

Abstract
Mycoplasma genitalium (M.genitalium) is a sexually transmitted pathogen. Detection of this microorganism in clinical specimens by culture is rather difficult and time consuming. The aim of this study was to produce polyclonal antibody against a synthetic peptide from P110 protein of M.genitalium in order to develop a diagnostic tool for detection of this microorganism in clinical specimens. A synthetic peptide from P110 protein was conjugated to Keyhole Limpet Hemocyanin (KLH) and used for immunization of a white New Zealand rabbit. The produced antibody was purified by affinity chromatography and its specific interaction with immunizing peptide was determined by ELISA. Immunoreactivity of the antibody was also tested by Western blotting in bacterial cell lysate prepared from M.genitalium G-37. To confirm its application as a diagnostic tool, indirect immunofluorescent staining method was performed on M.genitalium-infected PBMC using anti-P110 as the primary antibody. The results showed that produced antibody has excellent reactivity with immunizing peptide and also detected a single band of 110 kDa corresponding to P110 protein. M.genitalium-infected PBMC showed a bright fluorescent signal in IF staining. This antibody might be used as a tool in diagnostic applications.
Keywords: Antibodies, Mycoplasma genitalium, Peptides
Introduction
Mycoplasma genitalium is a clinically important sexually transmitted pathogen that was first isolated in urethral specimens from two men with non-gonococcal urethritis by culture in 1981 (1). It is one of the major causes of non-gonococcal urethritis in men (2–4). Association between M.genitalium with chronic persistent prostatitis has been shown (5, 6). The capacity of this microorganism for malignant transformation of benign human epithelial cells has been reported (7). The association between M.genitalium and urethritis, cervicitis, endometritis, salpingitis and Pelvic Inflammatory Disease (PID) has also been shown in women (8–10). Detection of this microorganism in clinical specimens by culture is rather difficult and time consuming. MG192 (mgp C) gene (a part of the MgPa operon), which encodes an immunogenic and cyto-adherence related protein, designated as P110, is a highly variable within and among in vitro cultured strains and in vivo specimens. However there are some regions that do not undergo variation (11–13). In this study we designed and used a synthetic peptide derived from constant part of P110 protein to produce polyclonal antibody in order to develop a diagnostic tool for detection of M.genitalium in clinical specimens.
Materials and Methods
Peptide design and conjugation
A 16-mer synthetic peptide, sequencing NPGNDSLLSTTDNNIA, from constant part of P110 protein of M.genitalium was selected as immunogen. A cysteine residue was added to the C-terminus end of peptide to facilitate the conjugation to carrier protein. Immunograde peptide was purchased from Thermo Electron Corporation (GmbH, Ulm, Germany) and was conjugated to Keyhole Limpet Hemocyanin (KLH) and Bovine Serum Albumin (BSA), separately as described elsewhere (14). The peptide-KLH and peptide-BSA conjugates were used for immunization and conjugation assessment, respectively.
Confirmation of peptide conjugation by SDS-PAGE
To check the efficacy of conjugation, 10 µg of peptide-BSA conjugate was mixed with 10 µl of sample buffer and boiled for 5 min. Electrophoresis was performed on 10% SDS-PAGE condition using mini-PROTEAN electrophoresis instrument (Bio-Rad laboratories, Philadelphia, PA) with 100 mA for 1 hr. The gel was stained with Coomassie Blue R-250 (Sigma, St. Louis, MO). The change in mobility shift of conjugated BSA represented the efficiency of conjugation (Figure 1).
Figure 1.
