

Abstract
The coupling of environmental sensing to flagella-mediated directed motility allows bacteria to move to optimum environments for growth and survival, either by sensing external stimuli (chemotaxis) or monitoring internal metabolic status (energy taxis). Sensing is mediated by transducer-like proteins (Tlp), either located in the membrane or in the cytoplasm, which commonly influence motility via the CheA-CheY chemotaxis pathway. In this study we have investigated the role of PAS-domain-containing intracellular Tlp-sensors in energy taxis of the food-borne pathogen Campylobacter jejuni, using plate- and tube-based assays utilising the conversion of the redox indicator dyes triphenyl tetrazolium chloride (TTC) and resazurin. Inactivation of the genes encoding the Campylobacter Energy Taxis system (CetA (Tlp9) and CetB (Aer2)) in C. jejuni strain NCTC 11168 resulted in reduced taxis. Inactivation of the cj1191c gene, encoding the CetB homolog CetC (Aer1), did not affect taxis per se, but the cetC gene complemented a cetB mutant in trans, indicating that CetC can form a functional signal transduction complex with CetA in the absence of CetB. Inactivation of both CetB and CetC resulted in greatly reduced taxis confirming the role of CetC in energy taxis. Inactivation of the cj1110c gene, encoding Tlp8 (CetZ), a cytoplasmic sensor with two PAS-domains, resulted in increased taxis, a phenotype opposite to that of CetAB. Inactivation of the cheA gene resulted in the same overall phenotype as the cetAB mutant in both wild-type and cetZ backgrounds, suggesting that both systems use the CheA system for signal transduction. Absence of both CetAB and CetZ resulted in the cetAB taxis phenotype, suggesting that CetZ is subordinate to CetAB. In conclusion, we present evidence that C. jejuni balances the input from two counteracting PAS-domain-containing sensory systems to position itself for optimal usage of energy resources.
Introduction
Bacteria are well adapted to survive in various niches, but long-term survival will often require transit through stressful environments. Consequently, bacteria have adopted a variety of mechanisms to sense and respond to these environmental stresses. Long-term genetic adaptations are possible by horizontal gene transfer, genomic deletions or reorganisation, or through random mutation and selection [1]. Relatively short-term adaptations are possible by sensing environmental changes and modulating gene expression, as typified by the two-component regulatory systems [2]. However, the most rapid adaptation a motile bacterium can make is to use taxis-controlled motility to move away from stress or move towards a more favourable locale [3].
Bacterial taxis encompasses sensing a specific environmental or internal signal, which triggers a signal transduction cascade, resulting in changing the direction of flagella rotation to affect a change in swimming direction. The paradigm of bacterial taxis is the chemotaxis system characterised in Escherichia coli [4]. Specifically, activation of a signal receptor results in auto-phosphorylation of CheA, with subsequent phosphotransfer to CheY. Phospho-CheY interacts with the flagella motor proteins, which alters the direction of flagella rotation from counter-clockwise (swimming phenotype) to clockwise rotation (tumbling phenotype). Accordingly, the population of bacteria change direction, albeit in a somewhat stochastic manner [5].
While the CheA-CheY phospho-relay pathway serves as the ‘master’ switch controlling taxis, the task of sensing a diverse set of environmental signals falls to a group of sensing proteins termed either methyl-accepting chemotaxis proteins (MCPs) [6] or transducer-like proteins (Tlps) [7]. Such sensing systems give bacteria the ability to exhibit taxis responses to gradients of small molecules including nutrients (chemotaxis), pH (pH taxis), light (phototaxis), magnetic fields (magnetotaxis), and intracellular energy status (aerotaxis, redox taxis, or energy taxis) [3]. Energy taxis allows bacteria to swim to environments that are optimal for energy generation, whether that means sensing an optimal oxygen concentration, appropriate electron acceptors, light, or redox-active compounds [8], [9]. For example, the E. coli aerotaxis sensor Aer mediates taxis to optimum oxygen concentrations through direct interaction with the respiratory chain [10], whereas the Azospirillum brasilense AerC protein is a redox sensor that mediates taxis to microaerobic conditions optimal for nitrogen fixation [11]. One of the protein domains associated with energy taxis is the PAS (Per-ARNT-Sim) domain, which is the most common sensing domain present in bacterial MCP proteins [6]. The PAS domain is found in all kingdoms of life, and has been described to be involved in sensing diverse stimuli including light, oxygen, redox potential, small metabolites, and proton-motive force [12], and is often involved in energy taxis [8], [9].
Many bacterial pathogens require directed motility for infection and accordingly possess functional taxis systems; the human pathogen Campylobacter jejuni is no exception. C. jejuni is leading cause of foodborne illness in the developed world (over 60,000 confirmed cases annually in the UK [13]), and is most often associated with the consumption or handling of under-cooked poultry products [14]. Despite its relatively small size, the C. jejuni genome encodes ten proteins containing MCP-domains [7], which is considerably more than the five, including Aer, found in E. coli. Indeed, based on the Pfam database [15], the MCP domain is the fifth most prevalent domain in C. jejuni. This is similar to the average number of MCPs predicted in bacterial genomes, but considerably more than the average number found in pathogens and commensals [6]. For C. jejuni, formate and asparate sensors have been identified [16], [17], as well as a proposed energy taxis system, CetAB (Campylobacter energy taxis proteins A and B) [18]. The CetAB system consists of two co-transcribed open reading frames [19] encoding a discrete membrane-bound MCP protein and cytoplasmic PAS-domain protein (CetA (Tlp9) and CetB (Aer2), respectively), which is atypical for such sensors, as these usually consist of a single protein with a modular architecture containing a C-terminal MCP domain, and an N-terminal ligand binding domain (LBR) [6].
Inactivation of cetA and cetB in C. jejuni strain 81–176 resulted in decreased motility in defined agar plates supplemented with pyruvate and fumarate [18]. The CetA and CetB proteins were further shown to directly interact, with CetB being associated with membrane-bound CetA [19]. Based on the structure and genetic organization of the cetAB genes, these have been proposed to constitute a new class of bipartite energy taxis systems, with a number of such systems found outside the genus Campylobacter [20].
In addition to cetAB, the C. jejuni genome contains two other genes encoding PAS-domain containing proteins implicated in taxis: cj1110c (tlp8), which encodes an MCP protein which contains two predicted PAS domains, and cj1191c (aer1), which encodes a homolog of CetB and contains a single predicted PAS domain. In this study, we have investigated the role of the PAS- and MCP-domain genes cetAB, cj1191c and cj1110c in C. jejuni energy taxis. We have used genetic mutation and complementation analyses, swarm plate assays, and a newly developed tube taxis assay utilising redox dyes to show that the CetAB system is required for efficient energy taxis in a redox gradient. We demonstrate that Cj1191c is involved in energy taxis, and further show that Cj1110c is a sensor involved in a CheA-dependent redox taxis response that is opposed, but subordinate to the response mediated by the CetAB system. These results suggest that Campylobacter has the ability to balance redox and energy status signals, via PAS-domain linked signal transduction, to mediate taxis to an optimal environment.
