, Central Kenya Rift, and calibration of the Gauss/Matuyama boundary. Earth Planet Sci Lett. 2006;247(1-2):41–60. [Google Scholar]
- 43.Ashley G. Orbital rhythms, monsoons, and playa lake response, Olduvai Basin, equatorial East Africa (ca. 1.85–1.74 Ma) Geology. 2007;35(12):1091–1094. [Google Scholar]
- 44.Osborne AH, et al. A humid corridor across the Sahara for the migration of early modern humans out of Africa 120,000 years ago. Proc Natl Acad Sci USA. 2008;105(43):16444–16447. doi: 10.1073/pnas.0804472105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Reed K, Rector A. African Pliocene paleoecology. In: Ungar P, editor. Evolution of the Human Diet. New York: Oxford University Press; 2007. pp. 262–288. [Google Scholar]
- 46.Bamford M, et al. Late Pliocene grassland from Olduvai Gorge, Tanzania. Palaeogeogr Palaeoclimatol Palaeoecol. 2008;257(3):280–293. [Google Scholar]
- 47.Wynn J. Carbon isotope fractionation during decomposition of organic matter in soils and paleosols: Implications for paleoecological interpretations of paleosols. Palaeogeogr Palaeoclimatol Palaeoecol. 2007;251(3-4):437–448. [Google Scholar]
- 48.Post-Beittenmiller D. Biochemistry and molecular biology of wax production in plants. Annu Rev Plant Physiol Plant Mol Biol. 1996;47(1):405–430. doi: 10.1146/annurev.arplant.47.1.405. [DOI] [PubMed] [Google Scholar]
- 49.Lamb A, Leng M, Umer Mohammed M, Lamb H. Holocene climate and vegetation change in the Main Ethiopian Rift Valley, inferred from the composition (C/N and δ13C) of lacustrine organic matter. Quat Sci Rev. 2004;23(7-8):881–891. [Google Scholar]
- 50.Ficken K, Li B, Swain D, Eglinton G. An n-alkane proxy for the sedimentary input of submerged/floating freshwater aquatic macrophytes. Org Geochem. 2000;31(7-8):745–749. [Google Scholar]
- 51.Petit J, et al. Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature. 1999;399(6735):429–436. [Google Scholar]
- 52.Lüthi D, et al. High-resolution carbon dioxide concentration record 650,000–800,000 years before present. Nature. 2008;453(7193):379–382. doi: 10.1038/nature06949. [DOI] [PubMed] [Google Scholar]
- 53.Hopley P, et al. High-and low-latitude orbital forcing of early hominin habitats in South Africa. Earth Planet Sci Lett. 2007;256(3-4):419–432. [Google Scholar]
- 54.Trauth M, Larrasoaña J, Mudelsee M. Trends, rhythms and events in Plio-Pleistocene African climate. Quat Sci Rev. 2009;28(5-6):399–411. [Google Scholar]
- 55.Kingston J. Orbital controls on seasonality. In: Brockman D, van Schaik C, editors. Seasonality in Primates. New York: Cambridge University Press; 2005. pp. 521–543. [Google Scholar]
- 56.Hay R, Kyser T. Chemical sedimentology and paleoenvironmental history of Lake Olduvai, a Pliocene lake in northern Tanzania. Geol Soc Am Bull. 2001;113(12):1505–1521. [Google Scholar]
- 57.Passey BH, Levin NE, Cerling TE, Brown FH, Eiler JM. High-temperature environments of human evolution in East Africa based on bond ordering in paleosol carbonates. Proc Natl Acad Sci USA. 2010;107(25):11245–11249. doi: 10.1073/pnas.1001824107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Huang Y, et al. Climate change as the dominant control on glacial-interglacial variations in C3 and C4 plant abundance. Science. 2001;293(5535):1647–1651. doi: 10.1126/science.1060143. [DOI] [PubMed] [Google Scholar]
- 59.Bobe R. The evolution of arid ecosystems in eastern Africa. J Arid Environ. 2006;66(3):564–584. [Google Scholar]
- 60.Ruddiman W. Earth’s Climate: Past and Future. New York: Freeman; 2001. [Google Scholar]
- 61.Clement A, Hall A, Broccoli A. The importance of precessional signals in the tropical climate. Clim Dyn. 2004;22(4):327–341. [Google Scholar]
- 62.Chiang J. The tropics in paleoclimate. Annu Rev Earth Planet Sci. 2009;37(1):263–297. [Google Scholar]
- 63.Vuille M, et al. Stable isotopes in East African precipitation record Indian Ocean zonal mode. Geophys Res Lett. 2005 doi: 10.1029/2005JD006022. [DOI] [Google Scholar]
- 64.Balas N, et al. The relationship of rainfall variability in West Central Africa to sea-surface temperature fluctuations. Int J Climatol. 2007;27(10):1335–1349. [Google Scholar]
- 65.Farnsworth A, et al. Understanding the large scale driving mechanisms of rainfall variability over Central Africa. In: Williams C, Kniveton D, editors. African Climate and Climate Change. New York: Springer; 2011. pp. 101–122. [Google Scholar]
- 66.Eltahir E, Gong C. Dynamics of wet and dry years in West Africa. J Clim. 1996;9(5):1030–1042. [Google Scholar]
- 67.Snodgrass J, Leonard W, Robertson M. The energetics of encephalization in early hominids. In: Hublin J, Richards M, editors. The Evolution of Hominin Diets. New York: Springer; 2009. pp. 15–29. [Google Scholar]
- 68.Wood B, Strait D. Patterns of resource use in early Homo and Paranthropus. J Hum Evol. 2004;46(2):119–162. doi: 10.1016/j.jhevol.2003.11.004. [DOI] [PubMed] [Google Scholar]
- 69.van der Merwe N, Masao F, Bamford M. Isotopic evidence for contrasting diets of early hominins Homo habilis and Australopithecus boisei of Tanzania. S Afr J Sci. 2008;104(3-4):153–155. [Google Scholar]
- 70.Cerling TE, et al. Diet of Paranthropus boisei in the early Pleistocene of East Africa. Proc Natl Acad Sci USA. 2011;108(23):9337–9341. doi: 10.1073/pnas.1104627108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bintanja R, van de Wal RS. North American ice-sheet dynamics and the onset of 100,000-year glacial cycles. Nature. 2008;454(7206):869–872. doi: 10.1038/nature07158. [DOI] [PubMed] [Google Scholar]
- 72.Chatfield C. The Analysis of Time Series. London: CRC Press; 2004. [Google Scholar]
- 73.Scargle J. Studies in astronomical time series analysis. Statistical aspects of spectral analysis of unevenly spaced data. Astrophys J. 1982;263(1):835–853. [Google Scholar]
- 74.Brodie CR, et al. Evidence for bias in C and N concentrations and δ13C composition of terrestrial and aquatic organic materials due to pre-analysis acid preparation methods. Chem Geol. 2011;282(3-4):67–83. [Google Scholar]
- 75.Mitchell T, Jones P. An improved method of constructing a database of monthly climate observations and associated high resolution grids. Int J Climatol. 2005;25(6):693–712. [Google Scholar]
- 76.Wood B. Colloquium paper: Reconstructing human evolution: achievements, challenges, and opportunities. Proc Natl Acad Sci USA. 2010;107(Suppl 2):8902–8909. doi: 10.1073/pnas.1001649107. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.