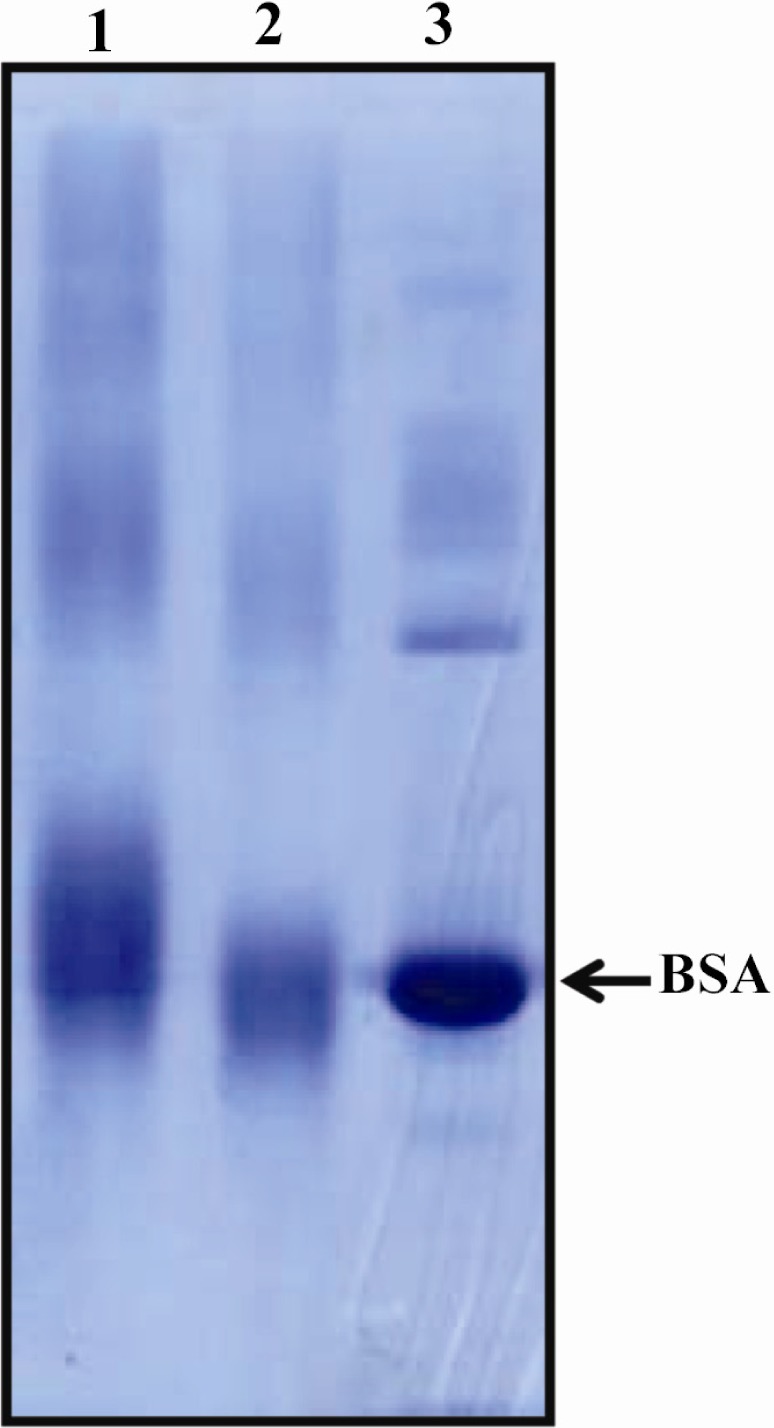
SDS-PAGE analysis of conjugated P110 peptide. Changing in mobility shift represents the efficacy of conjugation. Lane 1) Conjugated peptide-BSA. Lane 2) BSA-MBS (m-Maleimidobenzoyl- N -Hydroxysuccinimide Ester). Lane 3) BSA. Because of very high molecular weight of KLH, it is not possible to run the KLH conjugate on SDS-PAGE. In this context BSA conjugate was used for efficacy of conjugation
Immunization protocol
A female white New Zealand rabbit was immunized 3 times with one-week interval for each injection.
In first immunization, 100 µg KLH-peptide conjugate and 250 µl IMMACCEL (Pick cell Laboratories, Netherlands) was mixed with an equal volume of Freund's complete adjuvant (Sigma), and injected subcutaneously in 4-6 regions. For the subsequent immunizations, 500µg peptide-KLH and 250 µl IMMACCEL were admixed and injected with Freund's incomplete adjuvant (Sigma).