Materials and Methods
C. jejuni strains and growth conditions
Campylobacter jejuni strain NCTC 11168 and its isogenic mutants (Table 1)were routinely cultured in a MACS-MG-1000 controlled atmosphere cabinet (Don Whitley Scientific) under microaerobic conditions (85% N2, 5% O2, 10% CO2) at 37°C [21]. For growth on plates, strains were either grown on Brucella agar, blood plates (Blood Agar Base 2 (BAB), 1% yeast extract, 5% horse blood (Oxoid)), or BAB with Skirrow supplements (10 µg ml−1 vancomycin, 5 µg ml−1 trimethoprim, 2.5 IU polymyxin-B). Broth culture was carried out in Brucella broth (Becton Dickinson). An Innova 4230 (New Brunswick Scientific) incubator was used for aerobic culture at 37°C and a Sanyo MCO-20 AIC incubator was used for culture in 10% CO2 in air at 37°C.
Table 1. Bacterial strains used in this study.
| Strain | Descriptiona |
| E. coli strains | |
| Top10 | General cloning strain |
| C. jejuni strains | |
| NCTC 11168 | Wild-type C. jejuni [46] |
| 11168 ΔcetA | NCTC 11168 cj1190c::kanR |
| 11168 ΔcetB | NCTC 11168 cj1189c::kanR |
| 11168 ΔcetAB | NCTC 11168 (cj1189-90c)::kanR |
| 11168 ΔcetC | NCTC 11168 cj1191c::kanR |
| 11168 ΔcetZ | NCTC 11168 cj1110c::catR |
| 11168 ΔcheA | NCTC 11168 cj0284c::kanR |
| 11168 ΔcheA ΔcetZ | NCTC 11168 cj0284c::kanR cj1110c::catR |
| 11168 ΔflaAB | NCTC 11168 (cj1338-39c)::kanR |
| 11168 ΔcetA ΔcetZ | NCTC 11168 cj1190c::kanR cj1110c::catR |
| 11168 ΔcetB ΔcetZ | NCTC 11168 cj1189c::kanR cj1110c::catR |
| 11168 ΔcetAB ΔcetZ | NCTC 11168 (cj1189-90c)::kanR cj1110c::catR |
| 11168 ΔcetB ΔcetC | NCTC 11168 cj1189c::kanR cj1191c::catR |
| 11168 ΔcetA::cetAfdxApr* | NCTC 11168 cj1190c::kanR cj0046::cetAfdxApr* catR |
| 11168 ΔcetA::cetA cetApr * | NCTC 11168 cj1190c::kanR cj0046::cetA cetApr * catR |
| 11168 ΔcetA::cetABfdxApr* | NCTC 11168 cj1190c::kanR cj0046::cetABfdxApr* catR |
| 11168 ΔcetB::cetABfdxApr* | NCTC 11168 cj1189c::kanR cj0046::cetABfdxApr* catR |
| 11168 ΔcetAB::cetABfdxApr* | NCTC 11168 (cj1189-90c)::kanR cj0046::cetABfdxApr* catR |
| 11168 ΔcetB::cetB * | NCTC 11168 cj1189c::kanR cj0046::cetBfdxApr* catR |
| 11168 ΔcetB::cetC * | NCTC 11168 cj1189c::kanR cj0046::cj1191cfdxApr* catR |
| 11168 ΔcetAB::cetA-cetB * | NCTC 11168 (cj1189-90c)::kanR cj0046::cetA-cetBfdxApr* catR |
| 11168 ΔcetAB::cetA-cetC * | NCTC 11168 (cj1189-90c)::kanR cj0046::cetA-cj1191cfdxApr* catR |
| 11168 ΔcetZ::cetZ * | NCTC 11168 cj1110c::catR cj0046::cj1110cfdxApr* kanR |
kanR, kanamycin antibiotic resistance cassette; catR, chloramphenicol antibiotic resistance cassette;
denotes genetic complementation; fdxApr, gene under control of the fdxA promoter, cetApr, gene under control of the cetA promoter. Superscript text denotes the promoter used in the complementation cassette.
Construction of cetA (cj1190c), cetB (cj1189c), cetAB, cetC (cj1191c), cetZ (cj1110c), and cheA (cj0284c) mutants
Plasmids used in this study are listed in Table S1, whereas primers used are listed in Table S2. DNA fragments of the target gene and approximately 500 bp of flanking sequence on each side was PCR amplified using Phusion DNA polymerase (New England Biolabs) using the primers detailed in Table S2. These amplified fragments were purified (QIAgen PCR purification kit), digested and ligated into pNEB193 (New England Biolabs) also digested with the corresponding restriction endonucleases (see Table S2) and transformed into chemically competent E. coli strain Top10 (Invitrogen). Constructs containing an insert were selected on LB agar plates containing 100 µg ml−1 carbenicillin and 20 µg ml−1 X-gal. Constructs containing an insert were confirmed by restriction digest analysis and sequencing (TGAC, Norwich, UK). To make the ΔcetA, ΔcheA, and ΔcetC (Δcj1119c) insertional inactivation constructs, the BamHI-digested kanamycin cassette from pMARKan9 [22] was inserted into the naturally occurring BamHI and BglII sites in the cetA gene, the naturally occurring BamHI site in cheA, or the naturally occurring BglII site of cj1191c. Similarly, the BamHI-digested chloramphenicol cassette from pAV35 was ligated into the naturally occurring BglII site of cj1191c. To make the ΔcetB and ΔcetAB insertional-inactivation constructs, inverse-PCR primers (see Table S2) were used to amplify the regions flanking the target genes. These purified PCR products were digested with either BglII or BamHI, purified and ligated to the BamHI ends of the kanamycin cassette. To make the ΔcetZ (Δcj1110c) insertional-inactivation construct, a chloramphenicol cassette [22] with BamHI ends was ligated into the naturally occurring BglII site in cetZ. All ligation reactions were transformed into E. coli strain Top10 and positive transformants were selected for by plating on LB agar supplemented with 30 µg ml−1 kanamycin or 30 µg ml−1 chloramphenicol as appropriate. Plasmids with insert, and insert orientation, were verified with restriction digest analysis and sequencing. C. jejuni mutant strains were isolated after transformation of the C. jejuni NCTC 11168 wild-type strain with the plasmids by electroporation [23], followed by selection on plates with kanamycin, chloramphenicol or both antibiotics. Colonies were screened by PCR using primers that anneal outside of the cloned flanking regions in combination with antibiotic cassette-specific primers (Table S2).
Construction of cetA, cetB, cetC, cetZ, cetAB, cetA-cetB chimera and cetA-cetC chimera complementation constructs
C. jejuni mutants were complemented by supplying the genes of interest in trans using a cj0046 pseudogene-based delivery system [24]. To make the cetA complementation constructs (expressed from either the fdxA or native cetA promoter), the cetA gene was PCR-amplified (for primers see Table S2), digested with NcoI, and ligated into either pCfdxA or pC46 digested with Esp3I (Fermentas). To make the cetB complementation construct, the cetB gene was PCR-amplified (see Table S2), digested with PciI, and ligated into pCfdxA digested with Esp3I. To make the cetC complementation construct, the cetC gene was PCR-amplified (see Table S2), digested with BspHI, and ligated into pCfdxA digested with Esp3I. To make the cetAB complementation construct, a PCR-fragment was generated containing cetB and a C-terminal region of cetA encompassing a unique BamHI site. This PCR fragmented was digested with BamHI and PciI and ligated into plasmid pCASO12 (cetA-fdxApr) sequentially digested with BamHI and Esp3I, thus adding the cetB gene downstream of cetA. To make the cetA-cetB and cetA-cetC chimeric complementation constructs, the cetB or cetC PCR fragments, digested with either PciI or BspHI, were ligated into pCASO12 (see Table S1) digested with Esp31. To make the cetZ complementation construct, the cetZ gene was PCR-amplified (see Table S2), digested with PciI, and ligated into pKfdxA digested with Esp31. These complementation constructs were transformed into either the wild-type strain or the appropriate mutant background using standard electroporation methods. Complementation strains were confirmed by PCR using primers that anneal outside of the cj0046 flanking regions in combination with gene-specific primers (see Table S2).