The last immunization was perfumed using 1000 µg peptide-KLH together with 250 µl IMMACCEL and Freund's incomplete adjuvant. The IMMACCEL reduces the antibody production time in rabbit from standard 80-day protocol to 28 day without any difference in affinity or specificity (15).
Titration of antibody
Before each immunization and 7 and 14 days after the last immunization, blood was drawn by venipuncture of the rabbit ear and allowed to clot for periods of 2 to 3 hr at room temperature before preparation of serum. Titration of the specific polyclonal antibody was then performed as follow: A96-well ELISA plate was coated with 100 µl of the immunizing peptide (20 mg/ml in PBS) at 37 °C for one hr followed by overnight incubation at 4 °C. The plate was washed 3 times with PBS containing 0.05% Tween 20 (PBS-T) for 5 min. The wells were then blocked with 2.5% BSA at 37 °C for 1.5 hr. Wells were washed 3 times as above and rabbit serum was added to the wells in two fold serial dilutions starting from 1:100. The plate was incubated at 37 °C for 1.5 hr and washed again with PBS-T.
At the next step, 100 µl of 1:1000 dilution of HRP-conjugated sheep anti-rabbit immuneglobulin (Avicenna Research Institute, Tehran, Iran) was added to the wells and incubation was continued for 1 hr, at 37 °C. After washing, 100 µl of Tetramethylbenzidine (TMB) chromogen was added to each well and the plate was incubated at room temperature in a dark place. After 15 min, the reaction was stopped by adding 30 µl of stopping solution (0.16 M H2So4) to each well. The Optical Density (OD) of the reaction was measured at 450 nm by an ELISA reader. Negative controls included omission of coating layer, serum (as primary antibody) or combination of both (Figure 2).
Figure 2.

Kinetic evaluation of anti-P110 antibody production in serum of immunized rabbit. A white New Zealand rabbit was immunized with peptide P110-KLH conjugate. The reactivity of 1:1000 diluted sera from immunized rabbit with immunizing peptide was determined at different time intervals by ELISA. The specific anti-body titer was upraised in immunized rabbit over the time and reached to the plateau after 28 days
Antibody purification
Rabbit serum was filtered through 0.45 µm filter and antibody was purified by affinity chromatography column prepared by coupling immunogenic peptide to SulfoLink Coupling Resin (Thermo Scientific). The elution was performed using 0.1M glycine. HCl (pH = 2.6). The pH of eluted antibody was adjusted to 7.0 with 1 MTris.HCl pH = 9.0. The eluted anti-body was dialyzed overnight against PBS pH = 7.5. The reactivity of the antibody was measured by ELISA and its purity evaluated by SDS-PAGE (Figures 3 and 4).
Figure 3.

SDS-PAGE analysis of purified anti-P110 antibody. Polyclonal anti-P110 antibody was produced in rabbit and purified over peptide affinity column. The purity of purified antibody was assessed by SDS-PAGE. Lane 1) Purified antibody. Lane 2) A mixture of human IgG and BSA for comparison
Figure 4.

Titration of purified rabbit anti-P110 polyclonal antibody with immunizing peptide. Anti-P110 polyclonal antibody was purified over peptide affinity column and its reactivity with immunizing peptide was titrated by ELISA
M.genitalium cultivation and lysate preparation
M.genitalium G-37 (a gift from Dr. Jaume Piñol, Institut de Biotecnología i Biomedicina Universitat Autònoma de Barcelona) was cultured in SP-4 medium (16). The color changing (reddish yellow) in medium after 48-72 hr was evident for mycoplasma growth. The surface-attached mycoplasmas were harvested by scraping off in PBS. The bacterial cell lysate was prepared by sonication in 500 µl of lysis buffer containing 1% Triton X-100, 50 mM Tris-HCl, pH = 7.4, 150 mM NaCl, 5 mM EDTA, 1% protease inhibitor cocktail (Sigma) and 10% phosphatase inhibitor (Roche Diagnostics GmbH, Penzberg, Germany) on ice. The protein concentration of lysate was measured by BCA protein assay kit (Thermo Scientific).