Growth Curves
A 50 µl single-use glycerol stock, routinely stored at −80°C, was plated on a BAB plate with Skirrow supplements and these cells were used to inoculate fresh Brucella broth. Cultures were grown in microaerobic conditions with shaking overnight at 37°C. The overnight culture was diluted to A600≈0.05 (∼1×107 CFU ml−1) in 50 ml fresh Brucella broth in a T75 filter screw cap tissue culture flask (TPP, Helena Biosciences) and grown in microaerobic conditions with shaking at 37°C. The A600 was monitored every 60 minutes for up to ten hours.
Autoagglutination assays and Light Microscopy
Autoagglutination (i.e. cell clumping and sedimentation) was measured as described previously by monitoring the decrease in A600 following incubation in a cuvette at room temperature in air [25]. For monitoring motility using light microscopy, 10 µl of overnight culture, mounted between a glass slide and coverslip, was examined using an Eclipse 50i microscope and videos acquired using a Coolpix 4500 digital camera (Nikon).
Tube taxis assay
Tube taxis assays were prepared by mixing 0.4 g agar with 100 ml Brucella broth, followed by autoclaving. Media was cooled to ∼50°C and 1 ml of 1% filter sterile 2,3,5-triphenyltetrazolium chloride (TTC, Sigma) was added to the media. To make Resazurin tubes, 500 µl of 0.02% resazurin was added to 100 ml Brucella broth containing 0.4 g agar prior to autoclaving. After cooling to ∼45°C, 10 ml of media was added to sterile 15 ml Falcon tubes (Corning) and allowed to set. If not used directly, tubes were stored at 4°C for up to one month. A 50 µl single-use glycerol stock was plated on a BAB plate with Skirrow supplements, grown for 24 hours, and cells resuspended in 2 ml sterile PBS. The top of the agar was inoculated with 50 µl of this cell suspension. The tubes were re-capped and incubated at 37°C in either: air, 2% O2/10% CO2/88% N2, 5% O2/10% CO2/85% N2, or in air/10% CO2. For culture in 2% O2/10% CO2/88% N2 and 5% O2/10% CO2/85% N2, tubes were incubated in gas jars containing a modified atmosphere supplied by an Anoxomat AN2CTS (Mart Microbiology B.V). For incubation in air/10% CO2, tubes were incubated in a Sanyo MCO-20AIC incubator. Tubes were photographed after 24, 40, 48, 64, and 72 hours. The dye front was measured from the top of the agar using the ImageJ software and expressed as a percentage of the wild-type strain. Each strain was tested for significance using a one-sample t-test compared to a hypothetical value of 100 (GraphPad Prism 5.01).
Swarm plate assays
All swarm plate assays were carried out using square 10 mm2 petri plates (Sterilin) inoculated with wild-type and three to four test strains. Swarming in rich media was assessed using Brucella broth supplemented with 0.01% TTC and 0.4% agar (Oxoid). Energy taxis assays were modified from a previous study [18], by replacing the chemically defined medium with the cell culture media DMEM and MEMα (Life Technologies). DMEM supplemented with 0.4% (w/v) agar was autoclaved and allowed to cool to ∼50°C. This medium was supplemented with 0.01% (w/v) TTC and 5% (v/v) MEMα medium (DMEM/MEMα). This medium was poured directly into plates and allowed to set for 24 hours at room temperature prior to inoculation. Plates were inoculated with 5 µl of overnight culture, and incubated in microaerobic conditions at 37°C. For any given assay, Brucella and DMEM/MEMα plates were set-up concurrently using the same overnight culture. Following inoculation, the number of viable cells in the inoculum was assayed using standard plating methods: using 96 well plates, cells were serially diluted to 1×10−8 then 5 µl of each dilution was spotted onto a Brucella plate. Swarm plates were photographed using a CCD camera and gel documentation system (U:Genius, Syngene) after 24 and 48 hrs and halo area measured using ImageJ (version 1.41; National Institute of Health [http://rsbweb.nih.gov/ij/]). For each plate, halo size was expressed as a percentage of the corresponding wild-type and each strain was tested for significance using a one-sample t-test, compared to a hypothetical value of 100 (GraphPad Prism 5.01).
Chemotaxis assays
Chemotaxis assays were adapted from [26]. A cell suspension was made by resuspending cells growing on a Skirrow plate in 2 ml sterile PBS. Two hundred µl of cell suspension was mixed with 30 µl 1% TTC (filter sterile), and 3 ml 0.4% agar/PBS and quickly mixed by hand before pouring on a sterile Brucella plate. Flame-sterilized tweezers were used to place a filter paper disc (6 mm Grade AA, Whatman) on the plate. Ten µl of either 1 M L-fucose (filter sterile) or PBS (negative control) was pipetted onto the filter discs. Plates were incubated in microaerobic conditions and halos visually inspected after 24 hours.
Results
The CetAB system is required for efficient swarming motility
To investigate cet-dependent energy taxis in C. jejuni, single cj1190c (cetA), cj1189c (cetB) and double cetAB mutants were constructed by replacing the genes (Fig. 1) with an antibiotic cassette (see Materials and Methods). These mutants showed no growth defects in Brucella broth, either by A600 in Brucella broth or viable counts (data not shown). Cells exhibited normal swimming behaviour, as assessed using light microscopy, showed wild-type autoagglutination, and possessed bi-polar flagella as shown using microscopy. However, in a standard swarming motility assay in Brucella soft agar, the ΔcetA and ΔcetB mutants showed a swarming defect (40% and 50% of wild-type respectively, Fig. 2 and Fig. 3A), with the ΔcetAB double mutant displaying most pronounced defect (30% of wild-type, Fig. 2 and Fig. 3A). However, these phenotypes were less severe than a non-motile ΔflaAB mutant, where halos were 10% that of the wild-type (Fig. 2, and Fig. 3A). Assays were conducted with the wild-type strain and the ΔcetAB strain in the presence and absence of the redox indicator TTC (2,3,5-triphenyltetrazolium chloride [27]), which is reduced by C. jejuni to TPF (1,3,5-triphenylformazan), a red insoluble precipitate. No difference in the swarming phenotype was observed with or without TTC (data not shown). The presence of TTC in the agar plate aided visualisation and documentation of the halo, so all subsequent assays were performed in media supplemented with TTC at a final concentration of 0.01%.
Figure 1. Genetic and domain organization of the cetC, cetAB and cetZ genes and encoded proteins in C. jejuni NCTC 11168, respectively.
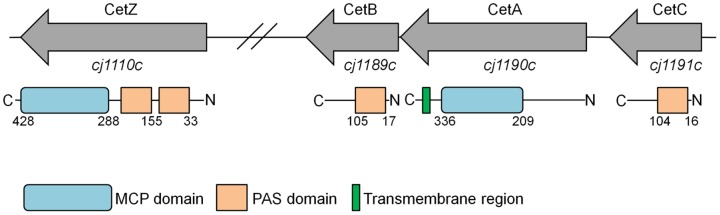
Numbers in the domain diagram show the regions of alignment according to Pfam identification.
Figure 2. Representative swarming and taxis assays.

Representative swarming plates showing wild-type, ΔcetA, ΔcetB, ΔcetAB and ΔflaAB strains after 24 hours incubation at 37°C in microaerobic conditions in: A) Brucella (0.4% agar, 0.01% TTC) and B) DMEM supplemented with 5% MEMα (0.4% agar, 0.01% TTC). C) Representative composite image of parallel TTC- and resazurin-based tube assays. Brucella soft agar was supplemented with either 0.01% TTC (T) or 0.0001% resazurin (R) and were incubated after inoculation at the top at 37°C in air and photographs taken at the times indicated. Asterisks are used to show the edge of the dye front. D) DMEM/MEMα swarming assay showing halo formation of the wild-type, ΔcheA, ΔcetZ, and ΔcetZ::cetZ* strains. E) Representative tube assay after 40 hrs incubation in air (37°C) showing migration of wild-type, ΔcetA, ΔcetB, ΔcetAB, ΔcetZ (cj1110c), ΔcetA ΔcetZ, ΔcetB ΔcetZ, and ΔcetAB ΔcetZ strains. Asterisks are used to show the edge of the dye front.
Figure 3. CetABC and CetZ play opposing roles in energy taxis.

A) Results from swarming assays in Brucella (light grey) and DMEM/MEMα media (dark grey). B) Initial 24 hour (white) and final (black) 72 hour time points from the tube assay. Strain designations are shown on the Y-axis. All data are expressed as a percentage of the wild-type and error bars show the standard deviation from at least three biological replicates. Asterisks denote statistically significant results compared to the wild-type (one-sample t-test, p<0.05).
Redox dyes can be used to visualise and track taxis and motility of C. jejuni
To investigate the effect of selected C. jejuni mutants in energy taxis, two different assays were developed. Firstly, swarming in defined media supplemented with defined carbon sources was investigated using soft agar plates containing DMEM supplemented with 5% MEMα (DMEM/MEMα). DMEM does not support growth of C. jejuni, while MEMα does support growth [22]. Cells inoculated into DMEM soft-agar, supplemented with defined carbon sources (MEMα), therefore migrate away from the point of inoculation as the carbon sources are utilized, resulting in a swarming halo. Cells that lack the ability to move towards these carbon sources can be said to have a defect in energy taxis. To extend these assays, we also developed a tube-based taxis assay using 15 ml Falcon tubes, where TTC-supplemented Brucella soft agar was inoculated with a cell suspension at the top. In this assay, movement of cells is visible as a dye-front that migrates away from the point of inoculation over time (Fig. 2C). Cell migration through the agar was shown to be dependent on growth, as no migrating dye-front was observed in tubes incubated below the minimum growth temperature for C. jejuni (sub 35°C) or in tubes supplemented with the bacteriostatic antibiotic chloramphenicol (data not shown). Time-lapse photography was used to show that the dye-front progressed evenly over time. Migration of the dye-front was influenced by the oxygen-tension, with tubes incubated in air with 10% CO2 showing the greatest level of dye migration, while tubes incubated at 2% O2-10% CO2 showed the lowest level of dye migration (Fig. S1). Cell migration was independent of gravity, with the same migration observed if tubes were incubated up-side-down. All further assays were carried out at 37°C in air. A non-motile ΔflaAB mutant did not form halos in the DMEM/MEMα swarm plates, and this behaviour was mimicked in the tube assays, as dye migration was only 20% that compared to the wild-type strain (Fig. 2B, Fig. 3).
The benefit of using the tube assay was demonstrated when the agar was supplemented with the redox indicator resazurin (7-hydroxy-3H-phenoxazin-3-one 10-oxide) [28]. This blue dye is non-reversibly reduced to pink resorufin, then further reversibly reduced to colourless hydroresorufin [29]. Parallel assays in tubes supplemented with TTC and resazurin showed that as the cells migrated through the agar (as shown in the TTC tubes), the resazurin in the supplemented tubes was being reduced, as shown by a colour change from pink to yellow (i.e. colourless, and therefore the colour of the Brucella medium, Fig. 2C). Thus, as a result of cell growth and metabolic activity, both energy and redox gradients are established in the tubes. Comparison of swarm plates using four different strains, with and without resazurin, showed no difference in motility; hence we concluded that the presence of resazurin, like TTC, does not influence motility and cell migration (data not shown).
The CetAB system is required for efficient energy taxis
The two taxis assays described in the previous section were used to assess whether the swarming phenotype of the ΔcetAB mutant was due to an energy taxis defect. The ΔcetAB mutant showed both poor swarming and poor migration in the tube assay (Fig. 2B, Fig. 2E, and Fig. 3). In DMEM/MEMα medium, the halo size of the cetAB mutant was 20% that of the wild-type, whilst in the tube assay, dye migration was 50% that of the wild-type (Fig. 3). Disrupting cetA and cetB individually also resulted in migration defects, as the ΔcetA strain showed 26% halo swarming and 50% migration compared to the wild-type, and the ΔcetB strain showed 50 to 60% swarming and migration compared to wild-type (Fig. 2E, and Fig. 3).
To show that the taxis phenotypes observed for the ΔcetB strain was due to the cetB gene, a complementation strain was constructed in which the cetB gene was expressed from the C. jejuni fdxA promoter [30]. These gene-promoter fusions were inserted into the chromosome in the pseudogene cj0046c [24], resulting in the ΔcetB::cetB* strain. Complementation of the ΔcetB strain by cetB resulted in wild-type migration in the tube assays (a small but significant increase was observed after 24 hrs in the tube assay) and wild-type halo sizes in all swarm plates suggesting that expression of CetB could rescue the migration defects observed in the ΔcetB mutant (Fig. 3). A complementation strain was also constructed in which the cetA gene was expressed from the fdxA promoter. Reasoning that stoichiometry between the CetA and CetB proteins may be critical for correct functioning of the sensing complex, cetA was also expressed from its native promoter. A strain expressing cetA from the fdxA promoter showed the same level of swarming and migration as the ΔcetA strain (Fig. 3). When cetA was expressed from its own promoter, migration was 80% of that of the wild-type strain in the early stages (24 hrs) of the tube assay (although significantly different from the wild-type) but subsequently fell to levels seen in the ΔcetA mutant (Fig. 3B). In the DMEM/MEMα swarming assays, this strain showed a slight increase in halo size compared to the ΔcetA mutant, but could not restore wild-type swarming (Fig. 3A).
Given that cetA and cetB are co-transcribed [19], disrupting cetA is likely to disrupt cetB expression. Indeed, there was no significant difference in taxis between the ΔcetA and ΔcetAB mutants in both the swarm plate and tube assays. This may account for the failure to complement a ΔcetA mutant with cetA alone. Thus, a complementation construct, containing both cetA and cetB, in the same transcriptional arrangement as the original locus, was made and transformed into the ΔcetA, ΔcetB, and ΔcetAB backgrounds. Strains complemented with the cetAB genes showed normal growth, and microscopy revealed robust swimming motility in liquid media and presence of bipolar flagella. In both the swarm plate assays and tube assays, halo size and migration of strains transformed with cetAB were indistinguishable from the ΔcetA and ΔcetAB mutants (Fig. 3). However, the ΔcetB mutant complemented with cetAB showed wild-type levels of swarming and migration in the tube assays. Indeed, after 24 hours in the tube assay, the ΔcetB::cetAB strain showed 120% mean migration compared to the wild-type, similar to levels seen in the ΔcetB::cetB* strain (Fig. 3B). This suggests that at least CetB is expressed and functional when downstream of cetA, and supports the findings that cetA and cetB are co-transcribed. Successful complementation with cetB but limited success with cetA could be due to the requirement for a delicate balance between the two components for a functioning sensing complex or due to technical issues with over-expression of the membrane-bound CetA. Previous attempts to complement an 81–176 ΔcetA strain with cetA were also unsuccessful [20]. Given that cetAB are co-transcribed and complementation with cetA was unsuccessful, it is not possible to fully discriminate the cetA and cetAB phenotypes. However, disruption of this locus clearly has a striking effect on taxis.