Western blotting
Twenty µg of M.genitalium lysate was run on a 10% SDS-PAGE (100 V for 2 hr). After electrophoresis, resolved proteins were transferred onto Immobilon-PVDF membrane (Millipore Corporation, USA). The membrane was blocked for overnight at 4 °C with 5% non-fat milk in PBS plus 0.05% Tween 20 (PBS-T). All antibody incubations were performed in PBS-T containing 3% non-fat milk. Filter was incubated with 10 µg/ml of anti-body 1.5 hr at room temperature. After extensive washing with PBS-T, the filter was incubated with peroxidase-conjugated sheep anti-rabbit immunoglobulin (Avicenna Research Institute) for 1 hr at room temperature followed by washing and developing with ECL chemiluminescence detection system (GE Healthcare, Uppsala, Sweden) (Figure 5).
Figure 5.

Western blot analysis of purified anti-P110 antibody. Bacterial cell lysate from M.genitalium G-37 was prepared by sonication and subjected to Western blot. Polyclonal anti-P110 antibody detected a specific band of 110 kDa. Lane 1) Protein marker (Fermentas). Lane 2) Reducing conditions. Lane 3) Non-reducing conditions
Immunofluorescent (IF) staining
The Peripheral Blood Mononuclear Cells (PBMC) from a healthy donor were separated by ficoll density gradient centrifugation (17). Freshly-prepared PMBC were cultured (2-3×104 cells/well) on an 8-well laminated glass slides (Paul Marienfeld GmbH & Co. KG, LaudaKönigshofen, Germany) in RPMI 1640 contain 10% FBS (Invitrogen, Carlsbad, CA) with subsequent incubation in moisturized and 5% CO2 conditions for overnight. The cultured and harvested M.genitalium was dispersed through 25-gauge needle and passed through 0.45 µm filter (18) followed by infecting the isolated PBMC. The infected cells were seeded on glass slide and incubated at 37 °C for 3 hr. The slides were washed by Tris-Buffered Saline (TBS), pH = 7.4, air-dried at room temperature for 15 min, followed by permeabilization by acetone (at -20 °C) for 2 min. The slides were kept at 4 °C for 30 min for drying. As negative control, few PBMC-cultured slides, were prepared without infecting by M.genitalium.
The uninfected and M.genitalium-infected slides were washed with TBS containing 0.1% bovine serum albumin (TBS-BSA) for three times, each 3 min. Slides were then blocked with 5% sheep serum for 10 min at room temperature. The primary anti-P110 antibody was diluted in TBS-BSA to a final concentration of 10 µg/ml and added to the slides. Slides were incubated at room temperature for 60 min followed by three times (3×3 min) washing steps with TBS-BSA. Slides were incubated at room temperature with Fluorescein Isothiocyanate (FITC)-conjugated sheep anti-rabbit polyclonal antibody (1:100 TBS-BSA) (Avicenna Research Institute) for 45 min. After washing with TBS-BSA, the nuclei were counterstained by 4’,6-diamidino-2-phenylindole dihydrochloride (D API) (Sigma) at concentration of 1 µg/ml for 5 min, then the slides were washed, mounted in TBS-glycerol 80% and examined under a fluorescence microscope (Olympus, Tokyo, Japan) (Figure 6).
Figure 6.