Cj1191c (CetC) functionally complements a ΔcetB mutant and is required for robust taxis in concert with CetB
The genomes of all sequenced C. jejuni strains (except C. jejuni subsp. doylei 269.97) are annotated to contain a gene upstream of cetA, which encodes a protein that is 64% identical to CetB and is also predicted to encode a PAS domain protein (70% identity over the predicted PAS domain region), see Fig. 1 and Fig. S2. To determine the role of this protein in energy taxis, the cj1191c gene was inactivated by insertion of a kanamycin cassette. The Δcj1191c strain had the same growth-rate as the wild-type strain and did not have a motility defect as shown with light microscopy and AAG. Microscopy showed normal cell morphology and bipolar flagella. The Δcj1191c strain also showed wild-type levels of taxis towards L-fucose in the chemotaxis swarming assay (data not shown). In all swarm plate assays and the tube assay, halo size and migration were the same as wild-type (Fig. 3). Given the similarity to CetB, to ability of Cj1191c to complement the ΔcetB mutant was investigated by introducing the cj1191c gene, under control of the fdxA promoter, into the cj0046 pseudogene in the ΔcetB mutant. Swarming halos and migration in the tube assay of the ΔcetB mutant complemented with cj1191c was equivalent to the wild-type showing that, like CetB, Cj1191c can also rescue the ΔcetB phenotype (Fig. 3). Therefore, in a ΔcetB mutant, a CetA-Cj1191c complex may contribute to signal transduction and taxis. To address this question, a double ΔcetB Δcj1191c mutant was constructed by transforming the kanamycin-resistant ΔcetB mutant with a Δcj1191c::chloramphenicol disruption construct. This double mutant had no growth defect, possessed bi-polar flagella, and showed robust swimming in liquid media. However, in the both the swarm plate and tube assays, the ΔcetB Δcj1191c double mutant showed greatly reduced halo size (around 10% of the wild-type) and migration (Fig. 3). In Brucella swarm plates, halos from the ΔcetB Δcj1191c mutant were significantly smaller than ΔcetA, ΔcetB, ΔcetAB, and Δcj1191c mutants. In DMEM/MEMα swarm plates, the same trend was observed (with the exception of the ΔcetAB strain, Fig. 3A). In the tube assays too, the ΔcetB Δcj1191c double mutant showed significantly less migration than the ΔcetA, ΔcetB, ΔcetAB, and Δcj1191c mutants (Fig. 3B). Thus, while a Δcj1191c mutant is not impaired for taxis, the data suggests that the Cj1191c protein can form a functional sensing complex with CetA, and that Cj1191c in concert with CetB is required for efficient energy taxis. Hence we propose to rename it CetC.
Chimeric CetA-CetB and CetA-CetC proteins cannot rescue the ΔcetAB phenotype
The cetA and cetB genes are co-transcribed [19] and together are homologous to the E. coli aerotaxis sensor Aer. To investigate if a CetA-CetB chimeric fusion protein formed a functional sensing protein, an in-frame cetA-cetB fusion construct, expressed from the fdxA promoter, was generated. In light of the observations that both CetB and CetC were able to complement the ΔcetB mutant (Fig. 3), a cetA-cetC fusion construct was also generated. Both constructs were introduced into a ΔcetAB strain and swarming and tube migration assessed. In all the swarm plate assays, halos from the strains transformed with the chimeric constructs were the same as the ΔcetAB strain (Fig. S3, A). In the tube assays, the strains containing the chimeric constructs showed a trend towards reduced migration, even compared to the ΔcetAB strain (significantly different at 40 and 48 hrs, as determined by a t-test, Fig. S3, B). These strains were not impaired for growth and retained the ability to autoagglutinate. It is not clear why these fusion constructs fail to rescue the cetAB phenotype, but this could be due to incorrect folding of these chimeric proteins, or due to the reasons given previously for the cetA complementation.
Cj1110c (CetZ) encodes a chemotaxis protein that has an opposing role to CetAB
In addition to the PAS-domain proteins encoded by cetB and cetC, the C. jejuni NCTC 11168 genome also contains the cj1110c (tlp8) gene, which encodes a protein containing an MCP domain and two PAS domains [7], see Fig. 1. A prediction of transmembrane regions using the DAS algorithm recommended in [31], [32], suggested that Cj1110c does not contain a recognisable transmembrane region or periplasmic export signals, and is therefore most likely located in the cytoplasm. According to the proposed classification of MCP proteins, this places Cj1110c into the IVa topology group [6], and Class IV [32].
To investigate the role of this protein in energy taxis in C. jejuni, the cj1110c was disrupted by a chloramphenicol resistance cassette. The resulting Δcj1110c strain had a wild-type growth rate and did not have a motility defect, as determined using AAG and light microscopy, and had wild-type morphology including bipolar flagella. In the swarming plate assays, conversely to the phenotype shown by the cet mutants, halos from the Δcj1110c strain were larger than those of the wild-type (Fig. 2D, and Fig. 3A). In Brucella soft-agar, the mean halo size was 150% that of the wild-type, while in DMEM/MEMα, the halo size was 120% of the wild-type (Fig. 3A). Likewise, in the TTC tube assay, migration of the dye-front of the Δcj1110c strain was 150% that than the wild-type strain after 24 hours (Fig. 2E, and Fig. 3B). Complementation of the Δcj1110c strain with the cj1110c gene expressed from the fdxA promoter rescued and even reversed the Δcj1110c phenotype. The halo size from the swarm plate assays was 70–80% that of the wild-type (Fig. 2D, and Fig. 3A) and migration of the dye-front in the tube assay was 66% of wild-type migration after 24 hours (Fig. 3B). As all the phenotypes observed suggest an action opposite to that of CetAB, and hence we propose the name CetZ for Cj1110c.
The cet phenotypes are CheA-dependent and the CetAB system predominates over CetZ
To investigate if the taxis phenotypes observed in the ΔcetA, ΔcetB, and ΔcetZ mutants is dependent on a functioning chemotaxis pathway, a ΔcheA mutant was constructed. The ΔcheA strain showed the same growth rate as wild-type and exhibited swimming motility as shown with light microscopy (data not shown), and microscopy confirmed the presence of bi-polar flagella. When testing for chemotactic swarming using 1 M L-fucose as a chemo-attractant, halo formation was abolished in the ΔcheA strain. The ΔcetA, ΔcetB, or ΔcetAB strains showed wild-type halo formation towards fucose suggesting that the cet phenotype is not due to a chemotaxis defect. The ΔcheA mutant showed both reduced halo size in swarming assays (Fig. 2D) and migration in tube assays (Fig. 3B), suggesting that the phenotype observed in the ΔcetA, ΔcetB, and ΔcetAB mutants is dependent on a functioning chemotaxis pathway.
To investigate whether CetZ uses the chemotaxis signalling pathway, a ΔcheA ΔcetZ double mutant was also made by transforming the ΔcheA background with the cetZ disruption construct. The ΔcheA ΔcetZ strain did not exhibit increased halo size or increased migration in the TTC tube assay, as observed in the ΔcetZ strain (Fig. 3). In DMEM/MEMα swarm plates, the halo size of the ΔcheA ΔcetZ mutant was not significantly different to the ΔcheA strain, whilst in Brucella media, halo size was 80% of the wild-type (Fig. 3A). In the tube assays, migration of the ΔcheA ΔcetZ strain was not significantly higher than wild-type, as the ΔcetZ mutant, and showed the same trend as the ΔcheA mutant in the later time points (Fig. 3B). The CetZ system therefore appears to be unable to bypass the CheA-mediated pathway.
If both the CetABC and CetZ systems are mediated via the chemotaxis pathway, the predominance of one system over the other was investigated by testing double mutants (ΔcetA ΔcetZ, ΔcetB ΔcetZ, ΔcetAB ΔcetZ), constructed by inactivating the cetA, cetB, and cetAB genes in the ΔcetZ background. In all the swarming assays, halo sizes were less than 50% of the wild-type (Fig. 3A) and in the tube assays, migration of the dye front was most similar to the cet phenotype (Fig. 2E, and Fig. 3B). These data suggest that the CetAB system predominates over the CetZ system.
Discussion
In this work we have characterised two systems (CetABC and CetZ) that jointly govern energy taxis in C. jejuni, by opposing actions. Energy taxis influences movement of cells dependent on the intracellular metabolic status, and allows the cells to move towards the most favourable conditions for growth or respiration [33]. The observations that ΔcetA and ΔcetB mutants display reduced energy taxis assay in agar plates [18] have been extended by using a new, and convenient tube-based assay to track taxis as a function of the migration of a dye-front, in a column of media supplemented with a redox-indicator. The conversion of TTC in the media is a marker for metabolic activity and suggests that, as the cells grow and migrate, a local energy gradient is established as the nutrients are exhausted. Tubes supplemented with the redox indictor resazurin also show that cell growth and migration is accompanied by the formation of a redox gradient (Fig. 2C), most likely due to utilization of dissolved oxygen, the preferred respiratory electron acceptor in C. jejuni [34], [35]. We therefore suggest that these assays can be employed for studying energy taxis in C. jejuni. This simple assay may also be adaptable for studying taxis in other bacteria. The ease of set-up and data gathering make it an attractive and scalable method, especially when compared to more sophisticated but complicated assays reported, based on the use of fluorescently-labelled cells [17] or microscope video tracking of single cells [36].
The observations made with the cetABC and cetZ mutants in this study support the original hypothesis from which the cetAB genes derive their name [18]. Originally, ΔcetA and ΔcetB mutants in C. jejuni strain 81–176 showed reduced taxis towards both the carbon source sodium pyruvate and alternative electron acceptor fumarate. However, the specific effector of the CetAB system remains unknown, be it direct sensing of molecular oxygen (as in the case of HemAT in Bacillus subtilis [37]) or indirect redox sensing. We favour the second hypothesis, and propose that these proteins function via a redox mechanism, likely mediated through a redox-active co-factor, such as FAD, bound to the PAS domain of CetB or CetC. Given the fact that CetB is soluble in the cytoplasm, the reason of localising the CetA component in the membrane remains unknown. The topology of CetA has been delineated, with the N-terminal being cytoplasmic, and a loop region projecting into the periplasm [19]. The role of the periplasmic region in signal sensing, if any, remains an open question.
Disrupting the CetB-like protein (Cj1191c) upstream of the CetAB system alone had no discernable phenotype. However, a lack of both this CetC (Cj1191c) protein and CetB had a dramatic effect on taxis, while retaining swimming motility, suggesting that CetB and CetC may act in concert in signal transduction. Accordingly cetC (cj1191c) was observed to rescue the energy taxis phenotype of the cetB mutant suggesting that CetC alone can form a functioning signal transduction complex with CetA (Fig. 3). Ergo, why should the genome encode both a CetB and second ‘substitute’ PAS domain sensor? In microarray studies reporting transcriptional regulation of cetAB, cetC is not reported as being differentially regulated, whereas the cetAB genes display growth phase-dependent regulation of expression [38], [39], [40]. It is possible that cetC is constitutively expressed at a basal level such that the sensing complex can always function. Then, under appropriate stresses when the cetAB locus is up-regulated, CetB assumes the role of primary sensor. The fact that the ΔcetB energy taxis phenotype in the tube assay is less severe than that of the ΔcetA mutant (60% migration compared to 50% migration of wild-type) may be explained by formation of a functioning CetA - CetC complex. Moreover, this hypothesis may go someway to explain the epithelial cell invasion phenotypes described previously [20]. Only the ΔcetA strain, but not the ΔcetB strain, was shown to have an invasion defect for human epithelial cells. If signal transduction through the Cet complex is required for invasion, it is possible that in the ΔcetB mutant, CetC functions in place of CetB to form a functional signalling complex. The data from the tube assays containing the chimeric CetA-CetB and CetA-CetC constructs showed reduced cell migration even compared to the ΔcetAB mutant. However, in all experiments where complementation with cetA was attempted, wild-type taxis could not be restored, whether cetA alone (ΔcetA::cetA fdxApr*), as a transcriptional unit with cetB (11168 ΔcetA::cetAB fdxApr*) or as a chimeric construct fused to either cetB or cetC. CetA complementation therefore seems beyond the technical limitations of the genetic tools used in this study. Given that CetB and CetC are expressed as discrete proteins, the ability of the PAS domain proteins to transiently interact with CetA may be critical for signal transduction. Correct functioning of the Cet complex may require dynamic interaction between CetA and both PAS domain proteins.
The results of the energy taxis assays with the ΔcetZ (Δcj1110c) mutant clearly demonstrated an opposing phenotype compared to that observed in the ΔcetAB mutants, a phenotype that could be rescued, and even reversed, by complementation (Fig. 2D, and Fig. 3). Thus, if the CetAB system has a role in energy taxis, does the opposing function of CetZ control taxis away from favourable nutritional environments? We suggest that CetZ is a redox sensor, which mediates taxis away from high redox potentials, associated with high oxygen concentrations (Fig. 4). In addition to the data presented here, the work on the role of the CetZ-ortholog AerC in A. brasilense is particularly revealing [11]. This protein shares the same domain architecture of CetZ and functions to mediate taxis away from high oxygen tensions (and thus redox potentials) to microaerobic conditions (low oxygen tension/redox potential) to permit nitrogen fixation by the oxygen-sensitive nitrogenase complex [11]. Given its predicted cytoplasmic location, CetZ may sense a cytoplasmic redox-active metabolite, or use a redox-sensing cofactor, such as FAD (as is the case for E. coli Aer, the N-terminal PAS domain of Azotobacter vinelandii NifL [41] and A. brasilense AerC [11]), although direct interaction with the respiratory chain cannot be ruled out. PAS domains are known to be involved in direct protein-protein interactions, often facilitating homo- and hetero-dimer formation [12]. NifL from A. vinelandii contains two tandem PAS domains, the second PAS domain lacking any bound cofactor and involved in homodimer formation [42]. In the published C. jejuni protein-protein interaction network [43], CetZ shows interactions with 19 other proteins, including CheV (in C. jejuni, a hybrid CheW-response regulator protein), but no other components of the chemotaxis pathway or respiratory complex. Thus, the precise sensing mechanism used by CetZ requires further investigation.
Figure 4. Model of how the combined activity of the CetABC and CetZ systems control energy taxis in C. jejuni.