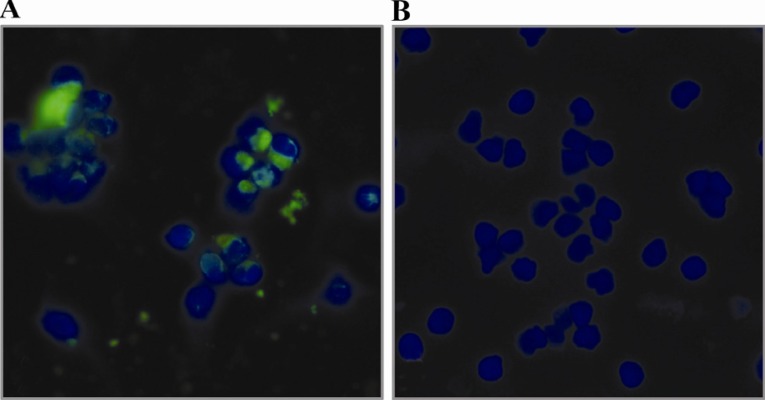
Immunofluorescent staining of M.genitalium infected PBMC using purified anti-P110 antibody. PBMC from a human healthy individual was infected with M.genitalium and the presence of organism in infected cells was monitored by indirect immunofluorescent staining using polyclonal anti-P110 antibody 3 hr after infection. DAPI was used for staining the nucleus (blue). The green speckled fluorescence signal shows the presence of M.genitalium in infected PBMC (A); Uninfected PBMC served as negative control (B)
Results
Peptide conjugation and antibody titration
Proper conjugation of peptide to carrier protein was assessed by SDS-PAGE electrophoresis. The change in mobility shift of peptide-BSA conjugate in SDS-PAGE gel represented the efficiency of conjugation (Figure 1). The presence of antibody against immunizing peptide in rabbit serum was evaluated by ELISA before and after immunization. Results indicated the presence of anti-body against immunizing peptide in rabbit serum which was upraised considerably and reached to the plateau at day 28 post immunization (Figure 2).
Evaluation of purified antibody
SDS-PAGE analysis of the purified anti-body revealed the presence of a single band of about 150 kDa indicative of the desired purity (Figure 3). Purified antibody exhibited excellent immunoreactivity with immunizing peptide in ELISA assay as well as suggesting its functionality (Figure 4).
Western blot analysis of anti-P110 antibody
The reactivity of the purified antibody to its corresponding native protein was determined by Western blot. The produced antibody recognized a single band of 110 kDa representtative of P110 protein of M.genitalium G-37 both at reducing and non-reducing conditions (Figure 5).
Immunofluorescence staining of M.genitalium-infected PBMC
To analyse the capability of produced anti-body to recognize M.genitalium in infected cells, indirect immunofluorescent assay was performed. Infected cells showed a bright speckled fluorescent signal located mainly in the cytoplasm of the cells 3 hr after infection (Figure 6).
Discussion
The association between M.genitalium and infections in men and women has been shown since its discovery in 1981 (19). Most mycoplasmas can grow in special culture media but M.genitalium grows rather difficult (20). Therefore, other diagnostic methods must be sought.
Antibodies could be produced virtually against any desired compound. The first step in the production of antibody is selection of a proper target. The decision about producing poly-or monoclonal antibodies depends on the application. The production of polyclonal antibodies is faster, less expensive, and less technically demanding than monoclonal anti-bodies. Monospecificity of monoclonal anti-bodies may limit their applications as the reactivity of these antibodies can be affected by a small change in the structure of an epitope including genetic polymorphism, glycolsylation, and denaturation. Monoclonal anti-bodies are also highly susceptible to small changes in pH and salt concentration. Modification of antibodies by covalently linking to a fluorochrome or radionuclide may alter their binding efficacy. The polyclonal antibodies bypass such problems with their reactivity to the vast variety of epitopes (21).
Seemingly, there is not any commercial antibody against M.genitalium available in the market. We produced polyclonal antibody against a synthetic peptide derived from the constant part of P110. There are several studies found in the literature indicating production of poly-or monoclonal antibodies against whole cell or recombinant proteins of M.genitalium for different purposes (22–26). The produced antibody showed specific interaction with immunogenic peptide by ELISA and it also recognized native P110 protein by Western blot and IF assay.
As designed peptide has been selected from the constant part of P110 protein, antigenic variations in M.genitalium do not impede the reactivity of this antibody. No cross-reactivity of this antibody with human cell lysates in both Western blot and immunofluorescence applications increases the specificity impact of this antibody making it suitable for detecting of this microorganism in clinical specimens. A clear example would be using urine sample from the infected patients for diagnostic IF assay.
Conclusion
Although sequence of this immunogenic peptide has theoretically no similarity to any other mycoplasma strains, the cross reactivity with other mycoplasma strains should be investigated in practice. Further inhibition assay studies are warranted to evaluate the capability of this antibody in preventing the infection of eukaryotic cells. In the case of functionality of the produced polyclonal anti-body in inhibition assay, this peptide and the epitopes thereof could be used for production of monoclonal antibody for potential therapeutic purposes.