A) Diagram showing our assumptions about the tube taxis assay (right), schematically displaying a hypothesis on role and sensing by the CetZ and CetABC systems. B) Model displaying possible signal transduction by the CetABC and CetZ system (left, adapted from [19]) and their interaction with the chemotaxis pathway via the CheA protein. The dominant CetABC pathway is shown by bold arrows (see discussion for more information). ETS, electron transport chain; OM, outer membrane; IM, inner membrane.
A double ΔcetAB ΔcetZ strain was used to demonstrate that the CetAB system pre-dominates over the CetZ system (Fig. 3), as this mutant displayed the reduced taxis of the ΔcetAB mutant. Thus, for C. jejuni, the ability to seek out an environment for optimal energy generation probably supersedes redox taxis. Indeed, energy taxis has been shown to be the primary mechanism Campylobacter uses to navigate to an optimal oxygen tension [33]. This perhaps reflects the ability of C. jejuni to utilize multiple electron acceptors with different redox potentials, ranging from the preferred electron acceptor oxygen (+820 mV) down to fumarate (+30 mV) [35]. Moreover, C. jejuni possesses two terminal oxidases: a lower affinity CioAB system and a high affinity cb-type cytochrome c oxidase [44], which may permit respiration at very low oxygen concentrations. It should also be noted that in E. coli too, the Aer system, which is most similar to the CetAB system, predominates over chemotaxis mediated by the Tsr, Tar, Trg, and Tap chemoreceptors [45].
The energy taxis phenotype of the cheA mutant showed 50–70% swarming/migration compared to the wild-type strain, suggesting that the CheA-CheY signal transduction pathway has an explicit role in energy taxis. However, the cheA phenotype was less striking than the cetAB phenotype, suggesting that taxis is not entirely abolished. So, while CheA is likely the primary pathway for energy taxis signal transduction, bypasses may exist, such as promiscuous phosphorylation of CheY.
In summary, we present data demonstrating that C. jejuni contains two taxis systems which have opposing roles, which are likely to function to maintain the cells at an optimum energy and redox balance. CetABC is an energy taxis system, utilizing two cytoplasmic sensing modules (CetB and CetC), which drives cells to favourable conditions for energy generation whilst CetZ drives cells away from high redox potentials (Fig. 4). Both pathways mediate their effects through CheA and the downstream chemotaxis system. The CetAB system predominates over CetZ, suggesting that while C. jejuni can fall back on sub-optimal electron acceptors for respiration, an exodus from low energy environments drives taxis permitting continued survival and proliferation.
Supporting Information
Taxis of C. jejuni NCTC 11168 in the TTC-based tube assay is dependent on the oxygen tension, as shown after quantification of energy taxis in different atmospheric conditions.
(PDF)
Genetic organization of the cetABCZ loci in the Campylobacter genus based on available genome sequences. Genes are drawn roughly to scale and additional flanking genes are hashed. Pseudogenes are shown by interrupted boxes.
(PDF)
Energy taxis phenotype of the C. jejuni Δ cetAB strain, and the Δ cetAB strain complemented with cetA - cetB and cetA - cetC chimeric constructs.
(PDF)
Plasmids used in this study.
(PDF)
Primers used in this study.
(PDF)
Acknowledgments
We thank Joseph Sundaradhas for technical assistance, and Duncan Gaskin and David Guy for construction of flagellar mutants. We thank members of the IFR Campylobacter group for helpful discussions, and also thank Maddy Houchen for microbiology media support and Gary Wortley for assistance preparing for time-lapse photography.
Funding Statement
This study was supported by the Biotechnology and Biological Sciences Research Council (BBSRC, http://www.bbsrc.ac.uk) through the Institute Strategic Programmes IFR/08/3 (Biology and Complexity of Foodborne Bacterial Pathogens) and BB/J004529/1 (Gut Health and Food Safety) to the Institute of Food Research. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Pallen MJ, Wren BW (2007) Bacterial pathogenomics. Nature 449: 835–842. [DOI] [PubMed] [Google Scholar]
- 2. Jung K, Fried L, Behr S, Heermann R (2012) Histidine kinases and response regulators in networks. Curr Opin Microbiol 15: 118–124. [DOI] [PubMed] [Google Scholar]
- 3. Krell T, Lacal J, Munoz-Martinez F, Reyes-Darias JA, Cadirci BH, et al. (2011) Diversity at its best: bacterial taxis. Environ Microbiol 13: 1115–1124. [DOI] [PubMed] [Google Scholar]
- 4. Baker MD, Wolanin PM, Stock JB (2006) Signal transduction in bacterial chemotaxis. Bioessays 28: 9–22. [DOI] [PubMed] [Google Scholar]
- 5. Lertsethtakarn P, Ottemann KM, Hendrixson DR (2011) Motility and chemotaxis in Campylobacter and Helicobacter . Annu Rev Microbiol 65: 389–410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Lacal J, Garcia-Fontana C, Munoz-Martinez F, Ramos JL, Krell T (2010) Sensing of environmental signals: classification of chemoreceptors according to the size of their ligand binding regions. Environ Microbiol 12: 2873–2884. [DOI] [PubMed] [Google Scholar]
- 7. Marchant J, Wren B, Ketley J (2002) Exploiting genome sequence: predictions for mechanisms of Campylobacter chemotaxis. Trends Microbiol 10: 155–159. [DOI] [PubMed] [Google Scholar]
- 8. Alexandre G (2010) Coupling metabolism and chemotaxis-dependent behaviours by energy taxis receptors. Microbiology 156: 2283–2293. [DOI] [PubMed] [Google Scholar]
- 9. Schweinitzer T, Josenhans C (2010) Bacterial energy taxis: a global strategy? Arch Microbiol 192: 507–520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Edwards JC, Johnson MS, Taylor BL (2006) Differentiation between electron transport sensing and proton motive force sensing by the Aer and Tsr receptors for aerotaxis. Mol Microbiol 62: 823–837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Xie Z, Ulrich LE, Zhulin IB, Alexandre G (2010) PAS domain containing chemoreceptor couples dynamic changes in metabolism with chemotaxis. Proc Natl Acad Sci? U? S? A 107: 2235–2240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Taylor BL, Zhulin IB (1999) PAS domains: internal sensors of oxygen, redox potential, and light. Microbiol Mol Biol Rev 63: 479–506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Tam CC, O'Brien SJ, Tompkins DS, Bolton FJ, Berry L, et al. (2012) Changes in causes of acute gastroenteritis in the United Kingdom over 15 years: microbiologic findings from 2 prospective, population-based studies of infectious intestinal disease. Clin Infect Dis 54: 1275–1286. [DOI] [PubMed] [Google Scholar]
- 14.Jacobs-Reitsma WF, Lyhs U, Wagenaar J (2008) Campylobacter in the Food Supply. In: Nachamkin I, Szymanski CM, Blaser MJ, editors. Campylobacter. Third ed. Washington, DC: American Society for Microbiology. pp. 627–644.
- 15. Finn RD, Mistry J, Tate J, Coggill P, Heger A, et al. (2010) The Pfam protein families database. Nucleic Acids Res 38: D211–222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Tareen AM, Dasti JI, Zautner AE, Gross U, Lugert R (2010) Campylobacter jejuni proteins Cj0952c and Cj0951c affect chemotactic behaviour towards formic acid and are important for invasion of host cells. Microbiology 156: 3123–3135. [DOI] [PubMed] [Google Scholar]
- 17. Hartley-Tassell LE, Shewell LK, Day CJ, Wilson JC, Sandhu R, et al. (2010) Identification and characterization of the aspartate chemosensory receptor of Campylobacter jejuni . Mol Microbiol 75: 710–730. [DOI] [PubMed] [Google Scholar]
- 18. Hendrixson DR, Akerley BJ, DiRita VJ (2001) Transposon mutagenesis of Campylobacter jejuni identifies a bipartite energy taxis system required for motility. Mol Microbiol 40: 214–224. [DOI] [PubMed] [Google Scholar]
- 19. Elliott KT, DiRita VJ (2008) Characterization of CetA and CetB, a bipartite energy taxis system in Campylobacter jejuni . Mol Microbiol 69: 1091–1103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Elliott KT, Zhulin IB, Stuckey JA, DiRita VJ (2009) Conserved residues in the HAMP domain define a new family of proposed bipartite energy taxis receptors. J? Bacteriol 191: 375–387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Reuter M, Mallett A, Pearson BM, van Vliet AH (2010) Biofilm formation by Campylobacter jejuni is increased under aerobic conditions. Appl Environ Microbiol 76: 2122–2128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. van Vliet AHM, Wooldridge KG, Ketley JM (1998) Iron-responsive gene regulation in a Campylobacter jejuni fur mutant. J? Bacteriol 180: 5291–5298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. van Vliet AHM, Wood AC, Henderson J, Wooldridge KG, Ketley JM (1998) Genetic manipulation of enteric Campylobacter species. Methods Microbiol 27: 407–419. [Google Scholar]
- 24. Thomas MT, Shepherd M, Poole RK, van Vliet AHM, Kelly DJ, et al. (2011) Two respiratory enzyme systems in Campylobacter jejuni NCTC 11168 contribute to growth on L-lactate. Environ Microbiol 13: 48–61. [DOI] [PubMed] [Google Scholar]
- 25. Golden NJ, Acheson DW (2002) Identification of motility and autoagglutination Campylobacter jejuni mutants by random transposon mutagenesis. Infect Immun 70: 1761–1771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Hugdahl MB, Beery JT, Doyle MP (1988) Chemotactic behavior of Campylobacter jejuni . Infect Immun 56: 1560–1566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Tengerdy RP, Nagy JG, Martin B (1967) Quantitative measurement of bacterial growth by the reduction of tetrazolium salts. Appl Microbiol 15: 954–955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Erb RE, Ehlers MH (1950) Resazurin Reducing Time as an Indicator of Bovine Semen Fertilizing Capacity. J? Diary Sci 33: 853–864. [Google Scholar]
- 29. O'Brien J, Wilson I, Orton T, Pognan F (2000) Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur? J? Biochem 267: 5421–5426. [DOI] [PubMed] [Google Scholar]
- 30. van Vliet AH, Baillon MA, Penn CW, Ketley JM (2001) The iron-induced ferredoxin FdxA of Campylobacter jejuni is involved in aerotolerance. FEMS Microbiol Lett 196: 189–193. [DOI] [PubMed] [Google Scholar]
- 31. Cserzo M, Wallin E, Simon I, von Heijne G, Elofsson A (1997) Prediction of transmembrane alpha-helices in prokaryotic membrane proteins: the dense alignment surface method. Protein Eng 10: 673–676. [DOI] [PubMed] [Google Scholar]
- 32. Wuichet K, Alexander RP, Zhulin IB (2007) Comparative genomic and protein sequence analyses of a complex system controlling bacterial chemotaxis. Methods Enzymol 422: 1–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Vegge CS, Brondsted L, Li YP, Bang DD, Ingmer H (2009) Energy taxis drives Campylobacter jejuni toward the most favorable conditions for growth. Appl Environ Microbiol 75: 5308–5314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Weingarten RA, Grimes JL, Olson JW (2008) Role of Campylobacter jejuni respiratory oxidases and reductases in host colonization. Appl Environ Microbiol 74: 1367–1375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kelly DJ (2008) Complexity and Versality in the Physiology and Metabolism of Campylobacter jejuni In: Nachamkin I, Szymanski CM, Blaser MJ, editors. Campylobacter. Third ed. Washington, DC: American Society for Microbiology. pp. 41–61.
- 36. Howitt MR, Lee JY, Lertsethtakarn P, Vogelmann R, Joubert LM, et al. (2011) ChePep controls Helicobacter pylori Infection of the gastric glands and chemotaxis in the Epsilonproteobacteria . MBio 2: e00098–00011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Hou S, Larsen RW, Boudko D, Riley CW, Karatan E, et al. (2000) Myoglobin-like aerotaxis transducers in Archaea and Bacteria. Nature 403: 540–544. [DOI] [PubMed] [Google Scholar]
- 38. Stahl M, Friis LM, Nothaft H, Liu X, Li J, et al. (2011) L-fucose utilization provides Campylobacter jejuni with a competitive advantage. Proc Natl Acad Sci? U? S? A 108: 7194–7199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Wright JA, Grant AJ, Hurd D, Harrison M, Guccione EJ, et al. (2009) Metabolite and transcriptome analysis of Campylobacter jejuni in vitro growth reveals a stationary-phase physiological switch. Microbiology 155: 80–94. [DOI] [PubMed] [Google Scholar]
- 40. Holmes K, Tavender TJ, Winzer K, Wells JM, Hardie KR (2009) AI-2 does not function as a quorum sensing molecule in Campylobacter jejuni during exponential growth in vitro. BMC Microbiol 9: 214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Key J, Hefti M, Purcell EB, Moffat K (2007) Structure of the redox sensor domain of Azotobacter vinelandii NifL at atomic resolution: signaling, dimerization, and mechanism. Biochemistry 46: 3614–3623. [DOI] [PubMed] [Google Scholar]
- 42. Slavny P, Little R, Salinas P, Clarke TA, Dixon R (2010) Quaternary structure changes in a second Per-Arnt-Sim domain mediate intramolecular redox signal relay in the NifL regulatory protein. Mol Microbiol 75: 61–75. [DOI] [PubMed] [Google Scholar]
- 43. Parrish JR, Yu J, Liu G, Hines JA, Chan JE, et al. (2007) A proteome-wide protein interaction map for Campylobacter jejuni . Genome Biol 8: R130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Jackson RJ, Elvers KT, Lee LJ, Gidley MD, Wainwright LM, et al. (2007) Oxygen reactivity of both respiratory oxidases in Campylobacter jejuni: the cydAB genes encode a cyanide-resistant, low-affinity oxidase that is not of the cytochrome bd type. J? Bacteriol 189: 1604–1615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Taylor BL, Zhulin IB, Johnson MS (1999) Aerotaxis and other energy-sensing behavior in bacteria. Annu Rev Microbiol 53: 103–128. [DOI] [PubMed] [Google Scholar]
- 46. Parkhill J, Wren BW, Mungall K, Ketley JM, Churcher C, et al. (2000) The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 403: 665–668. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Taxis of C. jejuni NCTC 11168 in the TTC-based tube assay is dependent on the oxygen tension, as shown after quantification of energy taxis in different atmospheric conditions.
(PDF)
Genetic organization of the cetABCZ loci in the Campylobacter genus based on available genome sequences. Genes are drawn roughly to scale and additional flanking genes are hashed. Pseudogenes are shown by interrupted boxes.
(PDF)
Energy taxis phenotype of the C. jejuni Δ cetAB strain, and the Δ cetAB strain complemented with cetA - cetB and cetA - cetC chimeric constructs.
(PDF)
Plasmids used in this study.
(PDF)
Primers used in this study.
(PDF)