Acknowledgement
This work was supported by a grant from Avicenna Research Institute (ARI), Tehran, Iran and Tehran University of Medical Sciences, Tehran, Iran. We would like to express our gratitude to Dr. Jaume Piñol, Institut de Biotecnología i Biomedicina Universitat Autònoma de Barcelona, Spain for providing us the M.genitalium G-37 strain as well as great theoretical assistance in optimizing the culturing conditions.
References
- 1.Tully JG, Cole RM, Taylor-Robinson D, Rose DL. A newly discovered mycoplasma in the human urogenital tract. Lancet. 1981;317(8233):1288–1291. doi: 10.1016/s0140-6736(81)92461-2. [DOI] [PubMed] [Google Scholar]
- 2.Tully JG, Taylor-Robinson D, Rose DL, Cole RM, Bove JM. Mycoplasma genitalium, a new species from the human urogenital tract. Int J Syst Bacteriol. 1983;33:387–396. [Google Scholar]
- 3.Jensen JS, Orsum R, Dohn B, Uldum S, Worm AM, Lind K. Mycoplasma genitalium: a cause of male urethritis? Genitourin Med. 1993;69(4):265–269. doi: 10.1136/sti.69.4.265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Totten PA, Schwartz MA, Sjostrom KE, Kenny GE, Handsfield HH, Weiss JB, et al. Association of Mycoplasma genitalium with nongonococcal urethritis in heterosexual men. J Infect Dis. 2001;183(2):269–276. doi: 10.1086/317942. [DOI] [PubMed] [Google Scholar]
- 5.Kreiger JN, Riley DE. Chronic prostatitis: Charlottesville to Seattle. J Urol. 2004;172(6):2557–2560. doi: 10.1097/01.ju.0000144291.05839.a0. [DOI] [PubMed] [Google Scholar]
- 6.Mandar R, Raukas E, Turk S, Korrovits P, Punab M. Mycoplasmas in semen of chronic prostatitis patients. Scand J UrolNephrol. 2005;39(6):479–482. doi: 10.1080/00365590500199822. [DOI] [PubMed] [Google Scholar]
- 7.Namiki K, Goodison S, Porvasnik S, Allan RW, Iczkowski KA, Urbanek C, et al. Persistent exposure to Mycoplasma induces malignant transformation of human prostate cells. PloS One. 2009;4(9):6872. doi: 10.1371/journal.pone.0006872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Falk L, Fredlund H, Jensen JS. Signs and symptoms of urethritis and cervicitis among women with or without Mycoplasma genitalium or Chlamydia trachomatis infection. Sex Transm Infect. 2005;81:73–78. doi: 10.1136/sti.2004.010439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Taylor-Robinson D. Mycoplasma genitaliuman update. Int J STD AIDS. 2002;13(3):145–151. doi: 10.1258/0956462021924776. [DOI] [PubMed] [Google Scholar]
- 10.Cohen CR, Mugo NR, Astete SG, Odondo R, Manhart LE, Kiehlbauch JA, et al. Detection of Mycoplasma genitalium in women with laparoscopically diagnosed acute salpingitis. Sex Transm Infect. 2005;81:463–466. doi: 10.1136/sti.2005.015701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ma L, Jensen JS, Myers L, Burnett J, Welch M, Jia Q, et al. Mycoplasma genitalium: an efficient strategy to generate genetic variation from a minimal genome. Mol Microbiol. 2007;66(1):220–236. doi: 10.1111/j.1365-2958.2007.05911.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Iverson-Cabral SL, Astete SG, Cohen CR, Totten PA. mgpB and mgpC sequence diversity in Mycoplasma genitalium is generated by segmental reciprocal recombination with repetitive chromosomal sequences. Mol Microbiol. 2007;66(1):55–73. doi: 10.1111/j.1365-2958.2007.05898.x. [DOI] [PubMed] [Google Scholar]
- 13.Ma L, Jensen JS, Mancuso M, Hamasuna R, Jia Q, McGowin CL, et al. Genetic variation in the complete MgPa operon and its repetitive chromosomal elements in clinical strains of Mycoplasma genitalium. PLoS One. 2010;5(12):15660. doi: 10.1371/journal.pone.0015660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hadavi R, Zarnani AH, Ahmadvand N, Mahmoudi AR, Bayat AA, Mahmoudian J, et al. Production of monoclonal antibody against human nestin. Avicenna J Med Biotech. 2010;2(2):69–77. [PMC free article] [PubMed] [Google Scholar]
- 15.Litvinov S. New adjuvant for accelerated and enhanced antibody response. Nature Methods. 2009;6:10–11. [Google Scholar]
- 16.Tully JG, Rose DL, Whitcomb RF, Wenzel RP. Enhanced isolation of Mycoplasma pneumoniae from throat washings with a newly-modified culture medium. J Infect Dis. 1979;139(4):478–482. doi: 10.1093/infdis/139.4.478. [DOI] [PubMed] [Google Scholar]
- 17.Ferrante A, Thong YH. Optimal conditions for simultaneous purification of mononuclear and polymorphonuclear leucocytes from human blood by the hypaque-ficoll method. J Immunol Methods. 1980;36(2):109–117. doi: 10.1016/0022-1759(80)90036-8. [DOI] [PubMed] [Google Scholar]
- 18.Ueno PM, Timenetsky J, Centonze VE, Wewer JJ, Cagle M, Stein MA, et al. Interaction of Mycoplasma genitalium with host cells: evidence for nuclear localization. Microbiology. 2008;154(10):3033–3041. doi: 10.1099/mic.0.2008/020735-0. [DOI] [PubMed] [Google Scholar]
- 19.Jones L, Felblinger D, Cooper L. Mycoplasma genitalium: more prevalent than you think. Nurse Pract. 2009;34(8):50–52. doi: 10.1097/01.NPR.0000358664.73596.4c. [DOI] [PubMed] [Google Scholar]
- 20.Manhart LE, Kay N. Mycoplasma genitalium: Is it a sexually transmitted pathogen? Curr Infect Dis Rep. 2010;12(4):306–313. doi: 10.1007/s11908-010-0114-3. [DOI] [PubMed] [Google Scholar]
- 21.Lipman NS, Jackson LR, Trudel LJ, Weis-Garcia F. Monoclonal versus polyclonal antibodies: distinguishing characteristics, applications, and information resources. ILAR J. 2005;46(3):258–268. doi: 10.1093/ilar.46.3.258. [DOI] [PubMed] [Google Scholar]
- 22.Clausen HF, Fedder J, Drasbek M, Nielsen PK, Toft B, Ingerslev HJ, et al. Serological investigation of Mycoplasma genitalium in infertile women. Hum Reprod. 2001;16(9):1866–1874. doi: 10.1093/humrep/16.9.1866. [DOI] [PubMed] [Google Scholar]
- 23.Svenstrup HF, Fedder J, Abraham-Peskir J, Birkelund S, Christiansen G. Mycoplasma genitalium attaches to human spermatozoa. Hum Reprod. 2003;18(10):2103–2109. doi: 10.1093/humrep/deg392. [DOI] [PubMed] [Google Scholar]
- 24.Svenstrup HF, Nielsen PK, Drasbek M, Birkelund S, Christiansen G. Adhesion and inhibition assay of Mycoplasma genitalium and M. pneumoniae by immunofluorescence microscopy. J Med Microbiol. 2002;51(5):361–373. doi: 10.1099/0022-1317-51-5-361. [DOI] [PubMed] [Google Scholar]
- 25.Burgos R, Pich O, Ferrer Navarro M, Baseman J, Querol E, Pinol J. Mycoplasma genitalium P140 and P110 cytadhesinae reciprocally stabilizaed and required for cell adhesion and terminal organelle development. J Bacteriol. 2006;188(24):8627–8637. doi: 10.1128/JB.00978-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Opitz O, Jacobs E. Adherence epitopes of Mycoplasma genitalium adhesin. Microbiology. 1992;138(9):1785–1790. doi: 10.1099/00221287-138-9-1785. [DOI] [PubMed] [Google Scholar]